-
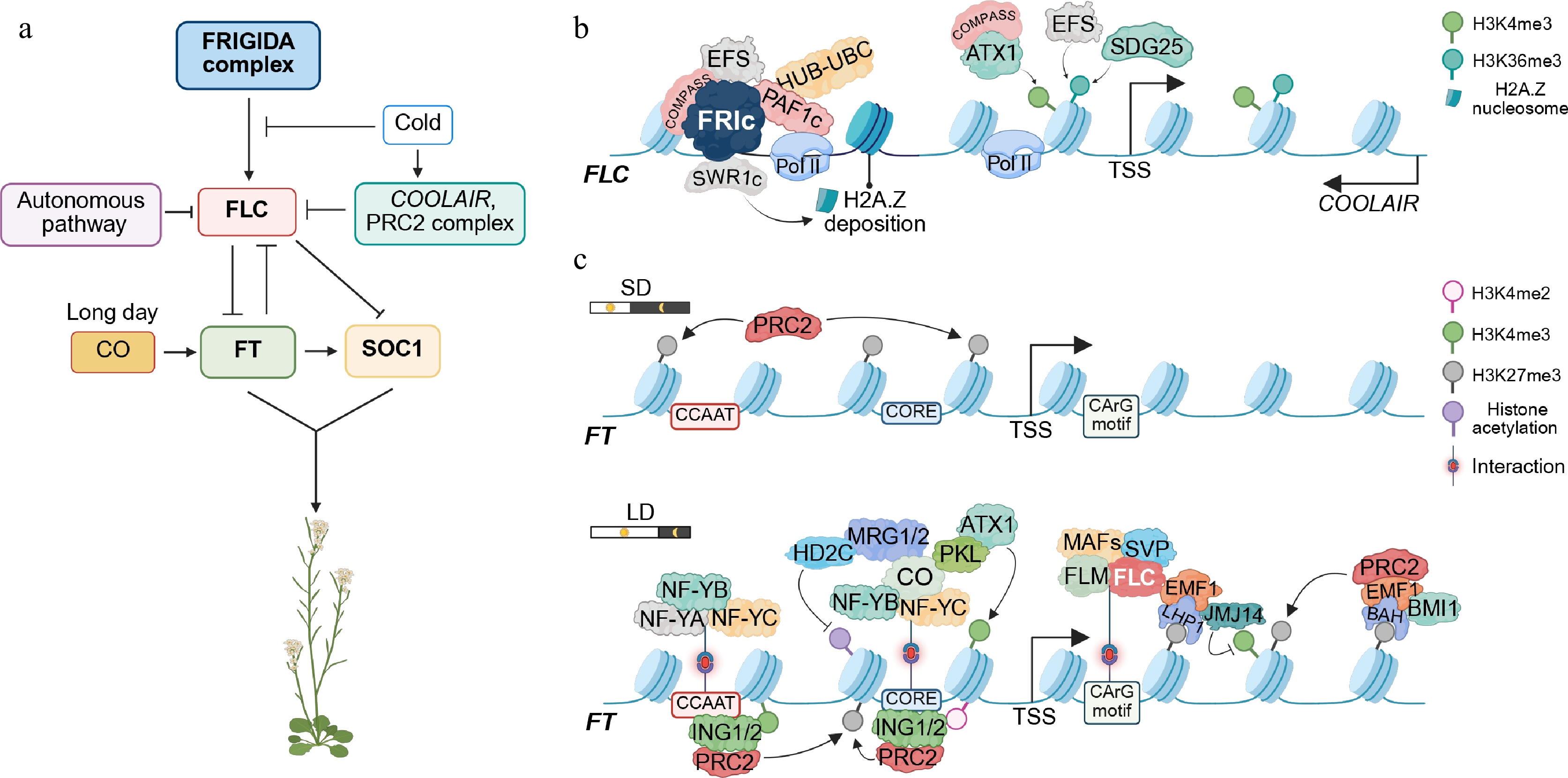
Figure 1.
Multilayered epigenetic regulation of the flowering genes FLC and FT in Arabidopsis. (a) Genetic circuitry overview. The FRIGIDA (FRI) complex delays flowering by activating the transcription of FLOWERING LOCUS C (FLC). Prolonged cold exposure (vernalization) induces COOLAIR, which silences FLC in parallel with Polycomb repressive complexes 2 (PRC2). The repression of FLC enables transcription of FLOWERING LOCUS T (FT), which subsequently upregulates the expression of SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1), APETALA1 (AP1), LEAFY (LFY), and SEPALLATA3 (SEP3), thereby specifying floral meristem identity and floral organ development. (b) FRIGIDA-dependent activation of FLC. The FRI complex (FRIc) recruits multiple chromatin remodelers, including the COMPASS-like complex (mediating H3K4me3, trimethylation of lysine 4 on histone H3), EARLY FLOWERING IN SHORT DAYS (EFS, also known as SDG25, a histone lysine methyltransferase responsible for H3K36me3), PAF1 complex-HUB-UBC module (PAF1c-HUB-UBC, mediating H2B mono-ubiquitylation), and the SWR1 chromatin remodeling complex (SWR1c, responsible for H2A.Z deposition), all of which establish a chromatin environment conducive to the activation of FLC expression. (c) Dynamic regulation of FT. Under short-day (SD) conditions, FT expression is constitutively repressed by Polycomb group (PcG) complexes, and thus, the floral transition is inhibited. Under long-day (LD) conditions, a trimeric NUCLEAR FACTOR Y (NF-Y) complex (composed of NF-YA, NF-YB, and NF-YC subunits) first binds to the CCAAT motif to increase chromatin accessibility. CONSTANS (CO) together with NF-Y binds the CORE element and recruits ARABIDOPSIS TRITHORAX 1 (ATX1), MORF-RELATED GENE 1/2 (MRG1, MRG2), and the chromatin remodeler PICKLE (PKL, a CHD3-type chromatin remodeler), depositing the activating mark H3K4me3 and inducing FT expression. Following FT activation at dusk, INHIBITOR OF GROWTH 1 (ING1) and ING2 recognize H3K4me2/me3 modifications on FT chromatin and recruit PRC2 to re-establish repression at night and into the early afternoon the next day. Conversely, the MADS-box proteins SHORT VEGETATIVE PHASE (SVP) and FLC occupy the CArG box, and Polycomb repressive complexes catalyze H3K27me3 deposition, repressing FT transcription. The key histone modifications are indicated on the right. Arrows denote positive effects; blunt lines denote repressive roles. Images were created by BioRender,
https://BioRender.com/6r3d71u . -
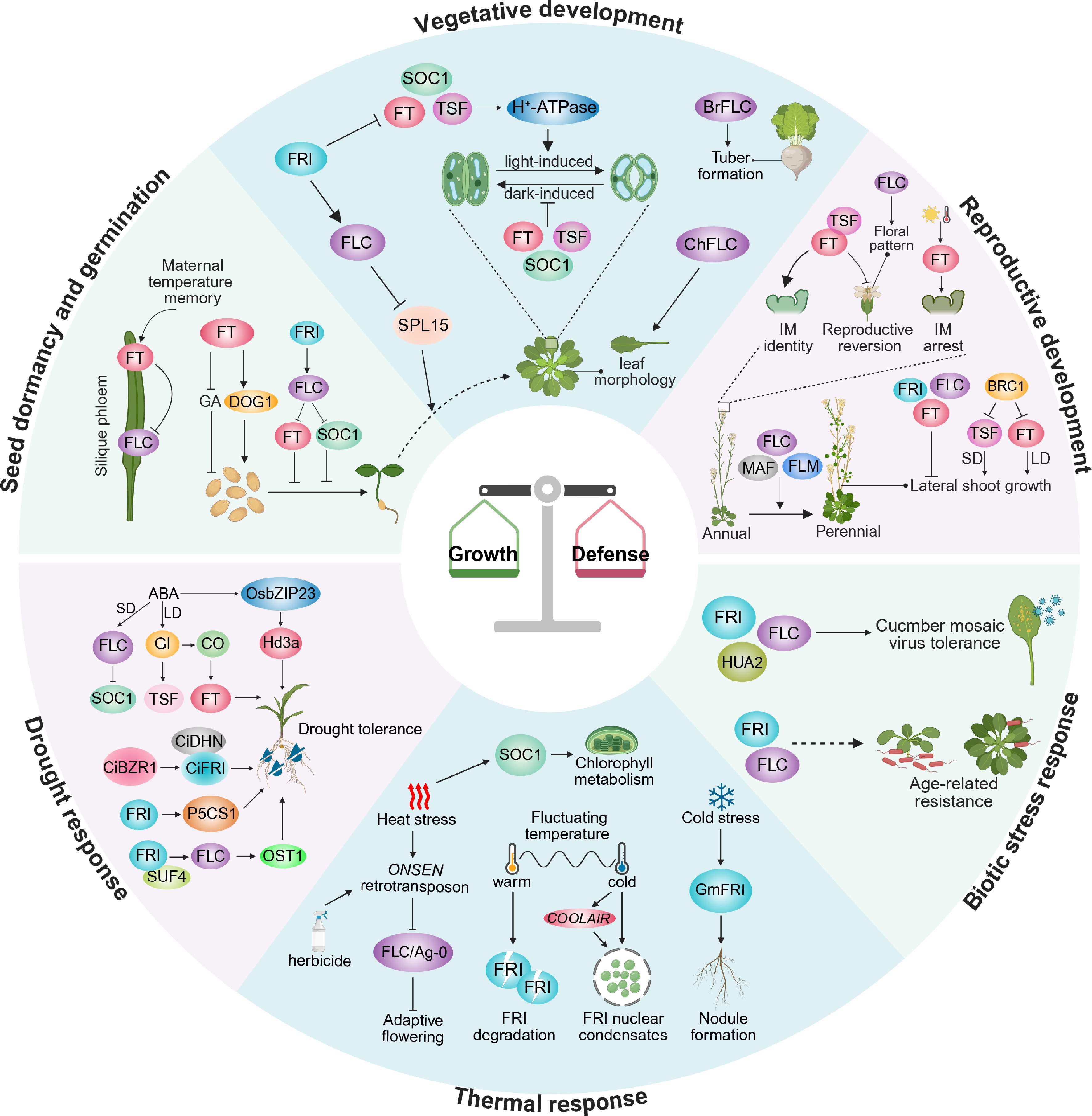
Figure 2.
Multifaceted roles of FRI, FLC, FT, and SOC1 in plant development and defense. The upper half focuses on their roles in growth and developmental transitions, and the lower half summarizes their regulation in response to abiotic environmental stresses and biotic challenges. Seed dormancy and germination: FT positively regulates seed dormancy by suppressing the GA pathway and activating the DELAY OF GERMINATION 1 (DOG1) pathway. Within the silique, FT mediates the control of offspring dormancy in response to maternal temperature signals through feedback inhibition of FLC. Conversely, FRI promotes the germination of non-vernalized maternal seeds by upregulating FLC. Vegetative development: FLC retards the transition from the juvenile to the adult phase by repressing SPL15 expression. FT, TSF, and SOC1 positively regulate stomatal opening. Furthermore, BrFLC correlates with tuber formation in turnips, whereas ChFLC regulates leaf size and complexity in Cardamine hirsuta. Reproductive development: FRI, FLC, and FT participate in regulating lateral shoot growth. BRC1 inhibits lateral shoot growth by repressing TSF or FT expression in a photoperiod-dependent manner. FT and TSF function redundantly to determine inflorescence meristem identity. Additionally, FT stabilizes the inflorescence and inhibits reproductive reversion, while also mediating the photo-thermal timing of inflorescence meristem (IM) arrest at the end of flowering. FLC, MAF and FLM are key factors maintaining perennial growth habit in perennial species. Drought response: FRI, FLC, FT enhance drought tolerance. In rice, the FT homolog Hd3a regulates drought escape, partially via an abscisic acid (ABA)-dependent pathway. Thermal response: SOC1 overexpression contributes to chlorophyll metabolism under heat stress. Heat shock activates the ONSEN retrotransposon located within the FLC intron 1, impairing FLC activity, and thereby promoting rapid adaptive flowering. Under fluctuating temperature conditions, warm destabilizes FRI, whereas cold promotes FRI nuclear condensation. In soybean, GmFRI positively regulates nodulation under cold stress. Biotic stress response: FRI, FLC and the flowering-time regulator HUA2 enhance plant tolerance to viral infection. Additionally, FRI and FLC promote age-related resistance (ARR) independently of the floral transition. Arrows denote positive effects; blunt lines denote repressive roles. Images were created by BioRender,
https://BioRender.com/cgkf6sf . -
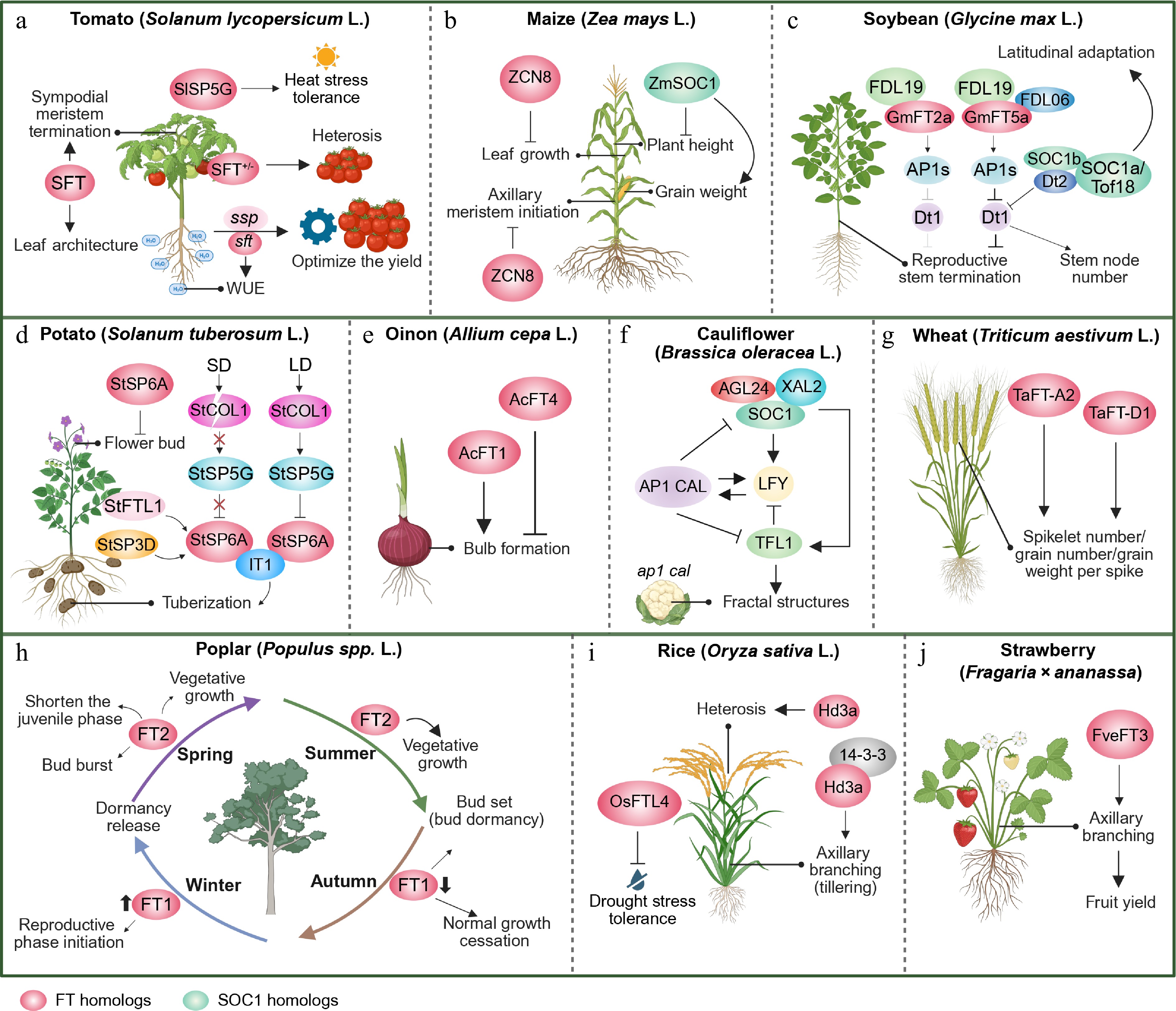
Figure 3.
Non-floral roles of FT and SOC1 family genes in major crops. (a) The tomato FT homologue SINGLE-FLOWER TRUSS (SFT) modulates sympodial meristem termination, leaf architecture, and water-use efficiency (WUE). SELF PRUNING 5G (SlSP5G) homologous to the TERMINAL FLOWER 1 (TFL1) promotes heat-stress tolerance; The combination of heterozygous sft and suppressor of sp (ssp) optimizes the fruit yield. (b) The maize FT homologue ZCN8 inhibits leaf growth and axillary meristem initiation, whereas the SOC1 homologue ZmSOC1 reduces plant height and enhances grain weight. (c) The soybean FT homologue GmFT5a rather than GmFT2a promotes stem termination through a specific interaction with FDL06 to inhibit Dt1-mediated reproductive stem growth. Tof18/SOC1a enhances latitudinal adaptation, and the SOC1s-Dt2 complex regulates stem node number by inhibiting Dt1. (d) Under long days (LD), CONSTANS-LIKE1 (StCOL1) activates the repressor StSP5G, which inhibits FT homologue StSP6A expression in potato; Under short days (SD), StSP6A triggers tuberization when the repressor is destabilized. Additionally, the florigen SELF PRUNING 3D (StSP3D) and FLOWERING LOCUS T-like 1 (StFTL1) are the novel tuberization signals. StSP6A also inhibits flowering buds. (e) Two onion FT homologs coordinate bulb formation with opposing functions: AcFT4 suppresses the formation of bulbs, while AcFT1 promotes it. (f) In cauliflower, the SOC1-centered floral gene network drives curd morphogenesis: loss of AP1 and CAL disrupts LFY expression maintenance and derepresses SOC1/AGL24, which induces ectopic TFL1 expression in floral primordia. TFL1 then further suppresses LFY, causing the primordia to lose floral identity, revert to inflorescence meristems, and ultimately form the characteristic cauliflower curds. (g) Two wheat FT homologs, TaFT-A2 and TaFT-D1, enhance spikelet number, grain number, and grain weight per spike, directly contributing to yield. (h) Two FT homologs act seasonally to coordinate poplar’s perennial growth cycle: FT2 promotes spring/summer vegetative growth, shortens the juvenile phase, and drives bud burst and bud set, whereas FT1 mediates autumn growth cessation, winter dormancy release, and the transition to reproduction. (i) The rice FT homolog Hd3a drives heterosis and regulates tillering (axillary branching) via 14-3-3 interaction, while OsFTL4 confers drought stress tolerance. (j) In strawberry, the FT homolog FveFT3 positively controls axillary branching, thereby increasing fruit yield. FT homologues are shown as pink circles and SOC1 homologues as light-green circles. Solid arrows indicate promotion/activation, while flat-ended (truncated) arrows denote repression/inhibition. Images were created by BioRender,
https://BioRender.com/xr70vik .
Figures
(3)
Tables
(0)