










-
Tea plants (Camellia sinensis) are one of the most economical woody plants[1]. The cultivation of tea originated in southwest China. However, unfavorable conditions and extreme climate, such as heavy metal toxicity, drought, high salinity, ABA and temperature stress, affect the quality and yield of tea during the growth of tea plants[2,3].
Transcription factors (TFs) play vital regulatory roles in plant response to diverse stresses[4]. Among them, members of the WRKY family, which are designated by the WRKYGQK signature domain, widely exist in plants. WRKY proteins usually contain a highly conserved WRKYGQK sequence and a zinc finger motif (either C2H2 or C2HC) at their C-terminus. Based on the number of WRKY domains and the pattern of the zinc finger motif, the WRKY family is further divided into groups I-III[5−7]. The first identified WRKY protein, SPF1, was isolated from sweet potato[8]. Subsequently, numerous WRKY proteins have been identified in different plant species[7,9−11]. Moreover, the majority of relevant studies have suggested that WRKY plays a vital role in plant growth and development. For example, Arabidopsis AtWRKY44 regulates epidermal color and root hairs[12,13]. AtWRKY53 and AtWRKY75 act as positive regulators of plant senescence[14,15].
Furthermore, WRKY proteins have been discovered to respond to abiotic stresses. Arabidopsis AtWRKY57 positively regulates drought stress[16]. AtWRKY63 plays an important role in plant respons to ABA and drought stress[17]. In rice, the homolog OsWRKY45 also acts in ABA signaling and drought tolerance[18].
Growing evidence suggests that group I WRKY proteins are involved in responses to abiotic stress. Gossypium hirsutum WRKY25 positively regulates the response to salinity stress in transgenic tobacco[19]. Similarly, wheat TaWRKY2 and TaWRKY19 regulate salt, drought and/or freezing stress responses in transgenic Arabidopsis plants[20]. Arabidopsis AtWRKY3 provides resistance to herbivory stresses, while AtWRKY25, AtWRKY26, and AtWRKY33 act synergistically in thermotolerance[21]. Jiang and Deyholos showed that Arabidopsis WRKY25 and WRKY33 function redundantly as positive regulators of the NaCl stress response[22]. Some recent studies have focused on WRKY factors in tea plants[23−26]. However, the physiological functions of CsWRKY proteins in abiotic stress are still poorly understood. Here, we aimed to provide a functional characterization of CsWRKY26, a member of the group I WRKY family in tea plants. CsWRKY26, was a close homolog of Arabidopsis AtWRKY26, AtWRKY25 and AtWRKY33, three well-characterized Group I WRKY family proteins with important roles in plant thermotolerance, osmotic stress responses, biotic stresses and camalexin biosynthesis[21,22,27−29]. In the present study, we cloned and characterized CsWRKY26 from tea plant cultivar ‘Longjing 43’. CsWRKY26 was significantly induced by multiple abiotic stresses. Overexpressing CsWRKY26 in transgenic Arabidopsis increased resistance to drought stress. These findings may provide new insights into the abiotic stress tolerance mechanism of CsWRKY26, which may serve as a target for molecular breeding for crop improvement and enhancement of stress tolerance.
-
The full-length WRKY26 was obtained by high fidelity PCR. Owing to its high sequence identity with A. thaliana WRKY26, the WRKY protein was named CsWRKY26. The gene contains a 1,674 bp open reading frame encoding a 557 amino acid protein, with a predicted isoelectric point of 8.77 and a molecular mass of 61.41 KDa.
CsWRKY26 shared high amino acid sequence similarity with AtWRKY26, AtWRKY25, and AtWRKY33 (49%, 46% and 48% respectively). Multiple sequence alignment showed that CsWRKY26 shared two typical WRKY DNA-binding domains, i.e., WRKYGQK and one CX1-5CX22-23HXH zinc finger motif (Fig. 1a), which categorized it as a group I WRKY superfamily member[5]. And the sequence of CsWRKY26 contained an extended C-terminal domain (CTD) compared to those of AtWRKY33.
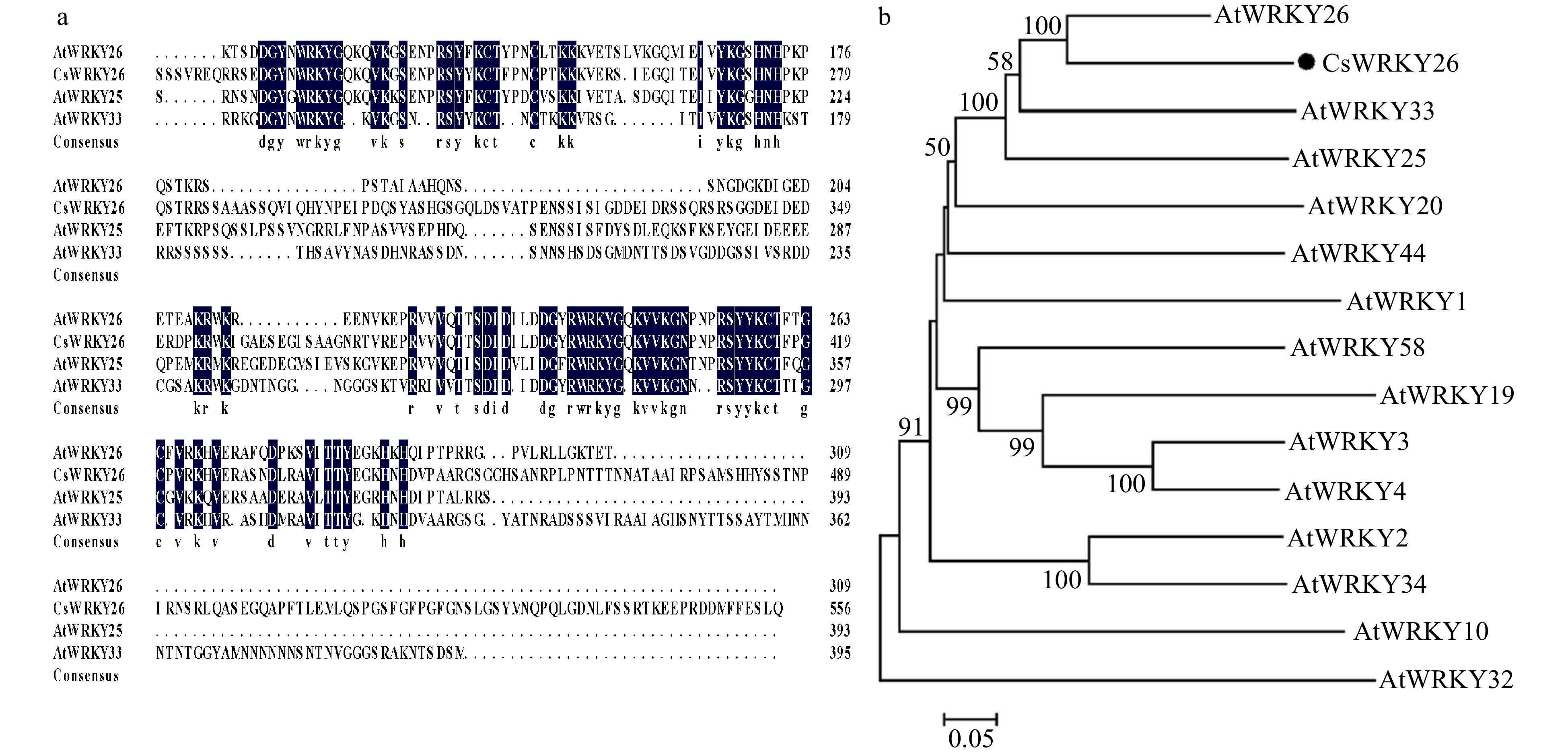
Figure 1. Sequence alignment and phylogenetic tree analysis of CsWRKY26. (a) Multiple sequence alignment of amino acid sequences of CsWRKY26 with homologous WRKYs from Arabidopsis. Sequences were aligned by the DNAMAN program. (b) Phylogenetic analysis of CsWRKY26 and Arabidopsis group I WRKY proteins, constructed using MEGA 6.0.
To analyze the evolutionary relationship of CsWRKY26 within group I WRKY subfamily members from A. thaliana, a phylogenetic tree was constructed using MEGA 6.0 software (Fig. 1b). Among these proteins, AtWRKY25, AtWRKY26 and AtWRKY33 contribute to plant defense stress and plant thermotolerance[21], these findings imply that CsWRKY26 is involved in stress response.
Expression patterns of CsWRKY26 under multiple stress treatments
-
Quantitative RT-PCR was performed to detect CsWRKY26 transcript abundance in leaves and roots under osmotic stress (Fig. 2). In leaves, CsWRKY26 was dramatically induced by NaCl and PEG treatments, the transcript levels rapidly peaked nearly 30 and 70-fold at 24 h post treatment, respectively. Additionally, CsWRKY26 was also induced both in sucrose and mannitol treatments. In roots, CsWRKY26 was significantly induced by osmotic stresses at the initial stages (1−3 h). Simultaneously, CsWRKY26 displayed a sustained increase during natural drought stress and rehydration processes. These results suggest that CsWRKY26 may play important roles in stress signaling, especially in response to osmotic stress.
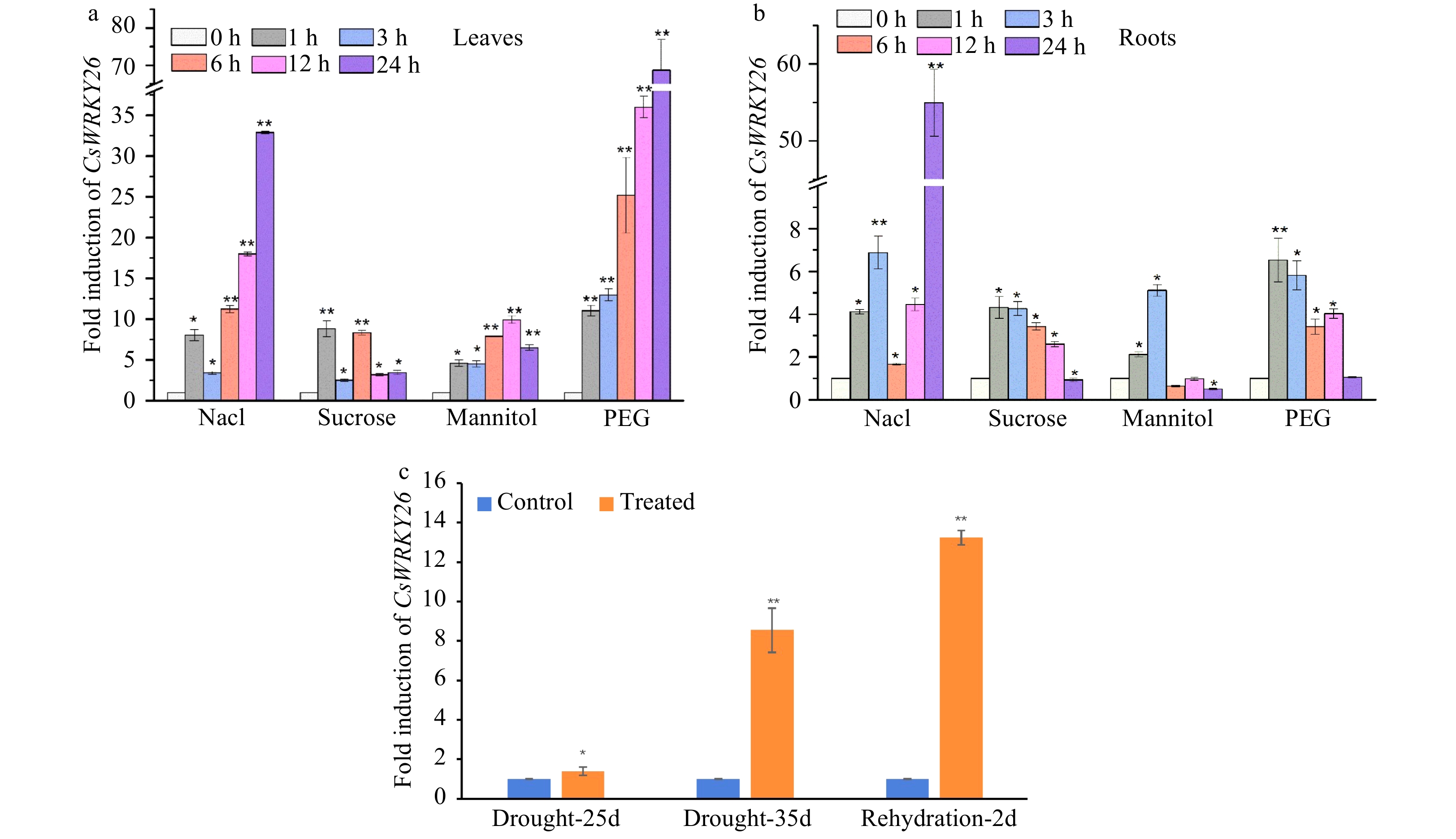
Figure 2. Expression patterns of CsWRKY26 gene in tea plants under different abiotic stress. (a) CsWRKY26 transcripts accumulate in leaves under 150 mM NaCl, 90 mM sucrose, 10% (W/V) PEG4000, 90 mM mannitol stresses. (b) CsWRKY26 transcripts accumulate in roots under the same stress as in (a). (c) The fold change of CsWRKY26 in leaves under natural drought stress. The relative expression levels of the CsWRKY26 gene were detected using a qRT-PCR approach. Asterisks indicate that the expression level is significantly different from the value of the control (*p < 0.05, **p < 0.01).
To investigate the association between CsWRKY26 and some signaling molecules, the expression of CsWRKY26 under salicylic acid (SA), abscisic acid (ABA), Gibberellic acid (GA3) and Methyl Jasmonate (MeJA) treatments was analyzed by qRT-PCR. As shown in Fig. 3, CsWRKY26 displayed upregulation to some extent under various hormone treatments, with results that could be divided into three expression profiles: (A) sustained upregulation, peaking at 6 h (GA3); (B) sharply increasing at the initial stage and then decreasing (ABA and MeJA); (C) rapidly increasing in the late stage (SA). It is worth noting that CsWRKY26 was significantly induced by ABA and MeJA treatments, with more than a 5-fold induction compared with that under normal conditions.
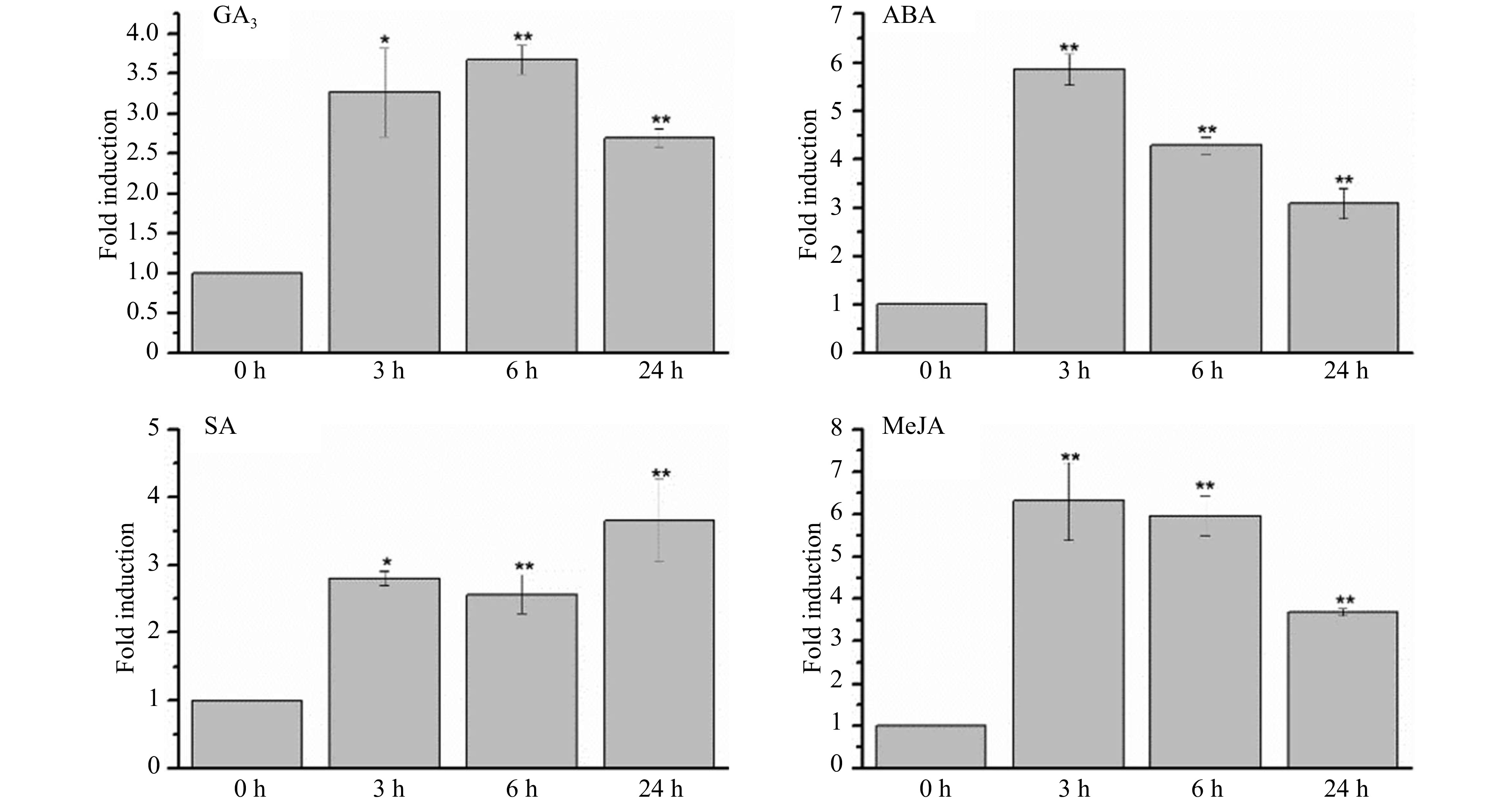
Figure 3. CsWRKY26 expression in leaves under various hormones treatments. The seedlings were treated with different hormones (100 µM GA3, 100 µM ABA, 5 mM SA and 50 µM MeJA), and leaves were harvested for expression analysis after 24 h of treatment. The level of CsWRKY26 expression were normalized to GAPDH and compared with control conditions. Data shown are averages of three biological replicates, with error bars representing SD. The asterisks indicate that the expression level is significantly different from the value of the control (*p < 0.05, **p < 0.01).
CsWRKY26 localized in the nucleus
-
Subcellular localization of CsWRKY26 was investigated. The full-length ORF of WRKY26 was fused to the N-terminal of enhanced green fluorescent protein (eGFP) and then transiently expressed in Nicotiana benthamiana epidermal cells (Fig. 4a). The control 35S::GFP showed green fluorescence throughout the whole cell. Confocal microscopy demonstrated that green fluorescence was detected in the nucleus when the CsWRKY26-fusion protein was transformed in tobacco. In addition, targeting ability was also determined in CsWRKY26 overexpression seedling roots (Fig. 4b), and confocal microscopy showed that CsWRKY26 is localized in the nucleus.
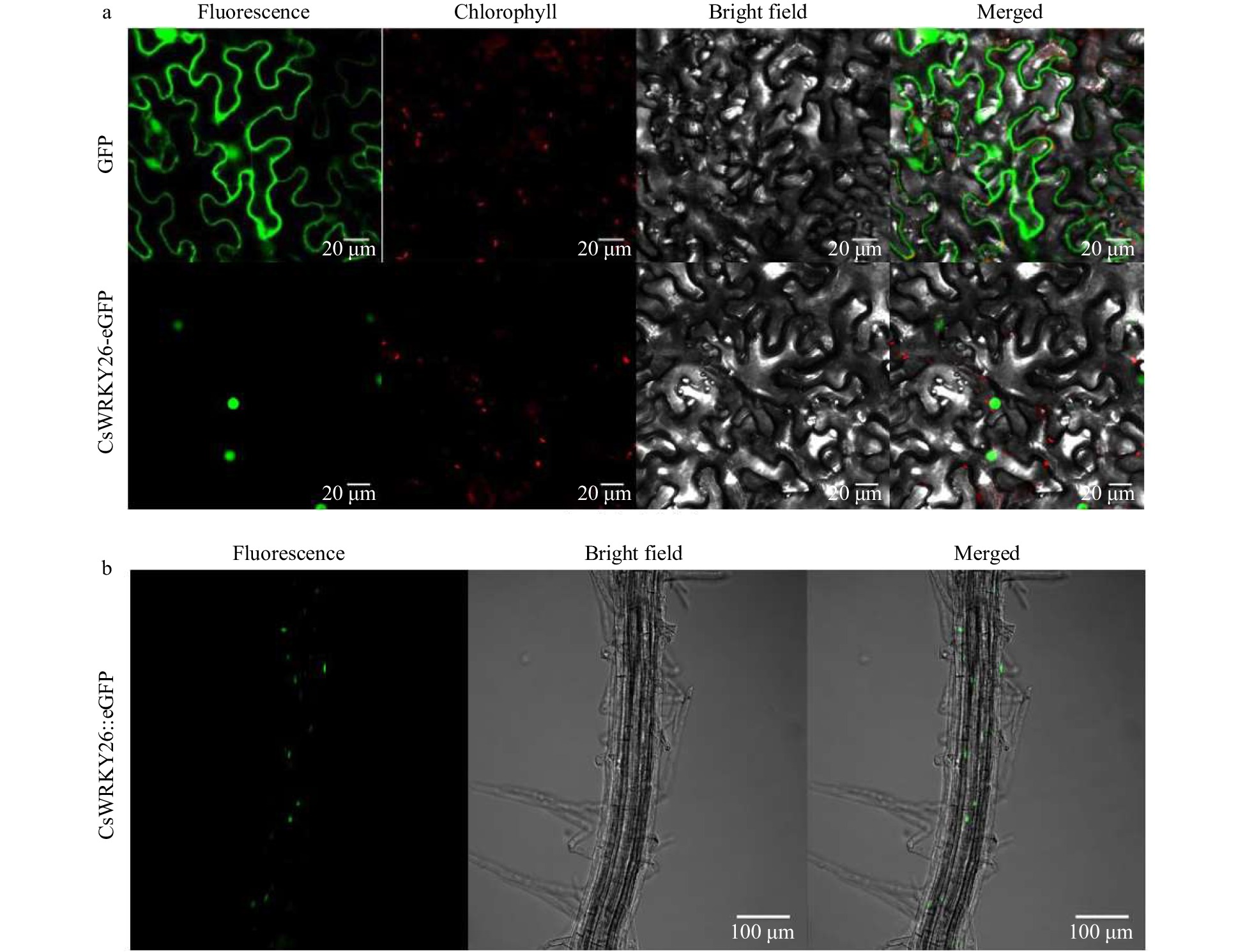
Figure 4. Subcellular localization analysis of CsWRKY26 protein. (a) GFP alone (upper panel) and CsWRKY26-eGFP (middle panel) were transiently expressed in tobacco epidermal cells. Representative images from left to right in each panel were taken under fluorescence, chlorophyll, transmitted light and an overlay of both channels. Scale bar = 20 µm. (b) Subcellular localization of CsWRKY26-eGFP in Arabidopsis roots. Representative images from left to right were taken under fluorescence, bright field and an overlay of both channels. Scale bar = 100 µm.
Tissue-specific expression analysis
-
The expression profiles of CsWRKY26 was assayed by qRT-PCR or β-glucuronidase (GUS) transcript under control of the CsWRKY26 promoter fragments (the 1.25 kb fragment upstream of the ATG start codon was fused to the GUS open reading frame).
CsWRKY26 exhibited high expression levels in leaves and roots, especially in older leaves, which were approximately 17 times higher than that in buds (Fig. 5a). Meanwhile, proCsWRKY26::GUS reporter gene fusion constructs were investigated by histochemical staining. Under normal conditions, we detected strong expression in the whole 7-d-old seedlings and in the roots and veins of leaves in both 10 and 20-d-old seedlings. Among inflorescence tissues, moderate to strong GUS expression was observed in filament, petal, stigma, ovary, sepal, and silique tissues (Fig. 5b). However, these mRNA accumulation patterns were inconsistent with the GUS expression patterns, indicating that regulatory elements beyond the promoter region may have affected this result or that the sensitivity of the reporter system may be insufficient to detect more moderate changes.
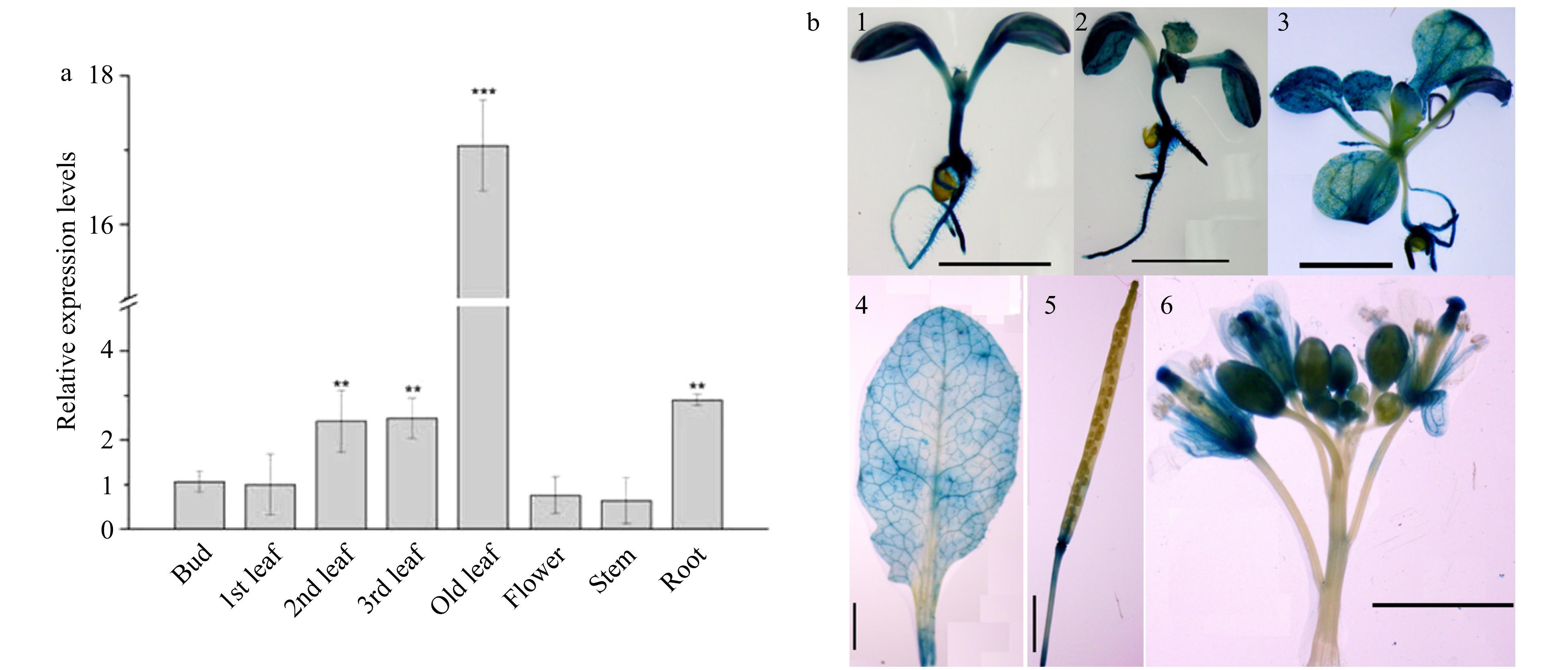
Figure 5. Tissue-specific expression patterns of CsWRKY26 gene. (a) Quantitative RT-PCR of CsWRKY26 expression in various tissues: bud, 1st leaf, 2nd leaf, 3rd leaf, old leaf, flower, stem and root. The transcript levels of CsWRKY26 gene in various tissues were related to GAPDH and compared with that of bud. Asterisks indicate that the expression level is significantly different from the value of the control (*p < 0.05, **p < 0.01). (b) Histochemical staining of transgenic Arabidopsis with ProCsWRKY26::GUS, (1) 7-d-old seedling, (2) 10-d-old seedling, (3) 20-d-old seedling, (4) mature rosette leaf, (5) silique, (6) flowers. Scale bars = 2 mm.
Overexpression of CsWRKY26 in Arabidopsis enhances tolerance to osmotic stresses
-
CsWRKY26 was induced by salinity and PEG treatments (Fig. 1). Thus, to investigate whether CsWRKY26 medicates the responses to osmotic stress, three independent transgenic Arabidopsis lines (OE1-3) and WT seedlings were subject to a series of abiotic stress. The root length of WT plants was slightly longer than those of CsWRKY26 transgenic plants under normal conditions (Fig. 6). There was no difference between WT and transgenic plants under salt stress. However, two lines showed longer root lengths than WT plants both in the presence of 0.3 μM ABA and 15% PEG6000. Thus, we speculate that CsWRKY26 may be involved in ABA-dependent drought-responsive regulatory pathways during early growth.
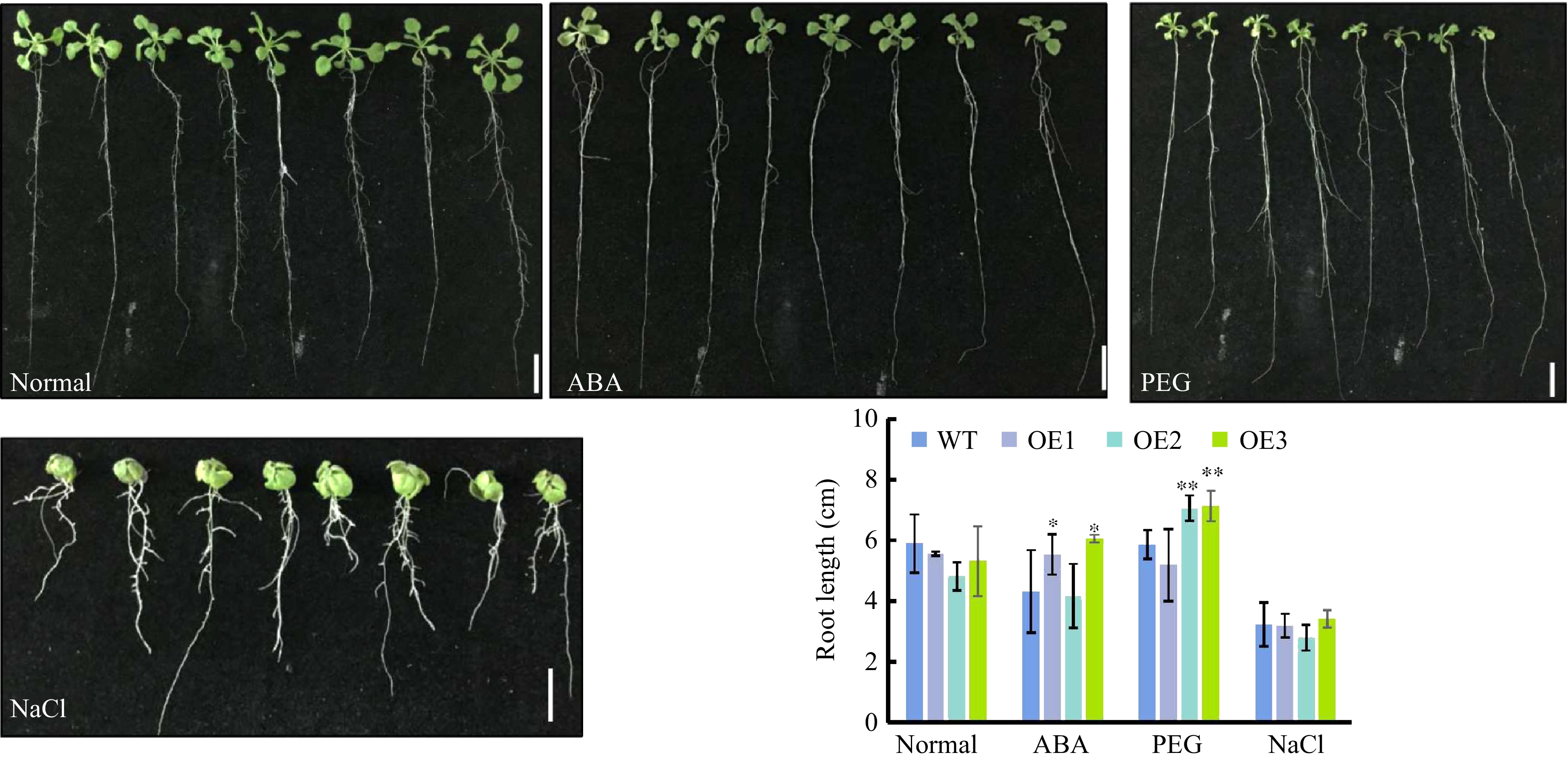
Figure 6. Root elongation of CsWRKY26-overexpressing Arabidopsis lines under various stress conditions. Four-d-old seedlings were transferred onto plates with or without 0.3 μM ABA, 15% PEG6000, 150 mM NaCl for 10 d, and numbers of root length were determined after 10 d of vertical growth (n > 12). Bars and error bars represent mean ± SE, scale bars =1 cm.
Transgenic Arabidopsis plants expressing CsWRKY26 showed improved drought tolerance
-
Under normal conditions and salt treatment, the growth vigor of CsWRKY26-overexpression plants was weaker than that of WT plants (Fig. 7). However, WT plants exposed to the 15% PEG6000 treatment for 7 d showed leaf curling and atrophy, while most of the transgenic lines showed undamaged leaves. Under drought stress for 15 d, leaves from WT lines were purple with severe wilting compared to those in the overexpression lines.
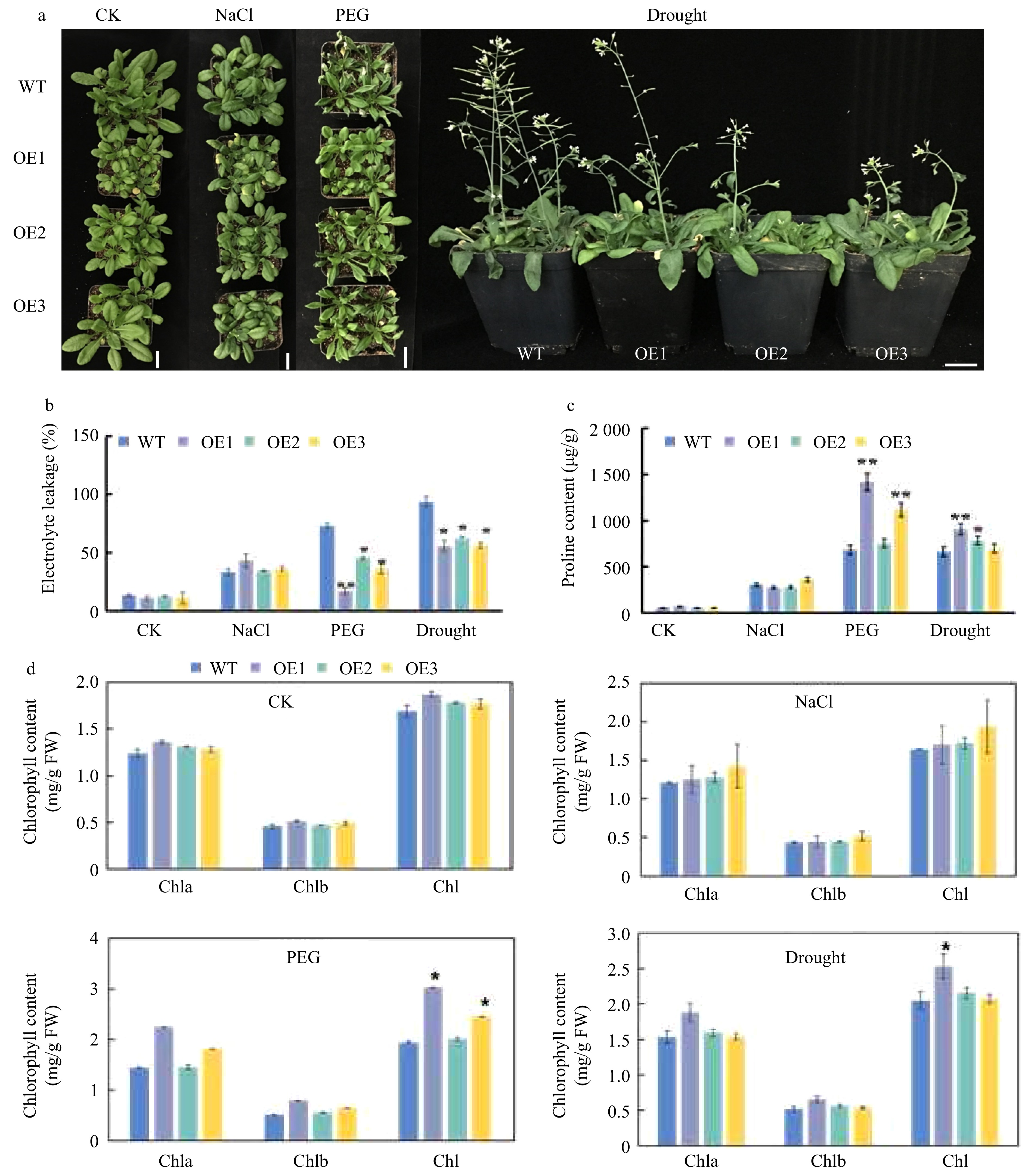
Figure 7. Phenotype of CsWRKY26-overexpressing transgenic plants and WT plants under different stress conditions. (a) Phenotype of WT and of CsWRKY26-overexpression Arabidopsis lines grown with or without exposure to salt treatment for 7 d, 15% PEG6000 treatment for 7 d, and a natural 15-d drought. (b−d) Determination of physiological indicators: relative electrolyte leakage, proline content, and chlorophyll content. Scale bars = 2 cm.
Electrical leakage was closely correlated with the degree of damage to cell membranes under various stresses. The relative electrical conductivity (REL) in transgenic plants under salt treatment was higher than that of WT plants, but there was no significant difference. Compared with WT plants, the transgenic plants had lower REL under PEG6000 and natural drought treatment. The chlorophyll content of WT and transgenic leaves under drought treatment were assayed. Both CsWRKY26-overexpression and WT plants exhibited increases in chlorophyll content, whereas chlorophyll content did not significantly differ between most transgenic lines and WT plants. Meanwhile, proline content varied among transgenic plants after salt treatment. The proline content in OE1 and OE2 lines were lower than that in WT plants, but these differences were not statistically significant. However, OE1 and OE3 lines showed significantly higher proline content compared with that of WT plants under drought stress (Fig. 7). Collectively, our results indicate that CsWRKY26 overexpression enhanced drought stress in transgenic Arabidopsis.
Expression of stress-related genes in CsWRKY26 transgenic Arabidopsis
-
To elucidate the regulation mechanisms underlying drought resistance in CsWRKY26 overexpression lines, we further detected the expression profiles of stress-responsive genes (namely DREB2A, RD29A, P5CS1, RD22, NCED3, ABI1, and ABI5) in 7-d-old vertically grown seedlings cultivated under drought. The expression levels of RD29A, NCED3, and ABI1/-5 were constitutively increased in transgenic plants under normal conditions compared to that of WT plants (Fig. 8a). Furthermore, the expression levels of NCED3, RD22, and P5CS1 were significantly elevated in OE1 and OE3 lines, which is consistent with the proline content increases in transgenic lines (Fig. 8b). In addition, the stress-related genes DREB2A and RD29A were significantly down-regulated, indicating that CsWRKY26 may be involved in either ABA-dependent or ABA-independent stress signal transduction pathways.
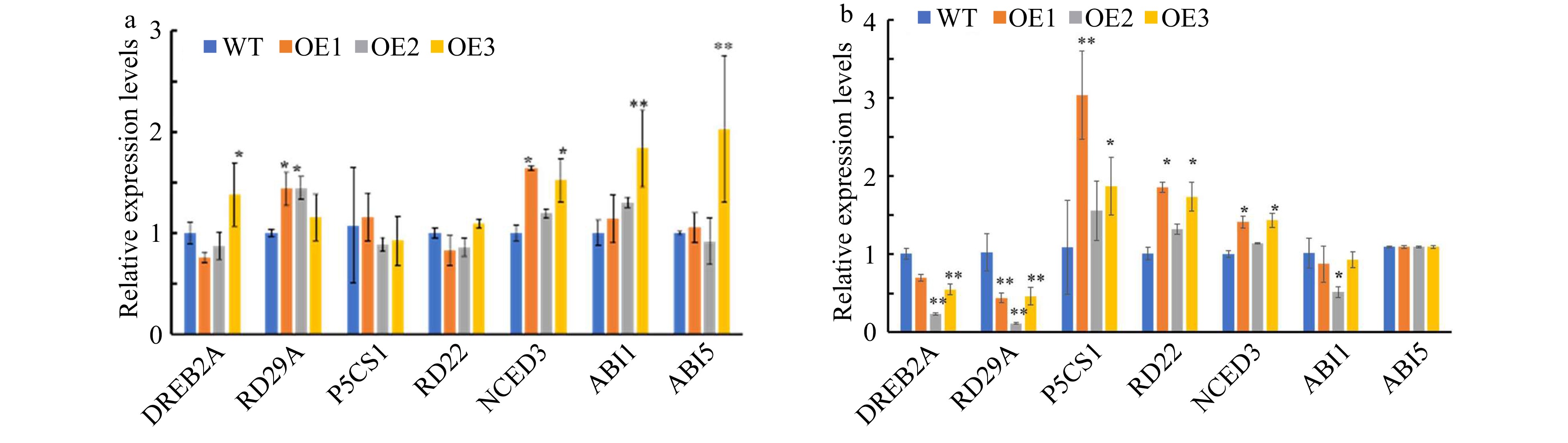
Figure 8. Analysis of expression levels for stress-related genes in CsWRKY26-overexpressing Arabidopsis lines and WT plants under normal and 15% PEG6000 conditions. Transcript levels of these genes in the transgenic lines and WT plants were determined by quantitative RT-PCR using Actin2 as a quantification control. Asterisks indicated that the expression level is significantly different from that of the control (*p < 0.05, **p < 0.01). (a) Normal conditions. (b) Simulated drought conditions induced by15% PEG6000.
-
WRKY transcription factors can act as transcriptional activators or inhibitors in response to stress stimuli[30]. Members of this family have been reported to participate in responses to multiple abiotic stresses, for example, Arabidopsis AtWRKY57/-63[16,17] and sunflower HaWRKY76 participate in drought stress responses[31], while Arabidopsis AtWRKY18/-25/-30/-60 participate in osmotic stress responses[22,32,33]. However, the functions of WRKY proteins in tea plants under abiotic stress have remained unclear.
In the present study, CsWRKY26 was cloned from tea plants. Two typical WRKY domains and a group I CX1-5CX22-23HXH zinc finger motif (Fig. 3a) were observed in the CsWRKY26 sequence[5]. CsWRKY26 showed high amino acid sequence identities with AtWRKY25, -26, and -33. Notably, the sequence alignment revealed an extended C-terminal domain (CTD) compared to those of AtWRKY33. Previous studies have shown that the CTD in AtWRKY33 plays a vital role in plant stress response[34]. Arabidopsis AtWRKY25 or AtWRKY33 overexpression increases the salt tolerance and sensitivity to ABA of transgenic plants[22]. Thus, it is likely that the CTD of CsWRKY26 plays vital roles in plant response to stress.
Here, CsWRKY26 was significantly expressed in salt- and drought-induced tea leaves (Fig. 2). This induction is similar to that of soybean GmWRKY16[35], indicating that CsWRKY26 in tea plants may participate in coping with drought stress. Previous studies have shown that many WRKY TFs are involved in diverse hormone signaling pathways. For example, OsWRKY24/-53/-70 inhibited ABA and GA signal transduction in rice aleurone cells[36], while banana MaWRKY31/-33/-60/-71 positively regulated drought tolerance in its ABA-dependent pathway[37]; similarly, Arabidopsis AtWRKY28/-46 activated the transcription levels of related genes in the SA signaling pathway[38]. CsWRKY26 was significantly induced by ABA and MeJA hormones (Fig. 2), indicating that it may participate in one or more phytohormone signaling cascades.
Drought is a major abiotic factor limiting growth, adversely affecting both plant growth and economic benefits. The role of WRKYs in drought stress responses have been reported in many plants. Boea hygrometrica BhWRKY1 regulated the transcription level of galactosidase BhGolSl (a key rate-limiting enzyme in the oligosaccharide synthesis pathway) in the ABA-dependent signaling in response to drought stress; thus, the content of osmotic substances was increased[39]. In tobacco, expression of cotton GhWRKY41 or tomato SpWRKY1 conferred tolerance to drought by regulating stomatal conductance and reactive oxygen level[40,41]. Overexpression of cotton GhWRKY17 increased sensitivity to drought in tobacco by impairing ABA-induced stomatal closure and reducing the level of ABA[42]. Arabidopsis AtWRKY40/-18/-60 proteins act as negative regulators of ABI4 and ABI5 genes in the ABA signaling pathway[32]. Rice OsWRKY45 confers resistance to drought stress[18]. In the present study, CsWRKY26 overexpression promoted root elongation in transgenic Arabidopsis treated with 0.3 µM ABA or 15% PEG6000 (Fig. 6). Moreover, NCED3 (9-cis epoxycarotenoid dioxygenase 3), a key enzyme in the ABA synthesis pathway that is involved in drought stress responses[43], was induced in CsWRKY26 transgenic plants under PEG-simulated drought conditions (Fig. 8). Similar functions were also found in other WRKY genes. For example, overexpressing AtWRKY57 in Arabidopsis increased tolerance to drought by enhancing NCED3 expression and increasing accumulation of ABA content[16].
Studies have reported that osmotic stress imposed by drought are transmitted through at least two pathways: ABA-dependent and ABA-independent signaling pathways[6,16,18,32,42−45]. Notably, in our research, some related genes involved in those two pathways, such as RD29A, RD22, and DREB2A, were found to be significantly induced by drought stress, showing that CsWRKY26 may be involved in the abscisic acid signaling pathway (Fig. 8). Dehydration response element binding (DREB) TFs play important roles in plant stress response. Dubouzet et al.[46] showed that OsDREB1A and OsDREB2A, which specifically bound to DRE, were significantly up-regulated in transgenic rice lines, and conferred drought tolerance to rice. However, the expression levels of DREB2A in CsWRKY26-overexpressing transgenic lines were significantly reduced compared to that of WT plants, which may relate to sampling time (Fig. 8). Miao et al.[47] showed that the expression of DREB2A was significantly up-regulated in the early stage during drought stress but later down-regulated in MaASR1 transgenic Arabidopsis lines. Proline plays a critical role in plant osmotic stress tolerance by stabilizing subcellular structures and maintaining cellular functions[48]. Here, Proline content significantly increased in WT and transgenic plants under drought stress. Meanwhile, P5CS1 expression was significantly elevated, consistent with the proline accumulation observed in transgenic plants (Fig. 7). Similar regulatory mechanisms have been found in other studies, which have confirmed that the adaptability of similar transgenic plants to drought stress is related to the dehydration avoidance mechanism of proline metabolism regulation[49].
In addition, WRKY TFs form a complex network with other TFs to regulate plant stress. For example, Glycine max GmWRKY27 interacts with GmMYB174 to suppress GmNAC29 expression and enhance stress tolerance in soybean plants[50]. AtWRKY33 acts downstream of the mitogen activated protein kinase genes MPK3 and MPK6 in reprogramming camalexin biosynthesis genes[51]. Andreasson et al.[52] reported that MPK4 was found to interact with MKS1, a VQ motif protein substrate of MPK4 substrate, that in turn interacts with Arabidopsis WRKY25 and WRKY33. Zhou et al. (2015)[34] reported that WRKY33 is an evolutionarily conserved WRKY transcription factor with a broad and critical role in plant stress responses. However, most reports indicate that WRKY25 and WRKY33 may be involved in diverse defense mechanisms against most stresses[22,27−29]. Notably, the MBS motif was found in the CsWRKY26 region, implying that CsWRKY26 may interact with MYB TF to participate in drought stimuli (unpublished). Therefore, further research should focus on the components related to WRKY26 during drought stress in tea plants.
-
WRKY proteins play a vital role in response to abiotic stress. This study has identified a CsWRKY26 gene from tea plant cultivar ‘Longjing 43’. CsWRKY26 was induced by sodium chloride (NaCl), polyethylene glycol (PEG), drought and different hormones treatments. CsWRKY26-overexpression transgenic Arabidopsis lines exhibited tolerance to drought stress. The findings reported here shed new light on the abiotic stress tolerance mechanism of CsWRKY26, which may serve as a target for molecular breeding of drought resistance tea cultivar.
-
‘Longjing 43’ (LJ43), a very popular and famous green tea (in particular for Longjing tea) cultivar bred by the Tea Research Institute of the Chinese Academy of Agricultural Sciences (TRICAAS), was used in this study. The buds, leaves, stems, flowers and roots were collected for tissue-specific analysis from seven-year-old tea plants, grown in China National Germplasm Hangzhou Tea Repository of the TRICAAS.
Two-year-old rooted cuttings, similar in both size and growth stage, were grown in pots inside the greenhouse of TRICAAS. These cuttings were subject to various abiotic stresses, such as natural drought, 150 mM NaCl, 10% (W/V) PEG4000, 90 mM sucrose or mannitol, and different hormones including 100 μM abscisic acid (ABA), 100 μM gibberellic acid (GA3), 50 μM Methyl Jasmonate (MeJA) and 5 mM salicylic acid (SA) treatment. Different treatments were performed as previously described[53]. For various osmotic treatments, the functional leaves (the third leaves from two-year-old rooted cuttings of tea plants) and roots were sampled at 0, 1, 3, 6, 12, or 24 h. For phytohormone treatments, the functional leaves were harvested at 0, 3, 6 or 24 h post-treatment. For natural drought stress, the functional leaves were harvested from two-year-old rooted cuttings of tea plants by withholding water for 25 d, 35 d and re-watering after 2 d. All samples were immediately frozen in liquid nitrogen and stored at −80 °C before analysis. All treatments contained at least three biological replicates.
Healthy leaves from tobacco (Nicotiana benthamiana) grown for 2−4 weeks were used for protein subcellular localization. The seeds of Arabidopsis thaliana were surface sterilized, vernalized for 3 d at 4 °C, and plants were grown on 1/2 Murashige and Skoog (MS) medium. Plants were analyzed on plates or transplanted to a sterilized soil 7 d after germination. Arabidopsis plants were grown in growth cabinets at 22 ± 1 °C under long day (LD, 16 h /8 h day/night cycle) photoperiods, while tobacco plants where maintained at 24 ± 2 °C under a photoperiod of 12 h /12 h day/night. The light intensity was at approximately 108 μmol/m2s.
Cloning of CsWRKY26 genes and phylogenetic analysis
-
Total RNA was extracted from different tissues of LJ43 using QIAGEN RNeasy Mini Kit and a reverse transcription reaction was performed using a FastKing gDNA Dispelling RT SuperMix Kit (KR118). The full-length sequence of CsWRKY26 was cloned using KOD DNA polymerase. The specific primers are listed in Supplemental Table S1. Polymerase chain reaction (PCR) product was purified from agarose gel, and the purified product was cloned into the pEASY-Blunt simple vector (Transgen Biotech Co., Ltd., Beijing, China). Multiple sequence alignments of CsWRKY26 in C. sinensis and Group I WRKY members in A. thaliana were performed in MEGA6.0 by the neighbor-joining method with 1000 bootstrap replicates.
Expression profiles analysis of CsWRKY26
-
The expression analysis of CsWRKY26 were detected by real-time quantitative polymerase chain reaction (qRT-PCR) or histochemical staining of β-glucuronidase (GUS). For GUS staining, the homozygous promoter-WRKY26::GUS (proCsWRKY26::GUS) transgenic Arabidopsis plants were immersed in 0.5 mg/mL XGluc (5-bromo-4-chloro-3-indolylglucuronide) solution in 100 mM sodium phosphate buffer (pH 7.0), 10 mM EDTA, 1 mM K4Fe(CN)6, 1 mM K3Fe(CN)6, 0.5% Triton X-100, and incubated at 37 °C in the dark as previously described[54]. Tissue was cleaned in 70% ethanol before the visualization.
The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene was used as the housekeeping gene. And the gene expression levels were calculated using the 2−ΔΔCt or 2−ΔCt method[55,56].
Generation of CsWRKY26 overexpression lines
-
For overexpression analysis, the 35S::CsWRKY26 plasmid was generated, carrying the coding sequence of WRKY26 in the binary destination vector pH7FWG2.0. Floral dip-transformation of the plasmid into Arabidopsis cv. Columbia (Col-0) resulted in at least three independent transgenic Arabidopsis lines selected by growth on plates containing 30 μg mL–1 hygromycin B[57].
For subcellular localization prediction of CsWRKY26 protein, the pH7FWG2.0-CsWRKY26 was expressed in tobacco leaf epidermal cells. Plants were incubated for 48 h before fluorescence imaging using a Nikon C2-ER confocal laser microscope. In addition, GFP signals were also detected in the young roots of CsWRKY26 overexpression lines.
For promoter-β-glucuronidase (GUS) fusion, the promoter region of WRKY26 from C. sinensis was amplified by high fidelity PCR. The identified fragment was inserted into the pBI101.3-GUS plus vector. The recombinant proWRKY26::GUS was introduced into the A. tumefaciens strain GV3101 using the freeze-thaw method and introduced into Arabidopsis using the floral dip method[57]. Homozygous transgenic lines were used in subsequent experiments. The amplification primers are listed in Supplemental Table S1.
Abiotic stress analysis of CsWRKY26-overexpression transgenic plants
-
The seeds of wild type (WT) and CsWRKY26 overexpression lines were planted on 1/2 MS medium for 4 d, and then transferred to 1/2 MS or 1/2 MS medium containing 150 mM NaCl, 15% PEG6000 or 0.3 μM ABA for 10 d. For osmotic stress assay, 4-week-old transgenic plants and WT plants were irrigated with a final concentration of 250 mM NaCl for 7 d. To elevate drought tolerance, 4-week-old plants were stopped irrigation for 15 d or subjected to 15% PEG6000 for 7 d.
Measurement of physiological index
-
The sterilized and vernalized seeds of WT and CsWRKY26 overexpression plants were germinated on 1/2 MS medium for 7 d, and then transferred to 41 mm Jiffy seedling blocks. Plants were grown in a light incubator for three weeks. After a 7-d period of salinity, 15% PEG6000 treatment, or natural drought stress treatment for 15 d, the rosette leaves were collected for measuring the REL, proline and chlorophyll content. The REL was measured as described by Wang et al.[58]. Proline accumulation was assayed using the proline test kit cat. A107 (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The chlorophyll content from each sample was estimated as previously described by Li et al.[59].
Analysis of stress related genes by qRT-PCR in transgenic plants
-
Four-d-old seedlings were placed vertically in 1/2 MS medium with or without 15% PEG6000 for 7 d, and then sampled for RNA isolation. Actin-2 gene act as the reference control gene, and the 2−ΔΔCt method was used to evaluate the relative expression level. All qRT-PCR experiments contained three biological replicates.
Statistical analysis
-
Each experiment described in the research contained at least three biological replicates. Statistical analysis was performed using the one-way analysis of variance (ANOVA). And the Built-in Functions in Excel 2019 was used for statistical analysis.
- This work was supported by China Agriculture Research System of MOF and MARA, the Chinese Academy of Agricultural Sciences through the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2017-TRICAAS) to LC.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer sequences used for PCR.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen W, Zheng C, Yao M, Chen L. 2021. The tea plant CsWRKY26 promotes drought tolerance in transgenic Arabidopsis plants. Beverage Plant Research 1: 3 doi: 10.48130/BPR-2021-0003

The tea plant CsWRKY26 promotes drought tolerance in transgenic Arabidopsis plants
- Received Date: 08 March 2021
- Accepted Date: 18 May 2021
- Published Online: 28 May 2021
Abstract: WRKY proteins participate in various plant physiological processes, especially in response to stress. However, limited information is available for WRKY proteins in tea plants. Camellia sinensis WRKY26 (CsWRKY26), a group I WRKY member, was characterized in this study. Tissue specificity of CsWRKY26 expression revealed that its transcripts are abundant in leaves and roots. Moreover, CsWRKY26 transcripts were markedly induced by treatments with sodium chloride (NaCl), polyethylene glycol (PEG), drought and different hormones. Ectopic expression of CsWRKY26 conferred ABA insensitivity and tolerance to transgenic Arabidopsis plants subjected to PEG treatments. When exposed to 15% PEG, the expression levels of some stress-tolerant genes, such as responsive to dehydration 22 (RD22), 9-cis-epoxycarotenoid dioxygenase 3 (NCED3), and Δ1-pyrroline-5-carboxylate synthetase 1 (P5CS1), were up-regulated in transgenic plants compared to wild-type (WT) plants. Under natural drought stress, the growth performance of transgenic lines was better than those of WT plants, consistent with their significantly lower relative electrolyte leakage (REL) and higher proline content. These findings indicate that CsWRKY26 could enhance drought tolerance in Arabidopsis. This study provides a potential strategy for future crop improvement research.
-
Key words:
- WRKY26 /
- Camellia sinensis /
- drought stresses /
- Arabidopsis