









-
Salinity and drought stresses are major environmental factors limiting plant growth, development and agriculture productivity. In the process of plant evolution, several response strategies to withstand these stresses were established, including osmoregulation through accumulating compatible solutes and protective proteins, ion balance via regulating ion absorption and transportation, and an antioxidant system for scavenging reactive oxygen. These strategies can be further excavated following the identification of several stress tolerant genes, which can perform important roles for genetic improvement and molecular breeding. Over the past 20 years, a large number of genes conferring salt and drought stress tolerance have been identified in model plant Arabidopsis and crops.
Systematic and accurate strategies of forward and reverse genetics have been established for important obtainment of plant genes mined in previous reports. However, the forward genetics method can be only applied to plants having genome information or mutants; therefore, most plants still utilize reverse genetics to explore function genes, like transcriptomics, proteomics and cDNA expression library screening. A functional screening-based approach named FOX (Full-length cDNA Over-expression) hunting system can detect and identify functional genes through heterologous overexpression of full-length cDNA libraries in microorganisms or in the model plant Arabidopsis[1−6]. Yeast as a unicellular eukaryote, was widely used for functional analysis or screening of plant stress-tolerant genes. Through FOX yeast system, three novel salt tolerant genes (coiled-coil protein, thaumatin-like protein and short peptide of 32 residues) are isolated from halophyte Salicornia europaea L.[1], and 12 drought tolerant genes are obtained in potato[2]. In a previous study, 18 salinity-tolerance genes and five Cd-tolerance genes were obtained from seashore paspalum through the yeast FOX system[3]. Following a similar approach, several stress tolerant genes were also screened in other plants like rice, zoysiagrass and physic nut[4−6].
As an excellent leguminous forage crop, alfalfa (Medicago sativa L.) is widely cultivated throughout the world and performs important functions in animal production and soil fertility improvement. However, drought and soil salinity largely inhibit the productivity and quality of alfalfa. Genetic transformation of several tolerant genes including CsLEA, ZxNHX and ZxVP1, can improve salt- and drought-tolerance in alfalfa[7,8]. Based on the strategies of transcriptomics and proteomics, the molecular mechanisms of alfalfa responding to salt and drought stress have been excavated[9,10]. In this work, we aim to screen potential salt and drought tolerant genes of alfalfa using the FOX hunting system, and detect their expression patterns under salt and drought stress conditions.
-
The seeds of alfalfa 'No.2 gongnong', bred by Jilin Academy of Agricultural Sciences and well known for its strong salt and drought tolerance[11], were germinated and grown in pots filled with mixed soil (nutritional soil : peat : sand = 1:1:2) for 40 d, the uniform shoots with 8 cm length were selected and cultivated hydroponically on 1/2 Hoagland nutrient solution for 20 d. Thereafter, the plants were hydroponically treated by salt stress with 250 mM NaCl and drought stress with 20% PEG6000, respectively. The experiment was performed in a climate-controlled room with 14 h/d photoperiod, photosynthetically active radiation of 850 μmol photons m−2 s−1, day/night temperature of 25/20 °C and relative humidity of 60%.
Leaves and roots were collected at 0, 3, 6, 12, 24, or 48 h, and each sample of total RNA was isolated using OMEGA plant RNA kit. Five micrograms for each RNA was picked and mixed for the library construction. The remaining RNA samples were frozen in liquid nitrogen and stored at −80 °C for qRT-PCR analysis of the candidate genes expression.
Construction cDNA yeast expression library
-
The construction of the alfalfa cDNA expression library was executed following the methods in our previous studies[3,4]. The mixed total RNA was purified with Dynabeads® mRNA Purification Kit and the first cDNA was synthesized with primer Biotin-attB2-Oligo(dT) following reverse transcription reaction. Full length first-strand cDNA was enriched with Cap antibody and further generated to double-stranded cDNA, which thereafter was connected with attB1-adapter using DNA ligase. BP recombination reaction with jointed cDNA and pDONR/Zeo entry vector was performed and transformed to DH10B competent cell by electroporation. Thousand-fold diluted library Bacilli was plated on solid medium (LB + 50 mg L−1 Zeocin) and cultured overnight until the clones appeared. PCR detection of 24 clones was performed with pDONR/Zeo vector universal primer pair (F1/R1: TCCCAGTCACGACGTTGTAAAACGACGGCCAGTCTT/AGAGCTGCCAGGAAACAGCTATGACCATGTAATACGACTC). The plasmids of above constructed entry library were extracted and recombined to yeast expression vector pDEST52 through a LR reaction. The reaction products were transformed into DH10B competent cells, and cultured overnight on solid medium (LB + 50 mg L−1 Ampicillin). PCR analyses of 24 clones were detected with pDEST52 vector universal primer (F2/R2: CGGTAGGTATTGATTGTAATTCTG/GCGTGAATGTAAGCGTGAC). The yeast expression library plasmids were further isolated.
Yeast transformation, candidate gene hunting and sequence analysis
-
The pDEST52 expression library plasmids were respectively transformed into salt-sensitive yeast mutant strain G19 and drought-sensitive yeast mutant strain Hog1 using PEG-lithium acetate-based transformation protocols. The transformed G19 was plated on SD solid medium with 2% galactose lacking uracil and histidine including 500 mM NaCl at 30 °C for 3−5 d until single clones appeared. The transformed Hog1 was cultivated on SD solid medium with 750 mM sorbitol lacking uracil at 30 °C for 3−5 d. Surviving single clones on SD solid medium with 500 mM NaCl or 750 mM sorbitol were grown overnight, and then streaked with different dilution production (102, 103, 104, and 105) on salinity or osmotic stress treated SD solid medium as well as control solid medium. The plasmids of transformed yeast clones exhibiting salinity or drought tolerance were separated using OMEGA yeast plasmid extraction kit and transformed into TOP10 E. coli cells for sequencing. Sequences were analyzed following BLASTX ( http://blast.ncbi.nlm.nih.gov/Blast.cgi).
qRT-PCR expression analysis of candidate salinity and osmotic stress tolerant genes
-
The accuracy of qRT-PCR analysis is dependent on the stability of reference genes. Following several known reports, we selected six candidate reference genes, (EF1a, ACT, TUB, 18S rRNA, GAPDH, UBC2) and evaluated their stability in alfalfa under salt- and drought-treated conditions through the 2−ΔΔCᴛ data analysis of RefFinder ( www.heartcure.com.au/reffinder) software. The primers of six reference genes were shown in Supplemental Table 1.
cDNA was synthesized as the template of leaves or roots RNA using PrimeScript RT reagent kit with gDNA eraser, and then 10-fold dilution of cDNA was applied for template of the qRT-PCR reaction. The primers of candidate salt and osmotic stress tolerance genes were designed with Primer Premier 5.0 software, following the principles of primer length 18−23 bp, GC content 45%−55%, melting temperature 55−60 °C, and amplified fragment length 80−300 bp. A LightCycler 480 II master instrument was adopted to perform the qRT-PCR of 15 μL reaction mixture containing 7.5 μL 2 × SYBR I master, 2 μL of diluted cDNA, 0.3 μL each primer and 4.9 μL ddH2O. Three steps of reaction procedure were adopted, including an initial 5 min denaturation of 95 °C, 40 cycles amplification (95 °C 10 Sec, 55 °C 10 Sec and 72 °C 10 Sec), and production of melting curves with 60−95 °C. Relative expression levels of the target genes were calculated using the 2−ΔΔCᴛ method[12]. The data of three biological repeats were used for significance analysis following one-way detection of variance (ANOVA) of SPSS statistical software.
Generation of ERVT or CIPK11 transgenic Arabidopsis and tolerance analysis
-
The full-length ORFs of ERVT and CIPK11 were PCR amplified with primers pair ERVT-ORF-F/ERVT-ORF-R (GTCGACATGGGAATGAAACAAGTTATTAAG/GATATCTCTTTTTAGTCTTTGGTTTAGTTGC) and CIPK11-ORF-F/CIPK11-ORF-R (GTCGACATGGCGAATGAAGAGTCAACAAAC/GATATCCCGAGAAACCGGTACTTCCT), including enzyme digestion connector, and then connecting to the same digested entry vector pENTR1A (Invitrogen, USA). The pENTR1A-ERVT and pENTR1A-CIPK11 constructions were performed for LR recombination reaction with pEarlyGate103 expression vector, and then introduced into Arabidopsis accession Columbia (Col-0) via an Agrobacterium tumefaciens EHA105-mediated floral dip method[13]. Transformed progenies were screened by culturing on 1/2 MS medium agar plates containing 20 mg/mL glufosinate-ammonium and PCR detection. For tolerance evaluation, the 30 seeds of transgenic and wild-type plants were grown in each 1/2 MS medium agar plate containing 0 or 150 mM NaCl or 200 mM mannitol, respectively. The phenotype and survival rates of salt treatment and control plates were observed after 20 d, and the plants of drought stress treatment and control were measured after 12 d. For soil growth experimental, the uniformly growing seedlings of 10 d were planted in fritted clay-filled pot for 0 or 150 mM NaCl treatment (18 d) and watering or drought treatment (14 d). The phenotype and root length were detected. The growth condition was designed in a climate-controlled room with 12 h/d photoperiod, photosynthetically active radiation of 400 μmol photons m−2 s−1, day/night temperature of 25/20 °C and relative humidity of 60%.
Statistical analysis
-
A one-way analysis of variance (ANOVA) was performed to calculate whether treatment means were statistically different from one another (p = 0.05) using the SPSS v15.0 software. All analyses were performed based on three biological experiments and three technical replicates.
-
The RNA concentration and quantity reached 1,118 ng/μl and total 559 μg, presenting 2.16 of A260/A280 value (Supplemental Fig. S1). After mRNA extraction purification, A260/A280 value of mRNA was 2.05 and total mRNA quantity reached 5.42 mg (Supplemental Fig. S1), which was appropriate for library construction. Through 5'-capbody enrichment and recombinant technology, the cDNA entry library of pDONR/Zeo vector was successfully achieved. The results showed that the clone number of total entry library was approximately 9.6 × 106 (Supplemental Fig. S1), which could sufficiently represent most of genes expressed in alfalfa. Inserted gene sequence of random 24 clones was amplified and the fragment length ranged 0.6−4.0 kb with an average size at 1.35 kb (Supplemental Fig. S1). The expression library of pDEST52 vector was detected following the similar methods, revealing total 1.3 × 107 clones and 1.44 kb of average size (Supplemental Fig. S1).
Yeast transformation and identification of tolerant clones
-
Through yeast transformation in salt-sensitive G19 yeast competent cell, 18 salt-tolerant clones were picked and cultured overnight, and then different dilution products were grown on the control and salt-selection plates. The results showed 18 candidate salt-tolerant yeast grew more quickly than the yeast transformed by pDEST52 empty vector under culture medium containing 300 mM or 500 mM NaCl, while no differences were observed in the control plates (Fig. 1a). Similarly, 12 drought tolerant clones were selected and they all showed better growth than pDEST52 empty vector transformed yeast (Fig. 2a). The results indicated that the tolerances of these 30 yeast clones were dependent on the expressions of candidate tolerant genes of alfalfa.
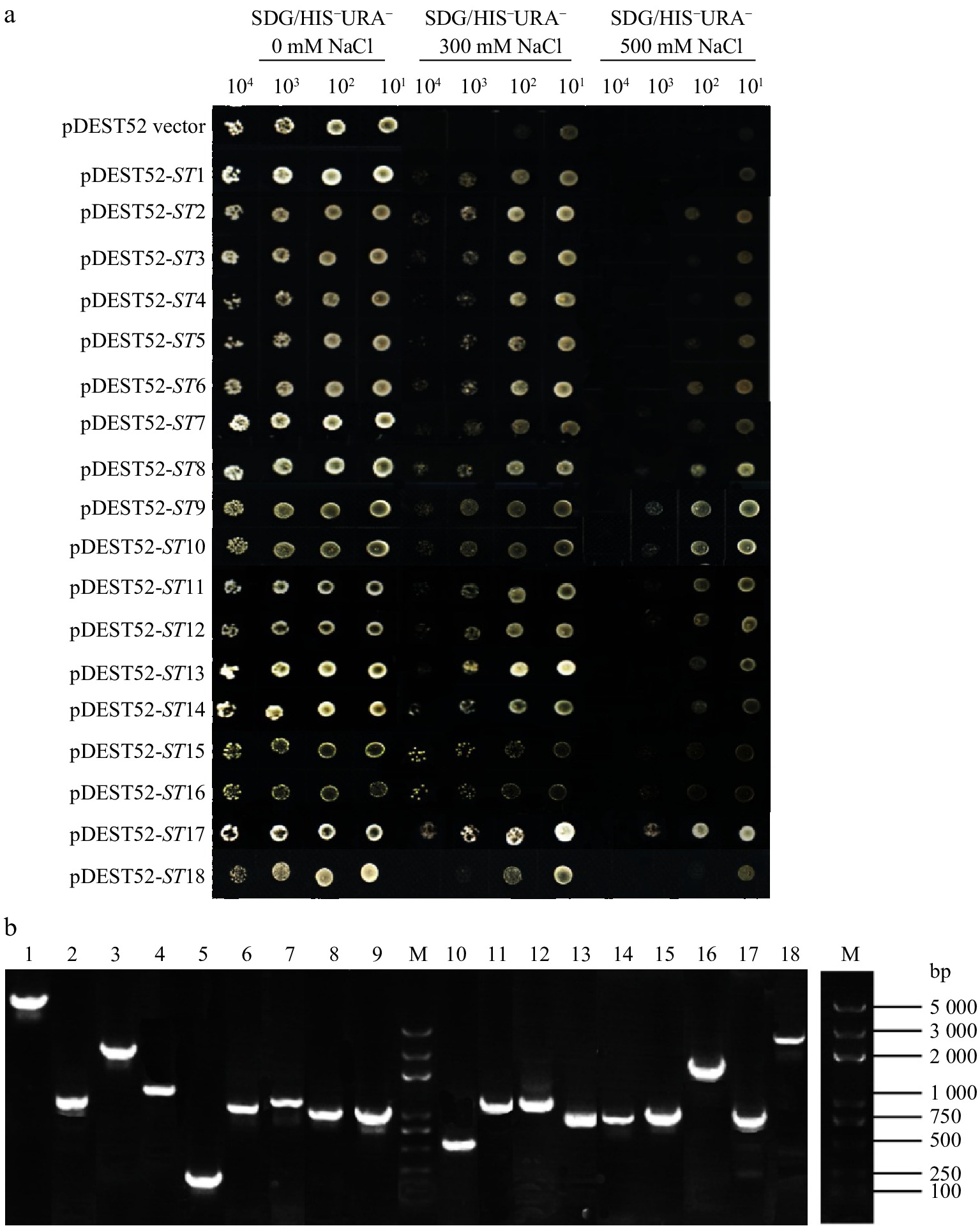
Figure 1. Salt-tolerance confirmation of 18 clones via library screening of alfalfa (a) and PCR detection of yeast plasmids (b).
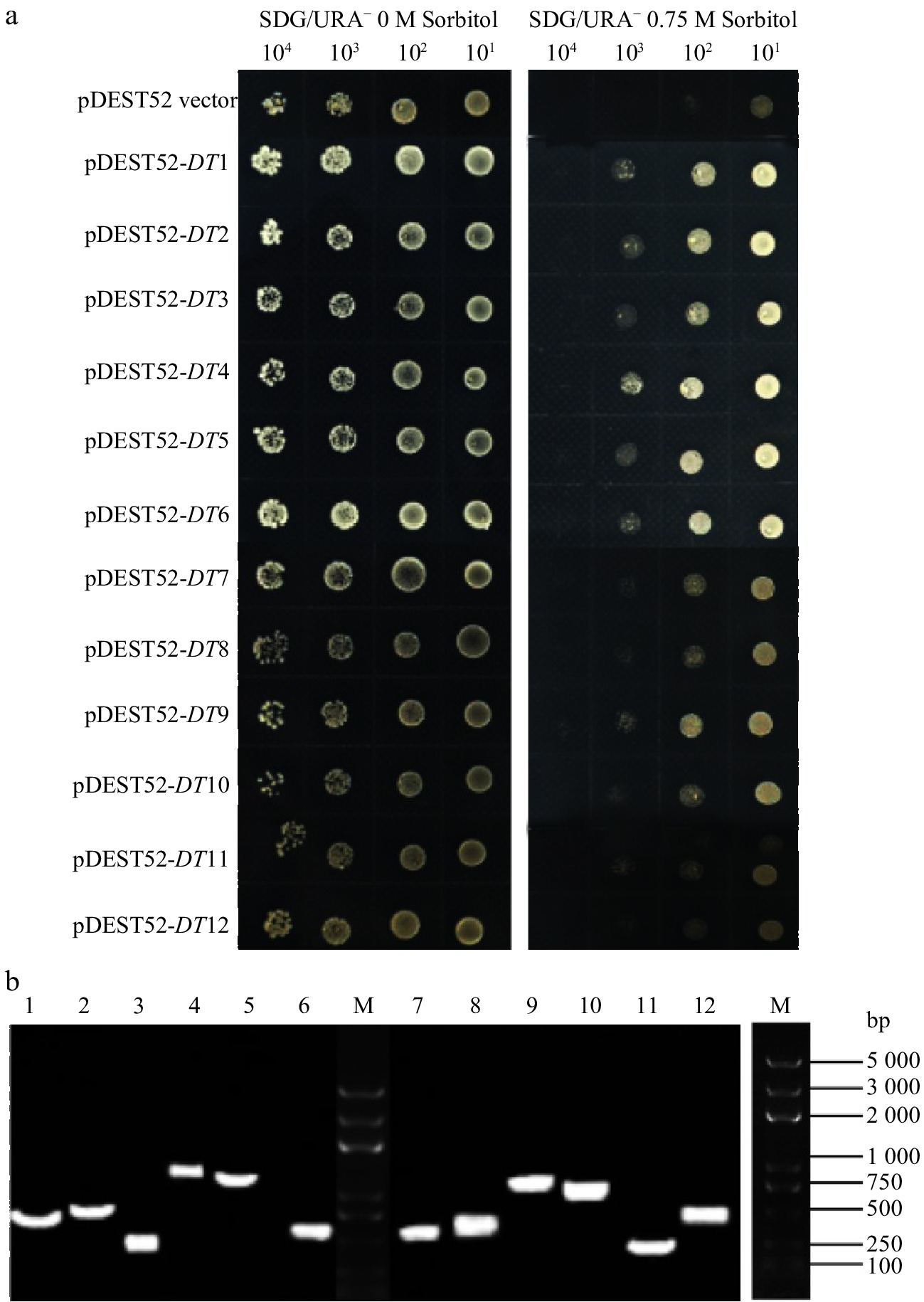
Figure 2. Drought-tolerance detection of 12 clones via library screening of alfalfa (a) and PCR analysis of yeast plasmids (b).
Plasmid rescue and sequence analysis of candidate tolerant genes
-
The plasmids of tolerant clones were isolated and detected, as shown in Fig. 1b and Fig. 2b. The nucleotide sequences of candidate tolerant genes were determined and analyzed (Table 1 & 2). The 18 salt-tolerant genes (Table 1) included one GRAS family transcription factor (ST4), five zinc finger proteins (ST7, ST9, ST13, ST14, ST17), seven proteases (ST2, ST5, ST8, ST11, ST12, ST15, ST18), one endoplasmic reticulum vesicle transporter protein (ST6), one transmembrane protein (ST16), one spatacsin carboxy-terminus protein (ST1) and one adaptor protein complex (ST3). The 12 drought tolerant genes (Table 2) participated in different biological function, containing ethylene-responsive transcription factor ERF071 (DT4), general transcription factor (DT10), three proteases (DT1, DT9, DT12), scarecrow-like protein 1 (DT2), lipase (DT5), peroxidase (DT3), Golgi transport (DT7), elongation factor (DT6), phosphoglucomutase (DT8) and Rubisco binding protein (DT11).
Table 1. Sequence analysis and function prediction of 18 salt tolerant candidate genes screened from cDNA expression library of alfalfa.
Salt tolerance clones Amino acid (aa) Protein function prediction GenBank accession number ST1 3167 Spatacsin carboxy-terminus protein (SCPT) MZ760872 ST2 358 L-allo-threonine aldolase-like protein (LA) MZ760873 ST3 1026 Adaptor protein complex AP-2, alpha subunit (acpAP-2) MZ760874 ST4 592 GRAS family transcription factor (GRAS) MZ760875 ST5 98 Cystatin (CYS1) MZ760876 ST6 353 Endoplasmic reticulum vesicle transporter protein (ERVT) MZ760877 ST7 409 Zinc finger CCCH domain protein (CCCH409) MZ760878 ST8 325 Alpha/beta hydrolase family protein (bHFP) MZ760879 ST9 325 Zinc finger, C3HC4 type (RING finger) protein (C3HC4) MZ760880 ST10 222 EARLY RESPONSE TO DEHYDRATION 14 (ERD14) MZ760881 ST11 403 PP2A regulatory subunit TAP46-like protein (TAP46) MZ760882 ST12 469 SCP1-LIKE SMALL PHOSPHATASE 4, SSP4 (SLP) MZ760883 ST13 299 Zinc finger B-box protein (ZFBP) MZ760884 ST14 379 Zinc finger CCCH domain protein (TZF1) MZ760885 ST15 326 Serine/threonine-protein phosphatase PP1 (PP1) MZ760886 ST16 724 Leucine zipper EF-hand transmembrane protein (LETM) MZ760887 ST17 377 Zinc finger CCCH domain protein (C3H49) MZ760888 ST18 901 Cytoplasmic-like aconitate hydratase (ACO1) MZ760889 Table 2. Sequence analysis and function prediction of 12 drought tolerant candidate genes screened from cDNA expression library of alfalfa.
Drought tolerance clones Amino acid (aa) Protein function prediction GenBank accession number DT1 422 CBL-interacting serine/threonine-protein kinase 11 (CIPK11) MZ760890 DT2 592 Scarecrow-like protein 1 (SCL1) MZ760891 DT3 326 Peroxidase 10 (POD10) MZ760892 DT4 259 Ethylene-responsive transcription factor ERF071 (ERF071) MZ760893 DT5 704 Triacylglycerol lipase 1 (TGL1) MZ760894 DT6 447 Elongation factor 1-alpha (GETP) MZ760895 DT7 265 Transport and Golgi organization 2 homolog (STT10) MZ760896 DT8 333 Cytoplasmic phosphoglucomutase (cPGM) MZ760897 DT9 274 Brassinosteroid-signaling kinase BSK7 (BSK7) MZ760898 DT10 281 General transcription factor IIE subunit 2 (TFIIE) MZ760899 DT11 350 Rubisco large subunit-binding protein subunit alpha, chloroplastic (Rubisco) MZ760900 DT12 432 Protease Do-like 1, chloroplastic (cPDL1) MZ760901 qRT-PCR analysis of candidate tolerant genes in response to salt or drought stress
-
Firstly, Ct values of six candidate reference genes (Supplemental Table S2) in all treated samples were obtained and their stabilities were assayed following the comprehensive analysis of RefFinder software. The results showed that EF1a and ACT exhibited the most stability under salinity stress, while EF1a and TUB revealed the most stable expression in drought treated samples (Supplemental Table S2). Therefore, we adopted EF1a for further qRT-PCR analysis of target genes in salinity or drought stressed samples.
The sequences of qRT-PCR primers were shown in Supplemental Table S3 and Supplemental Table S4, respectively. Salt treatment significantly induced the expression of 17 genes, except that CCCH409 (ST7) exhibited downregulation under salt stress (Fig. 3−5). Most salt-inducible genes reached a peak at 6 h and then declined, while C4HC4 (ST9) and ERD14 (ST10) genes revealed the highest expression at 3 h. Similarly, the expression of most drought tolerant genes appeared upregulated in response to PEG treatment, and only the expression of root CIPK11 (DT1) and the leaf Rubisco (DT11) decreased following PEG treatment (Fig. 6 & 7). These results indicated that the expression levels of screened tolerant genes were influenced by salt- or PEG-stress and these genes might function in stress tolerance regulation.
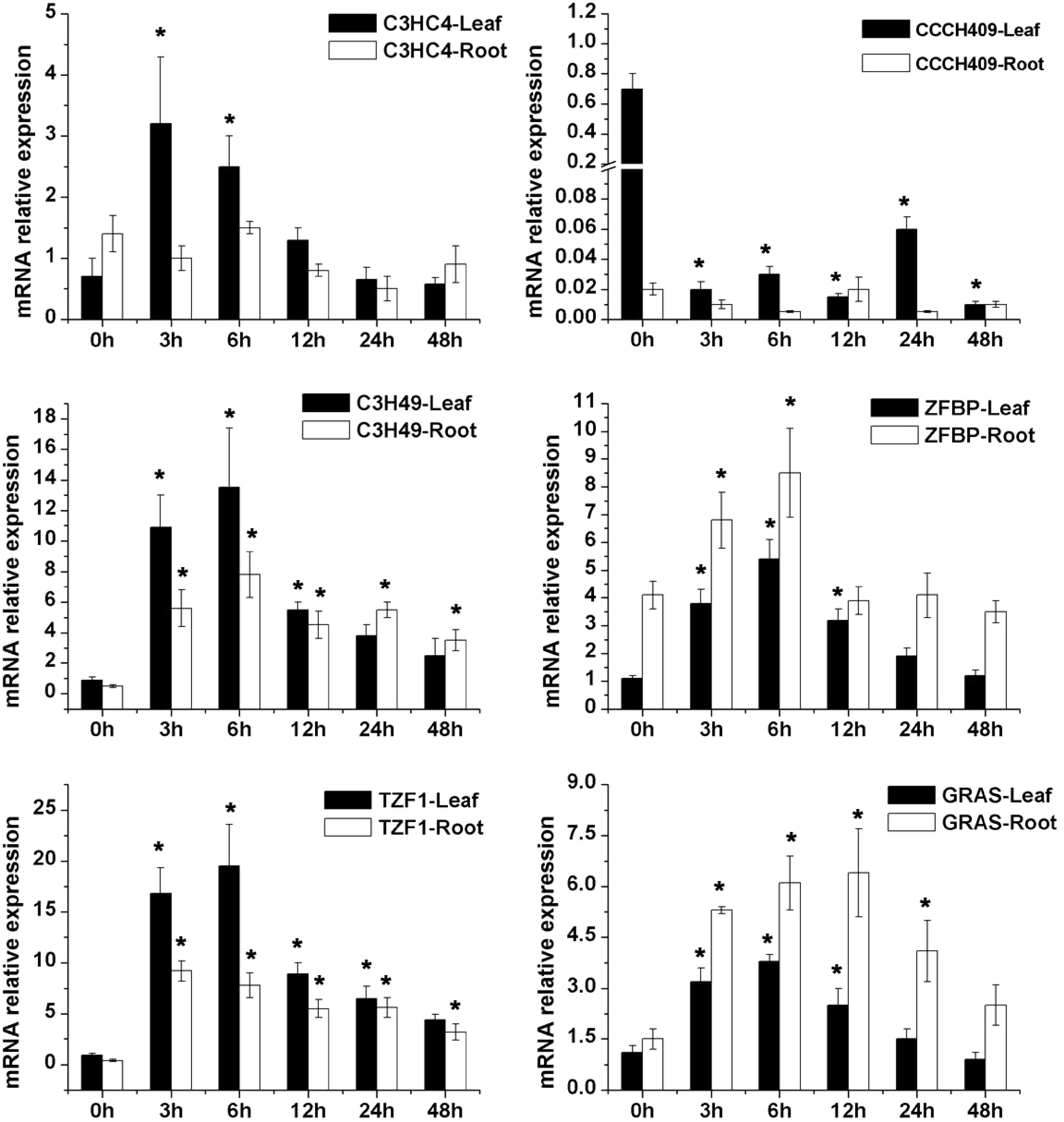
Figure 3. qRT-PCR analysis of six candidate salt-tolerant transcriptional factor of alfalfa including a GRAS family gene (GRAS) and five zinc finger protein (C3HC4, CCCH409, C3H49, ZFBP and TZF1) under 250 mM salt stress conditions. * Represent significant difference (p < 0.05) of salt stress samples for 3, 6, 12, 24 and 48 h compared to control sample (0 h), respectively.

Figure 4. qRT-PCR analysis of seven candidate salt-tolerant genes of alfalfa involved in protein phosphorylation modification (SLP, TAP46, PP1) and enzymes (LA, ACO1, bHFP and CYS1) under 250 mM salt stress conditions. * Represent significant difference (p < 0.05) of salt stress samples for 3, 6, 12, 24 and 48 h compared to control sample (0 h), respectively.

Figure 5. qRT-PCR analysis of five candidate salt-tolerant candidate genes of alfalfa participating in vesicle transporter (ERVT), transmembrane protein (LETM) and other genes (SCPT, ERD14, acpAP-2) under 250 mM salt stress conditions. * Represent significant difference (p < 0.05) of salt stress samples for 3, 6, 12, 24 and 48 h compared to control sample (0 h), respectively.
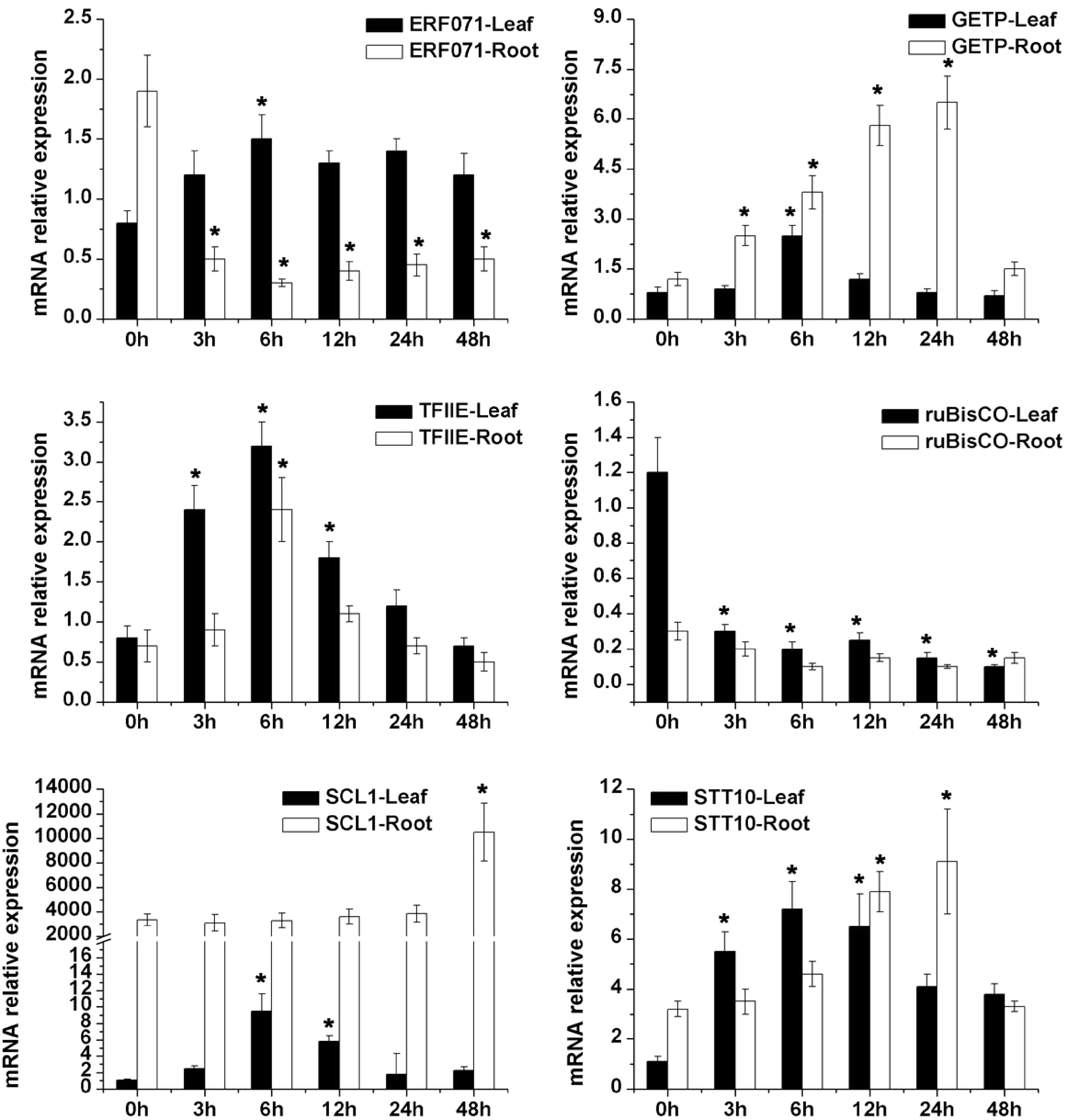
Figure 6. qRT-PCR analysis of six candidate drought-tolerant genes of alfalfa including transcriptional factor (ERF071, SCL, TFIIE, GETP), transport (STT10) and Rubisco under 20% PEG6000 simulated drought stress conditions. * Represent significant difference (p < 0.05) of drought stress samples for 3, 6, 12, 24 and 48 h compared to control sample (0 h), respectively.

Figure 7. qRT-PCR analysis of six candidate drought-tolerant genes of alfalfa including protein kinase (BSK7, CIPK11, cPGM), lipase (TGL1) and protease (cPDL1, POD10) under 20% PEG6000 simulated drought stress conditions. * Represent significant difference (p < 0.05) of drought stress samples for 3, 6, 12, 24 and 48 h compared to control sample (0 h), respectively.
Function identification of salt-tolerant ERVT and drought-tolerant CIPK11
-
To confirm the function of ERVT, three positive transgenic Arabidopsis plants (OX7, OX-8, OX-9) that overexpressed ERVT were used for salt tolerance identification. As shown in Fig. 8, the three transgenic lines exhibited better growth than the wild type plants under 150 mM NaCl stress. The survival rates and seedling growth of transgenic lines under salt stress were significantly higher than wild type plants (Fig. 8). In Fig. 9, we found three CIPK11-overexpressed transgenic plants showed higher survival rate and seedling growth under drought stress, comparing with wild type plants. These results indicated that ERVT or CIPK11 could enhance salt or drought tolerance respectively.
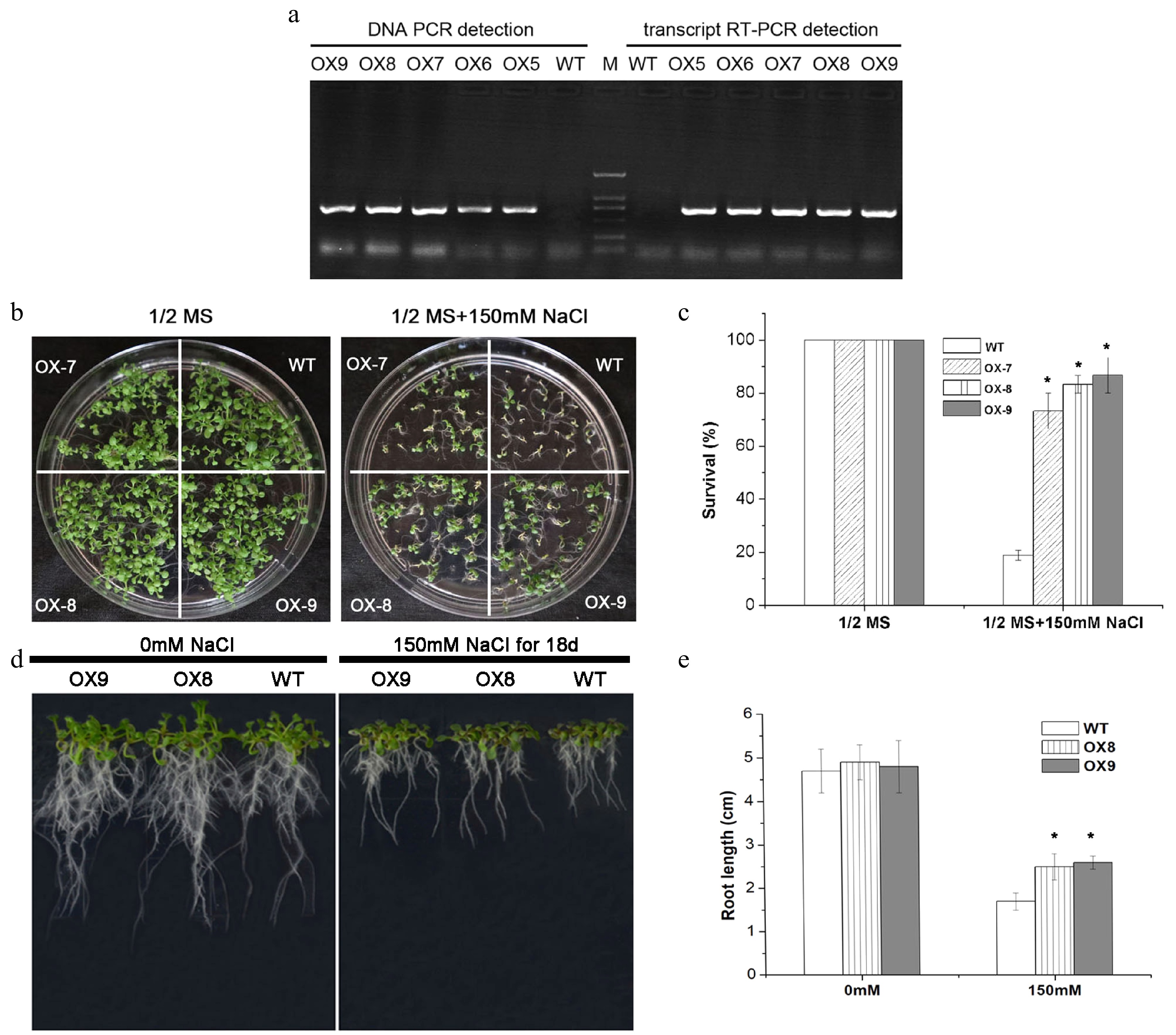
Figure 8. Salt-tolerance assays between wild type and ERVT transgenic lines of Arabidopsis. (a) Detection of PCR in DNA level and RT-PCR in mRNA level from wild type (WT) and transgenic plants (OX7, OX8, OX9). (b) Phenotype observation of plates. (c) Survival rate calculation under salinity stress with 0 and 150 mM NaCl. (d) Seedling growth in fritted clay under 0 and 150 mM NaCl treated conditions for 18 d. (e) Root length detection of seedlings. * Represent significant difference (p < 0.05) under salt stress compared to WT.

Figure 9. Drought-tolerance evaluation between wild type and CIPK11 transgenic lines of Arabidopsis. (a) Detection of PCR in DNA level and RT-PCR in mRNA level from wild type (WT) and transgenic plants (OX11, OX12, OX13). (b) Phenotype observation of plates. (c) Survival rate calculation under 0 and 200 mM mannitol simulated drought stress. (d) Seedling growth in fritted clay under normal watering and drought treated conditions for 14 d. (e) Root length detection of seedlings. * Represent significant difference (p < 0.05) under drought stress compared to WT.
-
Functional screening of cDNA expression library in yeast is a fast and effective strategy applied for excavation of stress tolerant genes, and some valuable functional genes were initially separated based on this system[1−6]. In this study, we also identified several candidate genes including known genes (TZF1, GRAS) and novel genes (ERVT, STT10). However, several stress tolerant marker genes such as SOS3, NHX1 and ERF1 were not screened in our work, likely due to the difference of these genes regulating pathway in plant and yeast. Therefore, FOX hunting system of model plants such as Arabidopsis or Medicago truncatula R108 will be used for identifying more salt- and drought-tolerant genes in our future research.
Candidate salt tolerant genes and their biological functions
GRAS and zinc finger protein
-
Previous studies demonstrated that plant transcription factor GRAS family members played multifarious roles in diverse physiological processes and environmental signals. A GRAS member BrLAS from Brassica rapa and rice OsGRAS23 were identified as a positive regulator of drought tolerance through increasing activities of antioxidant enzymes and decreasing accumulation of reactive oxygen species (ROS)[14,15]. JrGRAS2 contributed to improved heat tolerance in walnut by enhancing antioxidant systems and the expression of heat shock proteins (HSPs)[16]. GRAS gene of Poplar PeSCL7 and Vitis VaPAT1 conferred salt tolerance in transgenic Arabidopsis via modulating expression of stress-related genes[17,18]. In the present study, we obtained a GRAS family gene of alfalfa, which can be applied to salt-tolerant breeding as a potential target gene.
Zinc-finger protein (ZFP) is the largest family of DNA-binding transcription factors that also play essential roles in the eukaryote. Many CCCH-type zinc-finger proteins were identified for positively regulating plant salt stress tolerance through controlling the expression of stress-related genes, such as IbC3H18 of sweet potato[19], AtC3H17 of Arabidopsis[20], OsTZF1 of rice[21]. A recent study exhibited that a B-box zinc finger protein MdBBX10 from apple was found to be up-regulated under salt stress, and its overexpression could enhance plant salt tolerance by promoting the expression of ROS-scavenging genes[22]. In this study, we also separated a well-known salt tolerant gene TZF1 from alfalfa, which indicated the reliability of the FOX screening method. Simultaneously, four function-unknown zinc finger proteins were obtained and can be explored in future research.
Protease
-
The major plant serine/threonine protein phosphatases belong to the phosphoprotein phosphatase (PPP) family known to be composed of PP1, PP2 (PP2A), PP4, PP5, PP6, and PP7[23]. The latest research reveals that PP1 members are important regulators of plant immunity[24], no reports have been found demonstrating the relation between the PP1 family and salt tolerance. TAP46 is a protein phosphatase2A (PP2A)-associated protein that regulates PP2A activity and performs an essential function in plant growth and development[25]. A previous study discovered a PP2A-C5, the catalytic subunit 5 of PP2A, playing an important role in salt tolerance in Arabidopsis[26]. PP2A-B'γ can physically interact with the cytoplasmic form of aconitase hydratase (ACO), a central metabolic enzyme functionally connected with mitochondrial respiration, oxidative stress responses and regulation of cell death in plants[27]. However, whether TAP46-PP2A and PP2A-ACO pathways participate in salt-tolerant regulatory mechanism remains unclear. Our results provide several novel genes and research points for regulating plant salt tolerance.
A alfalfa Cystatin, a homologue of cystatin gene from sugar beet M14 enhancing plant salt tolerance, was also identified in this study[28]. Salt stress induced the alterations of Pol II-CTD phosphorylation at the target loci through SCP1-like small phosphatases (SSP4), indicating the correlation between SSP4 and salt tolerance[29]. Phytocystatins are plant proteins that inhibit cysteine proteinases, and show variable expression patterns during plant development as well as in response to abiotic stresses[30]. To date, a few α/β-hydrolase fold enzymes such as esterase, phospholipase D, OsPOP5 and IbMas have been shown to be involved in plant salt tolerance[31]. These candidate target genes separated from alfalfa can offer further explanation of the salt tolerance mechanism.
Transporter and transmembrane protein
-
Arabinogalactan protein-mediated vesicle trafficking from the apoplast to the vacuoles can contribute to sodium homeostasis during salt-adaptation to high salinity concentrations[32]. Arabidopsis synaptotagmin1 is enriched in endoplasmic reticulum-plasma membrane contact sites and confers cellular resistance to mechanical stresses[33]. Leucine zipper EF-hand transmembrane 1 (LETM1) superfamily proteins were proven to play roles in regulating mitochondrial-produced ROS in deoxynivalenol biosynthesis and the accumulation of mitochondrially encoded proteins[34,35]. Rice OsCCD1, a calcium-binding protein with one EF-hand motif, can enhance the tolerances to osmotic and salt stress[36]. Athb-12, a homeobox-leucine zipper domain protein from Arabidopsis, increases salt tolerance in yeast by regulating sodium exclusion[37]. Two ER vesicle transporter (ERVT) proteins (LB111423 and LB103860) and the vesicle-associated membrane protein family were enriched in the salt gland of L. bicolor[38]. We found two candidate salt tolerant genes encoding endoplasmic reticulum vesicle transporter protein (ERVT) and leucine zipper EF-hand transmembrane protein (LETM) of alfalfa induced by salt stress. In addition, we found ERVT could improve salt tolerance through genetic transformation in Arabidopsis, which indicated the reliability of the yeast screening strategy and it would be worth pursuing.
Candidate drought tolerant genes and their biological functions
Transcription factor
-
Previous investigations indicated that different ERF members might play diverse functions in plant responses to both abiotic and biotic stresses[39]. ERF74 acts as a switch controlling a RbohD dependent mechanism in response to different stresses and subsequently maintains ROS homeostasis in Arabidopsis[40]. Recent research revealed an ERF transcription factor TINY modulated brassinosteroid-regulated plant growth and drought responses in Arabidopsis[41]. The ERF transcription factor MdERF38 of apple can promote drought stress induced anthocyanin biosynthesis[42]. One hundred and fiftynine ERF genes were identified from alfalfa and four ERF members (MsERF013, MsERF089, MsERF136, MsERF140) were induced in expression level under drought stress[43]. We also found a drought stress-inducible ERF071 which positively regulated drought tolerance of yeast. These results exhibited the important regulatory pathways of ERF factors mediating drought stress tolerance.
SCARECROW-LIKE transcription factors (SCL) provided a major function for plant growth and development. The SCL23 regulatory module is also essential for specification of the endodermis (known as the bundle sheath) in leaves[44]. SCL3 acts as a positive regulator to influence the GA pathway by attenuating the DELLA repressors in the root endodermis[45]. SCL27 is critical for mediating GA-DELLA signaling in the coordinate regulation of chlorophyll biosynthesis and leaf growth under light[46]. The tomato DELLA protein PROCERA can promote stomatal closure and reduce water loss under water deficit stress[47]. We screened a SCL family factor SCL1 of alfalfa controlling drought tolerance, implying the candidate SCL-DELLA regulatory mechanism of drought tolerance.
Kinase, lipase and phosphoglucomutases
-
The CBL-CIPK signaling system is a basic calcium sensor that plays an important role in sensing adverse environmental stimuli. Wheat TaCIPK23 and TaCIPK10 positively regulates drought stress and resistance to Pst (Puccinia striiformis f. sp. tritici)[48,49]. Overexpression of pineapple AcCBL1 resulted in enhanced tolerance to salinity, osmotic, and fungal stress[50]. Apple MdCIPK13 and MdCIPK22 was required for the phosphorylation of MdSUT2.2 protein and then could enhance its stability and transport activity, promote sugar accumulation and improve salt and drought tolerance[51,52]. The plant brassinosteroid-signaling kinase (BSK) proteins belong to a family of receptor-like cytoplasmic kinases, and perform important roles in BR signal transduction and environmental adaptation[53]. Arabidopsis loss-of-function mutant bsk5 exhibited increased sensitivity to salinity and ABA[54]. However, the functions of BSK family members regulating osmotic stress tolerance are still unclear. The recent study showed that the concentrations of BR increased significantly with the extension of the drought period, revealing the relevance of BR signal and drought response[55]. Our results concluded that the CIPK and BSK pathways may participate in drought tolerance regulation of alfalfa.
Lipases are classified into phospholipases, lipolytic acyl hydrolases, galactolipases and triacylglycerol lipases based on the lipid substrates catabolized[56]. Several previous reports showed phospholipases, lipolytic acyl hydrolases and galactolipases can contribute to drought stress adaption[57−59]. A patatin domain triacylglycerol lipase (TGL) encoded by SDP1 can initiate storage oil breakdown in germinating Arabidopsis seeds[60], while little has been explored in the drought tolerance of TGL-type ligase, a candidate TGL-type ligase member found in this study will help further this research.
Phosphoglucomutases (PGM) belong to the phosphohexomutases superfamily and are highly specific in catalyzing the interconversion of glucose 1-phosphate (G-1-P) to glucose 6-phosphate (G-6-P). The lack of cPGM activity inhibited the growth of rosette and roots, resulting in reduced seed production[61]. Drought stress significantly induced the transcriptional expression of Chloroplastic protease Do-like (cPDL)[62]. However, the biological function of cPGM and cPDL mediated drought tolerance is unknown.
The expression pattern of these candidate tolerant genes
-
In previous studies[9, 10, 12], several candidate stress-tolerant genes were selected according to transcription expression of gene mRNA response to stress conditions. However, not all gene transcription regulated by stress had stress-tolerant functions. In this study, we firstly screened the candidate tolerant genes via yeast transformation system successfully applied in several plants, and then detected their mRNA transcriptional level response to salt or osmotic stress conditions. The expression changes of these genes could provide a foundation for tolerance function and further research ideas based on the upstream regulation network.
-
For the molecular explanation of alfalfa response to salt and drought stress, a highly efficient FOX hunting system was applied to identify tolerant genes. We obtained 18 potential salt-tolerant genes and 12 candidate drought-tolerant genes, including several known tolerant genes for representing the reliability for our experimental results, and novel unknown pathways for future research. Our results exhibited that transcription factors (GRAS and zinc finger), protein phosphorylation (PP2A regulatory module), vesicle trafficking and transmembrane transport might participate in salt-tolerant regulation of alfalfa. The transcription factors (ERF and SCL), protein phosphorylation (CIPK and BSK), TGL-type lipases and PGM-mediated glucose interconversion might contribute to the regulatory network of drought stress response in alfalfa. We also transformed ERVT and CIPK11 into the model-plant Arabidopsis and confirmed salt-tolerance and drought-tolerance function, indicating the effectiveness of the yeast screening approach. These conclusions can enrich the regulatory mechanism of plant abiotic tolerance and also provide new insights for molecular breeding of alfalfa.
- We thank Dr. Huazhong Shi (Department of Chemistry and Biochemistry, Texas Tech University) for providing yeast strains G19 (MATa ade2 ura3 leu2 his3 trp1 ena1::HIS3::ena4Δ). Hog1 mutant YSH444 (MATα leu2-3/112 ura3-1 trp1-1 his3-11/15ade2-1 can1-100 GAL SUC2 hog1::TRP1 mutant) is provided by Dr. Stefan Hohmann (University of Gothenburg). This work was supported by the program of National Natural Science Foundation of China (NSFC, 32071876, 31672193) and Open-end Funds of State Key Laboratory of Grassland Agro-ecosystems (SKGAE), Lanzhou University, China.
- The authors declare that they have no conflict of interest.
- Supplemental Table 1 Primer sequence of 6 reference genes used for stability analysis.
- Supplemental Table 2 Most stable and least stable combination of reference genes based on RefFinder analysis.
- Supplemental Table 3 qRT-PCR primer pairs of 18 salt tolerant candidate genes in alfalfa.
- Supplemental Table 4 qRT-PCR primer pairs of 12 drought tolerant candidate genes in alfalfa.
- Supplemental Fig. S1 Expression library procedure of alfalfa. (A) Alfalfa “No.2 gongnong”; (B) Analysis and detection of total RNA extracted from alfalfa mixed sample (roots, stems and leaves) by 1% Agarose gel electrophoresis; (C) Analysis and detection of mRNA purified from total RNA of alfalfa by 1% Agarose gel electrophoresis; (D) titer and inserted fragment detection of entry cDNA library; (E) titer and inserted fragment detection of pDEST52 expression cDNA librar.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Z, Zou Y, Li Y, Sun C, Liu Y, et al. 2021. Exploring the salt- and drought-tolerant genes of alfalfa through expression library screening strategy. Grass Research 1: 11 doi: 10.48130/GR-2021-0011

Exploring the salt- and drought-tolerant genes of alfalfa through expression library screening strategy
- Received Date: 11 May 2021
- Accepted Date: 15 November 2021
- Published Online: 29 November 2021
Abstract: Salinity and drought stress are major abiotic stresses negatively affecting the growth and productivity of alfalfa (Medicago sativa L.). Exploration of genes exhibiting superior tolerance to salinity and drought stress in alfalfa will help aid target molecular breeding and developing tolerant forages. In this study, we adopted a high-efficient yeast FOX hunting system for the identification of salinity and drought tolerant genes in alfalfa. Based on the Gateway-compatible vector system, a high-quality expression library was constructed, containing 1.3 × 107 clones with an average size of 1.44 kb. Through heterologous transformation of mixed library plasmid into salt or drought sensitive yeast mutants, monoclonal resistant strains were screened and tolerant genes were captured. Eighteen salinity-tolerance genes were obtained which were involved in several pathways, containing GRAS and zinc finger transcriptional factors, PP2A interaction module, ERVT vesicle transporter and LETM transmembrane protein. Twelve drought tolerance genes were separated, including ERF and SCL transcriptional factors, CIPK and BSK protein kinases, TGL-type ligase, cPGM and cPDL protease. The mRNA transcription levels of these tolerant genes were inducible or suppressible for response to salt or drought stress conditions following quantitative PCR detection, respectively. Furthermore, heterologous transformation of ERVT and CIPK11 can improve the salt and drought stress tolerance in Arabidopsis, which indicates the conservative function of the screening gene in yeast and Arabidopsis. Obtaining these candidate genes can provide new insights for future research with respect to plant salt and drought tolerance.
-
Key words:
- Alfalfa /
- yeast /
- cDNA library /
- salinity tolerance /
- drought tolerance