









-
The expansion of the ozone hole, which has been greatly accelerated by anthropogenic activities, has resulted in a steady increase in the amount of ultraviolet radiation reaching the earth's surface[1−3]. Enhanced ultraviolet radiation (wavelength range 200–400 nm), especially UV-B radiation (280–315 nm), damages plant proteins, photosynthetic pigments and membranes and affects plant growth, development and metabolism[4−6]. Therefore, enhancing plant tolerance to UV-B stress is critically important for improving plant productivity.
Plants have evolved a variety of mechanisms to deal with UV-B radiation stress. In environments that experience high levels of ultraviolet radiation, plants are often shorter and their leaves thicker and smaller, as these changes increase survival in such environments[7−9]. In addition, plants possess a powerful reactive oxygen species (ROS) scavenging system, including various non-enzymatic antioxidants (e.g., proline, ascorbic acid and carotenoids) and antioxidant enzymes (e.g., ascorbic acid peroxidase (APX), superoxide dismutase and catalase (CAT)), that remove excess ROS[10,11].
Another mechanism for protecting plants against UV damage is through the synthesis of secondary metabolites that reduce the amount of UV-B radiation entering plants[12]. Phenols are particularly important secondary metabolites because of their ability to remove excess ROS and absorb UV radiation[13−15]. Phenolic compounds are mainly synthesized in the cytoplasm and accumulate in vacuoles; their synthesis can be enhanced under strong ultraviolet and visible light conditions[16,17]. Polyphenols in apple are produced through a series of enzymatic reactions starting from phenylalanine, including PAL (EC: 4.3.1.24, phenylalanine ammonia-lyase), CHS (EC: 2.3.1.74, chalcone synthase), F3H (EC: 1.14.11.19, flavanone 3-hydroxylase), DFR (EC: 1.1.1.219, dihydroflavanol 4-reductase), ANS (EC: 1.14.20.4, anthocyanin synthase), UFGT (EC: 2.4.1.115, UDP-glucose: flavonoid 3-O-glucosyltransferase), FLS (EC: 1.14.11.23, flavonol synthase), ANR (EC: 1.3.1.112, anthocyanin reductase) and LAR (EC: 1.17.1.3, leucoanthocyanidin reductase)[18,19]. Previous studies have shown that MdLAR and MdANR genes are correlated with flavonoid accumulation in apples[20]. In addition, our previous study showed that the application of exogenous melatonin significantly increased the expression of MdCHS, MdF3H, MdANS, MdUFGT, MdLAR and MdFLS under UV-B stress which was consistent with the increase in phenolic compounds[21].
Melatonin is an indole that has been shown to improve plant stress resistance. For example, Li et al[22] demonstrated that 0.1 μM exogenous melatonin alleviated salt stress in apples by improving antioxidant activity and regulating ion balance. Moreover, melatonin can improve the function of stomata and the drought tolerance of apples[23]. The negative effects of replant disease[24] and alkaline stress[25] in apples can also be alleviated by exogenous melatonin. The pathway of melatonin synthesis in animals has been well studied[26]. With tryptophan as a precursor, melatonin is finally synthesized through the action of four enzymes: T5H (tryptophan 5-hydroxylase), TDC (tryptophan decarboxylase), AANAT (arylalkylamine-N-acetyltransferase) and HIOMT (hydroxyindole-O-methyltransferase)[26]. The pathways of melatonin synthesis in plants have also been described. The first two steps of the melatonin synthesis pathway in plants differ from those in animals, but the last two steps are similar[27]. AANAT and HIOMT in humans are thought to be key synthases that play a rate-limiting role in the last two steps of the melatonin synthesis pathway[28,29].
Previously, we found that the application of exogenous melatonin could improve the resistance of apples to UV-B stress by increasing the activity of antioxidant enzymes, and the content of phenolic compounds in leaves[21]. Some studies have shown that transgenic plants with AANAT and HIOMT genes from animals have increased melatonin levels and significantly improved stress resistance[30]. Here, the synthetic human melatonin synthase genes (AANAT and HIOMT) were transferred respectively into 'GL-3' apples, and the tolerance of apple plants with ectopic expression of AANAT or HIOMT under UV-B stress was analyzed to better understand the role of melatonin in UV-B stress tolerance. We found that the melatonin content and the tolerance to UV-B stress were noticeably increased by the ectopic expression of AANAT or HIOMT. Overall, the results of this study increased our understanding of the function of melatonin under UV-B stress from the perspective of melatonin endogenous metabolism.
-
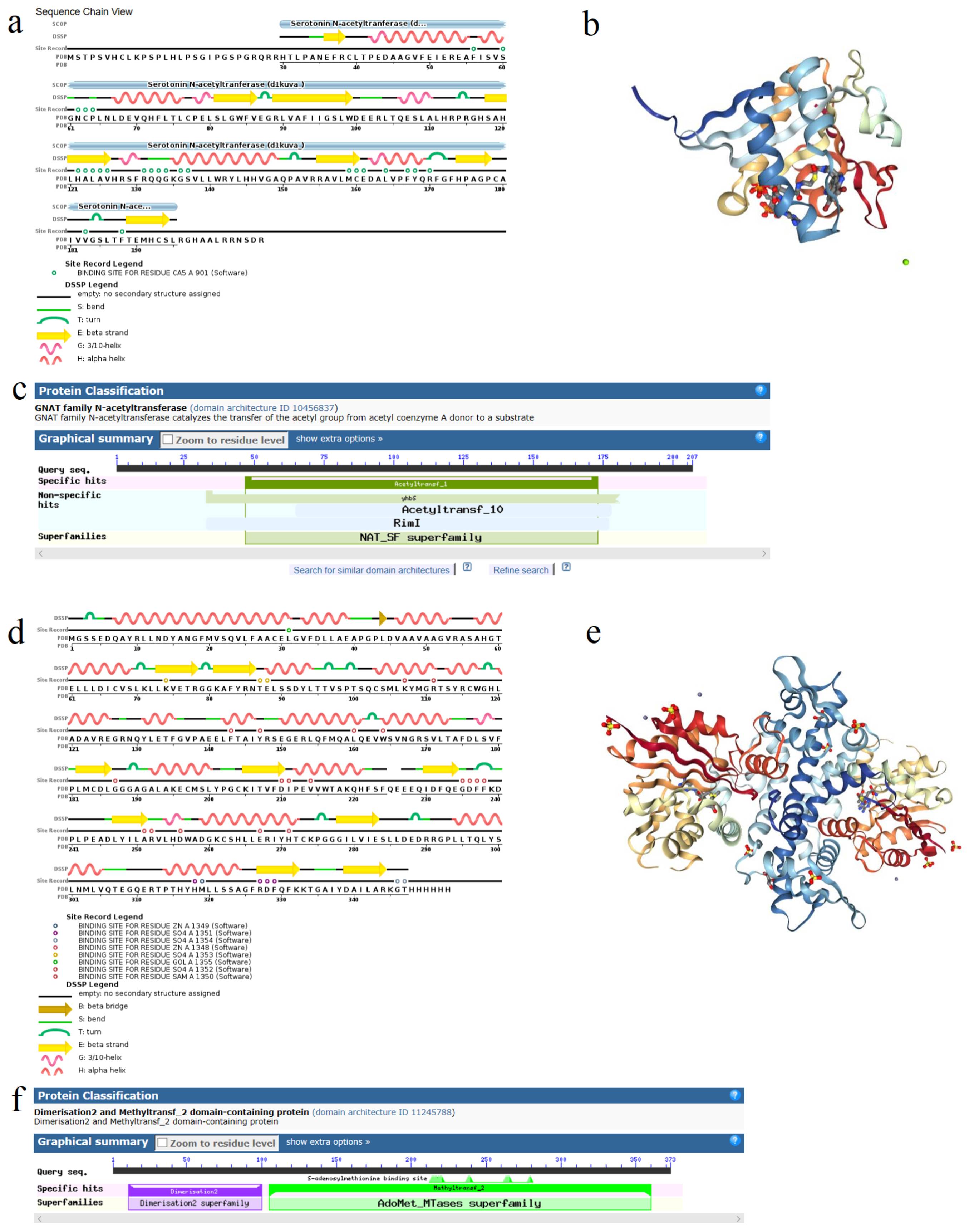
Sequence analysis revealed that AANAT and HIOMT genes encoded 207 amino acids and 373 amino acids, and the full lengths of their complete open reading frames were 624 bp and 1122 bp, respectively (Supplemental Fig. S1). According to protein sequence alignment, there was an Acetyltransf-1 conserved domain in the AANAT protein, which belongs to the NAT-SF superfamily and was related to acetyltransferase (Supplemental Fig. S2a−c); there were Dimerisation 2 and Methytransf-2 conserved domains in the HIOMT protein, which belongs to the Dimerisation 2 superfamily and Methytransf-2 superfamily, respectively, and were related to oxymethyltransferase (Supplemental Fig. S2d−f).
The plant expression vectors containing AANAT and HIOMT genes were transformed respectively into 'GL-3' apple by Agrobacterium-mediated leaf plate transformation. Three apple lines (A1, A3 and A6) with ectopic expression of AANAT and three apple lines (H1, H2 and H5) with ectopic expression of HIOMT were obtained. DNA and RNA identification analysis revealed that AANAT and HIOMT genes were successfully transformed into apple plants, respectively (Fig. 1b−e). Compared with WT (wild type), the levels of AANAT gene transcripts in A1, A3 and A6 were increased 45.32, 142.04 and 50.04-fold, respectively (Fig. 1b, d); the HIOMT gene transcript levels in H1, H2 and H5 were increased 52.06, 203.05 and 90.92-fold, respectively (Fig. 1c, e).
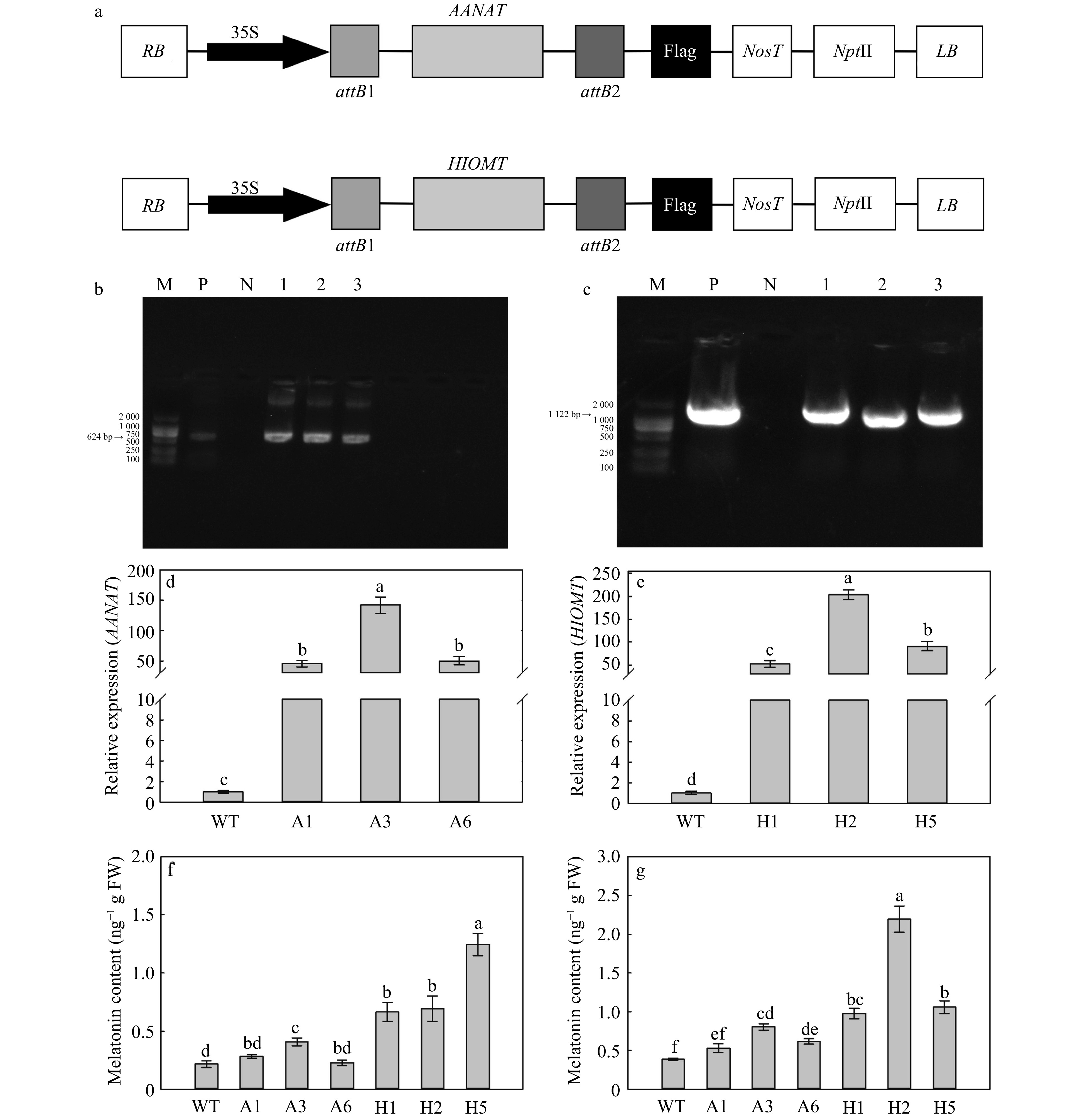
Figure 1. Identification and melatonin content of apple plants with ectopic expression of AANAT or HIOMT. (a) The structure of expression vectors of AANAT and HIOMT genes. PCR confirmation of apple plants with ectopic expression of AANAT (b) or HIOMT (c). Lanes: 1–3 (b) and 1–3 (c), transgenic lines to be tested; M, molecular marker DL 2000; P, positive control; N, negative control. Gene expression levels of AANAT (d) and HIOMT (e). Melatonin content under control (f) and UV-B conditions (g). Different letters indicate significant differences based on Tukey's multiple range test (p < 0.05). WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
The content of melatonin in transgenic plants was detected by HPLC-MS/MS. The content of melatonin was highest (1.24 ng g−1 FW) in H5 under normal growth conditions, which was 5.78-fold higher compared with that in the WT. The melatonin content was higher in most AANAT or HIOMT transgenic lines than in WT plants under both normal and UV-B conditions (Fig. 1f, g). In addition, the melatonin content was higher in HIOMT transgenic lines than in AANAT transgenic lines (Fig. 1f, g).
AANAT or HIOMT ectopic expression decreased the accumulation of
${{\text{O}}}_{\text{2}}^{\text{−}}$ and H2O2 -
After 15 d of UV-B treatment, the distribution of
${{\text{O}}}_{\text{2}}^{\text{−}} $ and H2O2 in mature leaves of apple plants with ectopic expression of AANAT or HIOMT was observed by NBT and DAB staining.${{\text{O}}}_{\text{2}}^{\text{−}} $ and H2O2 both accumulated in WT and transgenic plants after UV-B treatment. NBT staining showed that the number of blue spots on leaves was lower in most lines with ectopic expression of AANAT or HIOMT after UV-B treatment than in WT plants, especially for the A6 and H5 lines. These results indicated that${{\text{O}}}_{\text{2}}^{\text{−}} $ accumulation was lower in lines with ectopic expression of AANAT or HIOMT than in WT plants after UV-B stress (Fig. 2a). DAB staining showed that most of the lines with ectopic expression of AANAT or HIOMT had fewer yellow patches than WT after UV-B treatment, especially A6 and H2 lines. This indicated that AANAT or HIOMT ectopic expression decreased the accumulation of H2O2 in apple plants after UV-B stress (Fig. 2b).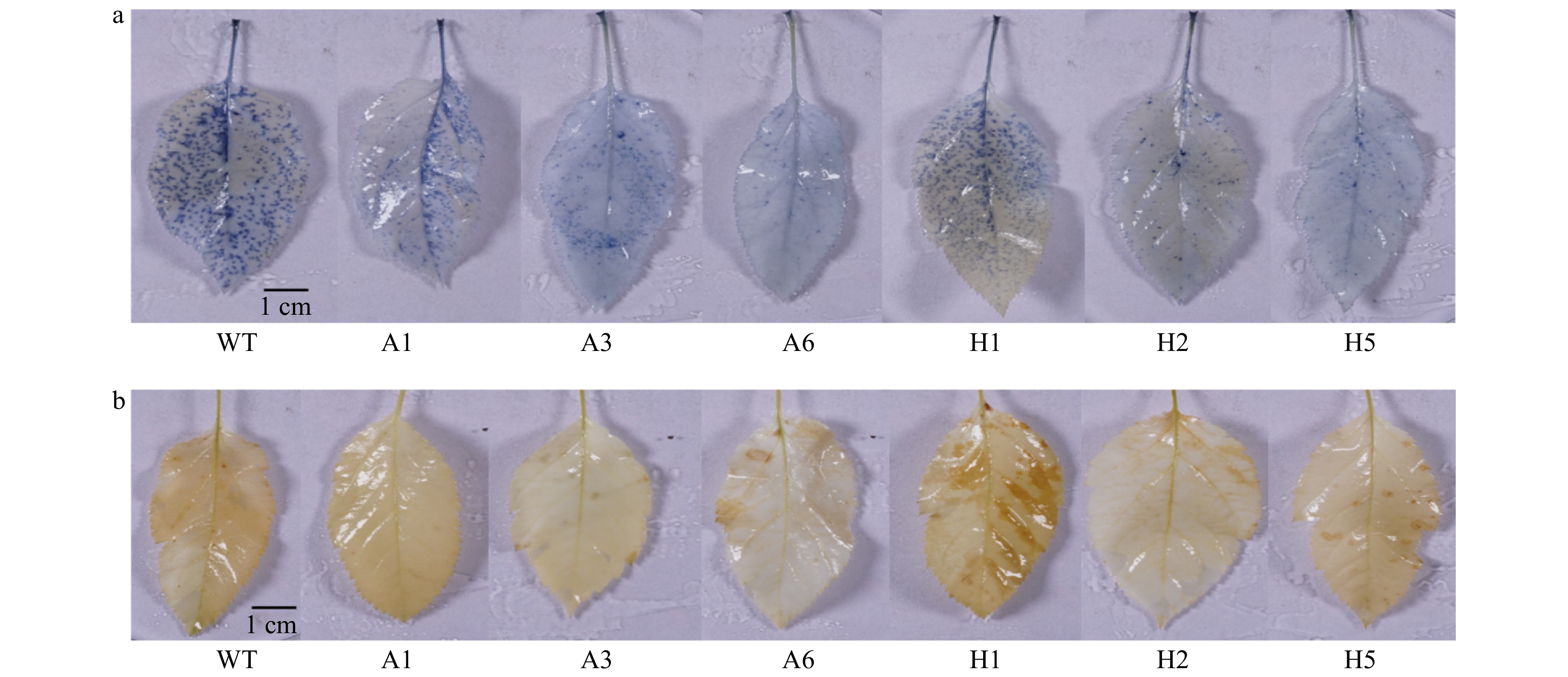
Figure 2. Chemical staining of
${{\text{O}}}_{\text{2}}^{\text{−}}$ (a) and H2O2 (b) in AANAT or HIOMT transgenic plants after UV-B treatment. WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.AANAT or HIOMT ectopic expression improved Fv/Fm and leaf area
-
Previous studies have found that high intensity UV-B can damage plant leaves, and long-term UV-B stress can alter the morphological structure of plants[21]. Therefore, we measured changes in the chlorophyll fluorescence of leaves of AANAT or HIOMT transgenic apples after 15 d of UV-B treatment. Before UV-B treatment, the fluorescence values of all genotypes were relatively high, and there were no significant differences observed among lines. After UV-B treatment, the Fv/Fm values of all genotypes were significantly reduced, and the spots reflecting the decrease in Fv/Fm values appeared in the chlorophyll fluorescence images (Fig. 3a). However, the Fv/Fm values were significantly higher in A1, H2 and H5 lines of transgenic plants than in the WT under UV-B stress (Fig. 3b).
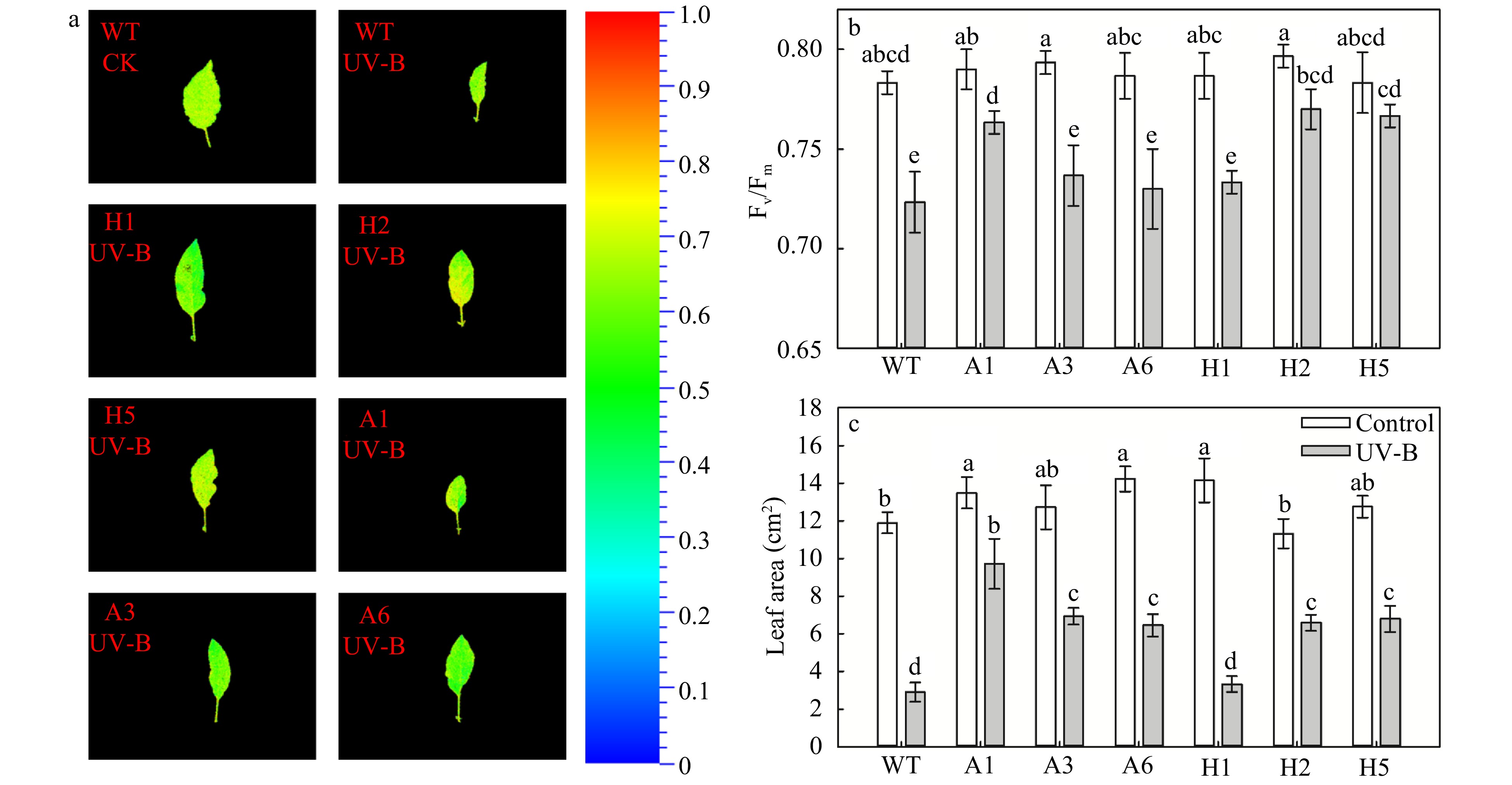
Figure 3. Analysis of chlorophyll fluorescence images (a), Fv/Fm (b), and leaf area (c) in AANAT or HIOMT transgenic plants after UV-B treatment. Different letters indicate significant differences based on Tukey's multiple range test (p < 0.05). WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
We also found that after 15 d of UV-B treatment, the area of the top leaves was reduced, and the leaves were damaged compared with the leaves before treatment. Nevertheless, the leaf area was significantly larger in most transgenic lines (A3, A6, H1, H2 and H5) than in the WT under UV-B conditions, and the effect of UV-B stress was the weakest on H1 (Fig. 3c).
AANAT or HIOMT ectopic expression affected stomatal properties
-
To characterize the effects of AANAT or HIOMT ectopic expression and UV-B treatment on the stomata, we examined the stomatal properties of leaves using SEM (Fig. 4a, b). In the absence of UV-B treatment, most of the AANAT or HIOMT transgenic lines did not significantly differ in stomatal length, width, aperture and density compared with WT plants (Fig. 4a−f). The stomatal length was significantly increased in H1 lines than in the WT (Fig. 4b, c). After 15 d of UV-B treatment, the stomatal length and width of WT plants were significantly decreased, and the stomatal density was significantly increased. However, the stomatal length and width were significantly higher, and the stomatal density significantly lower, in AANAT or HIOMT transgenic lines than in the WT under UV-B treatment (Fig. 4a−d, f). After UV-B treatment, the stomatal aperture did not significantly differ between transgenic lines and WT plants; the one exception was the stomatal aperture of H5, which was significantly higher compared with the WT (Fig 4b, e).
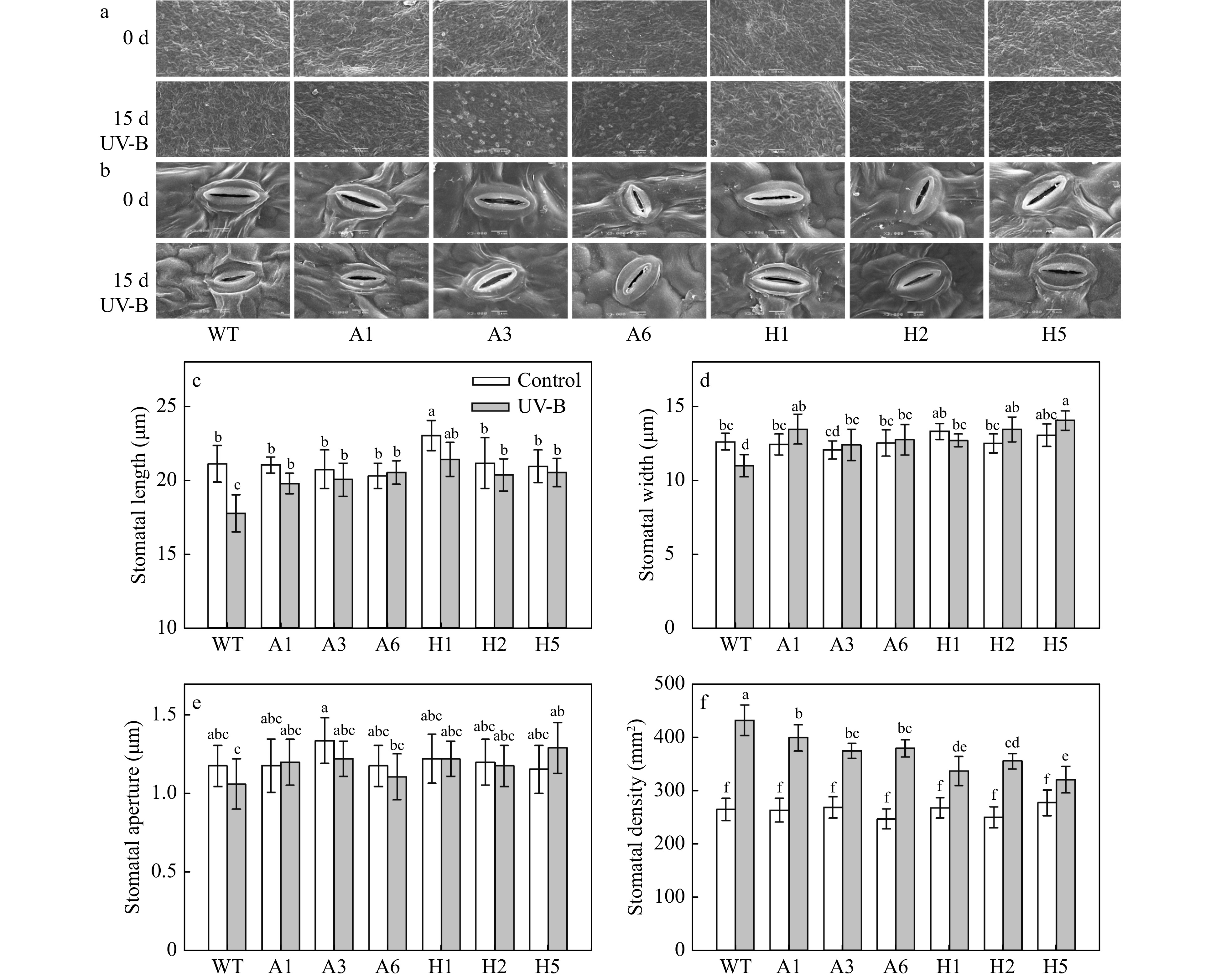
Figure 4. SEM images (a, 300×; b, 3,000×) and properties (c–f) of stomata from transgenic plants after UV-B treatment. (c) Stomatal length, (d) stomatal width, (e) stomatal aperture and (f) stomatal density. Different letters indicate significant differences based on Tukey's multiple range test (p < 0.05). WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
AANAT or HIOMT ectopic expression affected the content of phenolic compounds
-
Under UV-B stress, exogenous melatonin could increase the content of most phenolic compounds in Malus hupehensis and promote the expression of genes related to phenolic synthesis[21]. To further investigate the role of melatonin in tolerating UV-B stress, we measured the content of phenolic compounds in plants with ectopic expression of AANAT or HIOMT. Two benzoic acids (syringic acid and caffeic acid) and four hydroxycinnamic acids (gallic acid, chlorogenic acid, β-coumaric acid and ferulic acid) were detected by HPLC analysis; chlorogenic acid (hydroxycinnamic acid) was the most abundant after UV-B stress (Fig. 5a). In addition, UV-B treatment increased the content of syruvic acid, β-coumaric acid and ferulic acid (Fig. 5b, e, and f). Under UV-B stress, variation in the gallic acid content was not obvious (Fig. 5c). The content of caffeic acid in apple plants under UV-B treatment was decreased compared with normal conditions (Fig. 5d). However, the content of caffeic acid was higher in transgenic lines than in the WT (Fig. 5d). Moreover, the β-coumaric acid and ferulic acid content were significantly higher in lines with ectopic expression of HIOMT than in lines with ectopic expression of AANAT or WT plants under UV-B stress (Fig. 5e, f). In addition, variation in the sum of benzoic acids and hydroxycinnamic acids was consistent with variation in chlorogenic acid (Fig. 5g). Under normal conditions, there were no significant differences in the total content of benzoic acids and hydroxycinnamic acids between WT and transgenic plants (Fig. 5g); the exception was H1 line, in which the total content of benzoic acid and hydroxycinnamic acid was significantly higher compared with the WT. However, UV-B treatment could significantly increase the total content of these compounds in WT and transgenic plants. The total content of benzoic acids and hydroxycinnamic acids was markedly higher in A3, A6, H1 and H2 transgenic lines than in the WT.
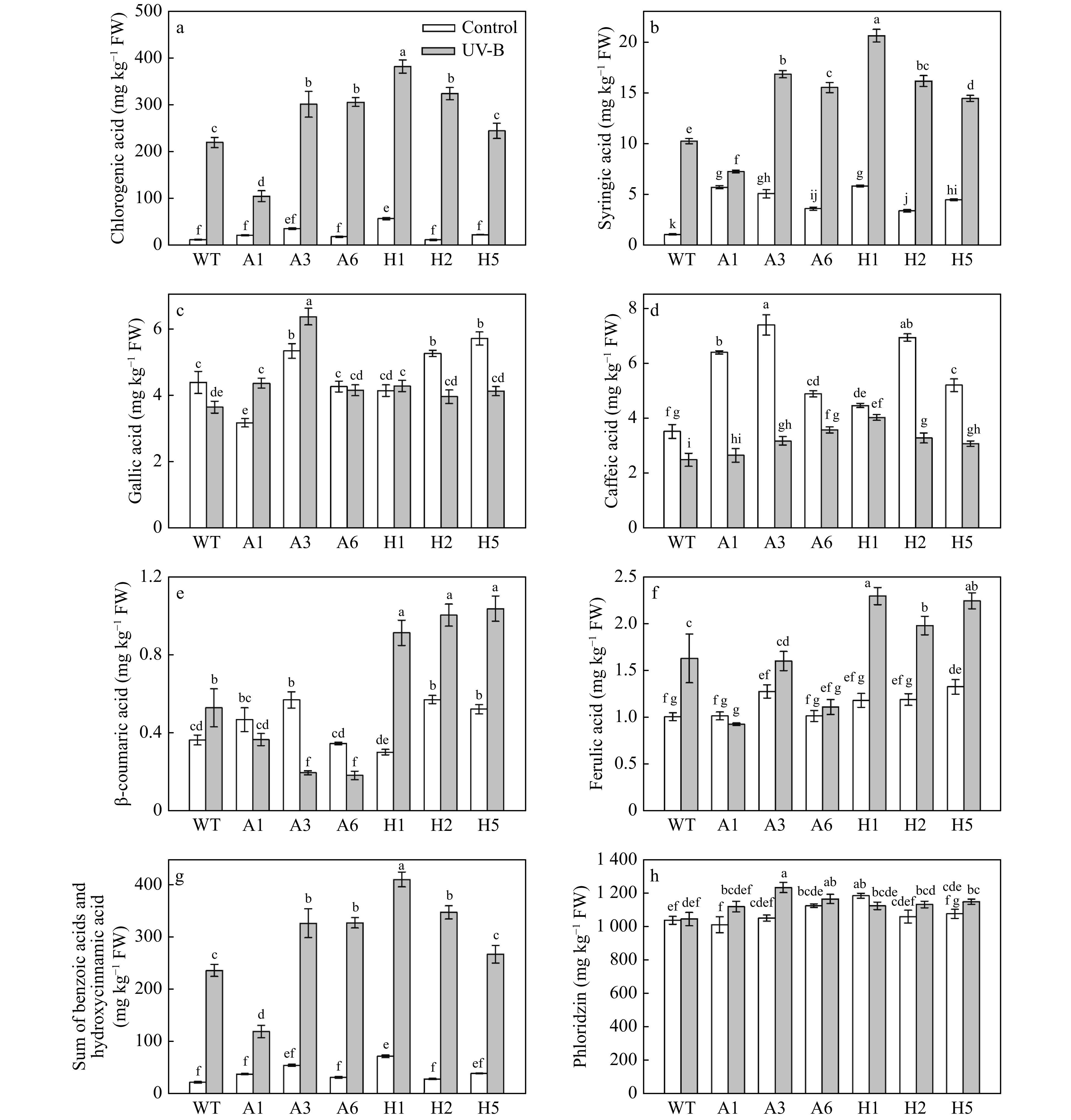
Figure 5. Analysis of benzoic acids, hydroxycinnamic acids and phloridzin in AANAT or HIOMT transgenic apple leaves after UV-B treatment. (a) Chlorogenic acid, (b) syringic acid, (c) gallic acid, (d) caffeic acid, (e) β-coumaric acid, (f) ferulic acid, (g) sum of benzoic acids and hydroxycinnamic acids and (h) phloridzin. Different letters indicate significant differences based on Tukey's multiple range test (p < 0.05). WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
Phloridzin is one of the main phenolic compounds in apple, and it was the phenolic compound with the highest content measured in this experiment. Here, phloridzin was the only one of the dihydrochalcones detected. Without UV-B treatment, the phloridzin content in most of the plants with ectopic expression of AANAT or HIOMT was increased compared with the WT (Fig. 5h). Moreover, the content of phloridzin was the highest in H1 (1,181.4 mg kg−1), which was 14.1% higher compared with that in the WT. Under UV-B stress, the phloridzin content in most transgenic apple plants was increased. The phloridzin content in A3 was the highest (1,230.8 mg kg−1), which was 17.5% higher compared with the WT. In addition, the phloridzin content of H1 was lower after UV-B stress compared with the treatment without UV-B, but it was still higher compared with the WT. The phloridzin content in WT leaves before and after UV treatment accounted for 58.7% and 50.7% of the total phenolic compounds, respectively. However, H1, which had the highest phloridzin content before UV treatment, accounted for 49.0% and 36.1% of the total phenolic content before and after UV treatment, respectively.
Next, the content of the four flavanols procyanidin B1, procyanidin B2, catechin acid and epicatechin was determined. After UV-B treatment, the procyanidin B1 and catechin acid content in WT plants and most transgenic lines were increased (Fig. 6a, c). However, the content of procyanidin B2 was decreased (Fig. 6b). The changes in epicatechin among treatments were subtle (Fig. 6d). Among the four flavanols measured, the content of procyanidin B1 was the highest, accounting for most of the total flavanols (Fig. 6a). The variation in total flavanol content was similar to the variation in procyanidin B1 (Fig. 6a, e). Under normal growth conditions, the total amount of flavanols was significantly higher in A1, A3, A6, H2 and H5 transgenic lines than in the WT (Fig. 6e). The A3 lines had the highest total flavanols (157.5 mg kg−1), which was 187.7% of the total flavanols in the WT. UV-B treatment significantly increased the total content of flavanols in apple. The total flavanol content was markedly higher in most transgenic lines compared with the WT; the content of H1 transgenic lines was the highest (386.9 mg kg−1).
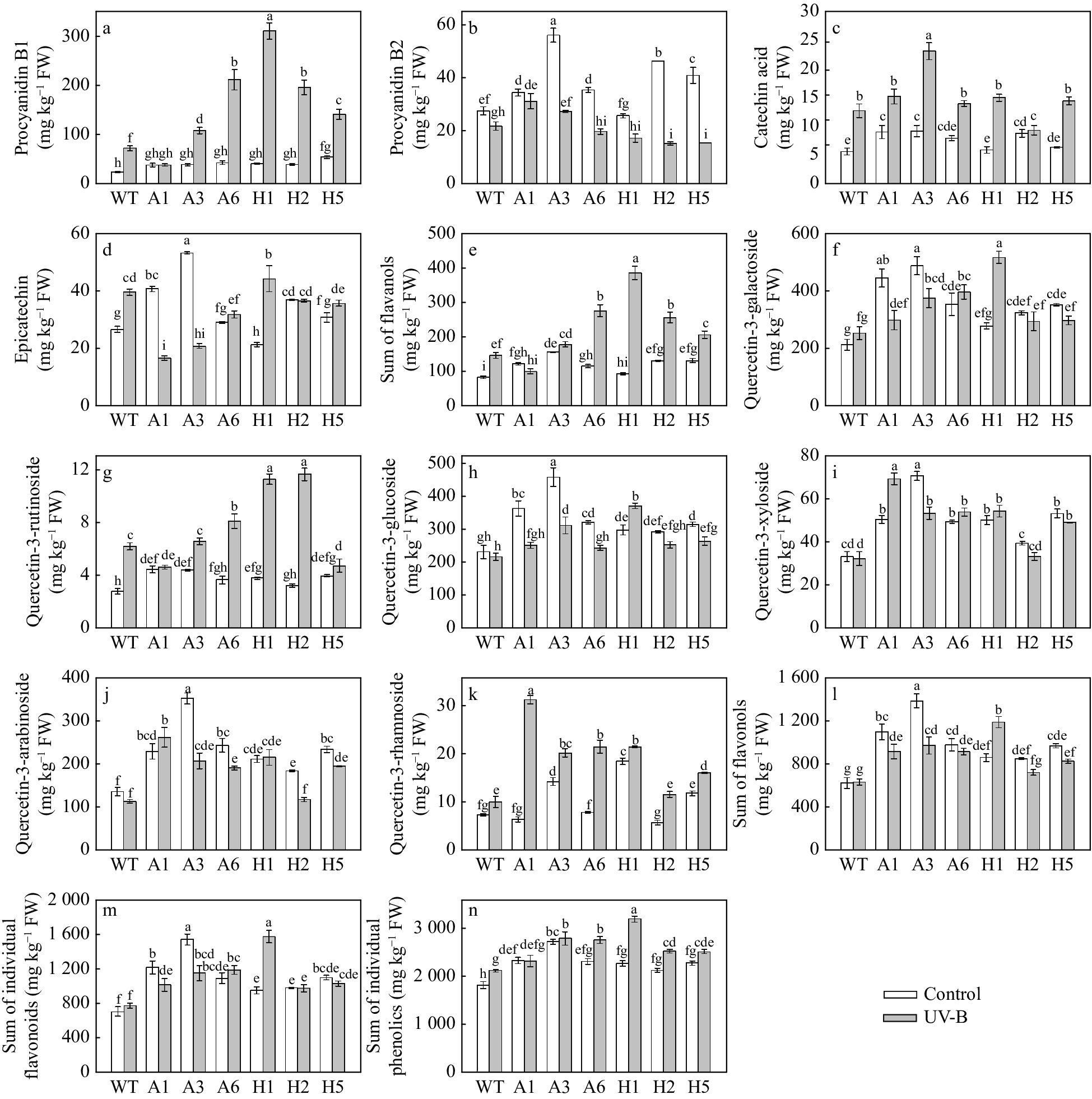
Figure 6. Analysis of flavanols, flavonols and sum of individual phenolics in AANAT or HIOMT transgenic apple leaves after UV-B treatment. (a) Procyanidin B1, (b) procyanidin B2, (c) catechin acid, (d) epicatechin, (e) sum of flavanols, (f) quercetin-3-galactoside, (g) quercetin-3-rutinoside, (h) quercetin-3-glucoside, (i) quercetin-3-xyloside, (j) quercetin-3-arabinoside, (k) quercetin-3-rhamnoside, (l) sum of flavonols, (m) sum of individual flavonoids and (n) sum of individual phenolics. Different letters indicate significant differences based on Tukey's multiple range test (P < 0.05). WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
Flavonols are important phenolic compounds in plants that play an important role in plant growth and development and resistance to environmental stress. Under normal growing conditions, quercetin-3-galactoside (A1, A3, A6, H2, H5) (Fig. 6f), quercetin-3-rutinoside (A1, A3, H1, H5) (Fig. 6g), quercetin-3-glucoside (A1, A3, A6, H1, H2, H5) (Fig. 6h), quercetin-3-xyloside (A1, A3, A6, H1, H5) (Fig. 6i), quercetin-3-arabinoside (A1, A3, A6, H1, H2, H5) (Fig. 6j) and quercetin-3-rhamnoside (A3, H1, H5) (Fig. 6k) content in some apple plants with ectopic expression of AANAT or HIOMT were significantly higher compared with WT plants. Quercetin-3-galactoside (Fig. 6f), quercetin-3-glucoside (Fig. 6h) and quercetin-3-arabinoside (Fig. 6j) content were relatively high among flavonols. Moreover, all three of these compounds were markedly increased in most transgenic lines under UV-B stress. After UV-B treatment, the quercetin-3-rutinoside and quercetin-3-rhamnoside content were increased in most transgenic lines, and the content of the other flavonols did not change consistently in the transgenic lines (Fig. 6f−k). However, flavonol content was still higher in most transgenic lines than in the WT under UV-B conditions (Fig. 6f−k). Total flavonols were markedly higher in plants with ectopic expression of AANAT or HIOMT than in WT plants under both normal and UV-B conditions (Fig. 6l). Total flavonol content in most lines was decreased under UV-B treatment compared with the treatment without UV-B; the exception was total flavonol content (412.3 mg kg−1) in H1 lines, which was increased in the UV-B treated samples (Fig. 6l).
We also determined the total flavonoid content and total phenolic content. The total flavonoid content in this experiment, including the sum of flavanols and flavonols, showed a pattern of variation similar to that for flavonols (Fig. 6m). The total phenolic content was markedly higher in most transgenic lines than in WT plants both with and without UV-B treatment (Fig. 6m). The total phenolic content (A6, H1, H2, H5) was significantly increased in WT plants and most transgenic lines under UV-B stress. Under normal conditions, A3 lines had the highest total phenolic content (as high as 2,643 mg kg−1). After UV-B treatment, the total phenolic content of H1 lines was the highest (as high as 3,106 mg kg−1).
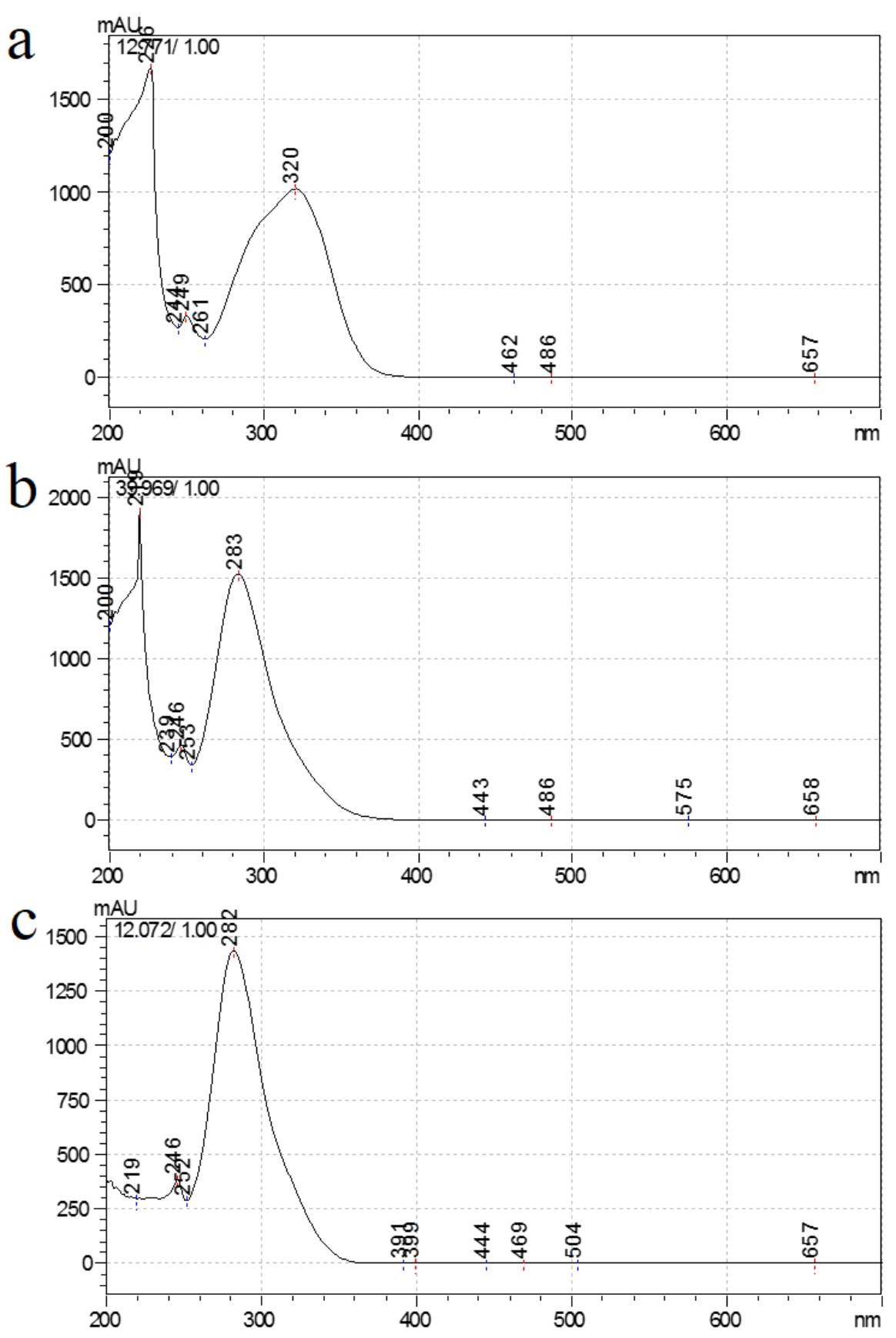
We also analyzed the UV–Vis spectra of chlorogenic acid, phloridzin and procyanidin B1, which were significantly increased in transgenic lines after UV-B stress. Chlorogenic acid had an absorption peak near 320 nm (Supplemental Fig. S3a). In addition, phoridin and procyanidin B1 had absorption peaks at 283 nm and 282 nm, respectively (Supplemental Fig. S3b, c). These three compounds show strong absorption of UV-B radiation. Therefore, the increase in the content of these three compounds may be related to the enhanced tolerance of transgenic lines to UV-B stress.
AANAT or HIOMT ectopic expression affected the expression of genes involved in antioxidant processes and the metabolism of phenolic compounds
-
To further study the defense mechanisms of plants with ectopic expression of AANAT or HIOMT against UV-B and the mechanism by which phenolic compounds change under UV-B stress, the genes of some antioxidant enzymes and enzymes related to phenolic metabolism were analyzed by qRT-PCR (Fig. 7).
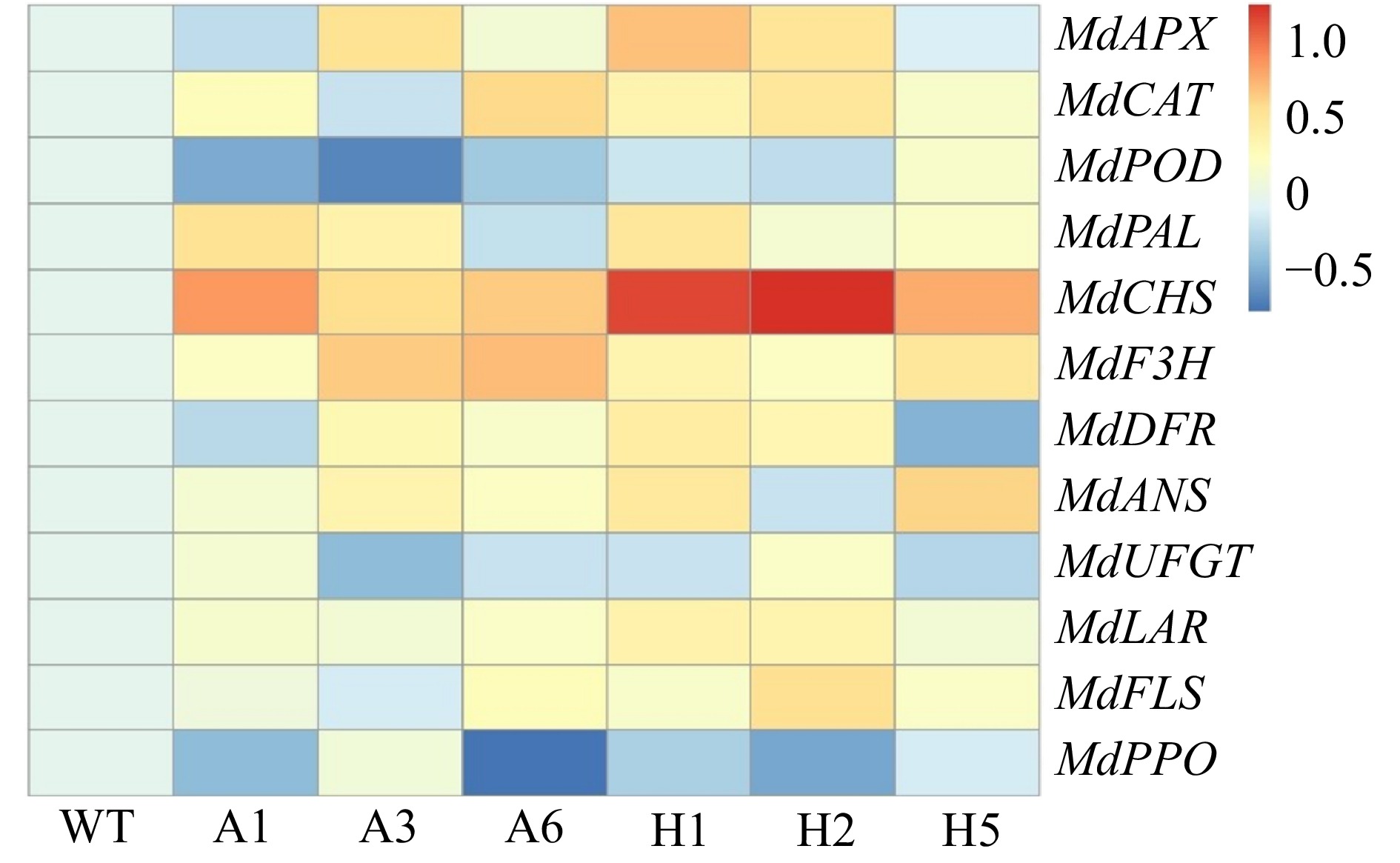
Figure 7. Heatmap showing transcripts of 3 antioxidant enzyme-related genes (MdAPX MdCAT and MdPOD) and 9 phenolic metabolism enzyme-related genes (MdPAL, MdCHS, MdF3H, MdDFR, MdANS, MdUFGT, MdLAR, MdFLS and MdPPO) in AANAT or HIOMT transgenic apple leaves after UV-B treatment. WT, wild type. A1, AANAT ectopic expression line 1. A3, AANAT ectopic expression line 3. A6, AANAT ectopic expression line 6. H1, HIOMT ectopic expression line 1. H2, HIOMT ectopic expression line 2. H5, HIOMT ectopic expression line 5.
After UV-B stress, the expression levels of antioxidant enzyme-related genes in transgenic lines were increased compared with WT plants. The expression levels of MdAPX and MdCAT in the WT and most lines with ectopic expression of AANAT or HIOMT were up-regulated, and the expression levels of MdAPX and MdCAT were significantly higher in lines with ectopic expression of AANAT or HIOMT than in the WT. However, the expression of MdPOD was down-regulated. These results indicated that MdAPX and MdCAT played a key role in the process of H2O2 scavenging under UV-B stress.
Under UV-B treatment, the transcripts of most phenolic metabolism-related enzymes, such as MdPAL, MdCHS, MdF3H, MdDFR, MdANS, MdLAR and MdFLS, were significantly increased compared with the WT; the exception was MdUFGT. Among the genes with up-regulated expression after UV stress, MdCHS was the most up-regulated. The transcription level of MdCHS was markedly higher in transgenic lines than in the WT. The expression levels of A1, A3, A6, H1, H2 and H5 lines were increased by 7.5, 4.0, 4.9, 15.1, 18.7 and 6.4-fold, respectively, compared with the WT. MdCHS is an upstream gene in phenolic synthesis, and the up-regulated expression of MdCHS may be related to the accumulation of a variety of phenolic compounds. In addition, the expression of MdPPO in most transgenic lines was markedly down-regulated compared with the WT under UV-B stress.
-
As a small indole compound with multiple functions, melatonin plays an important role in many aspects of plant growth and development[31]. Our previous studies have examined the effect of exogenous melatonin on the resistance of apple to photooxidative stress and UV-B stress[21,32]. However, the production and storage of melatonin requires dark conditions, which are not conducive to production. Therefore, finding a convenient and low-cost method that permits the continuous production of melatonin is critically important. Transgenic technology could potentially provide a solution to this problem[33,34].
Transgenic studies of enzymes related to melatonin biosynthesis have been carried out in many plants. The genes encoding AANAT and HIOMT enzymes have been most commonly used in transgenic research on melatonin[30,35], and other melatonin biosynthesis genes, such as T5H[33] and TDC[36], have also been studied. Most of the transformed genes are from animals, and research has primarily focused on the heterologous expression of melatonin synthesis genes[30]. Ectopic expression of the melatonin synthesis genes increases endogenous melatonin content in plants[28].
Although transgenic studies relating to melatonin have been carried out in a variety of plants (e.g., rice, switchgrass, tomato)[30,33,35], few studies have been conducted in apple. The low regeneration rate of apple leaves makes genetic transformation difficult and limits the development and application of apple transgenic technology. Here, transgenic-resistant apple buds were successfully obtained by genetic transformation according to human AANAT and HIOMT sequences (Supplemental Fig. S1 and S2). The AANAT and HIOMT genes were successfully transfected into the 'GL-3' apple genome by DNA and RNA detection, respectively (Fig. 1a−d). In apple lines with ectopic expression of AANAT or HIOMT, the transcripts of AANAT and HIOMT were increased by 45.32–142.04-fold and 52.06–203.05-fold, respectively (Fig. 1c, d). Wang et al[37] found that the transgenic tomato lines of oHIOMT and oAANAT had significantly higher melatonin content compared with the WT, and the content of melatonin was higher in the oHIOMT transgenic plants than in oAANAT transgenic plants. In this experiment, the melatonin content in most transgenic apple seedlings was increased (Fig. 1e), which was also consistent with the findings of Zhang et al[38]. In addition, ASMT, which encodes the last enzyme in rice melatonin synthesis and is a homolog of HIOMT, plays a rate-limiting role in the melatonin synthesis pathway[39]. Therefore, our results suggest that AANAT and HIOMT are rate-limiting enzymes in plants.
Previous studies have shown that UV-B radiation can cause oxidative stress in plants and increase the content of ROS, such as
${{\text{O}}}_{\text{2}}^{\text{−}} $ and H2O2[40]. By contrast, the${{\text{O}}}_{\text{2}}^{\text{−}} $ and H2O2 content were considerably lower in apple plants with ectopic expression of AANAT or HIOMT than in WT plants (Fig. 2); the transgenic lines thus maintained a strong scavenging ability of ROS under UV-B stress. Our previous research has shown that the application of exogenous melatonin dramatically inhibited the production of H2O2 and significantly increased the antioxidative activities of APX, POD and CAT[21]. The qRT-PCR analysis showed that UV-B stress dramatically increased the expression of MdAPX and MdCAT in most WT and transgenic lines (Fig. 8). Moreover, their expression levels under UV-B stress were markedly higher in transgenic lines than in the WT (Fig. 8). The similar results obtained in this study suggest that this pathway may also play a role in AANAT and HIOMT-mediated apple tolerance to UV-B stress.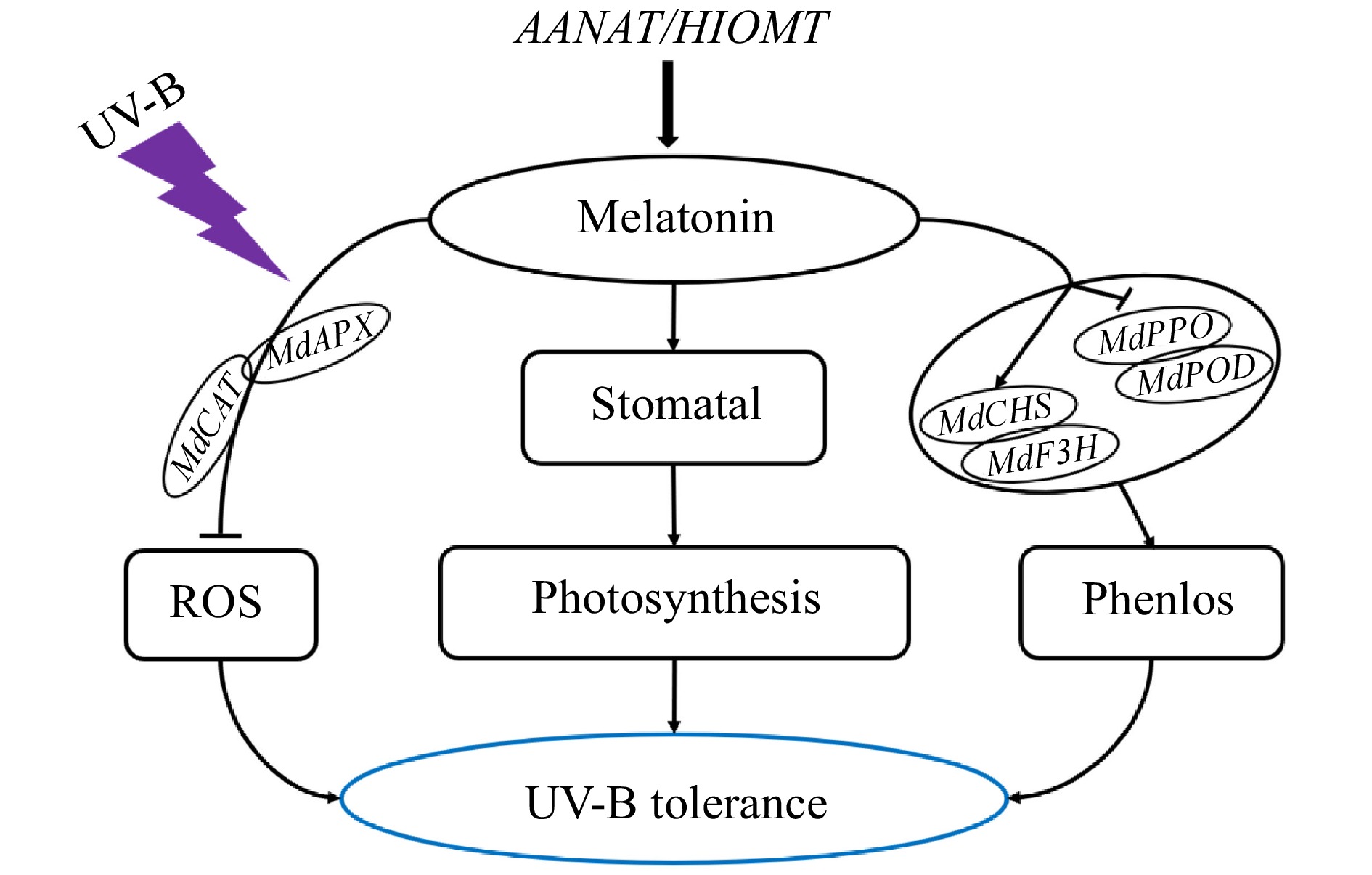
Figure 8. A model of the regulatory effect of AANAT or HIOMT on apple's UV-B stress response. Under UV-B stress, the melatonin content of apple lines with ectopic expression of AANAT or HIOMT was significantly increased. Thus, the expression of antioxidant enzyme-related genes was increased, and ROS levels were reduced; the stomatal shrinkage of transgenic lines caused by UV-B stress was alleviated, and the photosynthetic capacity was enhanced. In addition, the ectopic expression of AANAT or HIOMT not only increased the expression of genes related to phenolic synthesis but also inhibited the expression of genes related to phenolic degradation, thus increasing phenolic content. Therefore, the tolerance of transgenic apple plants to UV-B stress was improved.
Under environments with increased UV-B radiation, most plants show a reduced photosynthetic rate, which leads to decreased plant productivity[41,42]. Under UV stress, apple plants with ectopic expression of AANAT or HIOMT had higher photosynthetic capacity and could maintain higher Fv/Fm values compared with the WT (Fig. 3). These results suggested that AANAT or HIOMT ectopic expression could improve the tolerance of apple plants to UV-B stress by enhancing apple photosynthesis. The photosynthetic capacity and fluorescence parameters of Malus hupehensis hydroponic seedlings have been shown to improve through the application of exogenous melatonin under UV-B[21], salt[22] and drought[23] stress. The melatonin produced in apple plants with ectopic expression of AANAT or HIOMT might thus serve a continuous function in apple plants in the stress response, which produces an effect similar to exogenous melatonin application.
The stomatal sensitivity to the external environment increases as the strength of UV-B radiation increases, which limits the absorption and assimilation of CO2[43]. To determine why transgenic plants maintained high photosynthetic capacity under UV-B stress, the stomata of plants with ectopic expression of AANAT or HIOMT were studied by SEM. Under UV treatment, the stomatal length and width were significantly higher in lines with ectopic expression of AANAT or HIOMT than in WT plants (Fig. 4). Consistent with the previous study[23], the effect of drought stress on stomatal length and width was markedly weaker for apple leaves treated with exogenous melatonin than for untreated apple leaves. These results indicated that AANAT or HIOMT ectopic expression alleviated the negative effect of UV-B stress on the stomata of apple leaves, thereby mitigating the decrease in the photosynthetic rate caused by UV-B radiation.
Phenols are important secondary metabolites in plants that can act as antioxidants to scavenge excess ROS caused by external stress[44]. They can also accumulate on the surface of plants and reduce UV-B transmission through the plant epidermis and thus protect plants against UV-B radiation[12,45,46]. UV-B stress considerably increased the total amount of benzoic acid and hydroxycinnamic acid in lines with ectopic expression of AANAT or HIOMT compared with plants that were not treated with UV-B radiation, and the change in chlorogenic acid was the most significant (Fig. 5). As a phenylpropanoid compound, chlorogenic acid is produced by the shikimic acid pathway and has a strong scavenging effect on alkylperoxy radicals in plants[14]. UV-B treatment increased the accumulation of chlorogenic acid in apple plants, and the content of chlorogenic acid was significantly higher in most transgenic lines than in WT plants (Fig. 5a and Supplemental Fig. S3a). Therefore, AANAT or HIOMT ectopic expression may enhance the ability of apple plants to defend against UV-B by promoting the synthesis of chlorogenic acid. As a dihydrochalcone-like compound, phloridzin is widely present in Malus and accounts for more than 90% of the total soluble phenols in some apple leaves[47]. The observed changes in phloridzin content and chlorogenic acid were similar (Fig. 5h and Supplemental Fig. S3b). Furthermore, the expression of MdCHS (a key gene in phloridzin biosynthesis) was noticeably higher in transgenic lines under UV-B stress than in the WT. This also confirmed that AANAT and HIOMT may alleviate UV-B stress in apples by increasing the content of phloridzin, which has strong antioxidant capacity[48]. In addition, the study of flavonoids revealed that the accumulation of flavonoids was markedly higher in most lines with ectopic expression of AANAT or HIOMT than in the WT with or without UV-B treatment (Fig 6). The content of procyanidin B1 was higher, and the pattern of change was more pronounced (Fig 6a and Supplemental Fig. S3c). Procyanidin not only plays a role in the growth and development of plants but also has anti-UV[49], anti-bacterial[50], and antioxidant activities[49]. These functions may be responsible for the high accumulation of procyanidin in apple lines with ectopic expression of AANAT or HIOMT. Chlorogenic acid, phloridzin and procyanidin B1 all have strong absorption peaks in the ultraviolet band (Supplemental Fig. S3). After UV-B treatment, their increase can enhance the UV shielding of leaves and thus the tolerance of apple plants to UV-B stress[15].
After apples are damaged by high light or UV, phenolic compounds can remove reactive oxygen species and thus help alleviate the damage caused by light stress[40]. Under normal growth conditions, the total phenolic content was significantly higher in most transgenic lines than in WT plants, and the expression levels of genes involved in phenolic metabolism were also increased (Fig. 6 and 7). In addition, the tolerance of plants to UV-B was improved by the increase in melatonin content, the decreased degradation of flavonols and the increase in the synthesis of other phenolic compounds[37,51]. After UV-B treatment, the total phenolic content and the transcripts of genes related to phenolic synthesis were higher in most transgenic lines than in WT plants (Fig. 6 and 7). Polyphenol oxidase (PPO) and peroxidase (POD) have been shown to play an important role in the oxidative metabolism of phenols[52]. The expression of MdPPO and MdPOD in most lines with ectopic expression of AANAT or HIOMT was down-regulated (Fig. 7). The down-regulated expression of MdPPO and MdPOD indicated that apple plants with ectopic expression of AANAT or HIOMT under UV-B stress could not only promote the expression of phenolic synthesis genes but could also promote the accumulation of phenolic substances by slowing the degradation of phenolic substances[53]. These results suggested that the accumulation of phenolic compounds was promoted by the ectopic expression of AANAT or HIOMT. Moreover, transgenic plants maintained a high phenolic content even under UV-B stress, which enhances the tolerance of apple plants to UV-B radiation.
-
In this study, we analyzed the effects of AANAT or HIOMT ectopic expression on the tolerance of apple to UV-B stress (Fig. 8). The melatonin content of transgenic lines was higher, and that of lines with ectopic expression of HIOMT was significantly higher, than the melatonin content of lines with ectopic expression of AANAT. The resistance to UV-B stress mediated by AANAT and HIOMT was closely related to the improvement in antioxidant capacity, the maintenance of photosynthesis and the metabolism of phenols in transgenic apple plants. In addition, the limitation in the photosynthetic ability caused by stomatal shrinkage was alleviated through AANAT or HIOMT ectopic expression. Moreover, the increase in phenolic compounds in transgenic apple plants, especially chlorogenic acid, phloridzin and procyanidin B1, is critically important for the tolerance of transgenic apple to UV-B stress. Overall, this study provides evidence for the positive regulation of melatonin in apple UV-B stress. Generally, the results of this study indicate that melatonin could be used to enhance agricultural production.
-
AANAT (Accession # U40347) and HIOMT (Accession # M83779) in humans are key synthases in the melatonin synthesis pathway[28,29], while the last two steps of the melatonin synthesis pathway in plants and animals are the same[27]. Therefore, these two genes were selected for ectopic expression in apples in this study.
Gene sequence analysis was carried out on the NCBI website; the open reading frame of the genes was also translated into proteins on the NCBI website. The correlation analysis of the gene's structural domains was then carried out. Protein structure analysis and protein 3D structure prediction were performed in the RCSB PDB protein database. After obtaining AANAT and HIOMT sequences, the genes were synthesized by Genscript Biotech Corporation. The sequences of the synthesized genes were completely consistent with the original sequences of AANAT and HIOMT. The genes were then used in subsequent experiments.
Construction of vectors and generation of transgenic plants
-
'Gala' apple (M. domestica cv. Gala) used for heredity and transformation was a clonal GL-3 tissue-cultured vaccine. To construct the ectopic expression vector of apple lines, the coding regions of AANAT and HIOMT were inserted into the pGWB411 vectors driven by the CaMV 35S promoter, respectively. The vectors identified by sequencing were transformed in Agrobacterium tumefaciens strain EHA105. The transgenic apple plants were obtained as described previously[54]. The transgenic lines were preliminarily confirmed at the DNA level by polymerase chain reaction (PCR). AANAT and HIOMT ectopic expression were evaluated respectively by quantitative real-time polymerase chain reaction (qRT-PCR). Untransformed GL-3 lines (WT) were used as controls.
DNA extraction, RNA isolation and qRT-PCR analysis
-
Genomic DNA was extracted using the Wolact® Plant Genomic DNA Purification Kit (Wolact, Hong Kong, China). Total RNA was isolated using the Wolact Plant RNA Isolation Kit (Wolact, Hong Kong, China). One microgram total RNA was taken for the synthesis of the first-strand cDNA using the Thermo RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, MA, USA). qRT-PCR reactions were performed as previously described[55]. Transcripts of EF-1α in Malus were used to standardize the cDNA samples for different genes. All primer sequences used in this experiment are listed in Supplemental Table S1.
UV-B treatments
-
After 45 d on rooting media, WT and transgenic apple plants were transplanted into a growth chamber. After 50 d of growth, apple plants with consistent growth were selected from the seven apple lines (one WT line, three lines with ectopic expression of AANAT and three lines with ectopic expression of HIOMT) for UV-B experiments as described previously[21]. There were two treatments in the experiment: (1) control (CK), with eight fluorescent lamps (40 W, FSL Co., Ltd., Foshan, Guangdong, China) and a light intensity of 150 μmol m−2 s−1 and (2) UV-B treatment (UV-B), with four UV-B lamp tubes (40 W, Huaquiang Co., Ltd., Nanjing, Jiangsu, China), four fluorescent lamps, and a UV-B dosage of 0.45 W m−2. The photoperiod was 14 h of light (23–24 °C) and 10 h of darkness (16–18 °C). Water was added in the dark phase every day. The experiment lasted for 15 d. The samples were taken at 0 d and 15 d during UV-B treatment and stored at −80 °C after rapid freezing in liquid nitrogen. The physiological analyses were carried out on the last day of the experiment.
Measurements of melatonin content
-
Melatonin was extracted according to the method described in Pothinuch and Tongchitpakdee[56]. A total of 0.3 g of frozen leaf tissue was ground into a fine powder in liquid nitrogen with a mortar, and the frozen powder was suspended in 5 mL of methanol and treated with ultrasound (80 Hz) at 4 °C for 40 min. After centrifugation (4 °C, 15 min, 10,000 g), the supernatant was collected and dried with nitrogen. After resolution with methanol, the melatonin in the supernatant was further determined by high-performance liquid chromatography/tandem mass spectrometry (HPLC-MS/MS) based on the method of Zhao et al[57].
Chemical tissue staining of leaves
-
${{\text{O}}}_{\text{2}}^{\text{−}} $ was detected using the nitroblue tetrazolium (NBT) staining method. The leaves were soaked in HEPES buffer containing 1 mg mL−1 NBT (10 mM, pH = 7.5). After staining for 4 h, the stained leaves were rinsed with distilled water and transferred to 80% ethanol. The color was decolorized by boiling for 30 min. After the removal of chlorophyll, the leaves were preserved in 10% (v/v) glycerol and then photographed to detect${{\text{O}}}_{\text{2}}^{\text{−}} $ .H2O2 was detected using the diaminobenzidine (DAB) staining method. The leaves were immersed in tris-acetic acid buffer containing 1 mg mL−1 DAB (10 mM, pH = 5.0). They were then positioned in the dark at room temperature for approximately 8 h. After observation of brown spots, the stained leaves were washed with distilled water. The steps for decolorization, preservation, and photography were the same as those described for the NBT staining method.
Determination of chlorophyll fluorescence
-
Fv/Fm was measured using Open FluorCam FC 800-O following the method of Pérez-Bueno et al[58]. The data were analyzed by FluorCam7 software (PSI, Brno, Czech Republic).
Analysis of leaf area
-
Ten functional leaves were randomly selected from each treatment for leaf area measurements using an LI-3000C Portable Leaf Area Meter (LI-COR, USA) per the manufacturer's instructions.
Observations of leaf stomata
-
Mature leaves were collected from the same position of selected plants. After sampling, the leaves were fixed immediately in 4% glutaraldehyde solution prepared from 0.1 M phosphate buffered saline (PBS) (pH = 6.8). The samples were then rinsed with PBS five times (at 5, 10, 15, 20 and 30 min). Gradient dehydration was then carried out in ethanol, followed by vacuum drying and gold coating. Scanning electron microscopy (SEM) observations were performed using a Hitachi S-4800 microscope (Hitachi Ltd., Tokyo, Japan) at a magnification of 300× and 2,000×. Ten fields were randomly selected on the lower epidermis of leaves to record the number of stomata and calculate stomatal density. ImageJ was used to measure the aperture, width and length of stomata.
Determination of phenolic compounds
-
Phenolic compounds were determined by high-performance liquid chromatography (HPLC). Inertsil ODS-3 was used as a protective column (5.0 μm, 4.0 mm × 10.0 mm, GL Sciences Inc., Tokyo, Japan). The measured temperature was 30 °C, and the flow rate was 1.0 mL min−1. Two solvents (A, B) were used for gradient separation: solvent A consisted of 10% formic acid and water, and solvent B was an acetonitrile solution containing 10% formic acid and 1.36% water. The gradient was 95% A (0 min), 85% A (25 min), 78% A (42 min), 64% A (60 min) and 95% A (65 min)[15]. The post-operation time was 10 min. Procyanidin B1, procyanidin B2, epicatechin, syringic acid, catechin, gallic acid and phloridzin were measured at 280 nm; chlorogenic acid, caffeic acid, ρ-coumaric acid and ferulic acid were estimated at 320 nm; quercetin-3-rhamnoside, quercetin-3-glucoside, quercetin-3-galactoside, quercetin-3-xyloside, quercetin-3-arabinoside and quercetin-3-rutinoside were determined at 365 nm[21].
The UV–Vis spectra of chlorogenic acid, phloridzin and procyanidin B1 in leaves were determined using a Unispec-SC spectrometer (PP Systems, Hansatech, USA).
Statistical analysis
-
SPSS 20.0 and Excel software were used to statistically analyze the data. SigmaPlot 12.0 software was used to construct the graphical representations. Multiple comparison analyses were performed using the Tukey method (P < 0.05). Data were expressed as mean ± standard deviation.
- This work was supported by the National Natural Science Foundation of China (31972389), the earmarked fund for the China Agriculture Research System (CARS-27), the Key S&T Special Projects of Shaanxi Province (2020zdzx03–01-02) and Tang Scholar. The authors are grateful to Dr. Zhihong Zhang of Shenyang Agricultural University for donating the 'Gala' apple (M. domestica cv. Gala) plants.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Nucleotide sequences of AANAT (a) and HIOMT (b).
- Supplemental Fig. S2 Domain (a, c, d, f) and three-dimensional structure models (b, e) of AANAT (a–c) and HIOMT (d–f).
- Supplemental Fig. S3 The UV–Vis spectra of chlorogenic acid (a), phloridzin (b) and procyanidin B1 (c) in AANAT or HIOMT transgenic apple leaves after UV-B treatment.
- Supplemental Table S1 Sequences of primers used in qRT-PCR.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu X, Wei Z, Gao T, Zhang Z, Tan K, et al. 2021. Ectopic expression of AANAT or HIOMT improves melatonin production and enhances UV-B tolerance in transgenic apple plants. Fruit Research 1: 4 doi: 10.48130/FruRes-2021-0004

Ectopic expression of AANAT or HIOMT improves melatonin production and enhances UV-B tolerance in transgenic apple plants
- Received Date: 04 April 2021
- Accepted Date: 08 June 2021
- Published Online: 23 June 2021
Abstract: Melatonin is involved in plant responses to various environmental stresses. Although many studies have demonstrated that the tolerance of plants to stress is improved by exogenous melatonin, the role of endogenous melatonin metabolism in the response of apples to UV-B stress remains unclear. Here, the human melatonin biosynthesis-related enzyme genes AANAT or HIOMT were transformed into 'GL-3' apple, and the transgenic lines were treated with UV-B stress. The ectopic expression of AANAT or HIOMT significantly increased the melatonin content in apples. After UV-B stress, the tolerance of apple lines with ectopic expression of AANAT or HIOMT was markedly improved. The decrease in chlorophyll fluorescence, the generation of reactive oxygen species and the shrinkage of stomata caused by UV-B stress were alleviated by AANAT or HIOMT ectopic expression. In addition, the total phenolic content was markedly increased in the transgenic lines compared with the WT (wild type). The increase in phenolic compounds was related to the increase in benzoic acid, hydroxycinnamic acid, dihydrochalcones and flavanols, among which increases in chlorogenic acid, phloridzin and procyanidin B1 content were most prominent. Furthermore, the transgenic lines did not only promote the expression of genes related to phenolic synthesis under UV-B stress, but they also increased the accumulation of phenolic compounds by inhibiting the expression of MdPPO and MdPOD related to phenolic degradation. In summary, our results demonstrate that AANAT- or HIOMT-mediated melatonin synthesis improved the tolerance of apples to UV-B stress, mainly by scavenging reactive oxygen species, increasing photosynthetic capacity and increasing total phenolic content.
{kind=link}
{kind=link}
{kind=link}