-
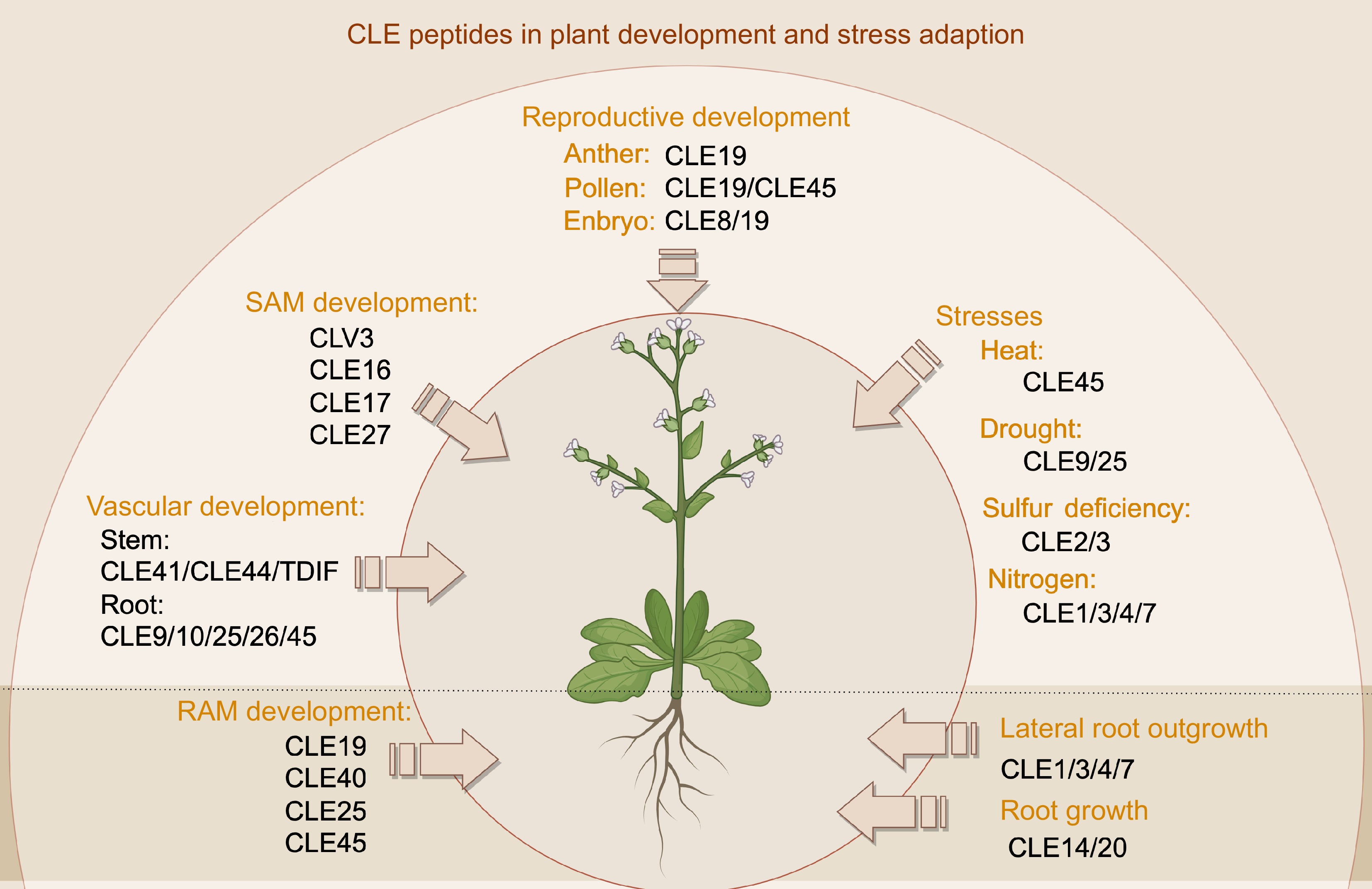
Figure 1.
CLE peptides' functions in plant development and stress adaption. CLE peptides play critical roles in both tissue development and stress response processes. They regulate key aspects of meristem maintenance, vascular and reproductive development, while also mediating plant adaptation to environmental stresses such as drought and temperature fluctuations.
-
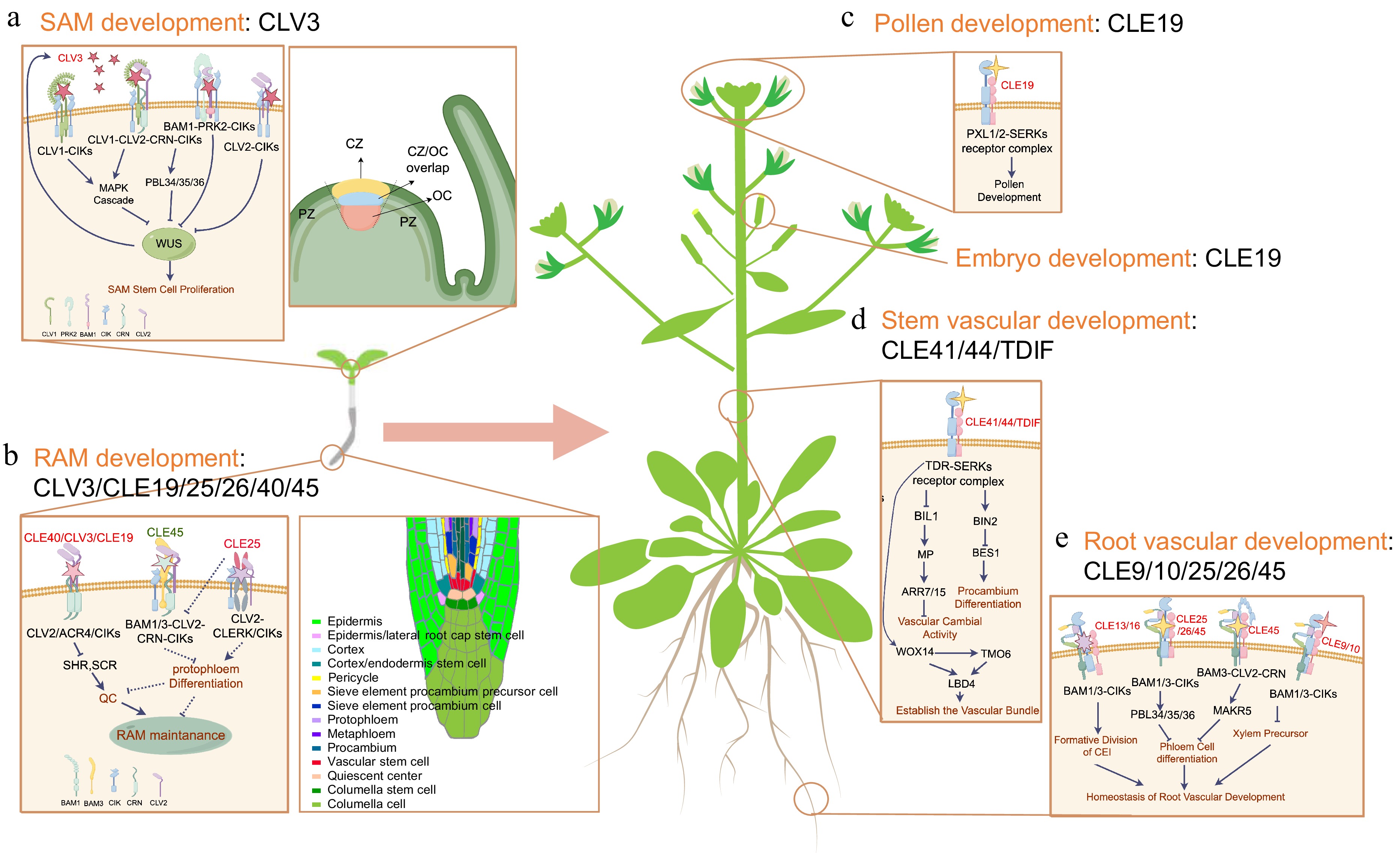
Figure 2.
CLE signaling pathways in regulating plant development. (a) CLV3-WUS feed-back regulatory loop in the organization of shoot apical meristem (SAM). Mature CLV3 peptide binds to the plasma membrane (PM)-localized receptor-coreceptor complexes to restricts WUS expression through the MAPK cascade, the receptor like cytoplasmic kinases PBS1-LIKE34/35/36 (PBL34/35/36), and other unknown downstream pathways. (b) CLE signaling pathways involved in root apical meristem (RAM) maintenance. The RAM is composed of quiescent center (QC) cells, stem cells surrounding the QC, and different cell layers derived from stem cells. CLE40/CLV3/CLE19 induce terminal RAM differentiation through CLV2-CRN receptors. CLE45 suppresses protophloem differentiation through the BAM1/3-CLV2-CRN-CIKs receptors and finally promotes RAM maintenance. CLE25 promotes the protophloem differentiation through CLV2/CLERK(CIKs) receptors and finally represses RAM maintenance. Moreover, CLE25 suppresses the activity of CLE45 receptors through OPS. These CLEs and related signaling pathways together maintain the RAM homeostasis. (c) CLE19 regulates pollen development through its PXL1/2-SERKs receptor-coreceptor complexes. (d) The CLE41/44/TDIF-TDR-SERKs peptide-receptor-coreceptor complex controls stem vascular development by coordinating the proliferation of vascular cells and establishing the precise position of vascular boundaries. CLE41/44/TDIF-TDR signaling interacts directly with the GSK3 family member BRASSINOSSTEROID-INSENSITIVE 2 (BIN2), and inhibits the activity of transcription factor BES1 to prevent procambium differentiation into xylem. Additionally, it inhibits the activity of another GSK3 family member BIN2-Like 1 (BIL1), which phosphorylates and activates the auxin response factor MONOPTEROS (MP), and subsequently induces the expression of ARR7 and ARR15 to repress vascular cambial activity. The WOX4-TMO6-LBD4 feed-forward loop acts downstream of CLE41/44/TDIF-TDR signaling to establish the vascular boundary. (e) CLE9/10/25/26/45 regulates the root vascular development through multiple receptor-coreceptor complexes, including BAM1/3-CIKs, BAM3-CLV2-CRN, and other downstream signaling components.
-
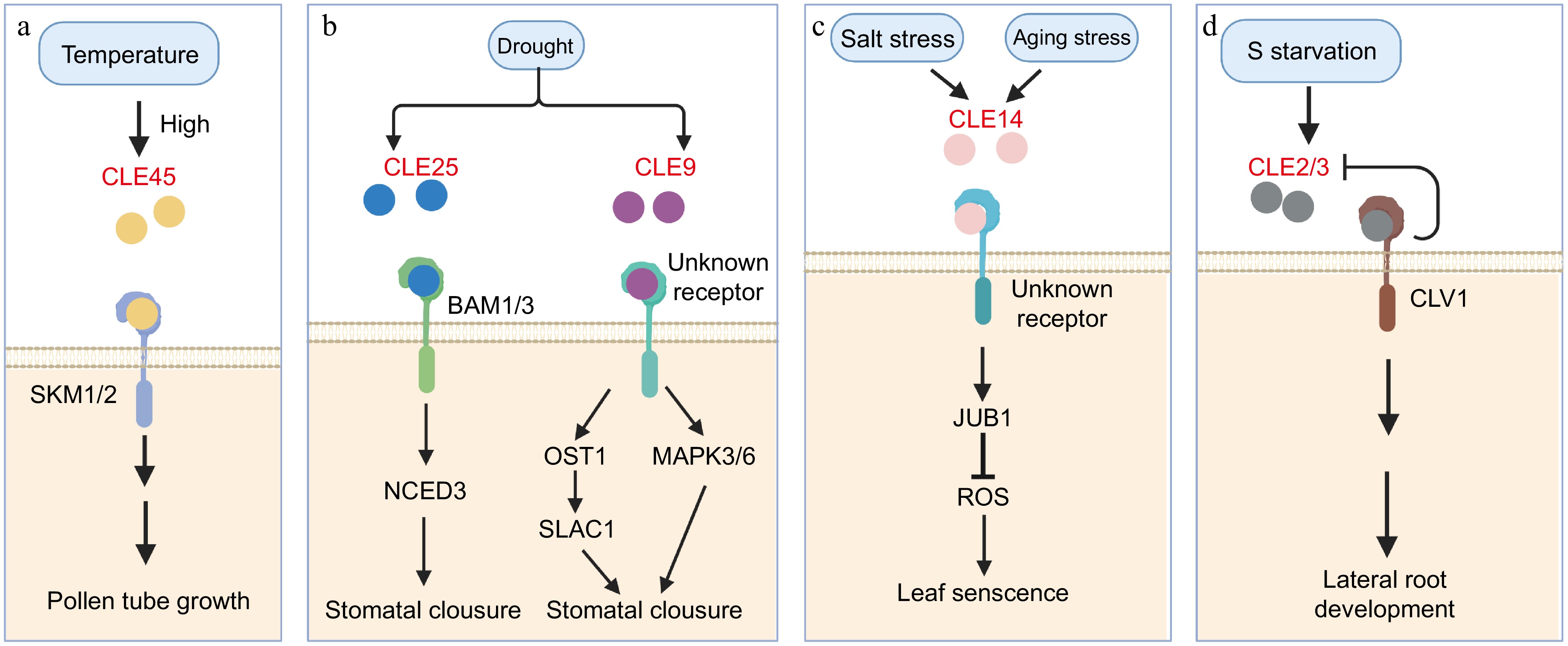
Figure 3.
CLE signaling pathway under different abiotic stresses. (a) The CLE45 peptide is perceived by the SKM1/2 receptors under high temperatures, regulating pollen tube growth and maintaining pollen viability. (b) The CLE25 peptide is perceived by BAM1/2 receptors, which regulate the NCED3 enzyme to modulate stomatal closure in response to drought stress. Additionally, CLE9 also responds to drought stress through unknown receptors, governing stomatal closure via distinct downstream signaling cascades. (c) CLE14 mediates responses to salt and aging stress by interacting with unknown receptors, regulating the transcription factor JUB1. JUB1 negatively regulates ROS levels, thereby influencing leaf senescence. (d) Sulfur (S) starvation induces CLE2/3 peptides, which are perceived by CLV1 to regulate lateral root development. In a feedback loop, CLV1 suppresses CLE2/3 expression to maintain root growth homeostasis.
Figures
(3)
Tables
(0)