-
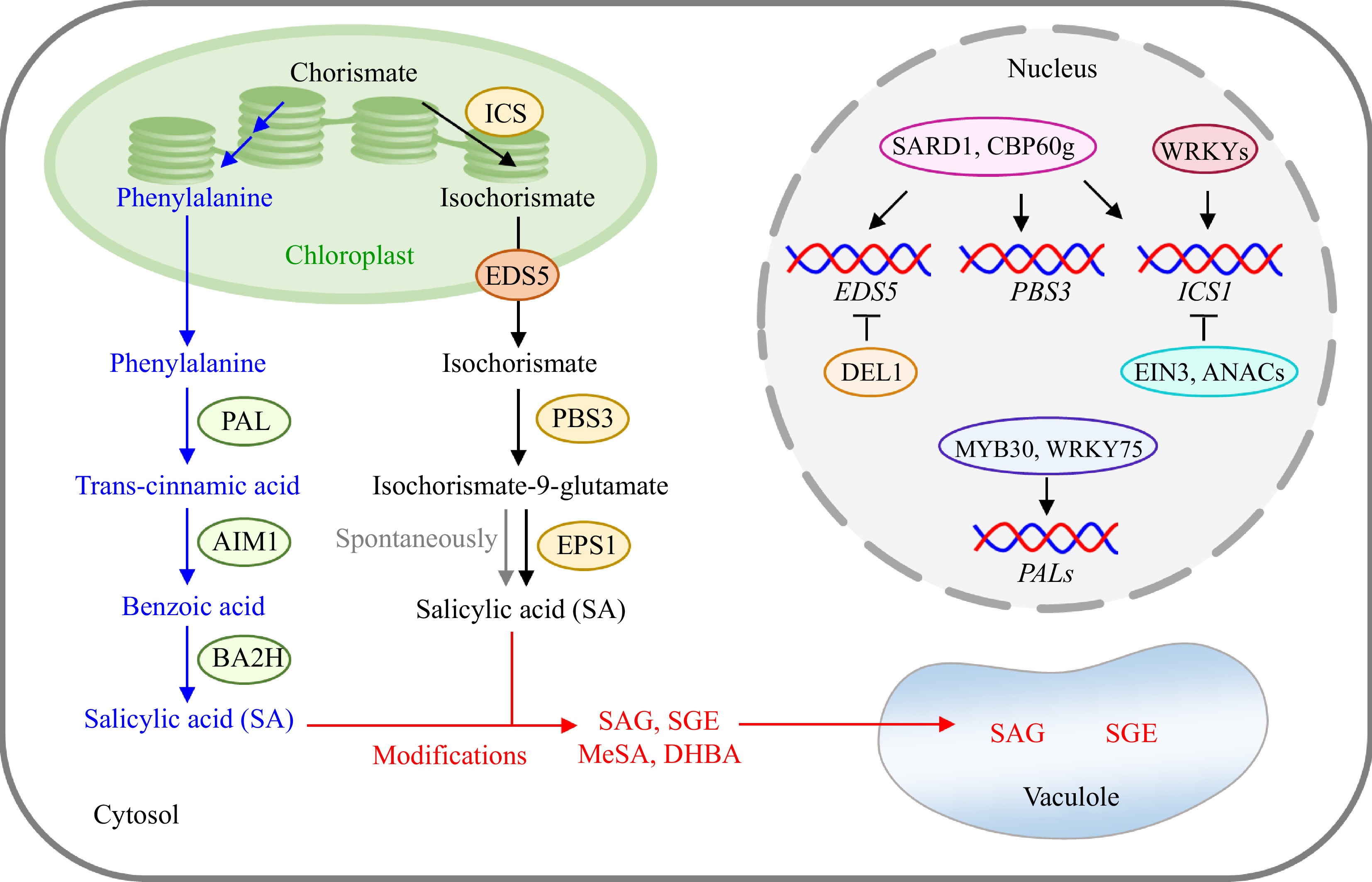
Figure 1.
Biosynthesis, metabolism, and regulation of SA in plants. Plants employ the precursor chorismate for SA production through two biosynthetic pathways mediated by ICS and PAL, respectively. On the one hand, SA is generally metabolized or conjugated to form other compounds (such as SAG, SGE, and MeSA) under various types of modifications to maintain SA at the basal level in vivo. SAG and SGE can be further stored in the vacuole. On the other hand, the biosynthesis of SA is also precisely regulated by many transcription factors. For instance, SARD1 and CBP60g are key regulators of ICS1, PBS3, and EDS5. The expression of ICS1 is also enhanced by some WRKYs and inhibited by EIN3 and ANACs. DEL1 is a well-documented repressor of EDS5. Additionally, PALs can be positively regulated by MYB30 in rice and WRKY75 in Tulip. PAL, phenylalanine ammonia-lyase; AIM1, abnormal inflorescence meristem 1; BA2H, benzoic acid-2-hydroxylase; ICS, isochorismate synthase; EDS5, Ehanced Disease Susceptibility 5; PBS3, AvrPphB Susceptible 3; EPS1, Ehanced Pseudomonas Susceptibility 1; SAG, SA glucoside; SGE, SA glucose ester; MeSA, methyl salicylate; DHBA, dihydroxybenzoic acid; SARD1, SAR-deficient 1; CBP60g, Calmodulin binding protein 60-like g; EIN3, ethylene insensitive 3; DEL1, atypical DP-E2F-like 1.
-
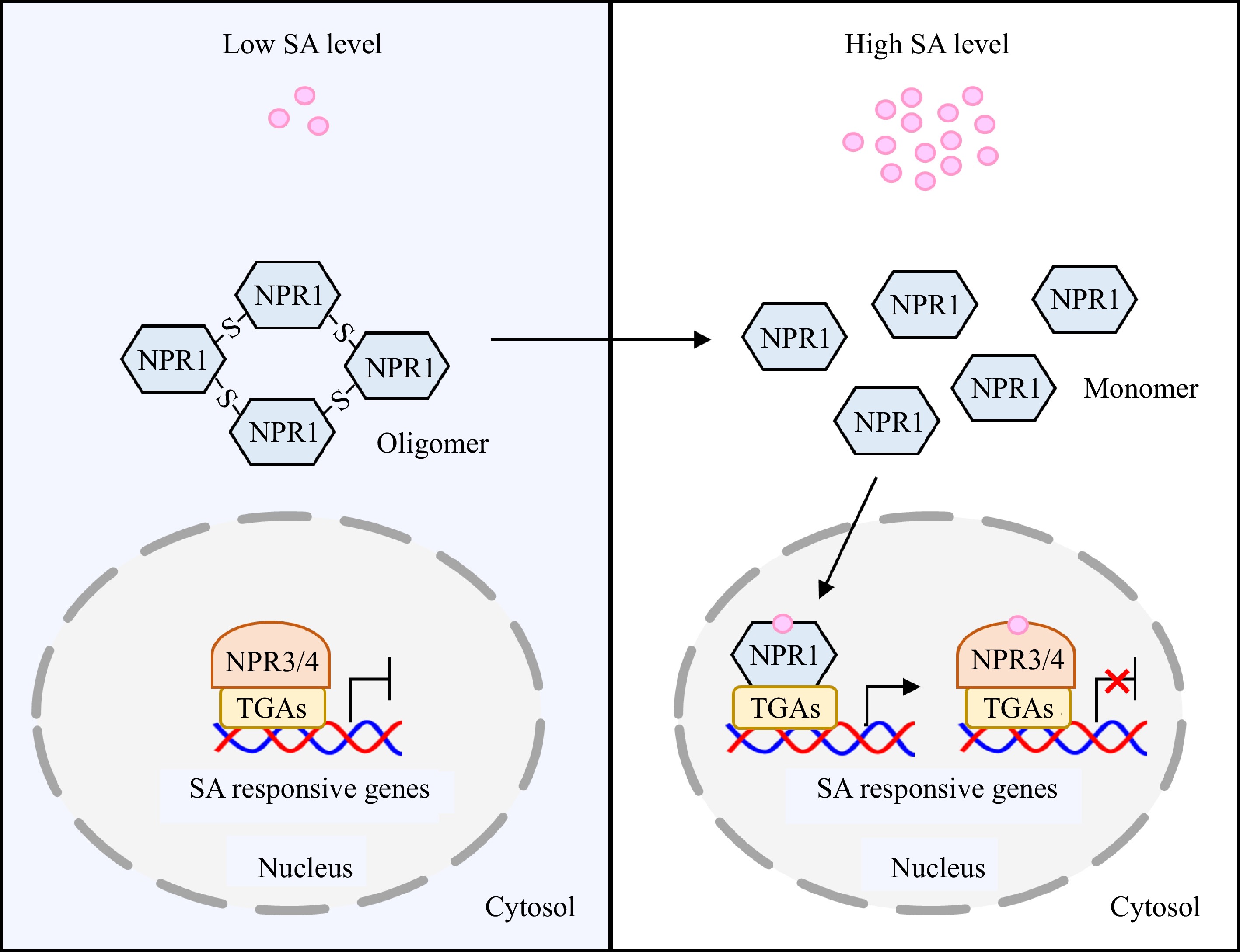
Figure 2.
NPRs-mediated SA signaling pathway in plants. At low intracellular SA level, NPR1 presents in the cytosol in its oligomeric form, while NPR3/4 interact with TGA transcription factors and inhibit the expression of SA-responsive genes. When intracellular SA level is elevated, the redox state transition induced by SA accumulation can trigger the conformational change of NPR1 from oligomer to monomer. The monomeric form of NPR1 enters into nucleus, and then interacts with TGAs to activate expression of SA-responsive genes. Meanwhile, SA binds to NPR3/4 and represses their activities, thereby alleviating the inhibitory effect of NPR3/4-TGAs on SA-responsive genes. NPR, nonexpressor of pathogenesis-related protein.
-
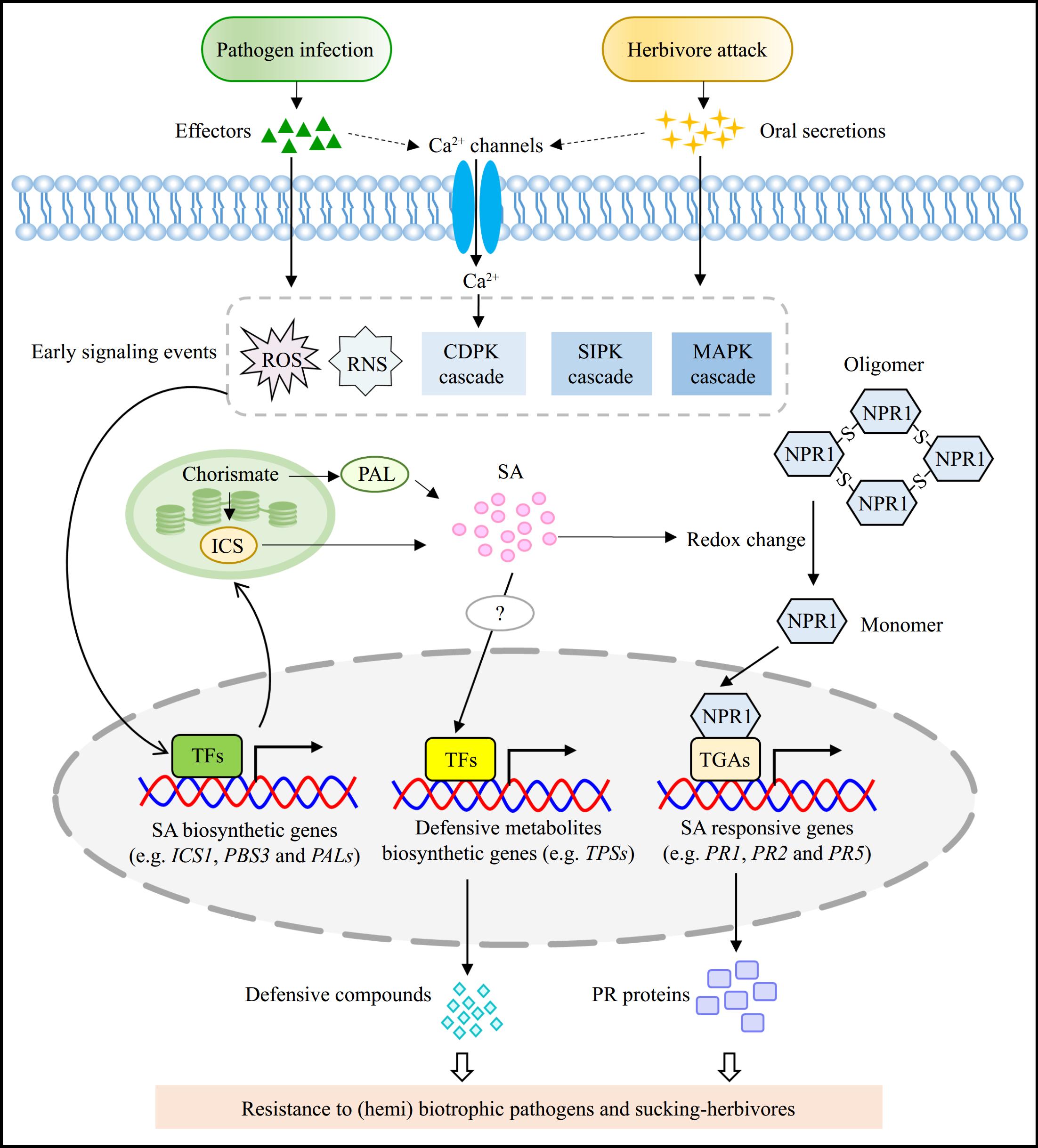
Figure 3.
SA-mediated defense responses against pathogens and herbivores. When pathogens and herbivores invade plants, they usually produce effectors and oral secretions, respectively. These effectors and oral secretions generally elicit a range of early signaling events, including the burst of ROS and RNS, an increase of intracellular calcium level, and the activation of several protein kinase-mediated cascades. These early signaling events subsequently activate the TFs to promote the expression of SA biosynthetic genes, thereby enhancing SA accumulation. The elevated intracellular SA level induces the redox change that triggers the transition of NPR1 from oligomer to monomer, allowing it to enter the nucleus and interact with TGAs to stimulate the expression of SA-responsive genes (e.g. PR1, PR2, and PR5). Additionally, SA accumulation also promotes the production of defensive metabolites, probably by activating TFs that can directly target to the promoter of biosynthetic genes associated with these defensive metabolites. However, the precisely regulatory mechanism of SA-mediated production of defensive metabolites remains to be investigate. These defensive metabolites and PR proteins contribute to improve the resistance of plants to (hemi)biotrophic pathogens and sucking-herbivores. ROS, reactive oxygen species; RNS, reactive nitrogen species; CDPK, Calcium-dependent protein kinase; SIPK, SA-induced protein kinase; MAPK, mitogen-activated protein kinase; TFs, transcription factors; PR, pathogenesis-related protein.
Figures
(3)
Tables
(0)