-
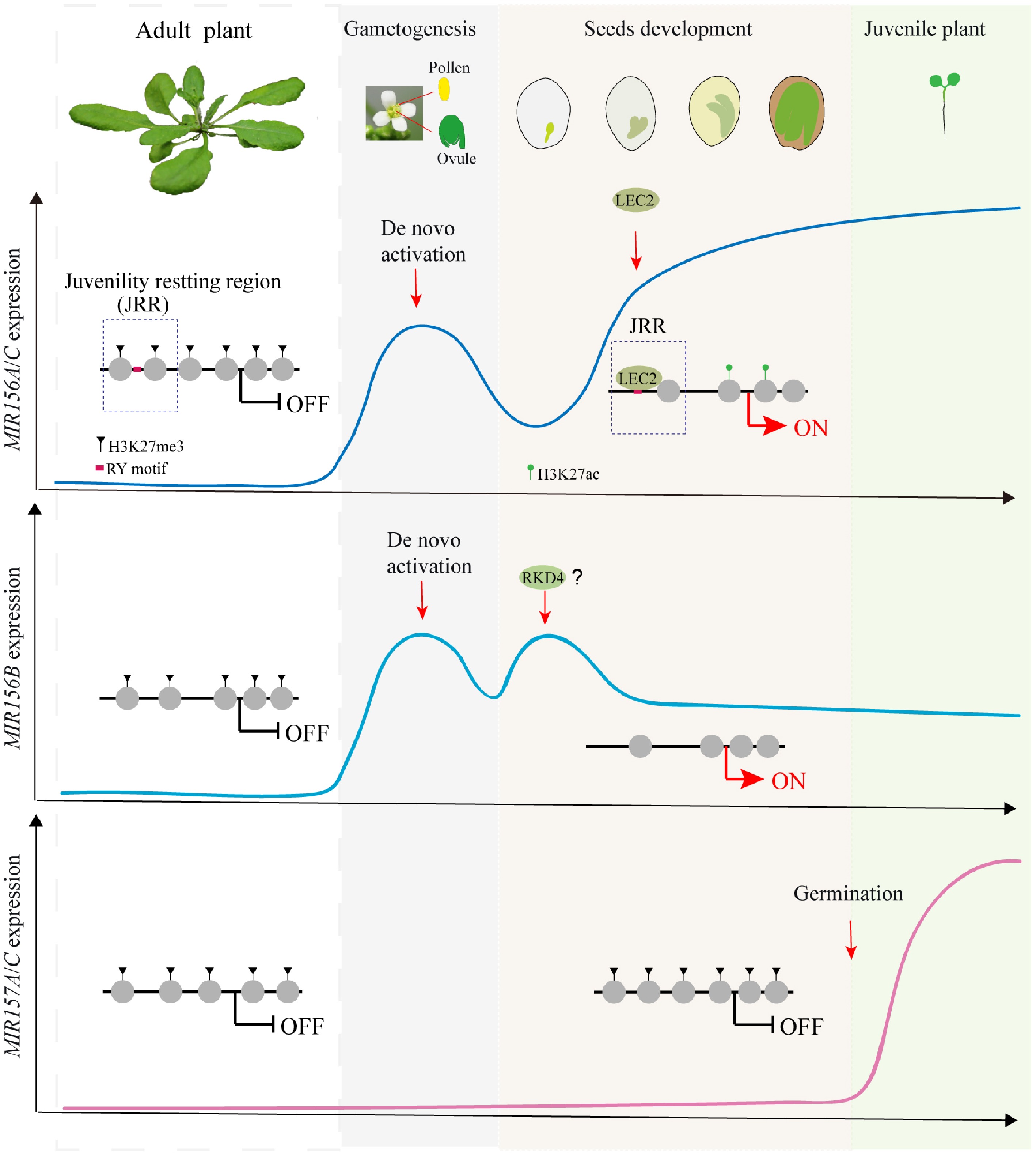
Figure 1.
Resetting of juvenility via reactivation of MIR156/157 during seed development in Arabidopsis. MIR156A–C and MIR157A/C play dominant roles in phase transition, but their reactivation occurs through distinct reprogramming routes. MIR156A–C are de novo activated in gametogenesis, with MIR156A/C subsequently reset from Polycomb repression to active chromatin by LEC2 through its binding to the RY motif in the juvenility resetting region during embryogenesis, whereas MIR156B, maintaining a constitutively open regulatory region throughout the life cycle, is reactivated earlier by other embryonic factors. MIR157A/C, constitutively marked by H3K27me3 and maintained in a closed chromatin state throughout embryogenesis, are reactivated upon germination.
-
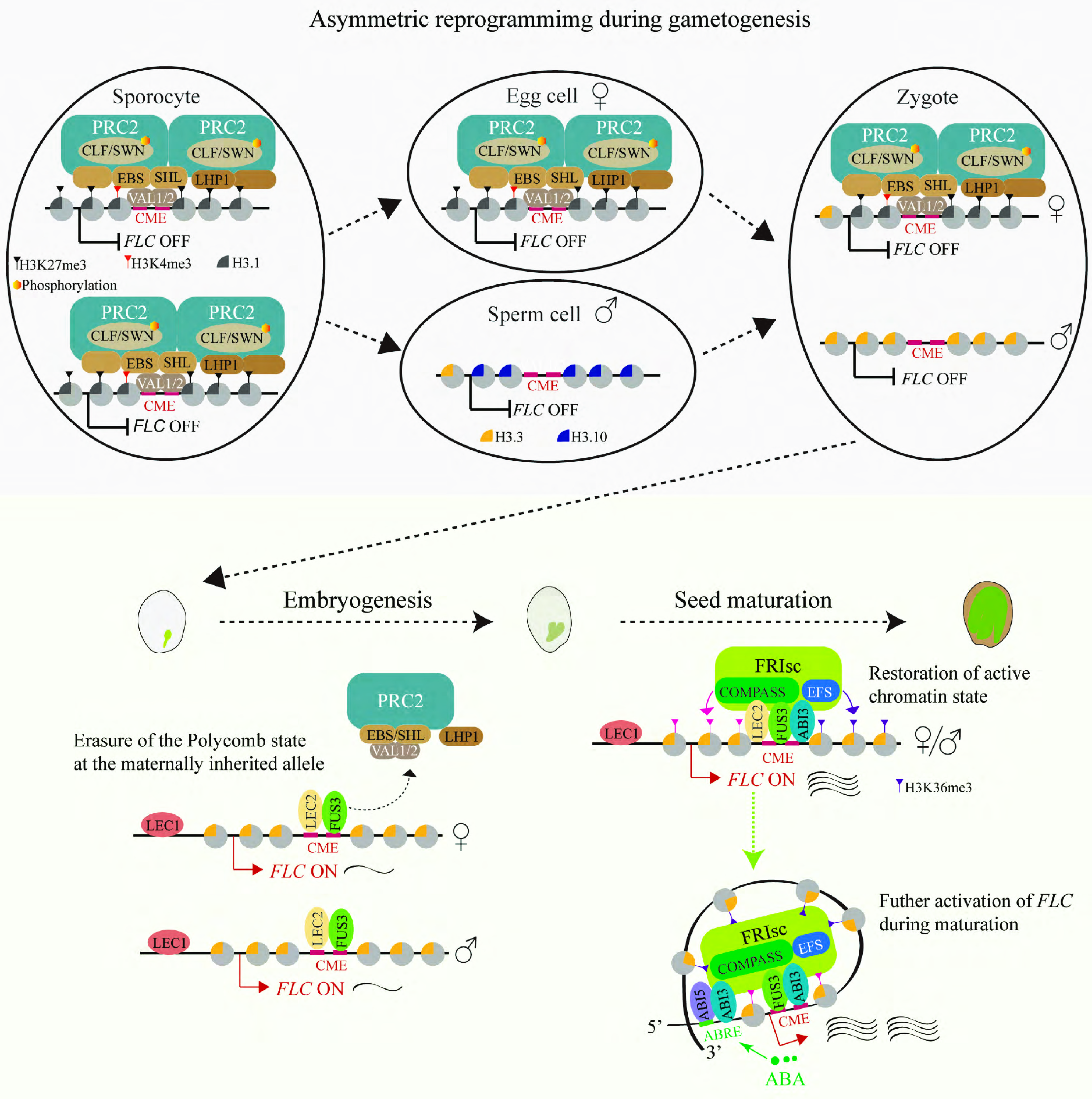
Figure 2.
Reprogramming of the vernalized state in Arabidopsis. During male gametogenesis, genome-wide chromatin reprogramming in sperm cells—mediated by active H3K27 demethylation and absence of core PRC2—depletes the vernalized state specifically at the paternal FLC allele. Histone variant H3.10 incorporation occurs in parallel but does not directly mediate H3K27me3 erasure at FLC. In contrast, the Polycomb-repressive vernalized state of FLC chromatin is transmitted to the zygote via the egg cell. After fertilization, LEC1 opens FLC chromatin, enabling B3-domain factors (LEC2, FUS3, and ABI3) to displace Polycomb-group factors. Displacement of the silencing machinery leads to progressive dilution of maternal H3K27me3 through rapid cell divisions. Concurrently, these factors recruit FRIsc, which deposits H3K4me3 and H3K36me3 to reset FLC to an active, mitotically stable state by the end of embryogenesis. ABA reinforces FLC activation in maturing seeds via the ABI5–ABI3 heterodimer, enhancing FRIsc recruitment and driving FLC upregulation. This active state is mitotically maintained in vegetative tissues, restoring the vernalization requirement in progeny.
-
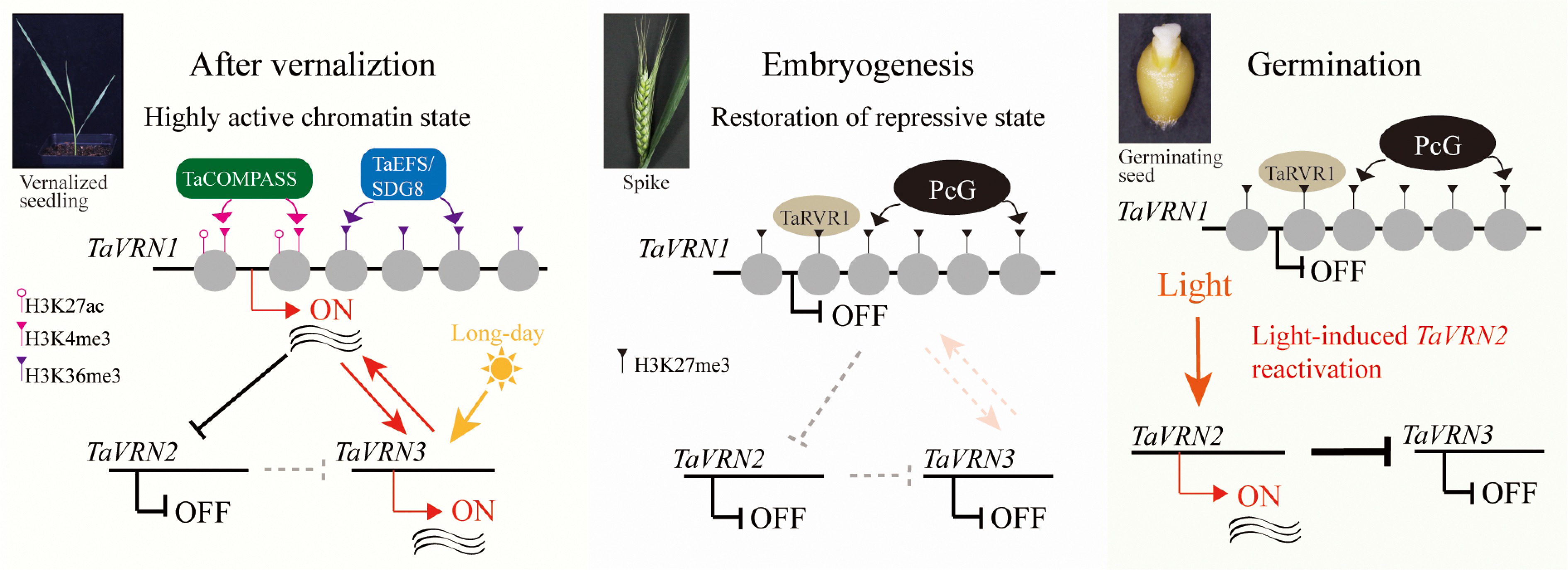
Figure 3.
Two-step restoration of over-wintering growth habit in winter wheat. Prolonged cold erases PRC2-mediated H3K27me3 at TaVRN1, enabling COMPASS-like and EFS to deposit H3K4me3 and H3K36me3, respectively, switching TaVRN1 to an active state. During embryogenesis, active marks are removed and H3K27me3 is re-deposited by Polycomb group (PcG) proteins, reinstating the Polycomb-repressed state partly via TaRVR1. Concurrently, TaVRN2 is light-reactivated during seed germination. Thus, cold-induced activation and embryonic resetting form a reversible chromatin switch that restores vernalization requirement in each generation.
Figures
(3)
Tables
(0)