










-
Due to its pleasant taste and health benefits, tea is one of the most popular drinks in the world. Numerous studies have demonstrated that specialized metabolites in tea plants (Camellia sinensis), including volatile compounds, amino acids, polyphenols, and caffeine, play critical roles in the determination of tea quality and function. For example, volatile compounds contribute to tea flavor[1], and amino acids and polyphenols affect flavor and are beneficial for human health[2,3]. As these specialized metabolites are important quality attributes of tea, their biosynthesis is a topic that attracts much attention. Accurate information about metabolite biosynthesis is fundamental for other lines of research on tea, such as molecular breeding.
Specialized metabolite biosynthesis is a hotspot in tea research. Some enzymes involved in the final biosynthetic steps of many specialized metabolites in tea have been elucidated, such as volatile synthases[4−7], glucosyltransferases[8−11], caffeine synthase[12], polyphenol oxidase[13], and ʟ-theanine synthase[14]. However, a common weakness of previous studies is the lack of gene function identification in vivo in tea plants. Most of these enzyme activities have been identified and analyzed in vitro. The activities of some enzymes, such as linalool synthase[4,15], farnesene synthase[6], glutathione transferase[16]and ʟ-theanine synthase[17] have been analyzed in vivo in other plants such as Arabidopsis and Nicotiana. One reason for the lack of homologous in vivo gene function identification is that it has been difficult to establish a mature genetic transformation system for tea plants. For decades, many efforts have been made to establish an Agrobacterium-mediated genetic transformation system in tea plants, but this has been largely unsuccessful due to large amounts of bactericidal polyphenols[18]. Although β-glucuronidase, a reporter protein, has been expressed successfully in somatic embryos derived from cotyledon explants, leaf explants, and hairy roots in Agrobacterium-mediated transformation systems[19−21], none of these methods has successfully generated transgenic plants for gene functional analyses or molecular breeding.
Compared with traditional genetic transformation systems, transient overexpression/suppression provides a convenient alternative method for the analysis of gene function in vivo. Synthetic antisense oligodeoxynucleotides (AsODNs) and protoplasts are widely used for transient expression analysis in planta. Previous studies have used AsODNs to repress gene expression[22,23]. Transient suppression mediated by AsODNs has been used successfully to analyze enzyme activity in tea plants. The mRNA levels of target genes encoding enzymes such as glucosyltransferase (UGT91Q2 and UGT74AF3)[8,24], α/β-farnesene/β-ocimene[25], and linalool/nerolidol synthase (CsLIS/NES)[15] were significantly reduced, and the contents of relevant metabolites were decreased. The protoplast-based expression system is not only useful for enzyme activity assay, but also for subcellular localization analysis. As several proteins can be co-expressed in protoplasts, such a system can also be used to study protein-protein interactions and transcriptional activation. However, a protoplast transient expression system has not been developed for tea plants as large-scale isolation of protoplasts has been unsuccessful. Previous studies have tried to isolate protoplasts from tea leaves, flowers, and radicles[26−28]. Peng et al. successfully obtained mesophyll protoplasts from young leaves of tea seedlings grown in the dark, but found that young leaves of tea plants grown in tea plantations were not suitable for protoplast isolation[28]. These strict conditions limited the feasibility of tea protoplast isolation. Moreover, no previous studies have reported the application of tea protoplasts for protein transient expression.
Although protoplasts have been obtained from tea flowers and roots[27,28], the metabolic pathway occurring in tea leaves received more attention. Therefore, it would be useful to establish a transient expression method using mesophyll protoplasts for analyses of specialized metabolite biosynthesis. Here, we present a simple and highly efficient method for the isolation of tea mesophyll protoplasts. We also report on the subcellular localization analysis of enzymes involved in specialized metabolite biosynthesis. These methods are widely applicable for further studies on gene function identification in vivo.
-
To establish an efficient mesophyll protoplast isolation procedure, we attempted to isolate protoplasts from seven different tea cultivars: ‘Yinghong 9’ (YH9), ‘Huangyu’ (HY), ‘Baihaozao’ (BHZ), ‘Jinxuan’ (JX), ‘Longjin 43’ (LJ43), ‘Fuding Dabaicha’ (FDDBC), and ‘Zhongbai 4’ (ZB4). These included two varieties: C. sinensis var. sinensis (BHZ, JX, LJ43, FDDBC, ZB4) and C. sinensis var. assamica (YH9, HY). Five were normal green cultivars (YH9, BHZ, JX, LJ43, FDDBC) and two were natural albino mutants (HY, ZB4). All tea shoots were collected in April, 2019. Those of ‘ZB4’ were collected at Jiande, Zhejiang, China, and those of the other lines were collected at Yingde, Guangdong, China. Briefly, 0.4 g shoot tissue (one newly emerged bud and one leaf) was cut into strips and incubated in enzyme solution (1.5% cellulase R-10, 0.1% macerozyme R-10, 0.5% pectolase Y-23)[28] for 9 h in the dark with gentle shaking (Fig. 1). The enzymatic digestion solution in the Petri dish was checked under a microscope to determine the protoplast yield. Among these cultivars, ‘YH9’ and ‘HY’ yielded few mesophyll protoplasts, while ‘BHZ’, ‘LJ43’, ‘FDDBC’ and ‘JX’ yielded more. The highest yield of mesophyll protoplasts was obtained from ‘ZB4’ (Fig. 1c). Comparison of the yields of mesophyll protoplasts from the different tea lines confirmed that ‘ZB4’ was the most suitable line for mesophyll protoplast isolation among those tested.
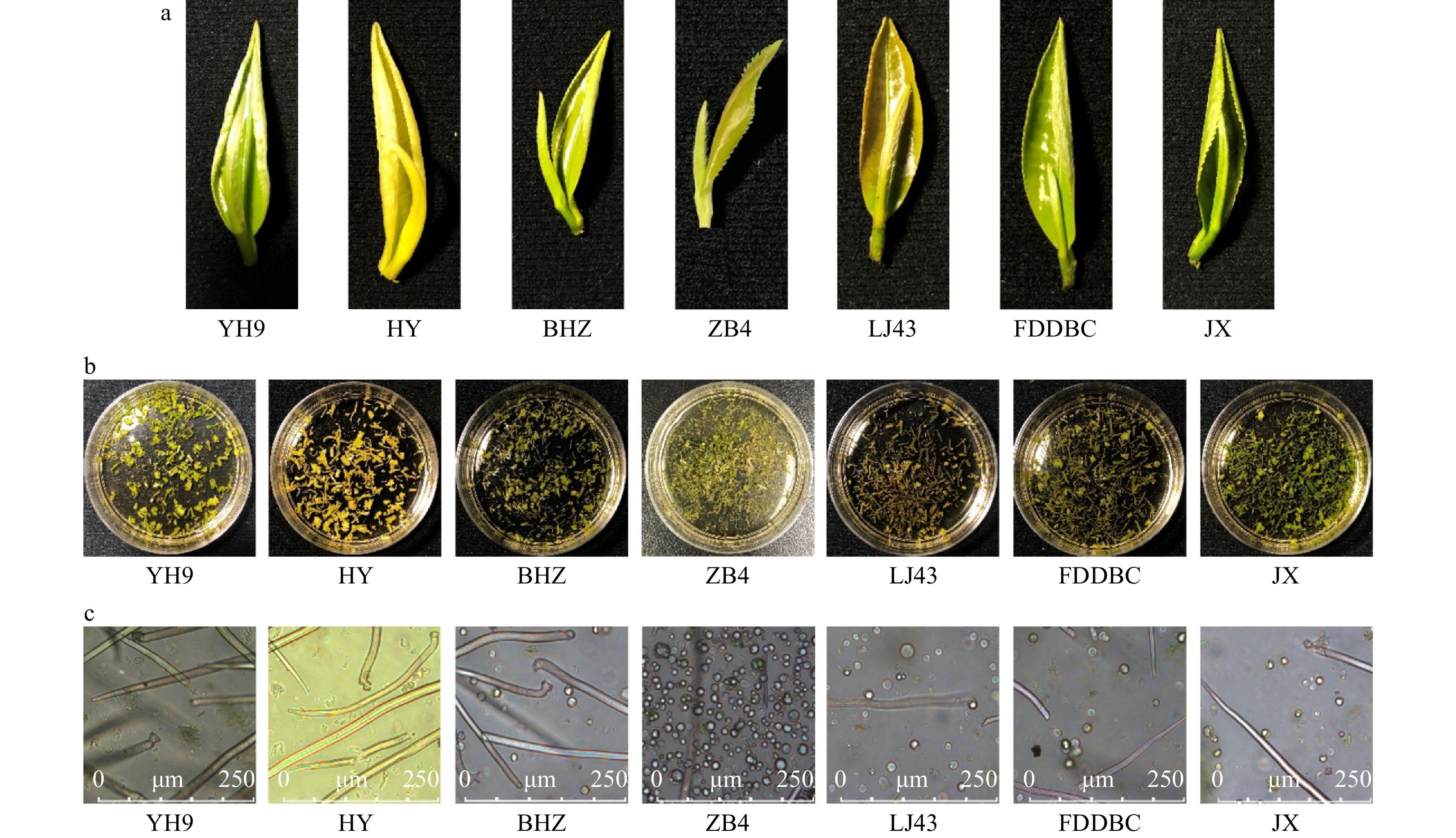
Figure 1. Protoplasts isolated from different tea plant cultivars. (a) Materials used for protoplast isolation. (b) Leaf strips in enzyme solution before enzyme digestion. (c) Isolated protoplast in enzyme solution. YH9, Yinghong 9; HY, Huangyu; BHZ, Baihaozao; ZB4, Zhongbai 4; LJ43, Longjin 43; FDDBC, Fuding Dabaicha; JX, Jinxuan.
The newly emerged leaves and mature leaves of ‘ZB4’ were thinner and softer than those of other lines. To determine why ‘ZB4’ was more suitable for mesophyll protoplast isolation, we analyzed the amounts of cellulose, pectin, and polyphenols in all the tested lines. Among all the lines, ‘ZB4’ had the lowest content of these cellular components (Fig. 2). The polyphenol content of ‘ZB4’ was about 8.90 mg/g F.W., significantly lower than in the other cultivars. To determine whether polyphenols were the critical factor affecting mesophyll protoplast yield, 0.4 mg/mL polyphenols was added to the enzyme solution without PVP. Few protoplasts were obtained after polyphenol addition, and the leaf strips in the enzyme solution containing polyphenols were barely digested. These results suggested that polyphenols affect the activity of cell wall degrading enzymes.
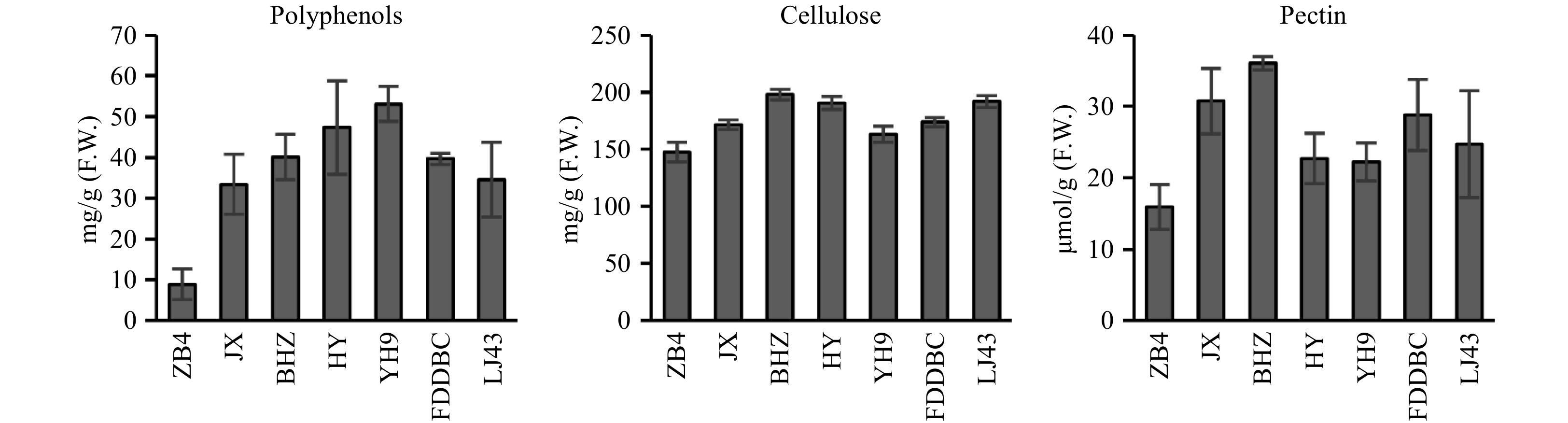
Figure 2. Polyphenols, cellulose and pectin content analysis of different tea plant cultivars. YH9, Yinghong 9; HY, Huangyu; BHZ, Baihaozao; ZB4, Zhongbai 4; LJ43, Longjin 43; FDDBC, Fuding Dabaicha; JX, Jinxuan.
Optimization of protoplast isolation conditions
-
We tested different enzyme combinations to improve the mesophyll protoplast yield from the first and second newly emerged leaves of ‘ZB4’ (Table 1). We found that the enzyme solution containing pectolase Y-23 became browner during the incubation period. This suggested that oxidation of polyphenols was occuring that had been released from the cells in this solution, possibly because of cell lysis. The original enzyme solution (1.5% cellulase R-10, 0.1% macerozyme R-10, 0.5% pectolase Y-23) yielded the lowest mesophyll protoplasts. The protoplast yield was significantly increased by raising the cellulase R-10 concentration to 3%, but was negatively affected by increasing the macerozyme R-10 concentration. Our results indicated that 3% cellulase R-10 and 0.3% macerozyme R-10 was the best combination of enzymes for isolating protoplasts from tea leaves. We also determined the effect of digestion time, from 3 to 18 h, on protoplast yield (Table 2). A few protoplasts were present in the enzyme solution after digestion for 3 h. The protoplast yield was increased after 6 h and the highest yield was around 12 h of digestion. The protoplasts lysed gradually after 12 h of digestion, which resulted in increased amounts of cell debris in the enzyme solution. Based on the above results, the best conditions for isolation of mesophyll protoplasts from ‘ZB4’ were 3% cellulase R-10 and 0.3% macerozyme R-10 with an incubation time of about 12 h.
Table 1. Protoplast yield in different enzyme solutions.
Cellulase
R-10Macerozyme
R-10Pectolase
Y-23Protoplast
(No./g F.W.)1.50% 0.10% 0.50% (4.64 ± 1.32) × 106 3.00% 0.30% (3.49 ± 0.46) × 107 1.50% 0.30% (1.18 ± 0.02) × 107 1.50% 0.60% (7.32 ± 1.49) × 106 3.00% 0.60% (3.02 ± 0.40) × 107 Table 2. Protoplast yield over different digestion times.
Isolation time Protoplast (No./g F.W.) 6 h (1.73 ± 0.32) × 107 9 h (2.27 ± 0.09) × 107 12 h (2.75 ± 0.49) × 107 15 h (2.05 ± 0.08) × 107 18 h (1.85 ± 0.14) × 107 Protoplasts, which were isolated from the first and second newly emerged leaves of ‘ZB4’ (Fig. 3a), ranged from 10 to 20 μm in size (Fig. 3e). The mesophyll protoplasts were round in W5 solution (Fig. 3e), suggesting that mannitol concentration was suitable for tea mesophyll protoplasts. We isolated mesophyll protoplasts from newly emerged tea leaves collected each month from early April to late October. High protoplast yields were obtained from all of these materials. Few new leaves emerged from November to March in the tea plantations. The yields of mesophyll protoplasts from dormant buds were very low.

Figure 3. Isolate protoplasts from ‘ZB4’. (a) Leaves used for protoplasts isolation. (b) Leaf strips before enzyme digestion. (c) Leaf strips after enzyme digestion (3% cellulase R-10 and 0.3% macerozyme R-10) for 12 h. (d) Purified protoplasts solution. (e) Purified protoplasts. Bar: 50 μm.
Transient expression and protein subcellular localization analyses in tea mesophyll protoplasts
-
The protoplasts were transfected with a 35S::YFP construct (pSAT6-EYFP-N1 4.6 kb) using a polyethylene glycol-mediated transfection approach[29]. About 35%–50% of the protoplasts were YFP-positive (Fig. 4), which indicates that a heterologous protein could be transiently expressed in these tea plant mesophyll protoplasts. Next, proteins related to the biosynthesis of specific tea metabolites in several different subcellular compartments were selected for transient expression in tea mesophyll protoplasts (Fig. 5). These proteins were (S)-linalool synthase CsSLIS[4], (R)-linalool synthase CsRLIS[4], (E)-nerolidol synthase CsNES[5], cytochrome c oxidase subunit 5b CsCOX5b, ATP synthase subunit d CsATPaseD, a basic helix-loop-helix transcript factor involved in jasmonate signaling (CsMYC2)[7], jasmonate-zim-domain protein 2 CsJAZ2[7], and polyphenol oxidase CsPPO3[13]. The predicted locations of these proteins were in different organelles and cellular compartments. In the subcellular localization analyses, CsSLIS and CsRLIS were located in the chloroplast, and CsNES was located in the cytosol and nucleus. These results were consistent with those of previous reports on their subcellular localization in Arabidopsis mesophyll protoplasts[4,5]. Cytochrome c oxidase subunit 5b and ATP synthase subunit d are well-known mitochondria-localized proteins[30]. In our analyses, the YFP fluorescence of CsCOX5b and CsATPaseD overlapped with the signal of MitoTracker Deep Red, confirming the mitochondrial localization of CsCOX5b and CsATPaseD in tea plant mesophyll protoplasts. CsMYC2 and CsJAZ2 are key regulators of jasmonate signaling[7]. As functional transcription factors, CsMYC2 and CsJAZ2 should be located in the nucleus. In our analyses, the YFP fluorescence of CsMYC2 and CsJAZ2 overlapped with the signal of the mCherry nuclear marker, confirming their nuclear localization. The above results showed that the transiently expressed proteins were correctly localized in the tea plant mesophyll protoplasts. We also used the isolated tea plant mesophyll protoplasts to analyze the subcellular localization of a polyphenol oxidase (CsPPO3) that plays an important role in polyphenol metabolism in tea plants. Although polyphenols are localized in the vacuole, the subcellular localization of CsPPO3 was previously unknown. In our analyses, the YFP-fused CsPPO3 was located in the chloroplasts of tea plant mesophyll protoplasts.

Figure 4. Transfection of tea plant mesophyll protoplast with 35S::YFP construct (pSAT6-EYFP-N1 4.6 kb). (a, c, e) Tea plant mesophyll protoplasts in bright field. (b, d, f) YFP expressed in tea plant mesophyll protoplasts.
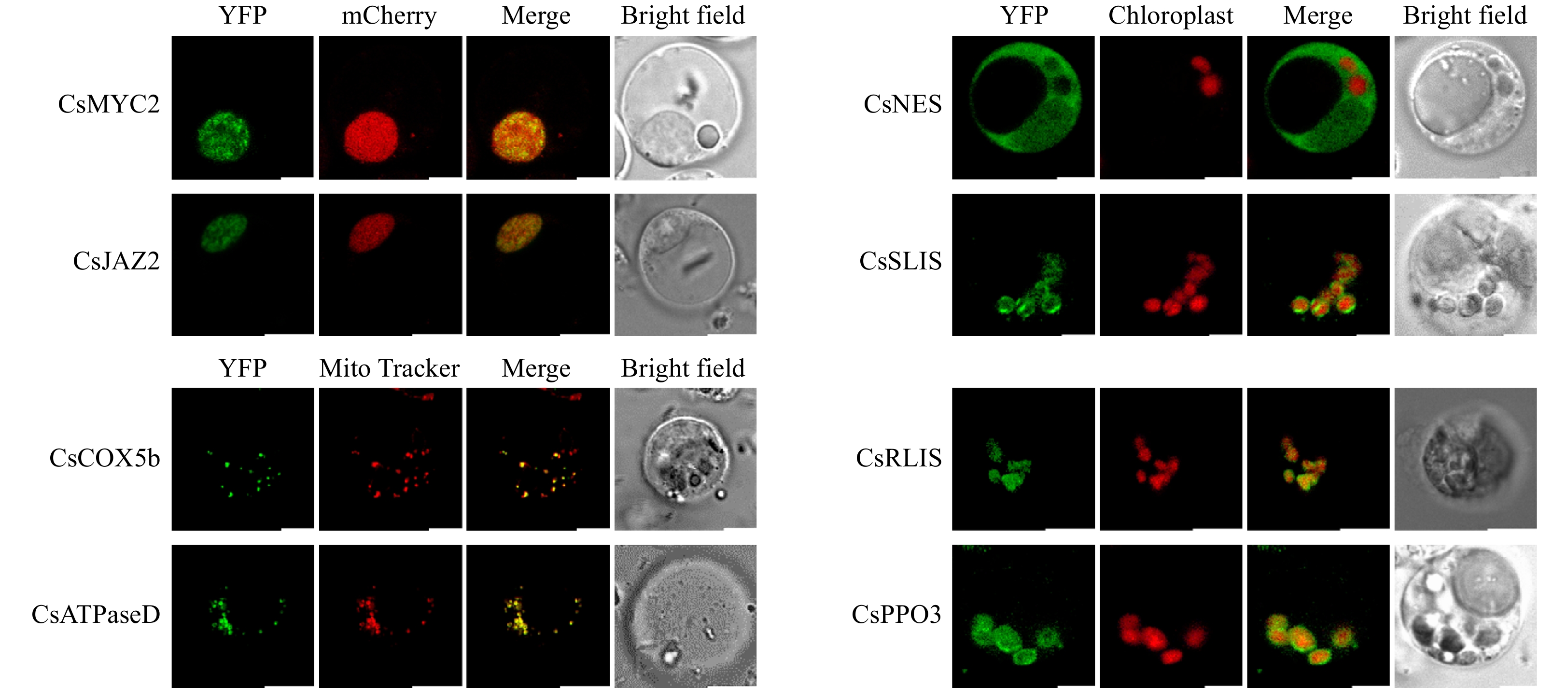
Figure 5. Subcellular localization analysis of different tea plant proteins. Bar: 5 μm.
We also attempted to assay CsNES activity using the tea plant mesophyll protoplasts. However, the results were inconsistent (Supplemental Fig. S1). After supplying farnesyl diphosphate (FPP), the amount of nerolidol formed in CsNES-overexpressing protoplasts was not always higher than that formed in protoplasts harboring the empty vector. These inconsistent results may have been due to the low transfection efficiency.
Elucidating specialized metabolite biosynthesis by isotopic feeding using tea mesophyll protoplasts
-
Stable isotopic tracing is an effective method to discover the metabolite biosynthesis pathway in plants. To evaluate the possibility of using tea mesophyll protoplasts to perform stable isotopic tracing, tea mesophyll protoplasts were used to feed with [2H5]-acetophenone. After feeding for 4 h, internal volatiles of protoplast was extracted and analyzed by GC-MS. It was shown that [2H5]-(S)-1-phenylethanol and [2H5]-(R)-1-phenylethanol, the representative aroma compounds in tea flowers, were detected (Fig. 6), suggesting that acetophenone was the precursor of 1-phenylethanol. This result was consistent with our previous report using tea flowers for isotopic feeding[31]. Our study suggested that tea mesophyll protoplast was also suitable for stable isotopic tracing.
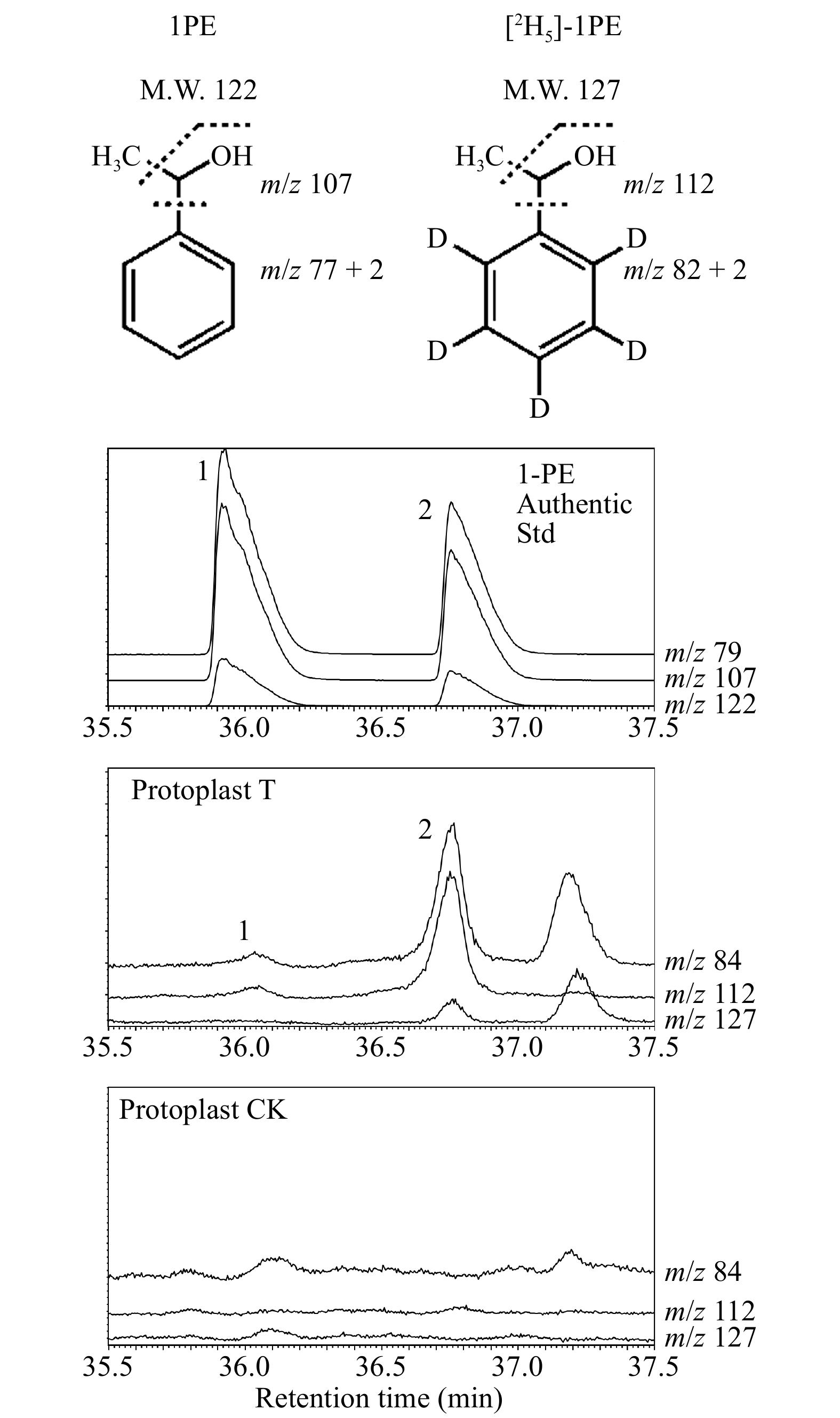
Figure 6. Stable isotopic tracing using tea mesophyll protoplasts.
1. (R)-1-phenylethanol; 2. (S)-1-phenylethanol. Protoplast T: protoplasts treated with 20 μL of 2.5 mM [2H5]-acetophenone (dissolved in 5% ethanol). Protoplast CK: protoplasts treated with 20 μL of 5% ethanol. -
Obtaining sufficient high-quality protoplasts is the first step in establishing an efficient transient expression system. Although protoplast isolation is now routine for a wide range of species, this technology has developed more slowly for woody plants than for herbaceous plants. Previous studies have reported the successful isolation of protoplasts from only a few woody plants, including Platanus orientalis[32], Broussonetia kazinoki[33], Populus[34], Morus alba[35], and Albizia julbrissin[36].
In most previous studies, woody plant protoplasts have been obtained from callus[36−38] or sterile seedlings[32,34,36,39,40]. Although tea leaves are covered by a thick layer of epicuticular wax, we established a highly efficient mesophyll protoplast isolation procedure by using newly emerged leaves from trees growing in tea plantations. Thus, there is a reliable source of materials for our experiments. It was notable that only newly emerged tea leaves were suitable for mesophyll protoplast isolation. We had found that when newly emerged leaves could not be found in a tea plantation (from November to March), mesophyll protoplast could not be isolated from dormant buds. Therefore, tea picking season would restrict the application of our mesophyll isolation method. Alternatively, we can use tea plant grown in the green house in winter. Previous studies have reported low yields of mesophyll protoplasts from many woody plants, for example, Magnolia (1.89 × 105/g F.W.)[41], grapevine (3.4 × 105/g F.W.)[42], and Albizia julibrissin (6.31 × 105/g F.W.)[36]. Compared with other woody plants, the tea line ‘ZB4’ showed a higher yield of mesophyll protoplasts (3.49 × 107/g F.W.), which guaranteed a sufficient amount for transfection studies.
Based on our results, the source material is an important factor for successful isolation of mesophyll protoplast isolation. Our results show that protoplast yields varied widely among different tea lines. The C. sinensis var. assamica cultivars ‘YH9’ and ‘HY’ were highly resistant to enzyme digestion. Compared with ‘ZB4’, the other C. sinensis var. sinensis cultivars yielded far fewer mesophyll protoplasts. Our analysis suggest that polyphenols might inhibit the enzymatic digestion of leaf tissues, which resulted in low yields of protoplasts. Therefore, the low polyphenol content could be one reason for the high mesophyll protoplast yield from ‘ZB4’. Relatively lower content of cellulose and pectin may also have contributed to the high protoplast yield from ‘ZB4’.
The enzyme treatment conditions are also important for successful protoplast isolation. Different plant species require different combinations of enzymes to degrade cell walls effectively. Our results showed that 3% cellulase was required for tea mesophyll protoplast isolation. This concentration is higher than those used to isolate protoplasts from woody plants in other studies, which suggests that there were high cellulose content in the cell walls of tea materials. Although a prolonged incubation time can increase tissue digestion, tea plant mesophyll protoplasts were prone to lysis in enzyme solution after 12 h of digestion. In general, 6 h digestion was enough to obtain sufficient protoplasts for further analyses.
Establishment of mesophyll protoplast-based homologous transient expression system and stable isotopic tracing system
-
Many methods have been used to overexpress genes to study their function in plants. These methods include biolistic bombardment, infiltration with Agrobacterium tumefaciens, and polyethylene glycol-mediated protoplast transformation. In tea plants, gene function has mainly been studied by expression in prokaryotic systems. Some heterologous in vivo systems, such as those based on tobacco and Arabidopsis, have also been used, although the heterologous protein may exhibit aberrant traits. For example, some Arabidopsis proteins expressed in tobacco are mis-localized[43]. Moreover, some of the specialized metabolite biosynthesis pathways that exist in tea are absent from model plants such as Arabidopsis and tobacco. Therefore, they are not suitable for studies on genes involved in these metabolic pathways. For these reasons, a homologous in vivo system is essential for studying the function of genes from tea plants. Given that rapid and reliable genetic transformation systems have not yet been established for tea plants, a protoplast-based transient expression system is a promising alternative.
A transient expression system for tea plant mesophyll protoplasts was established in our study. Using this system, we determined the subcellular localization of enzymes involved in the synthesis of specialized metabolites. The location of a protein in an organelle plays an important role in determining its function and which metabolic pathways it is involved in. The substrate of nerolidol synthase, FPP, is located in the cytosol, so nerolidol synthase should also be located in the cytosol for access to its substrate. Similarly, the substrate of linalool synthase, geranyl diphosphate (GPP), is located in the chloroplast, so linalool synthase should also be chloroplast-localized. In some cases, the enzyme and substrate show different sub-cellular compartmentalization. Polyphenols, the substrates of PPO, are mainly localized in the vacuole[44,45]. Galloylated catechins, the major polyphenols in tea plants, are also primarily located in the vacuole[46]. Our results show that CsPPO3 is located in the chloroplasts in tea mesophyll protoplasts. This location is consistent with the predicted plastidic localization of PPOs in plants[47]. This pattern of sub-cellular compartmentalization explains why only the wound edges in tea plants turn brown; i.e., because CsPPO3 and its vacuolar phenolic substrates will only come into contact when cells are disrupted. A nucleus-marker protein and CsMYC2/CsJAZ2 were co-expressed in the protoplasts, which confirms that the transfection efficiency was sufficient for co-localization analysis. All the proteins we analyzed were localized in the predicted organelle, which indicates that this transient expression system is mature and reliable enough for enzyme subcellular localization analysis. The biosynthesis of specialized metabolites in tea plants depends on the subcellular location of synthase and their substrates. The subcellular biosynthetic sites for specialized metabolites can also be studied using a non-aqueous fractionation approach[48−50]. The combination of tea mesophyll protoplast-based enzyme subcellular localization analyses and non-aqueous fractionation methods will shed new light on the biosynthesis of specialized metabolites within tea plant cells.
Besides the transient expression system, we also developed a tea mesophyll protoplast- based stable isotopic tracing system. Compared with plant tissues, chemical compounds could enter into cells more easily when mesophyll protoplasts were used. Thus, tracing time could be shortened and it would be beneficial to some self-degradable compounds. On other hand, using protoplasts for isotopic tracing required lower quantities of isotopically labeled compound, which would be a great advantage for some valuable isotopically labeled precursors.
Optimization of transfection procedure for tea mesophyll protoplasts to broaden applications
-
We attempted to assay CsNES activity by overexpressing its encoding gene in tea mesophyll protoplasts. However, the results were inconsistent, possibly because of the low transfection efficiency. The transfection efficiency using the 35S::YFP construct in our study was about 35%–50%, which is lower than those reported for other transient expression systems. For example, 50%–70% transfection efficiency was achieved in a rice protoplast system[51], and 70% transfection efficiency was achieved in Populus mesophyll protoplasts[34]. It has been reported that a transfection efficiency higher than 50% is necessary for reliable analysis[29]. The weakness of our tea mesophyll protoplast transient expression system is its low transfection efficiency. Because the transfection efficiency is affected by the plasmid concentration and quality, transfection time, transfection reagents, and ratio of plasmid to protoplasts, there are many factors that can be adjusted to improve this system. A high transfection efficiency will guarantee that the tea mesophyll protoplast system can be used for analyses of enzyme activity, protein-protein interactions, transcriptional activation, and protein overexpression. A protoplast system is also useful for RNA interference (RNAi) analyses to reduce the expression of certain target genes by transfection with double-stranded RNA[52]. Improving the transfection efficiency of the tea plant mesophyll protoplast system will make it suitable for such RNAi analyses.
-
For in-depth analyses of specialized metabolite biosynthesis pathways in tea, it is essential to establish a homologous in vivo gene function identification system. We have developed an efficient and simple method to isolate mesophyll protoplasts from tea leaves. Our results show that the isolated protoplasts can used for transient protein expression, and for subcellular localization analyses of various proteins. In future research, we intend to improve the transfection efficiency for broader applications of this system, for example, for analyses of enzyme activity and transcriptional activation. Our research represents significant progress in establishing an in vivo gene function identification system for tea. With continuing improvements to this transient expression system, it will become feasible to analyze gene function in vivo in tea plants.
-
‘Zhongbai 4’ (ZB4) tea plant cultivar was obtained in Jiande city, Zhejiang province, China. Other cultivars including ‘Yinghong 9’ (YH9), ‘Huangyu’ (HY), ‘Jinxuan’ (JX), ‘Longjin 43’ (LJ43), ‘Fuding Dabaicha’ (FDDBC) and ‘Baihaozao’ (BHZ) were obtained in Yingde city, Guangdong province, China in April, 2019. Cellulase ‘ONOZUKA’ R-10, macerozyme R-10, and pectolyase Y-23 were purchased from Yakult Pharmaceutical Ind. Co., Ltd., Japan. PEG4000 (cat. no. 81240), CaCl2 (cat. no. 223506), D-(+)-glucose (cat. no. G5767), NaCl (cat. no. S9888), KCl (cat. no. P3911), MgCl2 (cat. no. M9272), 4-morpholineethanesulfonic acid (MES, cat. no. M8250) and polyvinylpyrrolidone K30 (PVP, cat. no. 81420) were purchased from Sigma-Aldrich.
We suggested using the reagents we mentioned above to isolate tea mesophyll protoplasts. We did find that when another MES was used in the enzyme solution, the tea plant mesophyll protoplast was lysed during digestion. Also, mesophyll protoplast was lysed in MMG solution when another MgCl2 was used in MMG solution.
Tea protoplast isolation
-
To screen the best tea cultivar for mesophyll protoplasts isolation, one bud and newly emerged first leaves of different tea cultivars were used. They were cut into strips directly. After identifying that ‘ZB4’ was used for follow-up experiments, the tender tea leaves (newly emerged first and second leaves) of ‘ZB4’ were used for protoplast isolation. The tender tea leaves collected from early April to late October could be used for protoplast isolation. After removing the primary vein, the remaining leaves were cut into fine strips (0.5 mm or less). Leaf strips were transferred promptly and gently into the prepared enzyme solution. Enzyme solution contained various combinations of cell wall degrading enzymes, 20 mM MES pH 5.7, 0.4 M mannitol, 20 mM KCl and 4% PVP. Enzyme solution was incubated at 50 °C, 10 min for better dissolution. Leaf strips were digested at 25 °C with shaking of 40 rpm in the dark for the indicated time period. Generally, 0.4−0.5 g leaves were digested with 10 mL enzyme solution. After that, equal volume of W5 solution (2 mM MES pH 5.7, 154 mM NaCl, 125 mM CaCl2, and 5 mM KCl) was added into the enzyme solution and the mixture was filtered with nylon mesh (200 mesh/inch). The flow-through was centrifuged at 200 g for 2 min to pellet protoplasts. To evaluate protoplast yield under different isolation conditions, the protoplast was re-suspended with 5 mL W5 solution and counted with a hemocytometer.
Transfection of protoplasts, subcellular localization analysis and enzyme activity assay
-
The subcellular localizations of CsPPO3 (polyphenol oxidase GenBank Accession: MK977644), CsNES ((E)-nerolidol synthase, GenBank Accession: KY033151), CsSLIS ((S)-linalool synthase, GenBank Accession: AGX26045), CsRLIS ((R)-linalool synthase, GenBank Accession: MT178265), CsJAZ2 (jasmonate-zim-domain protein 2, GenBank Accession: MK336377), CsMYC2 (basic helix-loop-helix transcript factor involved in jasmonate signaling GenBank Accession: MK336383), CsCOX5b (cytochrome c oxidase subunit 5b GenBank Accession: XM_028237613) and CsATPaseD (ATP synthase subunit d, GenBank Accession: XM_028211204) were analyzed. Open reading frame of the above genes was constructed into pSAT6-EYFP-N1 vector. Protoplasts were transfected using procedures reported in Arabidopsis, with modification[29]. After filtering and centrifugation, the pelleted protoplasts were washed with W5 solution twice and re-suspended in W5 solution at 2 × 105 mL−1. The re-suspended protoplasts were kept on ice for at least 1 h. Tea plant mesophyll protoplasts sunk more slowly than Arabidopsis mesophyll protoplasts. As much of the W5 solution was removed as possible and the protoplasts were re-suspended at about 2 × 105 mL−1 in MMG solution (4 mM MES pH 5.7, 0.4 M mannitol, 15 mM MgCl2). The following items were added sequentially into 2-mL round-bottom tubes: 100 μL protoplasts, 10 μL plasmid (10 μg), 110 μL PEG solution (40% PEG 4000, 0.2 M mannitol, 100 mM CaCl2). The mixture was gently mixed by tapping the tube and was incubated at room temperature for 10 min. 440 μL of W5 solution was added to terminate the transfection and the mixture was centrifuged at 200 g for 2 min. The pelleted protoplasts were re-suspended in 500 μL W5 solution and incubated at 25 °C for 12−16 h. To stain mitochondria, the protoplasts were incubated with 50 mM MitoTracker Deep Red FM (Yeasen, Shanghai, China) for 30 min at room temperature. The fluorescence was observed under confocal microscope (LEICA-SP8 STED 3X, Germany). To assay enzyme activity of CsNES, CsNES/empty vector-overexpressed protoplasts were incubated with 2 μg farnesyl diphosphate (FPP) at 25 °C for 4 h. Afterwards, solid-phase microextraction (SPME, 2 cm–50/30 μm DVB/Carboxen/PDMS Stable Flex) was used to collect volatile compounds at 42 °C for 30 min. Then SPME was subject to GC-MS (GCMS-QP2010 SE, Shimadzu, Kyoto, Japan) analysis using a SUPELCOWAX 10 column (30 m × 0.25 mm × 0.25 μm, GL Sciences, Tokyo, Japan). The GC-MS program was as follows: 60 °C for 3 min; increased to 240 °C at 10 °C/min; and maintained at 240 °C for 10 min. Full scan mode (m/z 40–200) was used.
Polyphenols, cellulose, and pectin content analysis
Tea polyphenols
-
0.2 g finely powdered tea leaves were extracted with 5 mL pre-warmed 70% methanol for 10 min at 70 °C. After centrifuge at 3,500 g for 10 min, the supernatant was saved and used for polyphenol analysis. Polyphenols were analyzed using a tea polyphenols analysis kit (Leagene Biotechnology, Beijing, China) based on Folin-phenol method. Briefly, 25 μL samples were mixed with 125 μL Folin-phenol reagent. The mixture was incubated at room temperature for 5 min and then 100 μL chromogenic agent was added. After incubation at room temperature for 60 min, tea polyphenol content was calculated based on absorbance value of OD765 and the standard curve of tea polyphenols.
Cellulose
-
Cellulose was assayed using a cellulose content assay kit (Solarbio, Beijing, China). First, cell wall material (CWM) was extracted from 0.3 g finely powdered tea leaves according to the manufacture’s instruction. 5 mg CWM was homogenized with 0.5 mL ddH2O, then 0.75 mL concentrated H2SO4 was added. After keeping on ice for 30 min, the mixture was centrifuged at 8,000 g at 4 °C for 4 min. The supernatant was used for cellulose analysis based on absorbance value of OD620 according to the manufacture’s instructions.
Pectin
-
Pectin was assayed using a total pectin assay kit (Solarbio, Beijing, China) according to the manufacture’s instructions. 50 mg finely powdered tea leaves were extracted with 1 mL extraction buffer 1 (provided with the assay kit) at 90 °C for 30 min twice. After centrifugation at 5,000 g at 25 °C for 10 min, the pellet was hydrolyzed with extraction buffer 2 (provided with the assay kit) at 90 °C for 60 min. The mixture was then centrifuged at 8,000 g at 25 °C for 15 min. The resultant supernatant was used for pectin assay based on absorbance value of OD530 and standard curve of pectin.
Isotopic tracing with tea mesophyll protoplast
-
20 μL of 2.5 mM D5-Acetophenone (dissolved in 5% ethanol) was added into 1 mL tea mesophyll protoplasts (2 × 107 mL−1) for 4 h at 25 °C. Protoplasts treated with 20 μL of 5% ethanol was used as control. Then 1 mL n-hexane/ethyl acetate (1:1, v/v) was added to the protoplast solution and vortexed vigorously for 30 s. The upper phase was collected and concentrated under N2 to about 200 μL. 1 μL of extract was subjected to GC-MS (GCMS-QP2010 SE, Shimadzu, Kyoto, Japan) analysis using a InertCapTM CHIRAMIX column (GL Sciences, 30 m × 0.25 mm × 0.25 μm). The GC-MS program was: 60 °C for 3 min, 2 °C/min to 180 °C, 180 °C for 20 min.
- This study was financially supported from the National Natural Science Foundation of China (31870684), the Pearl River Science and Technology New Star Fund of Guangzhou (201806010018), the Youth Innovation Promotion Association of Chinese Academy of Sciences (Y821131001), the Regional Key Project of Science and Technology Service Network Plan of Chinese Academy of Sciences (KFJ-STS-QYZX-093), the National Key Research and Development Program of China (2018YFD1000601), and the Guangdong Provincial Special Fund For Modern Agriculture Industry Technology Innovation Teams (2020KJ120).
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Enzyme activity assay of CsNES using tea plant mesophyll protoplast.The results of five independent experiments are shown. y-axis is the peak area of formed nerolidol. CK: mesophyll protoplast transfected with empty vector; CsNES: mesophyll protoplast transfected with CsNES. All data are means of three independent replicates and error bars represent ±SD. * indicates statistically significant differences (p≤0.05). Three independent protoplast transfections were performed. Each transfected protoplasts were used for enzyme activity assay and was considered as one replicate. The experiments were performed for five times at different time to identify the stability of results.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou Y, Deng R, Xu X, Yang Z. 2021. Isolation of mesophyll protoplasts from tea ( Camellia sinensis ) and localization analysis of enzymes involved in the biosynthesis of specialized metabolites. Beverage Plant Research 1: 2 doi: 10.48130/BPR-2021-0002

Isolation of mesophyll protoplasts from tea (Camellia sinensis) and localization analysis of enzymes involved in the biosynthesis of specialized metabolites
- Received: 20 March 2021
- Accepted: 20 April 2021
- Published online: 14 May 2021
Abstract: Specialized metabolites in tea (Camellia sinensis) are fundamental quality factors. It is important to characterize gene function in vivo to identify key enzymes and reactions involved in the biosynthesis of such metabolites. Here we report a transient expression method to analyze gene function in isolated tea mesophyll protoplasts. This method is an alternative approach to traditional genetic transformation for studies on gene function in vivo. We screened several tea cultivars and different digestion conditions to optimize protoplast isolation. Digestion of newly emerged leaves of C. sinensis ‘Zhongbai 4’ with 3% cellulase R-10 and 0.3% macerozyme R-10 for about 12 h yielded approximately 107 mesophyll protoplasts. Genes encoding enzymes involved in secondary metabolite synthesis were transiently expressed in the protoplasts, and their subcellular locations were determined. With further improvements in the transfection efficiency, this transient expression system will contribute greatly to the analyses of in vivo gene function in tea.
-
Key words:
- Camellia sinensis /
- tea /
- protoplast /
- transient expression /
- subcellular localization