









-
In the phenylalanine metabolic pathway, F3'H catalyzes naringenin to produce dihydroquercetin, while F3'5'H catalyzes naringenin to produce pentahydroxyflavone. Subsequently, the products of naringenin catalyzed by F3H are further catalyzed by dihydroflavonol 4-reductase and anthocyanin synthase to produce cyanidin, which is scarlet red. Following continuous catalysis, naringenin catalyzed by F3'5'H produces delphinidin, which is blue[1−3] (Fig. 1). Thus, the amount of cyanidin synthesized is positively correlated with the expression of F3'H, and the amount of delphinidin biosynthesis is positively correlated with the expression of F3'5'H. For instance, in blue gentian, the inhibition of F3'5'H expression turned the flower color to red[4]. These catalytic differences are attributed to the number and position of hydroxyl substitutions in the B-ring of dihydrokaempferol.

Figure 1. Branch pathway of anthocyanin metabolism. F3H, flavanone 3-hydroxylase; DFR, dihydroflavonol 4-reductase; ANS, anthocyanidin synthase; 3GT, UDP-glucose: anthocyanidin 3-O-glucosyltransferase; MaT, malonyl-CoA: anthocyanin 3-malonyltransferase; F3'H, flavonoid 3'-hydroxylase; F3'5'H, flavonoid 3',5'-hydroxylase.
Both F3'H and F3'5'H are cytochrome P450 enzymes[1]. These enzymes diverged at the early stages of vascular plant evolution, as their amino acid sequence identity was < 50%. Phylogenetic analysis of Asteraceae plants shows that F3'5'H of Pericallis, Osteospermum, and Callistephus may have evolved from F3'H[5, 6]. Moreover, threonine and serine at the amino acid position 487 of F3'H in Gerbera hybrida and F3'5'H in Osteospermum hybrid are interchangeable. In other words, F3'H of Gerbera hybrida showed little F3'5'H activity and F3'5'H of Osteospermum hybrid showed strong F3'H activity in vitro after the amino acid substitutions[7]. Furthermore, due to the similar physicochemical properties between threonine and serine, F3'H and F3'5'H are interchangeable and each exhibit the same activity. Although demonstrated in an in vitro functional assay, the interchangeability of serine and threonine at the amino acid position 487 remains to be proven in vivo.
Chrysanthemums lack the F3'5'H gene and CmF3'H catalyzes cyanidin synthesis. Therefore, due to their inability to synthesize delphinidin, there are no blue or blue-purple chrysanthemums occurring naturally. As opposed to cyanidin, delphinidin harbors hydroxyl groups at the 3'-, 4'-, and 5'-positions of its B-ring. F3'5'H can add a hydroxyl group at the 3'-, and 5'-position of dihydrokaempferol B-ring to produce dihydromyricetin[8], which is a delphinidin precursor. Therefore, exogenous F3'5'H introduction is the gold standard for breeding blue ornamental flowers. For instance, the F3'5'H gene of pansy (Viola sp. ‘black pansy’) was overexpressed in rose and gerbera, which led to delphinidin glycoside synthesis[1, 9]. Following the specific expression of Campanula medium F3'5'H (CamF3'5'H) gene in tobacco, a large amount of delphinidin was accumulated, leading to a marked change in flower color[10]. Transformation of the CtF3'5'H gene in Verbena changed the flower color from pink to purple[11]. In addition, in a previous study, hairpin RNA (hpRNA) interference (hpRNAi) of chrysanthemum CmF3'H expression and pansy (Viola sp. ‘black pansy’) F3'5'H expression driven by the rose chalcone synthase promoter were employed; the authors found that the transgenic chrysanthemum lines accumulated approximately 80% delphinidin glycosides, and the flower color changed from red-purple to purple-violet[8]. Moreover, the CmF3H promoter linked to the 5′-untranslated region (UTR) sequence of the tobacco NtADH gene as a translation enhancer drove CamF3'5'H overexpression, leading to the production of 95% delphinidin and change in flower color from pink to blue[3]. Glycosylation and acylation of anthocyanins are the most common structural modifications for the production of blue anthocyanins, and these modifications are essential for the stability of color. Mutations in the glycosyltransferase gene AA7GT or acyltransferase gene AA7GT-AT conferred pink or red flower phenotype in Delphinium[12]. To date, two anthocyanin glucosyltransferases have been identified, namely acyl-glucose-dependent anthocyanin glucosyltransferase and UDP-sugar-dependent glucosyltransferases, the former confers the blue flower phenotype in Japanese gentian[13].
Transformation of genes regulating anthocyanin modifications may serve as a strategy to produce blue chrysanthemums. In a previous report, the A3'5'GT gene of Clitoria ternatea (CtA3'5'GT) and CamF3'5'H were co-transferred into chrysanthemum; the transgenic chrysanthemum lines accumulated a large amount of delphinidin 3',5'-diglycosylated derivatives and showed a pure blue flower phenotype[14]. However, the applicability of exogenous F3'5'H genes from other species of Asteraceae in the breeding of blue chrysanthemums remains unknown.
In Asteraceae, Senecio cruentus, Centaurea cyanus L., Callistephus chinensis (L.) nees, and Osteospermum hybrid possess delphinidin metabolic pathways and, consequently, produce bluish flowers[6]. In this study, we transform the OhF3'5'H or mutant CmF3’H (T485S) genes together with CtA3'5'GT driven by the CmF3H promoter respectively for exploring the delphinidin biosynthetic pathway and violet/blue flower phenotype of chrysanthemum.
-
To assess the functions of OhF3'5'H and mutant CmF3'H in the production of bluish chrysanthemum, we introduced CtA3'5'GT encoding a UDP-glucose: anthocyanin 3',5'-glucosyltransferase into the pORE-R4 (R4) vector. Next, we individually introduced the mutant CmF3'H (CmF3'Hm) (Supplemental Fig. S1) and OhF3'5'H genes into the R4-F3HP-CtA3'5'GT vector and then transformed these vectors into chrysanthemum. A total 18 R4-F3HP-CtA3'5'GT-F3HP-CmF3'Hm-positive and 11 R4-F3HP-CtA3'5'GT-F3HP-OhF3'5'H-positive transgenic lines were obtained respectively (Supplemental Fig. S2). Two positive transgenic lines were further selected to determine gene expression levels. Chrysanthemum endogenous CmFE1α was used as the internal control to analyze gene expression in two OhF3'5'H-positive lines (OH9-3 and OH23-8). OH9-3 and OH23-8 successfully expressed CtA3'5'GT and OhF3'5'H (Fig. 2). Cm-29 and Cm-38 transgenic lines also successfully expressed CtA3'5'GT and CmF3'Hm (Fig. 2).
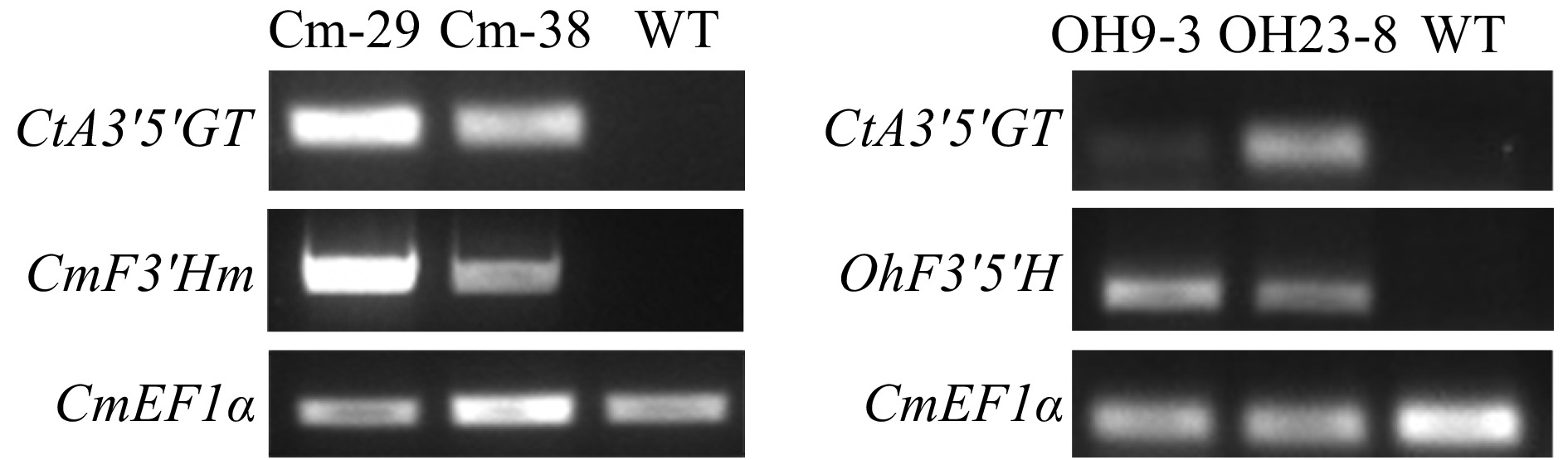
Figure 2. Gene expression analysis in OhF3'5'H and CmF3'Hm transgenic chrysanthemum lines.
The flower color of the transgenic chrysanthemum lines was changed (Fig. 3). According to the Royal Horticultural Society’s (RHS) Colour Chart, the ray florets of wild-type belonged to the red-purple group N74C (deep purplish pink), those of Cm-29 belonged to the purple group 77D (light purple), those of Cm-38 belonged to the purple group N76C (very pale purple), those of OH9-3 belonged to the purple-violet group N82D (light purple), and those of OH23-8 belonged to the violet group 84C (light purple).

Figure 3. Flower color phenotypes of transgenic chrysanthemum lines and comparison based on the RHS Colour Chart. (a) Comparison of flower color phenotypes between the OhF3'5'H transgenic chrysanthemum line and wildtype. (b) Comparison of flower color phenotypes between chrysanthemum CmF3'Hm transgenic chrysanthemum line and wild-type. Flower color of (c) OhF3'5'H and (d) CmF3'Hm transgenic chrysanthemum lines according to the RHS Colour Chart.
CIE L*a*b* values to quantify the colors
-
Changes in the flower color of Cm-29 and Cm-38 were analyzed with a colorimeter. Both lines showed a weaker brightness and lower red and blue chromaticity than the wild-type. Of note, the blue chromaticity of both lines was also lower than that of the wild-type, and the chromaticity angle was greater than 350°, indicating a color closer to red. OH9-3 and OH23-8 showed comparable ray floret color brightness to the wild-type, but the red and blue chromaticity was lower in both lines (Fig. 4).
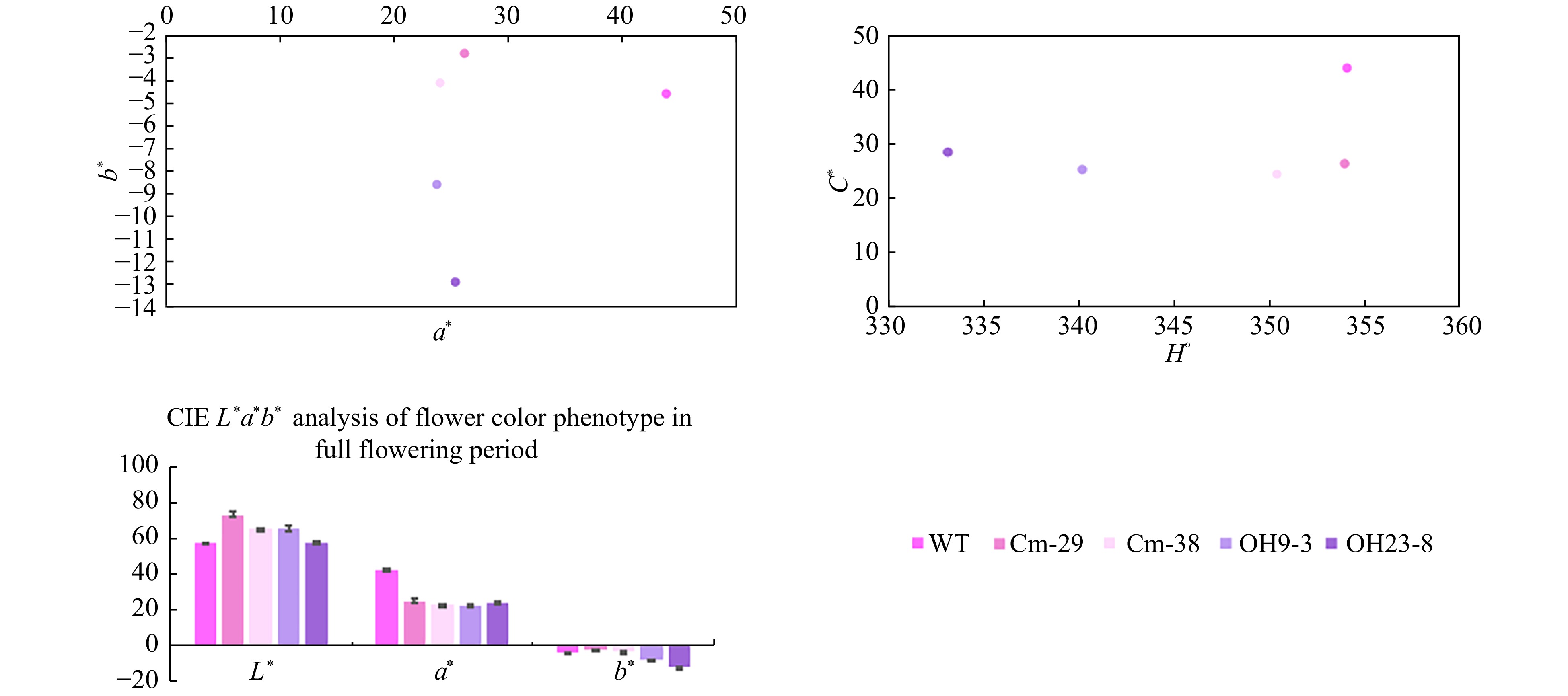
Figure 4. L*a*b* values of ray floret color in transgenic and wild-type chrysanthemum. L* value represents sample brightness; a* value > 0 indicates that the color is reddish; a* value < 0 indicates that the color is more green; b* value indicates the range from blue to yellow, and b* value > 0 indicates that the color is more yellow; b* value < 0 means the color is bluish; H° represents chromaticity angle, and the closer the value of H° is to 270, the more blue the color.
Mass spectrometry to analyze anthocyanin components and structures in the ray florets
-
Qualitative analysis of anthocyanin components in the ray florets of the wild-type and transgenic chrysanthemum lines was performed using high-performance liquid chromatography coupled to electrospray ionization with tandem mass spectrometry (UPLC-ESI-MS/MS) to determine changes in flower color. Consistent with previous reports[14], wild-type chrysanthemum showed two anthocyanin components, namely A1 and A2 (Fig. 5a). Both A1 and A2 exhibited a characteristic molecular ion at m/z 287, corresponding to cyanidin aglycone (Fig. 6). Furthermore, the CmF3'Hm transgenic chrysanthemum lines only showed two anthocyanin components, namely A3 and A4 (Fig. 5b). Both A3 and A4 exhibited a characteristic molecular ion at m/z 287 (Fig. 6). In other words, these two substances are cyanidin derivatives and both contain cyanidin aglycone (Cy), indicating that CmF3'Hm serves no function in conferring violet/blue coloration in chrysanthemum. The weaker brightness and lower red chromaticity of Cm-29 and Cm-38 could be a result of CtA3'5'GT. While, the OhF3'5'H transgenic chrysanthemum lines showed five anthocyanin components, namely A3, A4, A7, A8, and A9 (Fig. 5c, Table 1). A7, A8, and A9 exhibited characteristic molecular ions at m/z 303, corresponding to delphinidin aglycon (Dp), and these components may thus be delphinidin derivatives (Fig. 6). These results indicate that OhF3'5'H functioned well to produce blue color in chrysanthemum.

Figure 5. Mass spectrometric analysis results of anthocyanins of transgenic and wild-type chrysanthemums. (a) Wild-type chrysanthemums with red-purple flowers accumulated A1 and A2. (b) CmF3'Hm transgenic chrysanthemum lines with purple flowers accumulated A3 and A4. (c) OhF3'5'H transgenic chrysanthemum lines with purple-violet flowers accumulated A3, A4, A7, A8 and A9.
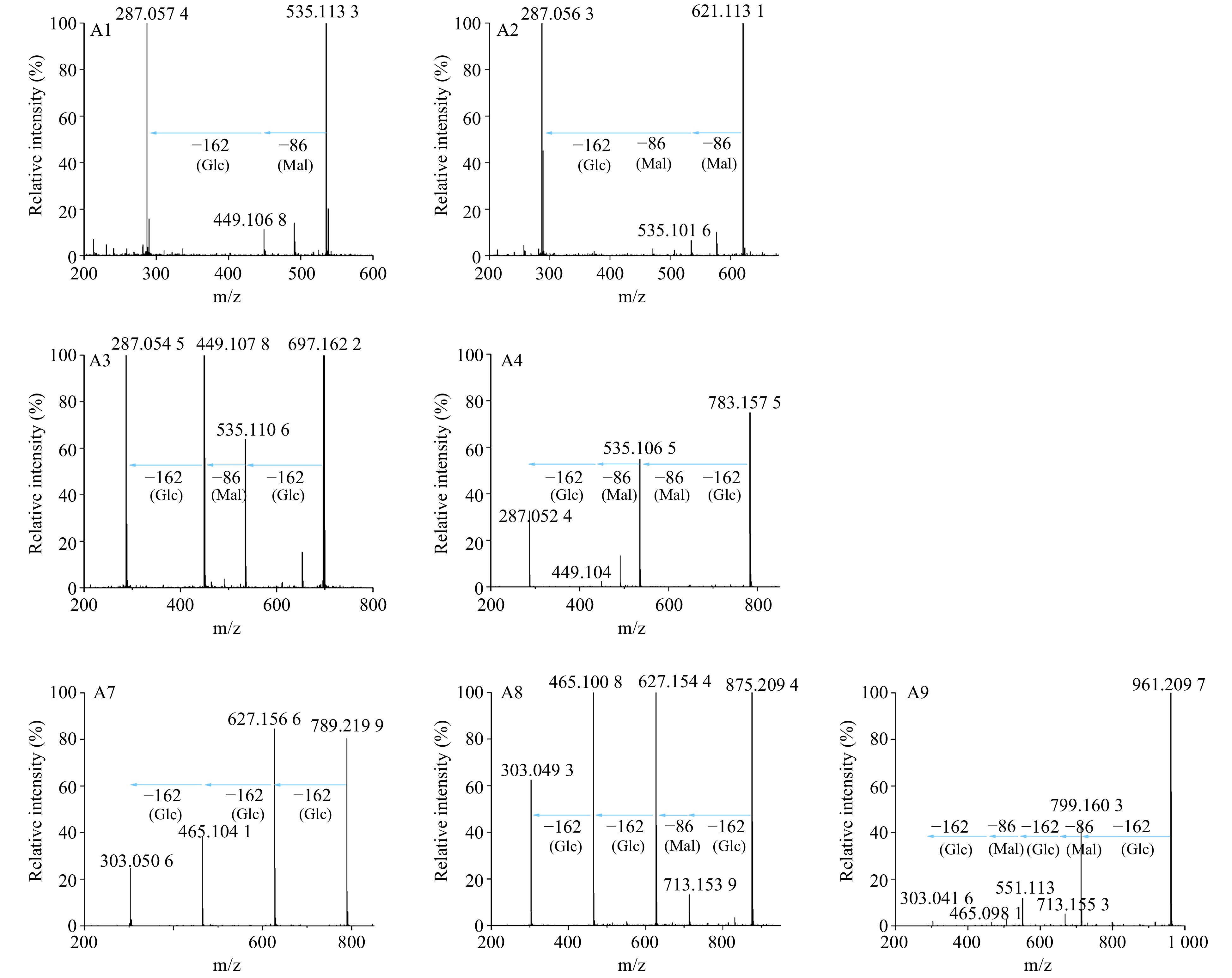
Figure 6. MS/MS analysis of anthocyanins in transgenic and wild-type chrysanthemums. Mal represents malonyl group, Glc represents glucosyl group.The product ions of A1 at m/z = 535[M]+ were 449(−Mal), 287(−Mal, −Glc; Cyanidin aglycone). The product ions of A2 at m/z = 621[M]+ were 535(−Mal), 287(−2×Mal, −Glc). The product ions of A3 at m/z = 697[M]+ were 535(−Glc), 449(−Glc, −Mal), 287( −2×Glc, −Mal). The product ions of A4 at m/z = 783[M]+ were 535(−Glc, −Mal), 449(−Glc, −2×Mal), 287(−2×Mal, −2×Glc). The product ions of A7 at m/z = 789[M]+ were 627(−Glc), 465(−2×Glc), and 303(−3×Glc; delphinidin aglycone). The product ions of A8 at m/z = 875[M]+ were 713(−Glc), 627(−Glc, −Mal), 465(−2×Glc, −Mal), and 303(−3×Glc, −Mal). The product ions of A9 at m/z = 961[M]+ were 799(−Glc), 713(−Glc, −Mal), 551(−2×Glc, −Mal), 465(−2×Glc, −2×Mal), and 303(−3×Glc, −2×Mal).
Table 1. Analysis of anthocyanin mass spectrometry data for transgenic chrysanthemum.
Strain Composition Retention time (min) Molecular ion (m/z) Fragment
ion (m/z)Estimated result WT A1 5.79 535 535, 449, 287 Cyanidin-3-O-(6"-malonyl) glucoside A2 6.86 621 621, 535, 287 Cyanidin-3-O-(3",6"-dimalonyl) glucoside Cm A3 4.30 697 697, 535, 449, 287 Cyanidin-3-O-(6"-malonyl) glucoside-3'-O-glucoside A4 5.06 783 783, 535, 449, 287 Cyanidin-3-O-(3",6"-dimalonyl) glucoside-3'-O-glucoside Oh A3 4.30 697 697, 535, 449, 287 Cyanidin-3-O-(6"-malonyl) glucoside-3'-O-glucoside A4 5.06 783 783, 535, 449, 287 Cyanidin-3-O-(3",6"-dimalonyl) glucoside-3'-O-glucoside A7 1.18 789 789, 627, 465, 303 Delphidin-3,3',5'-tri-O-glucoside A8 2.31 875 875, 713, 627, 465, 303 Delphidin-3-O-(6"-malonyl) glucoside-3',5'-di-O-glucoside A9 2.84 961 961, 799, 713, 551, 465, 303 Delphidin-3-O-(3",6"-dimalonyl) glucoside-3',5'-di-O-glucoside Analysis of ray floret color showed that the CmF3'Hm transgenic lines showed lower red and blue chromaticity than the wild-type. Analysis of anthocyanins in petals revealed two components, namely A3 and A4 (Fig. 5b). Flowers of the OhF3'5'H transgenic line showed lower red chromaticity but higher blue chromaticity than wild-type flowers. Anthocyanin analysis revealed five components, namely A3, A4, A7, A8, and A9, in this line (Fig. 5c). The skeleton of both cyanidin aglycone and delphinidin contains a benzo-oxygen-containing heterocyclic structure connected to an aromatic structure. This structure shows π–π conjugation to form a π1617 delocalized large π bond. In this structure, the more the glucosides connected to the delocalized π-bond structure, the stronger the molecular polarity. The accumulation of three glucosyl groups makes A7 more polar, and it was the first to be separated in reversed-phase chromatography. However, A8 with one Mal-group and A9 with two Mal-groups showed reduced molecular polarity, and separation peaks appeared at 2.31 and 2.84 min, respectively. Since A7, A8, and A9 exhibited the characteristic ions at m/z 303, corresponding to delphinidin aglycon (Dp), A7 is delphinidin-3,3',5'-tri-O-glucoside, A8 is delphinidin-3-O-(6"-malonyl) glucoside-3',5'-di-O-glucoside, and A9 is delphinidin-3-O-(3",6"-dimalonyl) glucoside-3',5'-di-O-glucoside (Table 1). In addition, the CmF3'Hm and OhF3'5'H transgenic chrysanthemum lines both contained A3 and A4. Secondary mass spectra showed that A3 contains two Glc-groups and one Mal-group and a characteristic ion at m/z 287, corresponding to cyanidin aglycone (Cy); thus, A3 is cyanidin-3-O-(6"-malonyl) glucoside-3'-O-glucoside. A4 contained two Mal-groups, and its peak appeared before the peaks of the wild-type chrysanthemum anthocyanins A1 and A2. Thus, A4 is cyanidin-3-O-(3",6"-dimalonyl) glucoside-3'-O-glucoside. Based on our MS data and previous reports, A1 is cyanidin-3-O-(6"-malonyl) glucoside and A2 is cyanidin-3-O-(3",6"-dimalonyl) glucoside (Table 1).
-
In recent years, the introduction of exogenous F3'5'H has become an efficient strategy for the breeding of ornamental plants with blue flowers. The transformation of CamF3'5'H can efficiently alter the original metabolic pathways driving flower color in plants. For instance, the content of delphinidin derivatives in CamF3'5'H transgenic tobacco accounted for 99% of total anthocyanins. Moreover, CamF3'5'H and CtA3'5'GT were co-transformed into chrysanthemum and blue flowers were obtained[10]. In the present study, we explored the effects of transforming mutant CmF3'Hm and exogenous OhF3'5'H on anthocyanin metabolism in chrysanthemum. Simultaneously, we introduced CtA3'5'GT to modify the anthocyanidin skeleton, hoping to obtain endogenous 3',5'-diglucosylated delphinidin through the intermolecular interaction of flavonoid glycosides with flavonoids.
According to a previous phylogenetic study, F3'5'H of Pericallis, Osteospermum, and Callistephus may have evolved from F3'H of other Asteraceae plants[6]. Moreover, threonine and serine of Gerbera hybrida and Osteospermum hybrid were interconvertible, and this change supported the F3'H of Gerbera hybrida with a little F3'5'H activity and the F3'5'H of Osteospermum hybrid with a high F3'H activity[7]. These results highlight great opportunities for using the endogenous F3'H of mutant chrysanthemum to alter its anthocyanin metabolism. However, the mutant CmF3'Hm did not lead to beneficial changes in delphinidin metabolism following co-transformation with CtA3'5'GT (Fig. 6). The weaker brightness and lower red chromaticity of Cm-29 and Cm-38 were due to CtA3'5'GT function, which is consistent with a previous report that the flowers of CtA3'5'GT transgenic chrysanthemum did not show a bluish hue and only produced A3 and/or A4 anthocyanin components[14]. Therefore, chrysanthemum CmF3'Hm obtained by site-directed mutagenesis does not possess F3'5'H activity and cannot be used for the breeding of plants with blue flowers. These findings indicate that other key amino acids of CmF3'H may be involved in F3'5'H activity, which warrant further research in the context of blue chrysanthemum breeding.
In OhF3'5'H and CtA3'5'GT transgenic chrysanthemum lines, the precursor components A1 and A2 can be catalyzed by OhF3'5'H to produce A5 and A6. Further, A5 and A6 can be modified via the addition of a glucosyl group by UDP-glucose: dependent anthocyanin 3′,5′-O-glucosyltransferase to generate A7, A8 and A9 (Fig. 7). Therefore, although the co-transformation of OhF3'5'H and CtA3'5'GT led to the accumulation of delphinidin derivatives (Fig. 6, Table 1), the bluish hue was not as good as that obtained previously through the transformation of CamF3'5'H and CtA3'5'GT[14]. This may be because the host variety used was different. Accordingly, additional transgenic lines would be beneficial for selection. In conclusion, OhF3'5'H effectively promoted the transition of chrysanthemum flower color to violet/blue and has potential applicability in violet/blue chrysanthemum breeding.
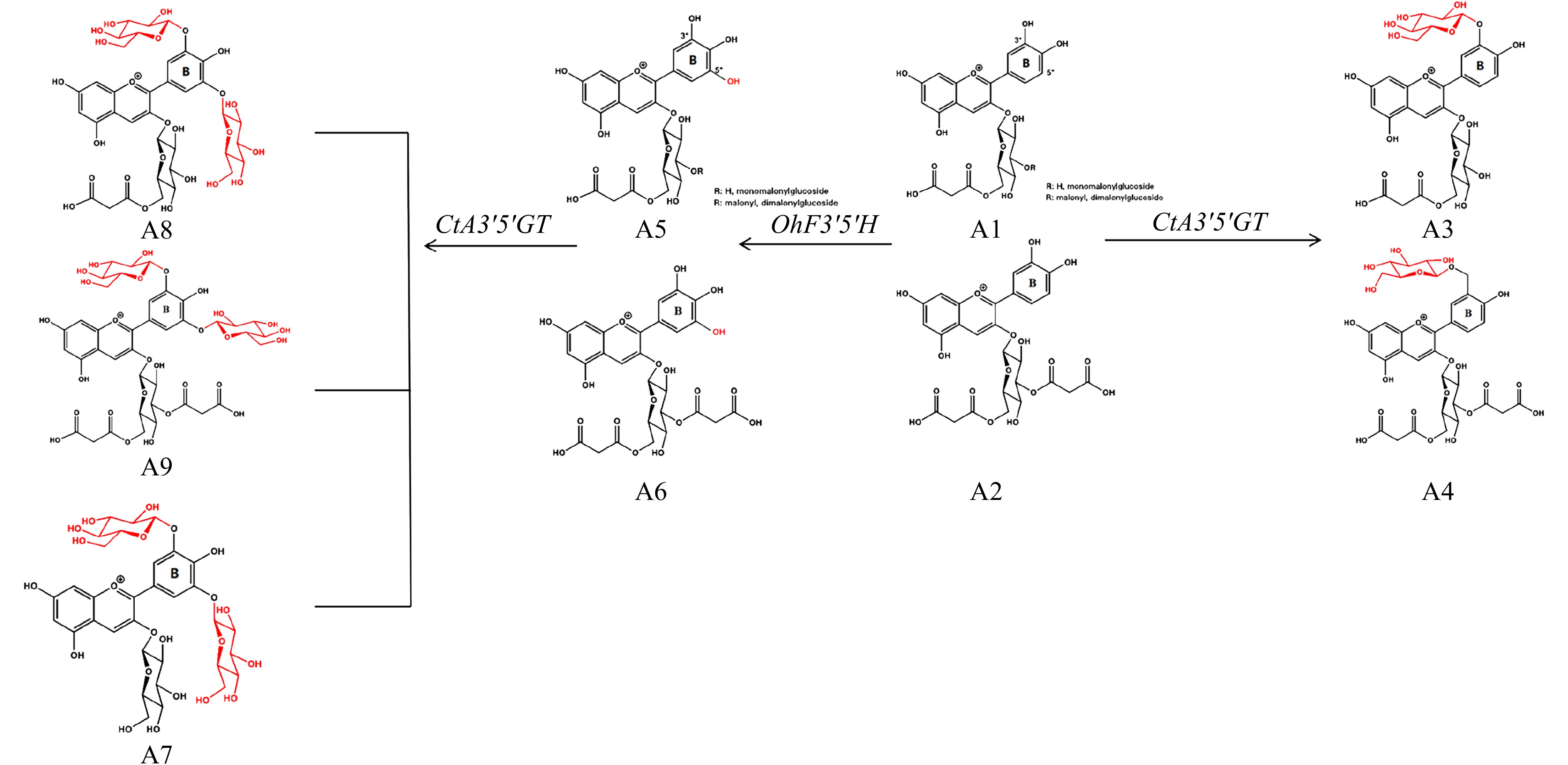
Figure 7. Metabolic modifications of endogenous chrysanthemum pigments by OhF3'5'H and CtA3'5'GT.
-
The chrysanthemum cultivar ‘Nannong Fencui’ with pink flowers was cultivated by the Chrysanthemum Laboratory at the Nanjing Agricultural University. Agrobacterium tumefaciens strain EHA105 containing a binary vector was used for agrobacterium-mediated transformation. Chrysanthemum leaves were used for stable gene transformation. The leaf tissues were cultured on a pre-culture medium (MS + 1.0 mg L−1 6-BA + 0.5 mg L−1 NAA) for 2−3 days. Following culture, the leaves were immersed in freshly prepared agrobacterium broth for 8−10 min, inoculated on a co-culture medium (MS + 1 mg L−1 6-BA + 0.5 mg L−1 NAA), and incubated in the dark at 28 °C for 3 days. Thereafter, the leaves were transferred to a decarboxylation medium (MS + 1 mg L−1 6-BA + 0.5 mg L−1 NAA + 500 mg L−1 Carb) and cultured for 5−7 days. Subsequently, the leaves were transferred to a selection medium (MS + 1 mg L−1 6-BA + 0.5 mg L−1 NAA + 300 mg L−1 Carb + 50 mg L−1 Kan) for 3−4 generations of subculture selection. When resistant buds to be differentiated grew to 2−3 cm in length, they were transferred to a rooting selection medium (MS + 30 mg L−1 Kan), and completely resistant plants were obtained. Following infection and transformation, the regenerated kanamycin-resistant tissues gradually grew into adult transgenic plants after four rounds of resistance screening. The transgenic plants were transferred to soil and cultivated in the greenhouse supplying either a 16-h or an 8-h photoperiod; in each case, the temperature was maintained at a constant 23 °C. Flower color was compared between the transgenic and wild-type plants.
Construction of transformation vectors
-
To the promoter sequence of CmF3H in NCBI (GenBank: FW570860.1), the 5′-UTR sequence of NtADH and a 3′-multiple cloning site (MCS) were added, and the construct was introduced in the intermediate vectors pYL322d2 (GenBank: KY420077.1) and pYL322d1 (GenBank: KY420076.1). The Xho I restriction site was introduced upstream, and the Pst I restriction site was introduced downstream. Plasmid construction was entrusted to General Bio company (Chuzhou, Anhui province, China), and the constructed plasmids pYL322d2 and pYL322d1 with the synthesized fragments were double digested with Xho I and Pst I. A gel recovery kit (AXYGEN) was used to recover the pYL322d2 vector and target F3'H promoter fragments. T4 DNA ligase (Fermentas) was used to ligate the two recovered products (10 μL reaction system: 1 μL of 10 × T4 DNA ligation buffer, 6 μL of F3'H promoter fragment, 2 μL of pYL322d2 vector fragment/pYL322d1 vector fragment, and 1 μL of T4 DNA ligase; ligated at 22 °C for 1 h), and DH5α competent cells were transformed with 5 µL of the ligation product to obtain d2F3HP- and d1F3HP-positive colonies. The Kpn I restriction site was introduced upstream, and the Sma I restriction site was introduced downstream of CtA3'5'GT fused with 35S PolyA (GenBank: AB115560.1). Plasmid construction was carried out by General Bio company (Chuzhou, Anhui province, China), and the construct was inserted between the Kpn I and Sma I sites of the d2F3HP vector; the vector thus obtained was labeled d2F3HP-CtA3'5'GT.
To the OhF3'5'H sequence in NCBI (GenBank: DQ257627.1), the Nos terminator sequence was added and Pst I and Spe I restriction sites were introduced upstream and downstream, respectively. Plasmid construction was carried out by General Bio company (Chuzhou, Anhui province, China) and the obtained construct was inserted between the Pst I and Spe I restriction sites of d1F3HP vector; the vector thus obtained was labeled d1F3HP-OhF3'5'H.
The d2F3HP-CtA3'5'GT vector was digested with Not I and Sma I. Simultaneously, the pORE-R4 empty vector (GenBank: AY562547.1) was digested, and the recovered F3HP-CtA3'5'GT fragment was ligated to this vector (GenBank: AY562547.1); the vector thus obtained was labeled R4-CtA3'5'GT. An upstream primer (ggactagtttacaaaaccatgtgcaagaat) was used for introducing the Spe I restriction site, and a downstream primer (aagaatgcggccgcgatctagtaacatagatgacaccgcgc) was used for introducing the Not I restriction site. d1F3HP-OhF3'5'H plasmid DNA was used as the template for PCR amplification. The reaction system (50 μL) contained 5.0 μL of 10 × PCR buffer, 1.0 μL each of upstream and downstream primers (10 μmol L−1), 5.0 μL (2 mmol L−1) of dNTP mix, 0.5 μL of Pfusion DNA polymerase, 2 μL of plasmid DNA template, and 35.5 μL of ddH2O; the reaction conditions were as follows: pre-denaturation at 95 °C for 2 min; 24 cycles of melting at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C 40 s; and final extension at 72 °C for 7 min. Following PCR, the products were subjected to 0.8% agarose gel electrophoresis for confirmation. Using the above restriction digestion and ligation system, the PCR product was ligated between the Not I and Spe I restrictions sites of the R4-CtA3'5'GT vector. After transforming into DH5α competent cells, the positive expression vector plasmid R4-OhF3'5'H-CtA3'5GT was obtained.
According to the sequence of chrysanthemum CmF3'H gene in NCBI (GenBank: AB523844.1), we site-specifically mutated the 1,534th base C in the cDNA sequence of CmF3’H to G, which is corresponding with a conservative Thr to Ser exchange at position 487 of recombinant G. hybrida F3'H conferred additional 5'-hydroxylation activity[7], and added the Nos terminator to this sequence entrusted to General Bio company. The Kpn I restriction site was introduced upstream, and the Sal I restriction site was introduced downstream. Using the above mentioned restriction digestion and ligation system, the product was ligated between the Kpn I and Sal I restriction sites of the d1F3HP vector; the vector thus obtained was labeled d1F3HP-CmF3'Hm. Then, we digested the above mentioned d2F3HP-CtA3'5'GT vector with Not I and Sma I and introduced the recovered F3HP-CtA3'5'GT fragment in the pORE-R4 vector; the vector thus obtained was labeled R4-CtA3'5'GT. An upstream primer (ccatcgatttacaaaaccatgtgcaagaat) were used for introducing the Not I restriction site, and a downstream primer (aagaatgcggccgcgatctagtaacatagatgacaccgcgc) was used for introducing the Cla I restriction site. The d1F3HP-CmF3'Hm plasmid DNA was as used as the template for PCR amplification using the above mentioned restriction digestion and ligation system. The PCR product was ligated between the Not I and Cla I restriction sites of the R4-CtA3'5'GT vector. After transforming into DH5α competent cells, the positive expression vector plasmid R4-CmF3'Hm-CtA3'5GT was obtained.
Total RNA extraction and cDNA synthesis
-
Total RNA was extracted from the petals of frozen transgenic chrysanthemum plants. A commercial kit from Hua Yueyang (Beijing, China) was used for total RNA extraction using DNase I to digest residual DNA, and plant RNA was obtained after elution. cDNA was synthesized from the extracted RNA (1,000 ng) using Prime ScriptTM RT-PCR Reverse Transcription Kit (TaKaRa).
RT-PCR
-
cDNA of R4-OhF3'5'H-CtA3'5'GT and R4-CmF3’Hm-CtA3'5'GT transgenic chrysanthemum lines as well as wild-type chrysanthemum was used as the template for PCR with the r-Taq PCR kit (TaKaRa); chrysanthemum CmEF1α was used as the internal control[15]. The following primer pairs were used: qCtA3'5'GT-F–qCtA3'5'GT-R and qOhF3'5'H-F–qOhF3'5'H-R for R4-OhF3'5'H-CtA3'5'GT transgenic lines and qCtA3'5'GT-F–qCtA3'5'GT-R and qCmF3’Hm-F–qCmF3’Hm-R (Table 2) for R4-CmF3’Hm-CtA3'5'GT transgenic lines. Following PCR, the products were against 0.8% agarose gel electrophoresis. Based on the brightness of amplification bands, differential expression of genes was analyzed between the transgenic and wild-type plants.
Table 2. Gene primers used to determine gene expression level in transgenic plants.
Primer Sequence (5'-3') qCtA3'5'GT-F GTCTTCAAACTCGCTCACGC qCtA3'5'GT-R CGGGCACATAACCCTCTGG qOhF3'5'H-F GGCTCCACCCATCAACACCA qOhF3'5'H-R TACCCGCACAAATCCTCCGC qCmF3’Hm-F GGCTCCACCCATCAACACCA qCmF3’Hm-R TACCCGCACAAATCCTCCGC CmEF1α-F TTTTGGTATCTGGTCCTGGAG CmEF1α-R CCATTCAAGCGACAGACTCA Color analysis
-
We selected ray florets from three different samples of each line as the test material. The RHS Colour Chart was used to compare floret colors of distinct phenotypes. A colorimeter (Konica Minolta/CR-10 Plus) was used to determine the L*a*b* values of ray florets. Measurement on each sample was repeated three times. The L* value represents the brightness of the color, the a* value represents the color spectrum from red to green, and the b* value represents the color spectrum from yellow to blue. Specifically, a* greater than zero indicates red, a* less than zero indicates green, b* greater than zero indicates yellow, and b* less than zero indicates blue. The hue angle (H° = arctan b*/a*) indicates color change. Specifically, H° = 0 or 360 indicates red, H° = 90 indicates yellow, H° = 180 indicates green, and H° = 270 indicates blue.
Sample preparation for and analysis of flavonoid pigments
-
Fresh chrysanthemum ray florets (0.25 g) were ground to a powder in liquid nitrogen. Next, 1 mL of solvent (methanol : water : formic acid : trifluoroacetic acid = 70:27:2:1) was added to the powder. The mixture was shaken well and stored in a refrigerator at 4 °C for 24 h in the dark. The mixture was centrifuged at 13,000 rpm for 10 min, and the supernatant was filtered through a 0.22 μm microporous membrane for further analysis. The anthocyanin extract was subjected to high-performance liquid chromatography (UPLC) coupled with mass spectrometry (G2-XS QTof, Waters) using the ACQUITY UPLC system and ACQUITY TQD tandem mass spectrometer, equipped with the ACQUITY UPLC BEH C18 (1.7 mm particle size; 2.1 × 100 mm) column. The mobile phase A was an aqueous solution containing 0.1% formic acid, and the mobile phase B was an acetonitrile solution containing 0.1% formic acid. The elution gradient was 5% B, 0.5 min; 5%−40% B, 20 min; 40%−95% B, 2 min; and 95% B, 2 min. The injection volume was 2 μL, and the flow rate was 0.35 mL·min−1. The ESI source in MS analysis was operated in the positive ion, MSe, and MS/MS acquisition modes, at a full mass scanning range of 50-1,200 m/z. Recalibration with Leucine–Enkephalin (m/z 556.2771) was performed for quality check. The ionization parameters were as follows: capillary voltage, 2.5 kV; sampling cone voltage, 40 V; source temperature, 120 °C; and desolventizing gas temperature, 400 °C. Mass Lynx 4.1 and Unifi were used for MS data acquisition and processing[14].
- This work was supported by the National Natural Science Foundation of China (31930100, 31902053), the earmarked fund for Jiangsu Agricultural Industry Technology System (JATS [2020] 402), a Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Comparison of mutant CmF3'Hm, chrysanthemum endogenous CmF3'H , and F3'5'H in Asteraceae OhF3'5'H , Osteospermum hybrid flavonoid 3'5'-hydroxylase gene (GenBank: DQ257627.1); PcF3'5'H , Pericallis cruenta flavonoid 3'5'-hydroxylase gene (GenBank: DQ257626.1); CmF3’Hm, mutant Chrysanthemum morifolium flavonoid 3'-hydroxylase gene; CmF3'H ,Chrysanthemum morifolium flavonoid 3'-hydroxylase gene (GenBank: FW570860.1).
- Supplemental Fig. S2 PCR results for transgenic chrysanthemum (A) OhF3'5'H and (B) CmF3'Hm levels in transgenic chrysanthemum.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Han X, Luo Y, Lin J, Wu H, Sun H, et al. 2021. Generation of purple-violet chrysanthemums via anthocyanin B-ring hydroxylation and glucosylation introduced from Osteospermum hybrid F3 ' 5 ' H and Clitoria ternatea A3 ' 5 ' GT . Ornamental Plant Research 1: 4 doi: 10.48130/OPR-2021-0004

Generation of purple-violet chrysanthemums via anthocyanin B-ring hydroxylation and glucosylation introduced from Osteospermum hybrid F3'5'H and Clitoria ternatea A3'5'GT
- Received: 26 February 2021
- Accepted: 06 May 2021
- Published online: 19 May 2021
Abstract: Chrysanthemums possess no metabolic pathway to synthesize delphinidin because of the lack of endogenous F3'5'H gene encoding the key enzyme in its biosynthetic pathway; therefore, there are no blue or blue-purple chrysanthemums occurring naturally. Currently, the introduction of exogenous F3'5'H into chrysanthemums is an efficient method for breeding bluish chrysanthemums. In this study, we explored the effects of the introduction of mutant CmF3'H (generated via site-directed mutagenesis, T485S, CmF3'Hm) and exogenous Osteospermum hybrid F3'5'H (OhF3'5'H) genes combined with Clitoria ternatea A3'5'GT (CtA3'5'GT) on delphinidin synthesis in chrysanthemum. Among the F3'5'H transgenic lines, those overexpressing endogenous CmF3'Hm could not generate blue flower color, although red color was changed to light pink due to CtA3'5'GT function. Meanwhile, OhF3'5'H introduction promoted the accumulation of delphinidin and its derivatives in chrysanthemum, changing the flower color from red-purple to purple-violet. These results indicate the applicability of exogenous OhF3'5'H and CtA3'5'GT transformation for promoting delphinidin synthesis during the molecular breeding of violet/blue chrysanthemums.
-
Key words:
- OhF3'5'H /
- CmF3'H /
- delphinidin /
- blue chrysanthemum
{kind=link}
{kind=link}