-
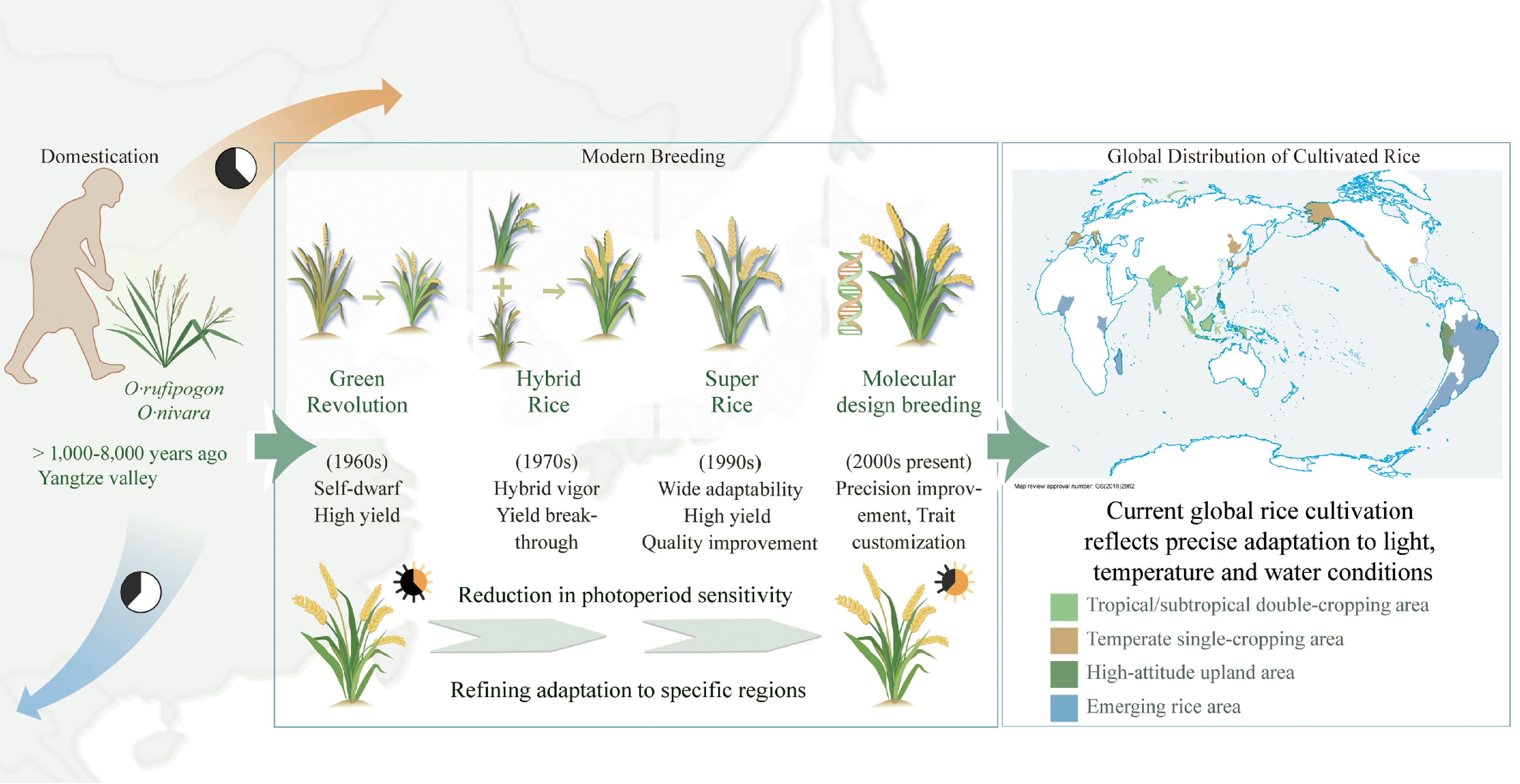
Figure 1.
From natural selection to artificial breeding: evolutionary adaptation and global expansion of rice. Rice has evolved from its wild progenitors (O. rufipogon and O. nivara) through successive breeding phases, including the Green Revolution (semi-dwarf, high yield), hybrid rice (heterosis utilization), super rice (yield and quality improvement), and molecular design breeding (trait-level precision). This evolutionary trajectory reflects a gradual reduction in photoperiod sensitivity and enhanced regional adaptability. The map illustrates current global rice-growing regions, including tropical and subtropical double-cropping, temperate single-cropping, high-altitude upland, and emerging areas, demonstrating rice's precise ecological adaptation to light, temperature, and water conditions.
-
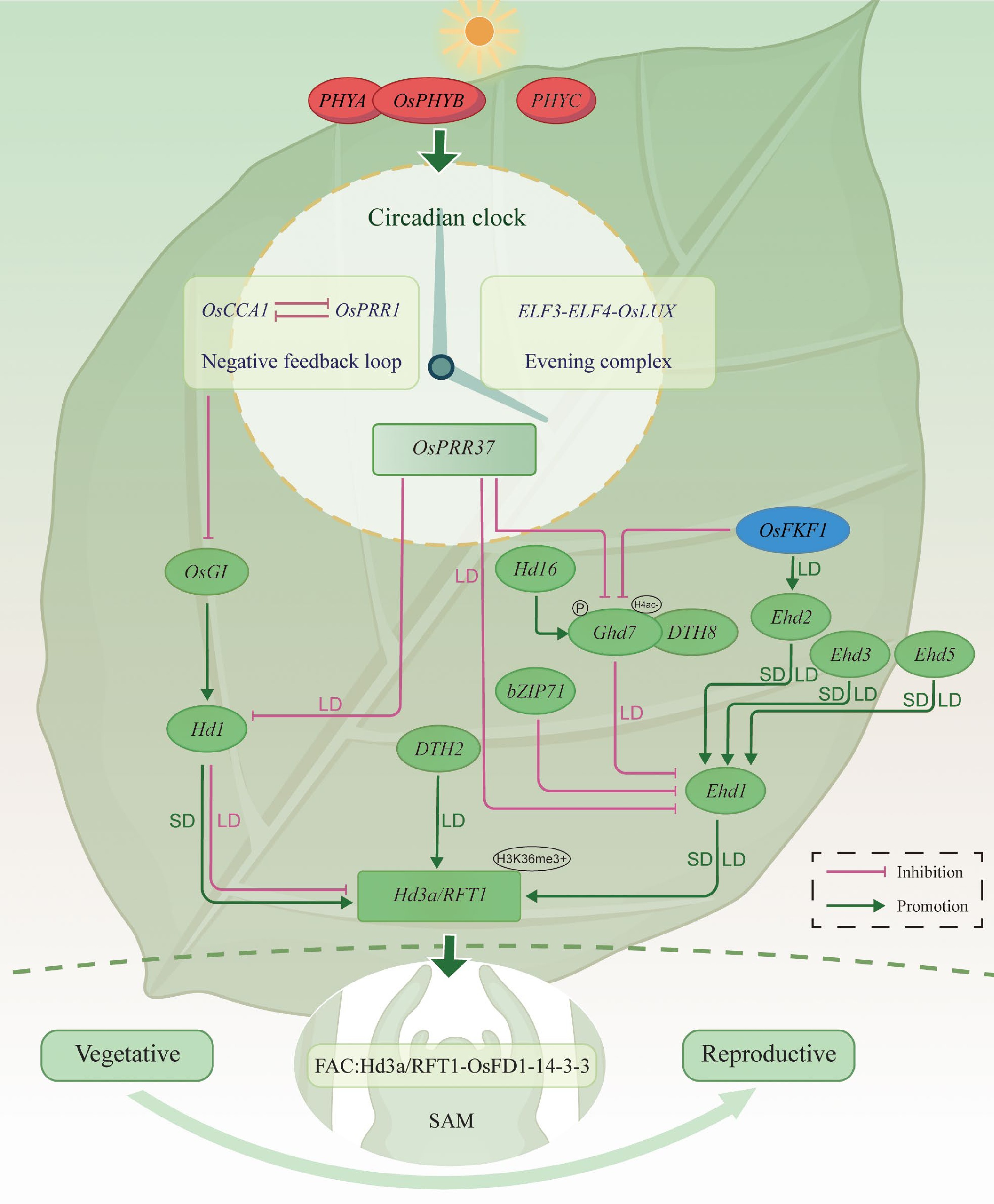
Figure 2.
Integrated regulatory network linking light signaling, circadian rhythm, and photoperiodic flowering in rice. Photoreceptors (PHYA, OsPHYB, PHYC) entrain the circadian clock (the OsCCA1-OsPRR1 negative loop and the ELF3-ELF4-OsLUX evening complex), which routes signals into the OsGI-Hd1-Hd3a/RFT1 and Ghd7-Ehd1-Hd3a/RFT1 axes. Small SD/LD labels on edges indicate predominant effects under short-day (SD) or long-day (LD): Hd1 activates Hd3a/RFT1 in SD but represses in LD; Ghd7, OsPRR37/DTH7, and DTH8 predominantly repress Ehd1 in LD; DTH2 promotes Hd3a/RFT1 under LD; Ehd2/3/5 promote Ehd1. Protein-protein interactions are overlaid, including the florigen activation complex (FAC; Hd3a/RFT1-14-3-3-OsFD1) at the SAM. Protein post-translational modification control is indicated by Hd16/CKI-dependent phosphorylation that potentiates Ghd7 repression under LD. Chromatin badges mark H3K36me3+ (activation) at Hd3a/RFT1 and H4ac− (deacetylation/raised repression threshold) at Ghd7. Green arrows denote promotion; magenta blunt lines denote repression. (Hd3a is SD-biased, whereas RFT1 is more LD-permissive).
-
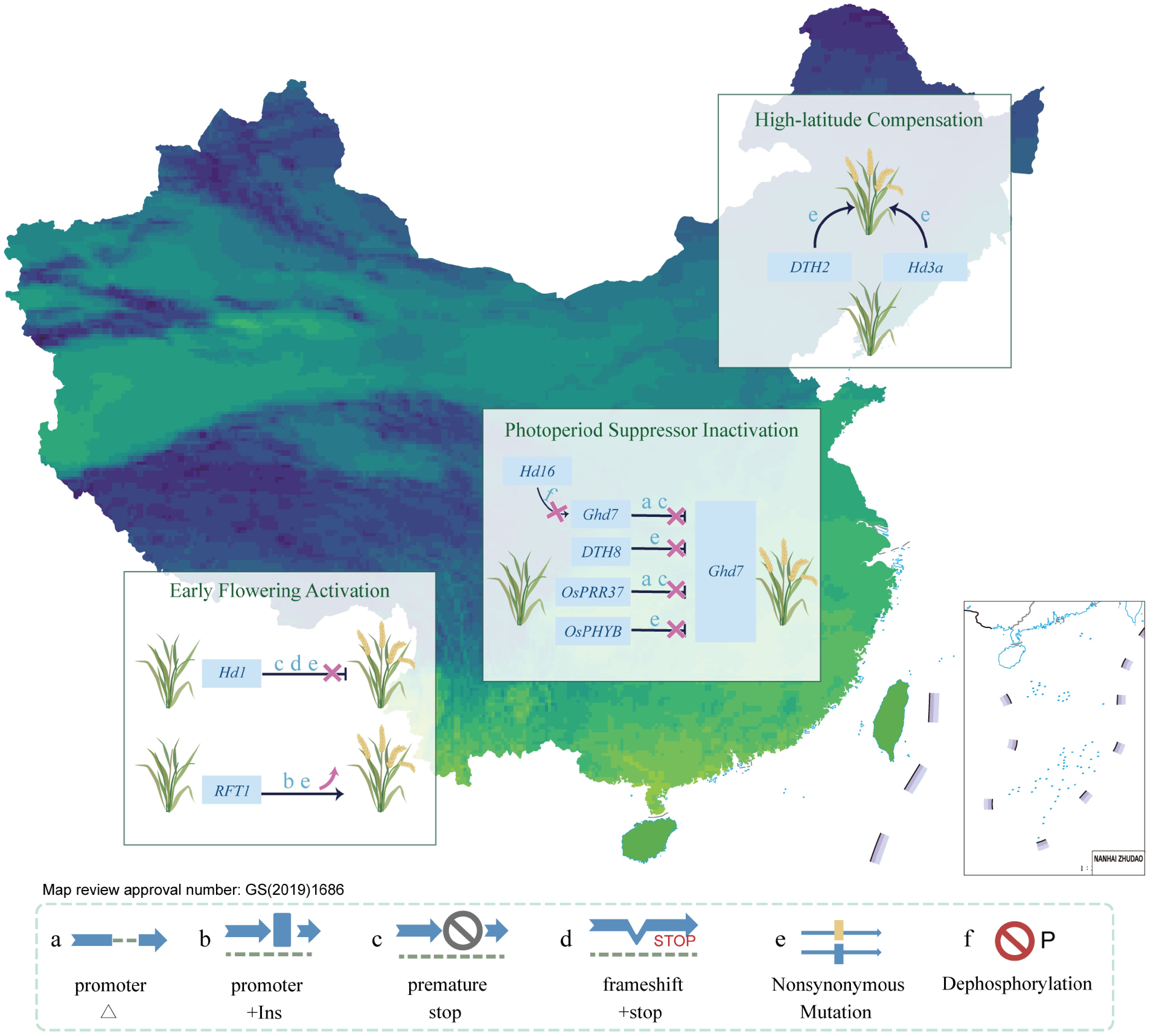
Figure 3.
Region-specific adaptation of rice flowering driven by regulatory gene variation. Region-specific allelic variation underlies the adaptive differentiation of heading date across China. In southern regions, loss-of-function mutations in Hd1 and upregulation of RFT1 promote early flowering under long-day conditions. In central areas, inactivation of repressors (Ghd7, DTH8, OsPRR37, OsPHYB) relieves photoperiod sensitivity. In northern high-latitude zones, compensatory mechanisms involving DTH2 and Hd3a ensure timely flowering. Major mutation types include: a, promoter deletion; b, promoter insertion; c, premature stop; d, frameshift + stop; e, nonsynonymous mutation; f, dephosphorylation site loss. These variations form the molecular basis for latitudinal flowering adaptation.
-
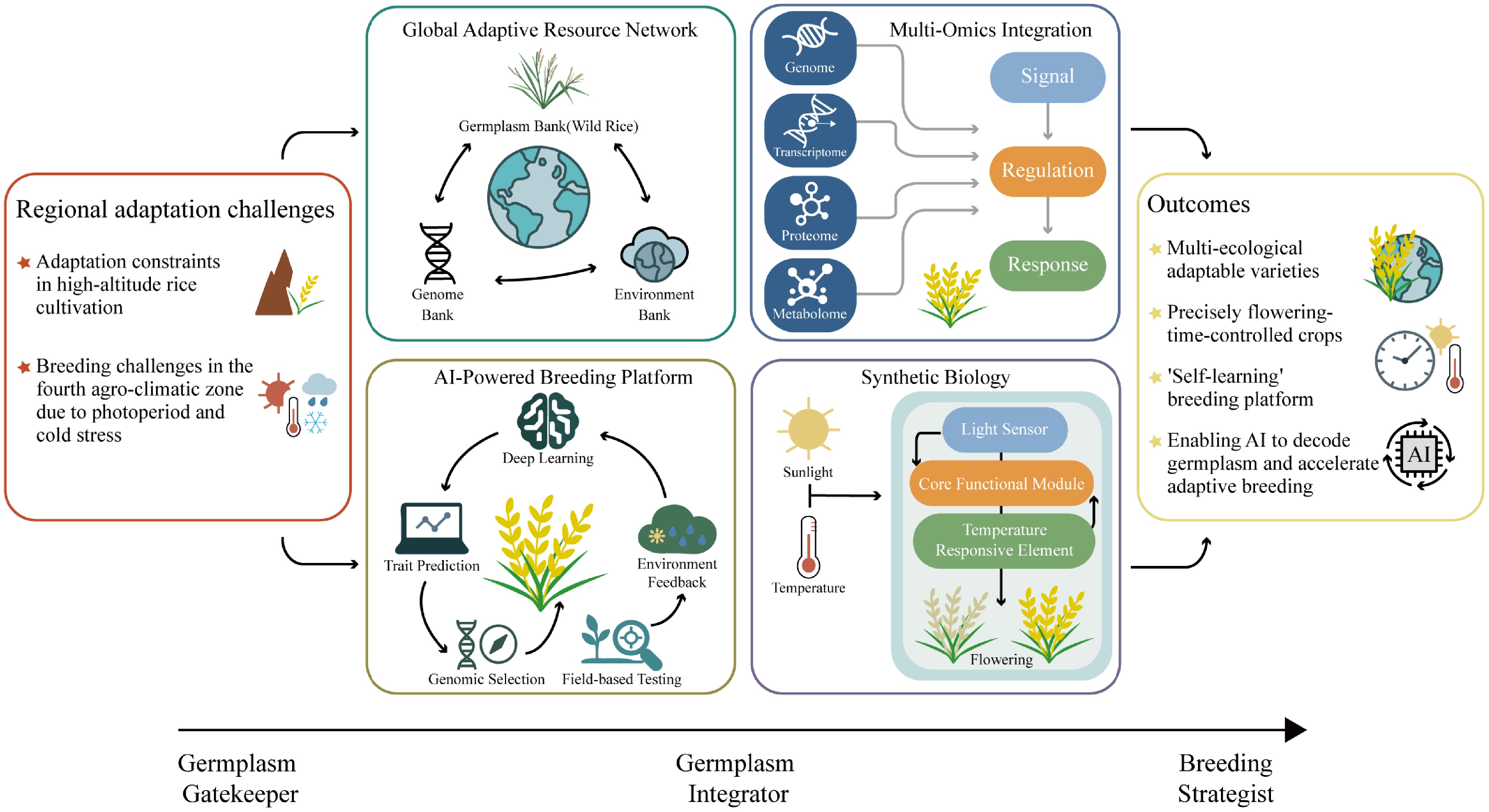
Figure 4.
A strategic roadmap for smart and climate-resilient rice breeding. The roadmap highlights a transition from germplasm gatekeeper to strategic, technology-enabled breeding. Central components include the construction of a global adaptive resource network that links germplasm, genomic, and environmental data; multi-omics integration to decode signal-regulation-response pathways; AI-powered breeding platforms for predictive selection and field feedback; and synthetic biology approaches for programmable control of flowering traits. Together, these components support the development of intelligent, adaptive, and future-oriented rice breeding systems.
Figures
(4)
Tables
(0)