









-
How do plants erase the memory of parental development and environmental experience to ensure that each new generation begins life with a fresh start? This fundamental question lies at the heart of epigenetic reprogramming. Epigenetic states—mediated by chromatin modifications, histone variants, remodeling, DNA methylation, and non-coding RNAs—serve as heritable yet reversible information carriers that precisely regulate gene expression in plants and animals[1−4]. Among these regulatory mechanisms, histone modifications play a central role by recruiting effector proteins that modulate transcriptional activity[5,6]. Activating marks such as H3/H4 acetylation (H3ac/H4ac) and trimethylation of histone H3 at lysine 4 and lysine 36 (H3K4me3 and H3K36me3) promote active transcription, with H3K4me3 specifically facilitating RNA polymerase II promoter-proximal pause-release and elongation beyond simply creating open chromatin accessibility[7,8]. In contrast, repressive marks such as Polycomb-mediated H3K27me3 contribute to chromatin compaction and gene silencing[9]. Notably, epigenetic states are highly dynamic throughout the plant life cycle[10]. This plasticity, mediated by dedicated enzymatic machinery—writers, readers, and erasers—underlies both cell fate changes and the establishment of epigenetic memory in response to developmental and environmental cues[11]. However, to ensure normal offspring development, such acquired states often require precise reprogramming in the germline, a process that faces a unique logistical challenge in plants[12,13]. Unlike in mammals, where the germline is sequestered early, the plant germline is specified late, derived from adult somatic cells only after major post-embryonic phase transitions, including seedling establishment, vegetative growth and development, and floral transition[10,14]. Consequently, the germline progenitor cells carry a legacy of somatic epigenetic commitments, necessitating extensive resetting during gametogenesis and early seed development to clear parental marks and restore totipotency[15−17].
Therefore, the faithful execution of the plant life cycle hinges on resetting multiple, interdependent, epigenetic programs. This imperative is illustrated by two paradigmatic needs: (1) resetting intrinsic developmental programs, such as the juvenile-to-adult phase transition governed by regulators like the miR156/miR172 module[18−21], which must be reversed each generation to ensure offspring restart from a default juvenile state; (2) erasing acquired environmental epigenetic 'memories', such as vernalized state, prolonged cold-induced stable chromatin changes such as silencing of FLOWERING LOCUS C (FLC) in Arabidopsis, or activation of VERNALIZATION1 (VRN1) in temperate cereals, which establishes a somatic 'memory' that must be cleared in offspring to restore developmental potential[22,23].
The collective need to reset both intrinsic developmental programs and acquired environmental 'memories' converges on a singular developmental window: seed development. Importantly, we distinguish two distinct phases of reprogramming: (1) gametophytic reprogramming, occurring in the haploid male and female gametes before fertilization, which prepares the parental genomes for the next generation; and (2) embryonic reprogramming, occurring after fertilization during diploid embryo development and seed maturation, which constitutes the core of what we term 'seed-based' resetting. Throughout this developmental continuum, genome-wide epigenetic reprogramming unfolds to set the stage for the locus-specific resetting of parental memories[24−26]. Thus, the seed serves as the indispensable hub where parental epigenetic information is comprehensively reprogrammed to establish the foundational totipotency of the offspring[17,27,28].
-
The juvenile-to-adult transition, known as vegetative phase change, is a critical developmental switch in plants[29]. In Arabidopsis, this phase change is marked by distinct morphological traits, including the initiation of abaxial trichomes, increased leaf length-to-width ratio and serration, and reduced cell size[20]. At the molecular level, the transition is orchestrated by a conserved microRNA pathway[30]. A decline in miR156/157 levels leads to the upregulation of their targets, the SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) transcription factors[18,19,31]. These SPLs act together with miR172 that represses AP2-like transcription factors to precisely time the phase change to the adult stage[32,33]. In Arabidopsis, miR156 is produced mainly from the MIR156A and MIR156C loci within an eight-member gene family (MIR156A–H), whereas miR157 is encoded by four genes (MIR157A–D) that function redundantly, but each with a weaker effect[20,31]. miR156/157 together regulate 10 of the 16 SPL genes, which are grouped into clades based on their DNA-binding domains. Among these, SPL9, SPL13, and SPL15 play particularly prominent roles in promoting the adult phase[21,34]. As the master regulator of the transition, the miR156/157-SPL pathway is itself tightly controlled at multiple levels. Beyond transcriptional regulation, epigenetic mechanisms are crucial for the timing of vegetative phase change[20,21]. Key MIR156 loci, especially MIR156A/C, are progressively silenced via H3K27me3 deposition by Polycomb-repressive complex 2 (PRC2)[21,35]. PRC1 reinforces this by maintaining H3K27me3 and catalyzing histone H2A monoubiquitination (H2Aub) at their promoters[36], while the active mark H3K4me3, deposited by ARABIDOPSIS TRITHORAX-RELATED PROTEIN 7 (ATXR7), declines[37]. Chromatin remodelers such as BRAHMA and PICKLE, along with the histone variant H2A.Z, further fine-tune their expression by modulating nucleosome positioning[21,38,39]. In parallel, SPL genes themselves are direct targets regulated by epigenetic modifications. They are repressed by PRC1-mediated H2Aub and activated by HISTONE ACETYLTRANSFERASE OF THE GNAT FAMILY 1 (HAG1)-mediated histone acetylation[40,41]. This dynamic chromatin landscape ensures precise developmental timing in response to endogenous and environmental signals, with established epigenetic states remaining stable until reset in the next generation.
The unique ontogeny of the plant germline, wherein gametes are formed from differentiated somatic cells of adult tissues, confers fundamental importance on epigenetic resetting[12]. In the absence of such an active mechanism, epigenetic information of parental age and maturation state would be heritable, resulting in a progressive decline in developmental plasticity across generations[17]. Research in Arabidopsis thaliana has elucidated a robust, multi-layered mechanism that ensures the reliable restoration of juvenility. This mechanism is centered on the tiered, de novo transcriptional reactivation of specific MIR156/157 gene family members, leading to a sophisticated, redundant, multi-pathway cascade (Fig. 1). The functional necessity of the miR156/157-driven resetting was revealed through the creation of high-order CRISPR/Cas9 mutants[28]. To overcome the limitations of partial T-DNA insertions[31], null alleles for all eight MIR156 genes (mir156oct) and all twelve MIR156/157 genes (mir156/157duodec) were generated, with small RNA sequencing to confirm their complete knockouts. These mutants exhibited dramatically precocious phenotypes. Most strikingly, the most severe mir156/157duodec mutant bolted without forming rosette leaves under long-day conditions[28]. Concomitant analyses confirmed the precocious activation of flowering pathway genes in shoot apices and even in embryos, where chromatin accessibility at flowering gene loci was significantly increased, providing direct molecular genetic proof—via complete loss-of-function mutants—that the reset is essential to suppress the default reproductive program[28,33].
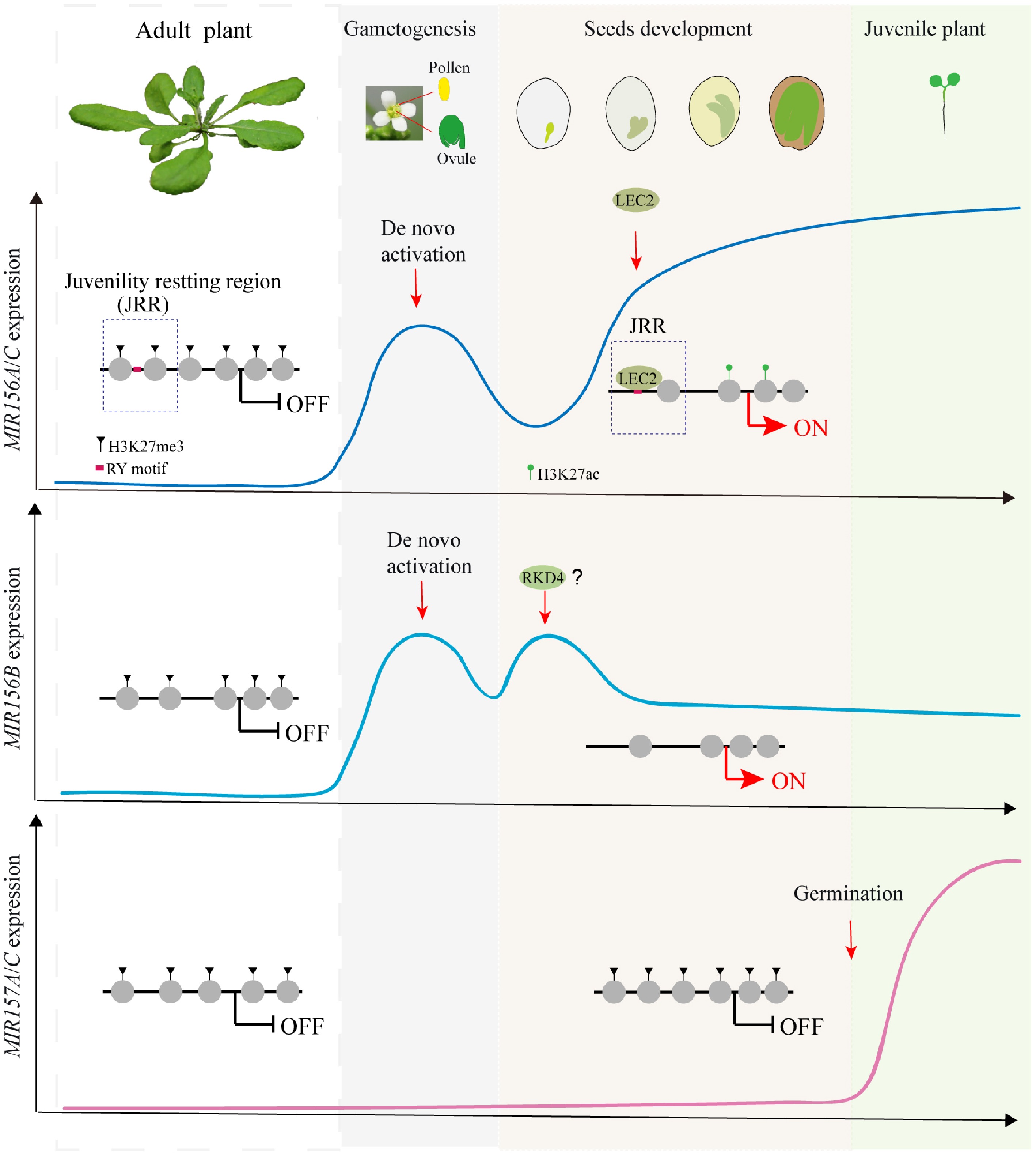
Figure 1.
Resetting of juvenility via reactivation of MIR156/157 during seed development in Arabidopsis. MIR156A–C and MIR157A/C play dominant roles in phase transition, but their reactivation occurs through distinct reprogramming routes. MIR156A–C are de novo activated in gametogenesis, with MIR156A/C subsequently reset from Polycomb repression to active chromatin by LEC2 through its binding to the RY motif in the juvenility resetting region during embryogenesis, whereas MIR156B, maintaining a constitutively open regulatory region throughout the life cycle, is reactivated earlier by other embryonic factors. MIR157A/C, constitutively marked by H3K27me3 and maintained in a closed chromatin state throughout embryogenesis, are reactivated upon germination.
Functional redundancy within the MIR156/157 gene family provides a robust mechanism for rejuvenation in plants. This process is primarily orchestrated by miR156, with the MIR156A-C clade playing a dominant role, as demonstrated by the similar phenotypes of the mir156abc and mir156oct mutants[28]. Molecular investigations, integrating ATAC-seq and ChIP-seq analyses, delineate the precise spatiotemporal dynamics of miR156 that accumulates at high levels in juvenile shoot apices, declines during vegetative maturation, and is robustly rebuilt during reproductive development, specifically within pollen, ovules, and embryos. In contrast, its close homolog, miR157, is not detected to be reactivated in these reproductive tissues in Arabidopsis; instead, as detailed later, its resetting is deferred until seed germination[28]. This expression divergence within the family highlights the precise regulatory partitioning essential for the resetting. Notably, this pattern exhibits species-specificity; for instance, in cotton, miR157 has been shown to directly regulate ovule development[42], indicating varied functional roles across plant species.
The reprogramming process initiates with the de novo activation of MIR156A, MIR156B, and MIR156C during gametogenesis. This early activation in parental reproductive tissues primes subsequent resetting in the next generation. Following fertilization, a second, critical wave consolidates the resetting during embryogenesis. This reprogramming cascade is underpinned by divergent molecular and epigenetic mechanisms (Fig. 1). Integrated ATAC-seq and ChIP-seq analyses revealed that the MIR156A and MIR156C loci undergo a precise seed-to-seed cycle of chromatin remodeling. The Juvenility Resetting Region (JRR), a conserved upstream regulatory sequence, is in closed chromatin marked by repressive H3K27me3 in adult tissues, but is reopened and marked by active H3K27ac in embryos and juvenile tissues[21,28]. This opening resets (or erases) the parental epigenetic memory, enabling de novo transcription. In contrast, the regulatory region of MIR156B remains constitutively open throughout the life cycle, and the loci of MIR157A/C do not exhibit this cyclical chromatin pattern, indicating that different family members employ distinct regulatory strategies to achieve a coordinated resetting[28].
The embryonic reprogramming of MIR156A/C is directly orchestrated by the master regulator of embryogenesis, LEAFY COTYLEDON2 (LEC2). Using ChIP-seq to map genome-wide binding and a dexamethasone-inducible LEC2 system to establish causality, LEC2 was shown to bind directly to JRR via the RY motif, a well-characterized motif recognized by the plant-specific B3 transcription factors[43,44]. Transient activation assays and a dexamethasone-inducible LEC2 system demonstrated that LEC2 drives MIR156A/C expression[28]. Genetically, LEC2 is required to maintain the JRR in an active state: its loss in lec2 mutants leads to chromatin closure featured by increased H3K27me3 and decreased accessibility, shutdown of MIR156A/C expression, and premature loss of juvenility[28]. Notably, LEC2 binding was not detected at the MIR156B, MIR157A, or MIR157C promoters, highlighting the specificity of this pathway. MIR156B is reset earlier, during early embryogenesis, potentially through the action of another transcription factor, RWP-RK DOMAIN-CONTAINING PROTEIN 4 (RKD4)[28,45]. Meanwhile, MIR157 follows a separate, later reprogramming route. MIR157A and MIR157C are activated specifically upon seed germination independently of light signals[28]. This post-germination activation follows the earlier embryonic resetting of MIR156, ensuring the completeness of the juvenile phase restoration.
-
Vernalization—the acquisition of flowering competence through exposure to prolonged seasonal cold (winter cold)—is a vital adaptive strategy in temperate plants, synchronizing reproduction with favorable spring growth conditions[46]. Prolonged exposure to cold establishes a chromatin-based 'vernalized state' at key flowering regulatory loci in crucifer plants, which is stably maintained during subsequent growth[22,23]. To ensure each new generation retains the requirement for vernalization, this environmentally acquired epigenetic state must be reset. In Arabidopsis thaliana, this is achieved through the resetting of the chromatin state at the potent floral repressor FLC during seed development[22]. This process provides a foundational paradigm for erasing the somatically-acquired 'memory of winter cold' and restoring developmental plasticity to each new generation.
In Arabidopsis winter-annual accessions, high expression of the MADS-box transcription factor FLC prevents flowering in the absence of cold[47,48]. Prior to cold, the scaffold protein FRIGIDA (FRI) recruits active chromatin modifiers, establishing a highly active FLC chromatin state marked by histone acetylation, H3K4me3, and H3K36me3[22,49]. Cold exposure triggers FRI sequestration into nuclear condensates, leading to rapid transcriptional downregulation[50,51]. Stable epigenetic silencing is then established by a cold-specific PRC2 that incorporates the cold-induced protein VERNALIZATION INSENSITIVE 3 (VIN3), along with its homolog VIN3-LIKE 1 (VIL1)/VERNALIZATION 5 (VRN5)[52,53]. The PRC2 complex is recruited to the bivalent Cold Memory Element (CME) in the first intron of FLC through the cooperative action of the bivalent readers EARLY BOLTING IN SHORT DAYS (EBS)/SHORT LIFE (SHL) and the B3 domain proteins VIVIPAROUS1/ABI3-LIKE1 (VAL1)/VAL2, the latter binding the RY motif within CME[54−56]. This coordinated recruitment then initiates H3K27me3 deposition. Upon return to warmth, a 'read-write' feedback mechanism promotes PRC2 and H3K27me3 spreading across the locus, conferring a stable, mitotically heritable, vernalized state that ensures flowering competence in spring[56−58]. Furthermore, Casein Kinase II (CK2) enhances this memory mechanism by phosphorylating and stabilizing the PRC2 catalytic subunits CURLY LEAF (CLF) and SWINGER (SWN), thereby promoting sustained H3K27me3 at FLC following prolonged cold exposure[59].
Following the transition to flowering, however, the somatically-acquired vernalized state must be erased in the progeny so each new plant can independently assess its environment. This resetting is not passive but a highly orchestrated, locus-specific reprogramming event embedded within seed development. The process begins before fertilization, with asymmetric reprogramming in the gametes. During male gametogenesis, sperm cells undergo extensive chromatin reprogramming—including replacement of histone H3 with a sperm-specific variant (H3.10), active H3K27 demethylation, and absence of core PRC2—which together mediate a genome-wide reduction of H3K27me3 (Fig. 2)[26,60]. However, it is important to note that H3.10 incorporation per se does not directly reset H3K27me3 at FLC; rather, the erasure at this locus is primarily attributed to active demethylation and PRC2 absence[26]. In contrast, the Polycomb-repressed state is more stable and can be transmitted through the maternal egg cell, resulting in delayed reactivation of the maternally inherited allele[61].
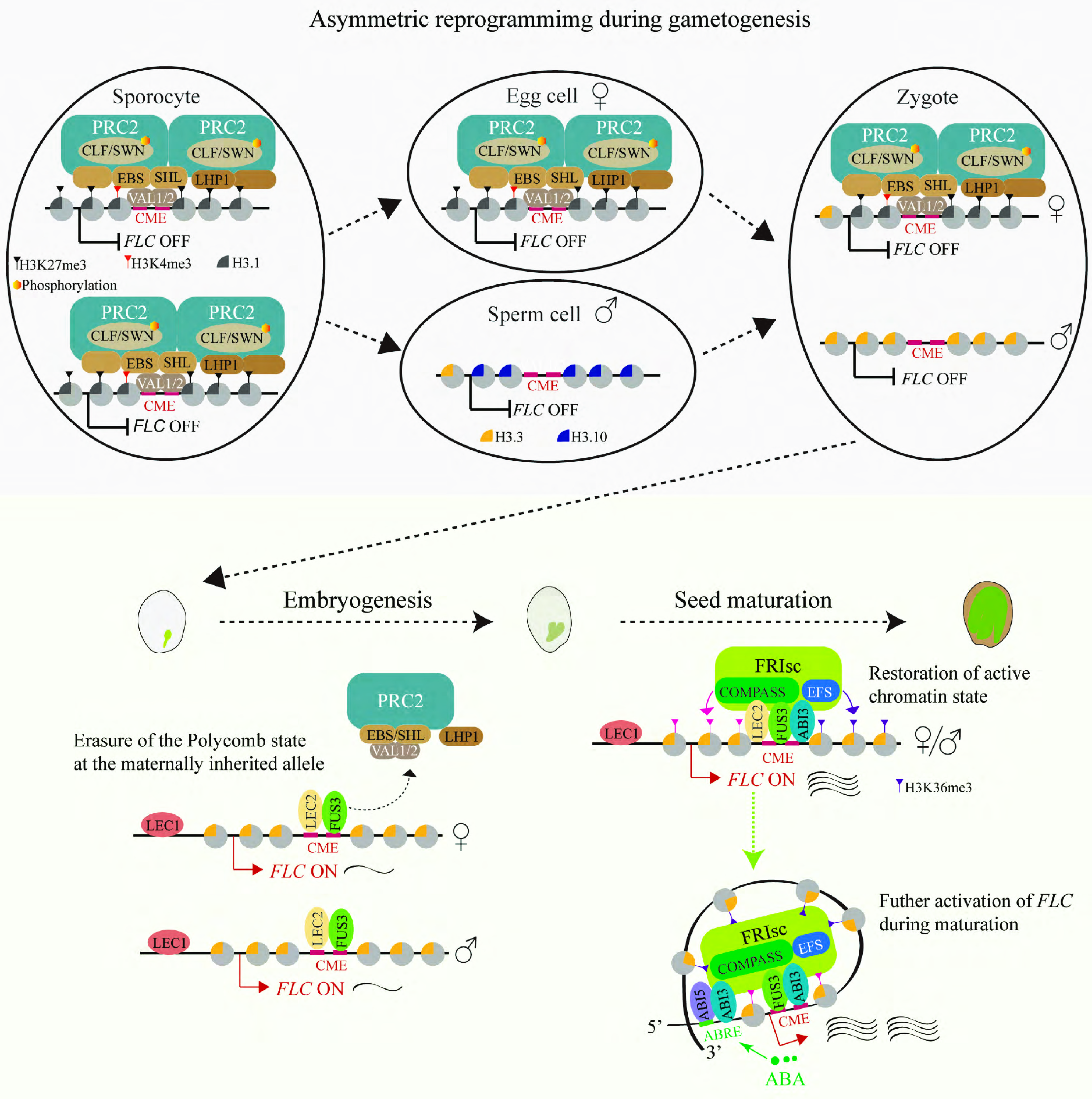
Figure 2.
Reprogramming of the vernalized state in Arabidopsis. During male gametogenesis, genome-wide chromatin reprogramming in sperm cells—mediated by active H3K27 demethylation and absence of core PRC2—depletes the vernalized state specifically at the paternal FLC allele. Histone variant H3.10 incorporation occurs in parallel but does not directly mediate H3K27me3 erasure at FLC. In contrast, the Polycomb-repressive vernalized state of FLC chromatin is transmitted to the zygote via the egg cell. After fertilization, LEC1 opens FLC chromatin, enabling B3-domain factors (LEC2, FUS3, and ABI3) to displace Polycomb-group factors. Displacement of the silencing machinery leads to progressive dilution of maternal H3K27me3 through rapid cell divisions. Concurrently, these factors recruit FRIsc, which deposits H3K4me3 and H3K36me3 to reset FLC to an active, mitotically stable state by the end of embryogenesis. ABA reinforces FLC activation in maturing seeds via the ABI5–ABI3 heterodimer, enhancing FRIsc recruitment and driving FLC upregulation. This active state is mitotically maintained in vegetative tissues, restoring the vernalization requirement in progeny.
Embryonic resetting of the vernalized state is driven by a sequential cascade of seed-specific transcription factors[22]. The pioneer factor LEAFY COTYLEDON 1 (LEC1)[62], which is expressed from the zygote stage onward, binds several CCAAT motifs in the FLC promoter to open the chromatin landscape, including that of the maternal allele[63]. This is followed by the sequential action of the B3-domain transcription factors LEAFY COTYLEDON 2 (LEC2) and FUSCA3 (FUS3) at the globular stage onward[44], followed later by ABSCISIC ACID-INSENSITIVE 3 (ABI3) at the transition stage onward[64]. These factors converge on the CME, where they perform a dual role: they physically displace the silencing machinery, VAL1/VAL2 in complex with PRC2, leading to a progressive dilution of the maternally inherited H3K27me3 through rapid cell divisions during embryogenesis[44,63,64]. Furthermore, these three transcription factors directly interact with and recruit the FRI super complex (FRIsc) to FLC chromatin[44,49,63,64]. Recruitment of FRIsc marks the pivotal transition from erasure to active re-establishment of a permissive state for transcription activation. FRIsc acts as a hub, assembling active chromatin modifiers like the COMPASS-like complex (depositing H3K4me3) and the H3K36 methyltransferase EARLY FLOWERING IN SHORT DAYS (EFS), which deposits H3K36me3[49,63,64]. This systematically overwrites the repressive landscape, constructing a chromatin environment for active transcription. By the end of embryogenesis, the Polycomb-repressed state is eliminated, and FLC chromatin is reset to an active state (Fig. 2)[44,64].
The mechanism is reinforced during seed maturation, integrating developmental cues with hormonal signaling. Accumulating abscisic acid (ABA) promotes binding of the ABA-responsive factor ABSCISIC ACID-INSENSITIVE 5 (ABI5) to the FLC promoter[64,65]. ABI5 forms a heterodimer with ABI3, and together with sustained FUS3 activity, they enhance FRIsc recruitment to both the proximal promoter and the CME, driving a significant upsurge in FLC expression in the mature seed (Fig. 2)[64]. This high-level expression in mature seeds may be adaptive, likely facilitating germination under the cool temperatures in a temperate autumn, thereby ensuring that winter-annual seeds germinate at the appropriate seasonal cue[22,66]. Ultimately, the highly active FLC chromatin state reconstructed during seed development is mitotically stable. Following germination, the reset active state is maintained in vegetative tissues, with FRI and its associated modifiers continually preserving it[63]. This restores the plant's inherent vernalization requirement. The progeny now possesses a high basal level of FLC expression, conferring the over-wintering growth habit.
-
While the seed-based resetting of FLC in Arabidopsis provides a detailed paradigm, resetting the vernalized state is widespread in winter annuals of diverse plant species[46]. Research in hexaploid bread wheat (Triticum aestivum) reveals a compelling case of convergent evolution, where this same imperative is achieved through a distinct, two-layered mechanism (Fig. 3)[67,68]. The transition to flowering in wheat is governed by a central regulatory module that integrates vernalization and photoperiod signals[23,69]. This module comprises three key genes: wheat VERNALIZATION1 (TaVRN1), TaVRN2, and TaVRN3 (the wheat ortholog of Arabidopsis florigen FLOWERING LOCUS T (FT))[70−72]. Before cold, the repressor TaVRN2 is highly expressed, repressing the expression of the florigen gene TaVRN3 and inhibiting flowering, while TaVRN1 is silenced. Prolonged cold represses TaVRN2 and activates TaVRN1 expression[69,71]. The TaVRN1 protein then further represses TaVRN2, creating a stabilizing feedback loop. With TaVRN2 suppressed, TaVRN1 cooperates with the photoperiod pathway to activate TaVRN3 expression in leaves; the resulting florigen moves to the shoot apical meristem to further amplify TaVRN1 expression on site, resulting in flowering[23,70,71,73]. Although cold-induced TaVRN1 activation is transmitted to the zygote and early embryos, and associated with a permissive chromatin environment, it is systematically reset during later embryogenesis, erasing the parental vernalized state[67,68]. Concurrently, TaVRN2—remaining silenced during seed development—is rapidly reactivated by light upon germination[67]. The resetting of these cold-induced active transcriptional states at TaVRN1 is governed by intricate chromatin dynamics[25,67,68].
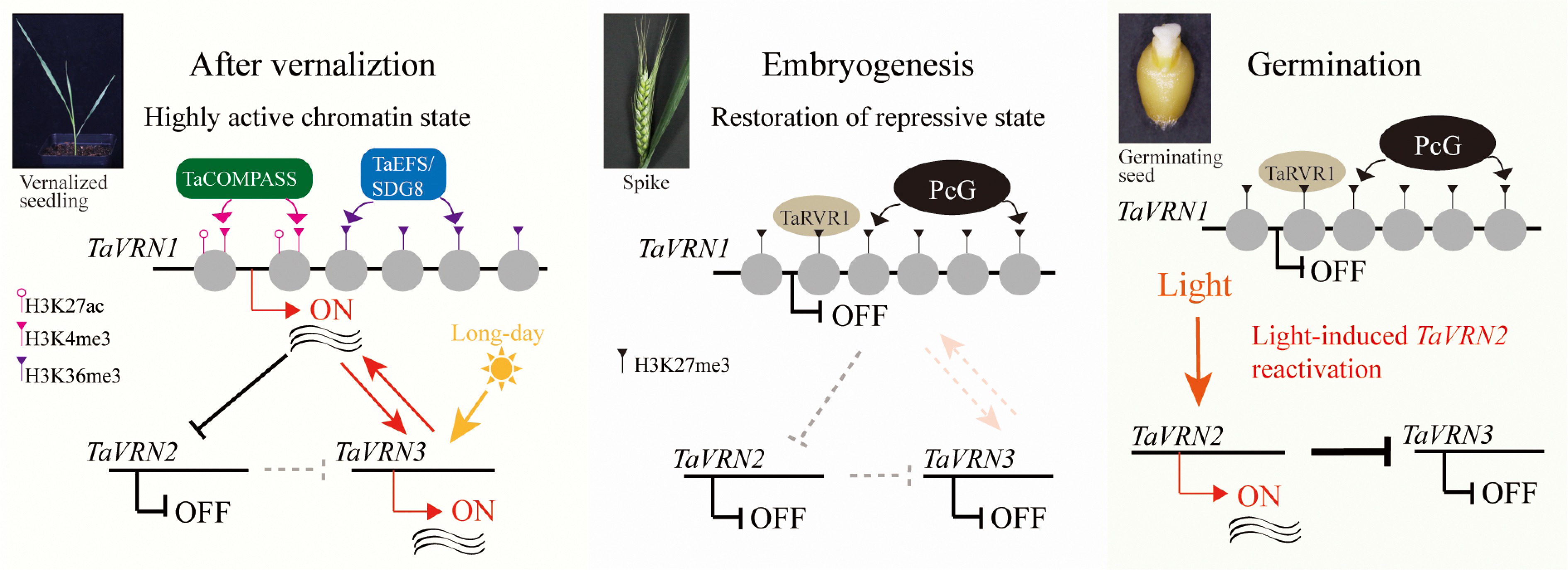
Figure 3.
Two-step restoration of over-wintering growth habit in winter wheat. Prolonged cold erases PRC2-mediated H3K27me3 at TaVRN1, enabling COMPASS-like and EFS to deposit H3K4me3 and H3K36me3, respectively, switching TaVRN1 to an active state. During embryogenesis, active marks are removed and H3K27me3 is re-deposited by Polycomb group (PcG) proteins, reinstating the Polycomb-repressed state partly via TaRVR1. Concurrently, TaVRN2 is light-reactivated during seed germination. Thus, cold-induced activation and embryonic resetting form a reversible chromatin switch that restores vernalization requirement in each generation.
During the establishment, maintenance, and resetting of the vernalized state in wheat, chromatin dynamics undergo programmed transitions genome-wide. The establishment phase is initiated by prolonged cold, which triggers a large-scale reorganization of chromatin states, most significantly within genomic domains marked by the repressive modification H3K27me3[68]. Genome-wide profiling of chromatin modifications reveals a co-regulated gene set that exhibits an antagonistic dynamic of decreasing H3K27me3 and increasing H3K36me3; the enrichment of floral development pathways within this gene set links this chromatin logic directly to the acquisition of flowering competency[68]. The maintenance phase entails the stable perpetuation of this active chromatin architecture to ensure the floral transition proceeds. During embryogenesis, a systematic reprogramming of the epigenome occurs, characterized by the genome-wide removal of active histone marks such as H3K36me3, H3K27ac, and H3K4me3, concerted re-deposition of repressive H3K27me3, and a decrease in chromatin accessibility. This collective re-establishment of the repressive landscape during late embryogenesis contributes to the restriction of totipotency and the establishment of the dormant seed state[25].
Global chromatin reprogramming converges with precise, locus-specific control, and the vernalized state is reset largely through repressing the core floral promoter TaVRN1. In seedlings without cold exposure, the TaVRN1 locus is enriched with repressive H3K27me3[67,68]. Following cold-induced activation, TaVRN1 expression persists throughout subsequent growth and development, is detectable in both male and female gametes, the zygote, and pre-embryos, but is silenced from the transition embryo stage onward[67]. Cold exposure triggers antagonistic histone modification dynamics: H3K27me3 progressively diminishes while active marks such as H3K4me3 and H3K36me3 accumulate[67,68]. The deposition of H3K36me3 is particularly critical for maintaining the active transcriptional state of TaVRN1 after cold exposure[68]. During embryogenesis, these active marks, including H3K36me3, H3K4me3, and H3K27ac, are progressively stripped away on TaVRN1 chromatin[68]. This is followed by the selective rebuilding of the H3K27me3 at the TaVRN1 locus specifically in winter wheat, which re-establishes the Polycomb-repressed state in the mature embryo[67,68]. The functional importance of these histone modifications is directly demonstrated by forward genetic studies. Accordingly, loss-of-function mutations in wheat histone H3K36 methyltransferase EFS/SET DOMAIN GROUP 8 (TaEFS/TaSDG8), which impairs H3K36me3 deposition, compromise the maintenance of the vernalized state, leading to reduced TaVRN1 expression and delayed flowering post-vernalization[68]. Conversely, genetic attenuation of the H3K27me3 'writer' complex via wheat FERTILIZATION-INDEPENDENT ENDOSPERM (TaFIE) mutation—a core PRC2 component essential for normal embryo establishment—reduces the vernalization requirement and elevates TaVRN1 expression at seedling stages, yet critically, does not prevent its initial silencing during embryogenesis[68]. This indicates that the function of locking in the silenced state (via PRC2) is separable from the mechanism of initial silencing, supporting a model where resetting is initiated by removal of active marks, with PRC2-mediated H3K27me3 deposition playing a subsequent locking role[67,68]. Consistent with this model, the wheat chromatin reader REPRESSOR OF VERNALIZATION 1 (TaRVR1)[74], a BAH-domain protein that recognizes H3K27me3, is a key executor of this resetting (Fig. 3). Knockout mutants of TaRVR1 in winter wheat exhibit a spring growth habit, with derepressed TaVRN1 in seedlings and failure to silence it during embryogenesis, accompanied by reduced H3K27me3 and elevated H3K4me3 at the locus[67].
To faithfully restore the obligate winter-annual growth habit, a complementary, environmentally-triggered mechanism resets the floral repressor TaVRN2[67]. Unlike the largely cell-autonomous epigenetic silencing of TaVRN1, the TaVRN2 is reset via a distinct, environmentally cued path. Upon germination, TaVRN2 undergoes rapid, light-triggered reactivation in an embryo-autonomous manner, specifically within the embryonic leaves, coleoptile, and scutellum[67]. This intrinsic, light-dependent reactivation restores the floral repression pathway in the newly-established seedling, thereby reinstating the vernalization requirement in each generation independent of parental history and solidifying the obligate growth habit. Thus, a two-step mechanism—embryonic resetting of epigenetic vernalized state (cold-induced TaVRN1 activation) and light-triggered reactivation of the floral repressor TaVRN2 during seed germination—ensures the re-establishment of vernalization requirement in each filial generation of winter wheat (Fig. 3)[67].
-
Seed development is a critical phase for epigenetic reprogramming, where erasure of parental marks and restoration of pluripotency are integrated into embryogenesis. Studies in the model plant Arabidopsis and the major crop wheat have elucidated the molecular epigenetic mechanisms underlying this coordinated process. Collectively, these studies illustrate that both juvenility and vernalization-mediated cold memory are reset through embryo-based, locus-specific epigenetic reprogramming. They function as dedicated circuits that reverse parental epigenetic states, whether repressive Polycomb silencing at MIR156 and FLC in Arabidopsis, or an active chromatin landscape at TaVRN1 in wheat[28,67,68]. Despite their distinct molecular routes, these pathways converge to fully erase parental somatic and environmentally-acquired imprints during seed development. This restoration of a developmentally plastic, juvenile ground state in each new generation underscores seed-based epigenetic resetting as a fundamental strategy for renewing the plant life cycle while preserving adaptive potential.
However, we must acknowledge important current limitations and unresolved questions in this field. First, the detailed mechanisms described for MIR156/157 and FLC represent in-depth case studies from Arabidopsis. Whether these paradigmatic loci are broadly representative of genome-wide resetting principles—and whether the regulatory logic of vernalization resetting defined in Arabidopsis and temperate cereals extends to other plant families—remains largely unknown. This gap is particularly significant given that many economically and ecologically important species, such as fruit trees[75,76], forage legumes[77], and ornamental plants[78,79], exhibit vernalization requirements, yet their underlying molecular networks and epigenetic dynamics are barely characterized. Systematic epigenomic profiling across seed development in diverse angiosperm lineages will be essential to determine the generality of these mechanisms and to identify whether novel regulators—including different transcription factors, chromatin readers/writers, or non-coding RNAs—have been independently recruited in different lineages. Second, it remains unclear whether environmentally induced epigenetic changes (e.g., stress memories) follow the same resetting logic as hardwired developmental programs such as juvenility. Some environmentally acquired states might escape full resetting, potentially contributing to transgenerational stress memory[10,80]. Third, the molecular triggers that specify which loci require resetting—and which transcription factors execute this process beyond the few known examples (LEC1 and B3 factors)—remain largely unidentified. Beyond these gaps, the epigenetic resetting strategies of perennial plants represent an additional frontier. Perennials, which carry years of somatic 'memory' across repeated seasonal cycles of growth and reproduction[81], likely employ dual, complementary systems to manage these long-term epigenetic states: a complete zygotic resetting during sexual reproduction to erase intergenerational 'memories', and a separate, cyclical somatic resetting (e.g., in meristems or buds) to revoke seasonal or age-related states[28,46,82].
Despite many open questions about the scope and generality of resetting, the extensive epigenetic reprogramming during seed embryogenesis demonstrates that the plasticity and dynamics of epigenetic states are fundamentally governed by chromatin-based mechanisms. A key mechanistic concept that merits expanded discussion is that of bivalent chromatin domains. Originally discovered in mammalian embryonic stem cells as regions co-marked by H3K4me3 and H3K27me3, bivalent domains are thought to maintain key developmental genes in a transcriptionally 'poised' state; silent, but ready for rapid activation upon differentiation cues[83,84]. In plants, the Cold Memory Element (CME) at FLC represents the best-characterized bivalent chromatin domain. The CME harbors both H3K4me3 and H3K27me3 prior to cold exposure, enabling the coordinated recruitment of both activating and repressive machinery. This bivalency nature underpins the CME's function as a plasticity hub that drives the switch from active transcription to Polycomb silencing, and, critically, back again during embryonic resetting[44,56]. It is tempting to speculate that similar bivalent domains may be a widespread feature at other key reprogramming loci across species, providing a general chromatin-based mechanism for implementing reversible epigenetic switches. Identifying such bivalent domains genome-wide, elucidating the 'writers', 'readers', and 'erasers' governing their dynamics, and determining their conservation across plant lineages represent pivotal future directions.
While this review has focused primarily on histone modification-mediated resetting, DNA methylation represents another major layer of epigenetic regulation that deserves consideration. In plants, DNA methylation occurs in three sequence contexts: CG methylation is enriched in gene bodies and is often associated with constitutively expressed genes, while CHG and CHH methylation is highly enriched over heterochromatic transposable elements and repeats, where they play a prominent role in silencing their expression at the transcriptional level[85,86]. Unlike the extensive reprogramming observed for histone modifications at developmental resetting loci, DNA methylation patterns undergo limited resetting during seed development. These patterns are stably propagated through mitosis and, in some cases, through meiosis, and certain methylated loci can escape reprogramming entirely, being transmitted transgenerationally as so-called epialleles—differentially methylated regions that are stably inherited across generations independent of genetic sequence changes[13,87−89]. Such epialleles have been increasingly documented across diverse plant species, where they can mediate heritable adaptive responses to environmental challenges such as climate variation and temperature stress[90]. This contrasts with the more thorough resetting of histone modification-based memory at key developmental loci such as FLC and MIR156. Understanding how these two layers of epigenetic information—histone modifications and DNA methylation—are differentially and coordinately regulated during seed development, and how their interplay shapes the balance between memory inheritance and developmental renewal, represents a critical frontier for the field[91].
The application of single-cell and spatial epigenomics[92,93], combined with advanced genome editing in non-model species and epigenome editing[94], will be essential to delineate the spatiotemporal dynamics of epigenetic resetting during seed development with unprecedented resolution. Ultimately, deciphering diverse resetting strategies will illuminate how plants balance the inheritance of adaptive information with the renewal of developmental potential—a balance that underpins their resilience in each generation. This fundamental understanding will, in turn, provide a rational framework for engineering crop productivity and sustainability in a changing climate.
-
The authors confirm their contributions to the paper as follows: draft manuscript preparation: Gao Z, Xu K, He Y; draft manuscript modification: Gao Z, He Y; manuscript and figure revision: Gao Z. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
This work was supported by grants from the Special Fund for Scientific Research of Shanghai Landscaping and City Appearance Administrative Bureau (G252415).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao Z, Xu K, He Y. 2026. Seed-based epigenetic resetting through histone modifications: erasing the past to renew the future in plants. Seed Biology 5: e019 doi: 10.48130/seedbio-0026-0017 

Seed-based epigenetic resetting through histone modifications: erasing the past to renew the future in plants
- Received: 01 April 2026
- Revised: 11 May 2026
- Accepted: 14 May 2026
- Published online: 14 July 2026
Abstract: Seed development serves as the indispensable hub for resetting epigenetic states acquired during growth and development as well as environmental responses, thereby restoring developmental plasticity in each new generation. Here we synthesize current understanding of how parental somatic memories—both developmental phase identity and environmental imprints—are mechanistically reprogrammed during seed development, with erasure of parental epigenetic states coupled to establishment of new chromatin configurations. In Arabidopsis thaliana, dedicated embryonic transcription factors orchestrate the reversal of Polycomb-mediated silencing at multiple key loci, controlling juvenility (MIR156A/C) and vernalization response (FLC), resetting these genes to a transcriptionally competent state. In hexaploid wheat (Triticum aestivum), a two-step mechanism resets the ‘vernalized state’ during seed development and seed germination. Together, these paradigmatic examples reveal that the renewal of the plant life cycle is fundamentally governed by embryo-based epigenetic circuits during seed development. Understanding of this regulatory logic provides a unifying framework for how flowering plants maintain both developmental flexibility and adaptive potential across generations.