









-
Donkey milk has been considered as an adequate alternative nourishment, in particular for people who experience cow milk protein allergy or hypoimmunity such as infants, lactating mothers, and the elderly[1]. According to the China Statistical Yearbook released in 2021, China raised 2.324 million donkeys in 2020, about 12.7% of which were from Xinjiang, China (
www.stats.gov.cn/sj/ndsj/2021/indexeh.htm ). As one of the major countries breeding donkeys, China produces 0.27 million tons of donkey milk per year[2]. With attractive health functions and highly digestive, donkey milk contains close to the same nutritional properties as human milk[1]. Recent studies demonstrated that the addition of donkey milk can improve sensory properties and acceptability of a cows milk cheese[3]. The lysozyme derived from donkey milk could significantly reduce the clostridial load on the skin of cows' teats before cleaning[4].Currently, commercial donkey milk is usually marketed as a pasteurized product where foodborne pathogens have been inactivated[5]. However, thermal processing denatures antimicrobial proteins and causes irreversible changes in heat-sensitive compounds such as vitamins and lipids[6]. Besides, some thermophilic and thermoduric bacteria such as Bacillus and aerobic spore formers show higher tolerance to heat and survive heating after thermal processing at 60–90 °C[7].
High hydrostatic pressure (HHP) as a non-thermal alternative to thermal pasteurization (generally 200–600 MPa) has similar elimination of microorganisms while almost completely retain the flavor and nutrition in milk with minimal heat[8−10]. The effects of HHP treatment on the processing of human milk at 425 MPa for 6 min allowed a similar elimination of the bacterial count below 3 cfu/mL as pasteurization, but lipase and Immunoglobulin G were better preserved in HHP processed samples compared to the pasteurized ones[11]. The good effectiveness of alkaline phosphatase in goat milk was well preserved after the treatment of HHP up to 500 MPa for 10 min, but it was completely inactivated after pasteurization[12]. Notably, revealing the microbial composition and tracing the microbial change in donkey milk before and after processing is necessary for ensuring its food safety. With the development of high-throughput sequencing (HTS), a deeper investigation was performed for the diverse bacterial communities in donkey milk[13]. Gram-negative bacteria are the most prevalent in donkey milk, and the phyla of Proteobacteria and Firmicutes are the frequent microbial composition in donkey milk[14−16]. Pseudomonas is the representative genus that contributed to the psychrotrophic spoilage of donkey milk[14,16]. The functional and the safety evaluation of lactic acid bacteria (LAB) isolated from donkey milk have been described previously, with Enterococci, Lactococcus, and Lactobacillus as the frequent genera[17−20].
Due to the potential presence of heat-resistance bacteria in donkey milk, which may lead to the failure of pasteurization, applying HHP processing to donkey milk has become an alternative strategy to ensure food safety[14]. The HHP treatment can be applied to inactivate Salmonella, E. coli, Shigella and S. aureus in raw milk at 300 MPa for 30 min, and its application can also improve the bacteriophage performance against S. aureus in milk[9,10,20]. In cow and goat milk, psychrotrophic bacteria were rarely found after processing at 450 MPa for 7 min, but up to 36 cfu/mL of psychrotrophic bacteria was recorded in the pasteurized sample at 72 °C for 15 s[21]. However, the assessment of microbiological quality in HHP treated donkey milk usually relied on traditional culture-dependent methods in previous studies, focusing on single investigations of pathogens, spoilage microorganisms or probiotic bacteria, which only showed limited indicators rather than the change of the overall microbial composition[5,18]. Although HTS has been applied to fresh or cold-stored donkey milk to obtain sufficient information to assess the safety of dairy products, the microbial composition in HHP treated donkey milk has not been described previously[5,14].
In this study, HTS technology was used to reveal the microbial composition in donkey milk, evaluating the effects of the HHP treatment on microbial quality in donkey milk. For further understanding, the surviving bacteria were identified based on plating and 16S rRNA genes to evaluate potentially probiotic and pathogenic organisms in donkey milk. This study provides further support for the control of microbial contamination and characterizes cultivable bacteria existing in donkey milk.
-
Fresh donkey milk was obtained from Urumqi, Xinjiang, China. A total of seven healthy Xinjiang breed donkeys were selected as the source of donkey milk. All donkey milk was transferred from bulk milk containers into 50 mL sterile tubes, transported to the laboratory with ice bags, and stored at −80 °C immediately. High-quality library preparation is critical for successful HTS analysis, but abundant antibacterial functional substances and low microbial population in donkey milk can limit the quantity and quality of DNA extraction[22,23]. To obtain the information on bacterial composition and communities comprehensively, fresh donkey milk was incubated at 37 °C with different times of 5, 8, 15, and 22 h for artificial bacterial enrichment, respectively[24]. The bacteria population in donkey milk was enumerated on Plate Count Agar (PCA) (Solarbio, China) after dilution with sterilized peptone water (Land Bridge, China). To comprehensively obtain the information on bacterial diversity in donkey milk, bacterial growth at an exponential phase was selected for further analysis. The change in the bacterial population before and after the treatment of HHP was expressed by plating counts. Student's t-test was used for the statistical significance of cell counts between the untreated and HHP treated donkey milk.
The treatment of HHP for donkey milk
-
A sample of 10 mL donkey milk was packed in a sterile polypropylene pouch and then hot sealed to ensure its vacuum. Based on the initial exploration of the effect of high pressure on the total bacterial count, we finally chose 500 MPa, 10 min as the optimum condition for further analysis[25]. Ultra-pure water was used as a pressure transmission medium. Donkey milk without the treatment of HHP was used as a reference (untreated group), and each experiment was set up with seven biological replicates. Cell counts before and after pressure treatment were enumerated by surface plating on the PCA agar plate (Solarbio, China).
DNA extraction and amplification for HTS sequencing
-
Microbial DNA used for HTS from donkey milk was extracted and purified by E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA, USA). The quality of the final DNA was checked by 1% agarose gel electrophoresis and the concentration was determined by NanoDrop2000 (Thermo Scientific, Wilmington, USA). The V3-V4 region of the 16S rRNA genes with the primers 338F and 806R was amplified by PCR amplification[26] (Supplemental Table S1). The PCR reaction mix and procedures for NGS are shown in Supplemental Table S1.
Table 1. Percentages of Gram-positive and Gram-negative genera in the untreated and HHP treated donkey milk.
Untreated donkey milk HHP treated donkey milk G− genera 98.47% G− genera 96.09% Pseudomonas 42.82% Pseudomonas 37.90% Enterobacteriaceae 26.20% Enterobacteriaceae 31.04% Acinetobacter 23.54% Acinetobacter 18.72% Erwinia 2.86% Enterobacter 3.69% Enterobacter 2.69% Erwinia 3.69% Enhydrobacter 0.34% Enhydrobacter 0.70% Chryseobacterium 0.02% Chryseobacterium 0.24% Christensenellaceae R-7 group 0.11% G+ genera 0.88% G+ genera 1.5% Rothia 0.41% Kocuria 0.50% Kocuria 0.18% Rothia 0.48% Macrococcus 0.13% Rhizobiaceae 0.32% Bacillus 0.12% Microbacterium 0.11% Rhizobiaceae 0.02% Macrococcus 0.05% Microbacterium 0.02% Bacillus 0.04% Bacteria isolation and identification from donkey milk
-
Donkey milk was diluted 10-fold serially with peptone water, and 100 μL dilution was inoculated onto Luria-Bertani (LB) agar (Solarbio, China) at 37 °C for 24 h, consequently colonies with different characteristics were randomly selected for the strain isolation. After further purification on LB medium, bacterial isolates were prepared as 25% glycerol stock and stored at −80 °C. DNA extraction was used by bacteria DNA kit (TIANGEN, China), and PCR processing was used for amplifying 16S rRNA genes with 27F and 1492R[27] (Supplemental Table S2). The PCR reaction mix and procedures for bacteria identification are shown in Supplemental Table S2. 16S rRNA genes sequencing analyzed by Sangon Biotech Co., Ltd (Shanghai, China) was confirmed using the NCBI BLAST search.
To verify potential fermentation performance, Bromcresol Purple Dextrose Broth (Hopebio, China) was used to evaluate the activity of acid and gas production from glucose for the identification of the characteristics.
Illumina MiSeq sequencing of 16S rRNA genes
-
Purified amplicons were pooled in equimolar and paired-end sequenced (2 × 300) on an Illumina MiSeq platform (Illumina, San Diego, USA) using the manufacturer's protocol provided by Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China). QIIME software (version 1.9.1,
http://qiime.org ) was performed to analyze sequences and Operational taxonomic units (OTUs). OTUs were clustered with a 97% similarity cutoff through UPARSE (version 7.1,http://drive5.com/uparse/ ). The raw reads have been submitted to the NCBI Sequence Read Archive (SRA) database, and the Accession Number was PRJNA838247.Statistical analysis of HTS sequencing
-
The online platform Majorbio Cloud Platform (
www.majorbio.com ) was used for sequencing analysis. Chimeric sequences were identified and removed through UCHIME. Raw fastq sequences were performed using fastp (https://github.com/OpenGene/fastp ,version 0.20.0) for quality control, and the DNA fragments were merged using FLASH (www.cbcb.umd.edu/software/flash , version 1.2.7). A total of 686,703 high-quality 16S rRNA gene reads were obtained with an average length of 443 bp. The average quality score was above 20 with over a 50 base pairs. OTUs were annotated through Ribosomal Database Project (RDP) Classifier (version 2.2,http://rdp.cme.msu.edu/ ) and compared to the Silva 16S rRNA database (v138) (Release 138,www.arb-silva.de ) at a confidence threshold of 70%. Alpha diversity was measured using the software Mothur (version 1.30.2,www.mothur.org ). Alpha diversity analysis was performed based on Wilcoxon's test. The coverage, diversity and richness of community were assessed by the Coverage index, Shannon index, and the Sob and Chao index, respectively. Beta diversity was evaluated by principal component analysis (PCoA) based on unweighted UniFrac distance. PCoA analysis was performed to compare the difference of microbial composition between the untreated and HHP groups. The analysis of similarities (ANOSIM) with 999 permutations was performed to obtain reliable p-values to assess the similarity. Besides, linear discriminant analysis (LDA) effect size (LEfSe) analysis was used to find biomarkers that contribute to group division. -
The microbial population calculated by plating counts during incubation are shown in Fig. 1a. With the total bacterial count of 5.71 log cfu/mL after 5 h, it was increased to 6.87 log cfu/mL after 8 h. After the incubation for 15 h, the total bacterial count was increased to a relatively stable amount of 8.09 log cfu/mL, and then it was only increased to 8.21 log cfu/mL after 22 h (Fig. 1a). The total bacterial count was only increased to 8.21 log cfu/mL after 22 h (Fig. 1a), where the number of dead cells also increased. As the number of dead cells in donkey milk was increased with the microorganisms entering a stable growth period, the samples of donkey milk enriched for 8 and 15 h were selected for the treatment of HHP[28]. In the donkey milk with pre-incubation of 8 or 15 h, the treatment of HHP caused significant reductions of bacteria, with the cell counts of about 3−4 log cfu/mL (student's t test, P < 0.001) (Fig. 1b). Considering the same significant differences in HHP treatment after 8 h or 15 h, and the smaller number of dead bacteria in the logarithmic phase, the samples of donkey milk incubated at 37 °C for 8 h were selected for further DNA extraction and HTS analysis.

Figure 1.
The total bacterial count at different incubation time at 37 °C. (a) Fresh donkey milk was incubated at 37 °C with different times of 5, 8, 15, or 22 h for artificial bacterial enrichment, respectively. (b) Samples incubated for 8 and 15 h were consequently treated by HHP at 500 MPa at 20 °C for 10 min. Student's t test was used for the statistical significance.
Microbial diversity based on alpha and beta analysis
-
Alpha diversity was used to evaluate the bacterial richness and evenness in the untreated or HHP treated donkey milk. Coverage index exceeding 0.99 indicated that the majority of microbial taxa were observed in both the untreated and HHP treated donkey milk (Fig. 2a). There was no significant difference in the Shannon index between the untreated and HHP treated donkey milk (P > 0.05), indicating that the treatment of HHP did not change the bacterial evenness in donkey milk (Fig. 2b). In the richness index of community distribution, the Sob and Chao index showed that the treatment of HHP changed the bacterial richness in donkey milk, and the difference was statistically significant (P < 0.05) (Fig. 2c & d).
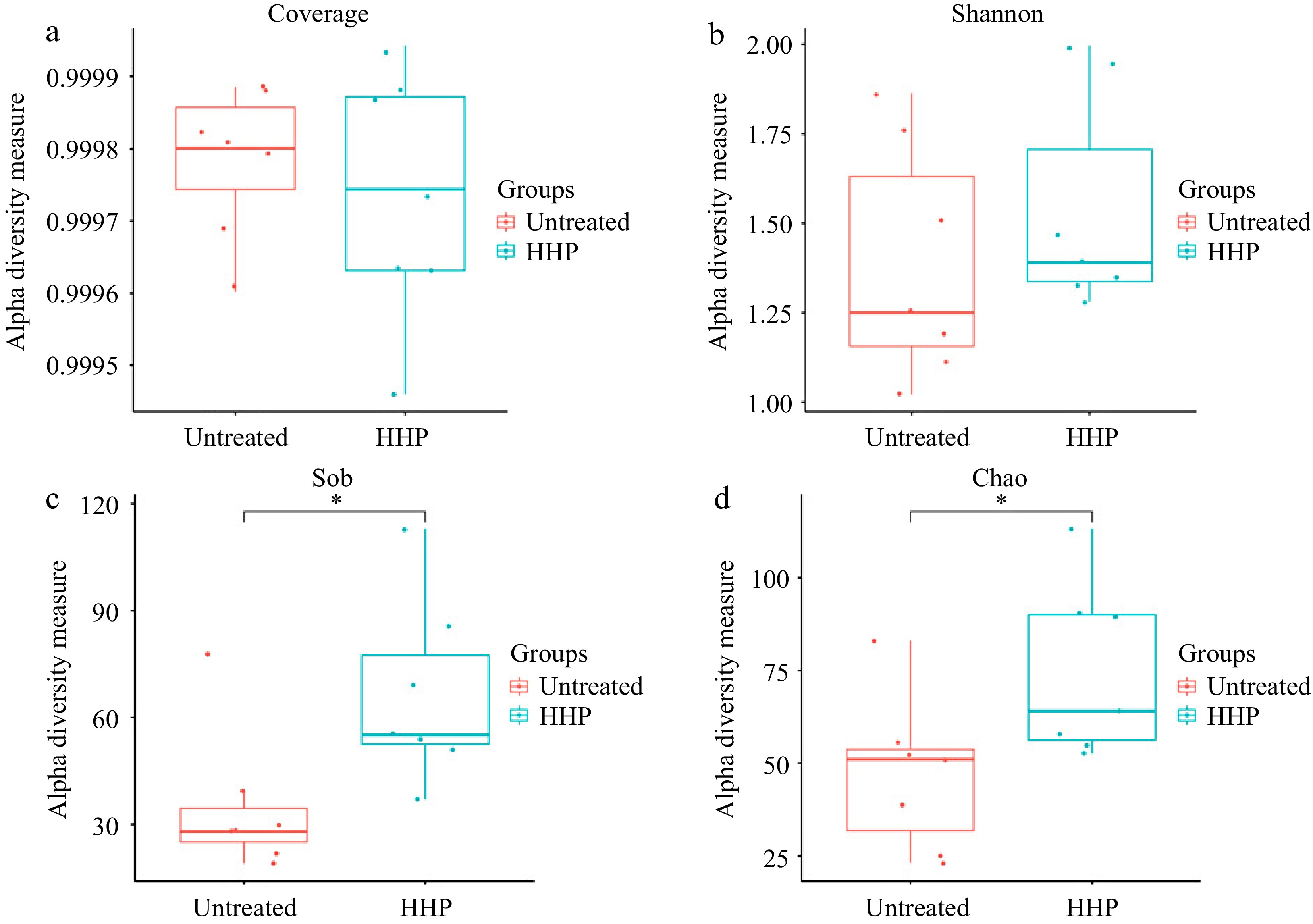
Figure 2.
(a) Alpha Diversity analysis of Coverage index, (b) Shannon index, (c) Sob index and (d) Chao index in untreated and HHP treated (500 MPa at 20 °C for 10 min) donkey milk. Wilcoxon test was used for the statistical significance. * means P < 0.05.
Principal component analysis (PCoA) based on unweighted UniFrac distance and Analysis of similarities (ANOSIM) generated by beta diversity are presented in Fig. 3. Beta analysis showed that the microbial composition in HHP treated donkey milk could be separated successfully from the untreated donkey milk at OTU level (Fig. 3). Comparing the two colors within the PCoA plot, the definitive separation between the untreated and HHP treated donkey milk demonstrated that the treatment of HHP significantly affected the microbial composition in donkey milk (Fig. 3a). For the ANOSIM analysis, the R value of 0.435 showed that the difference of the bacterial distribution between the untreated and HHP treated donkey milk was significantly greater than that within the two groups, confirming that the bacterial composition in the HHP treated donkey milk was significantly different from that in the untreated donkey milk (Fig. 3b, P = 0.006).
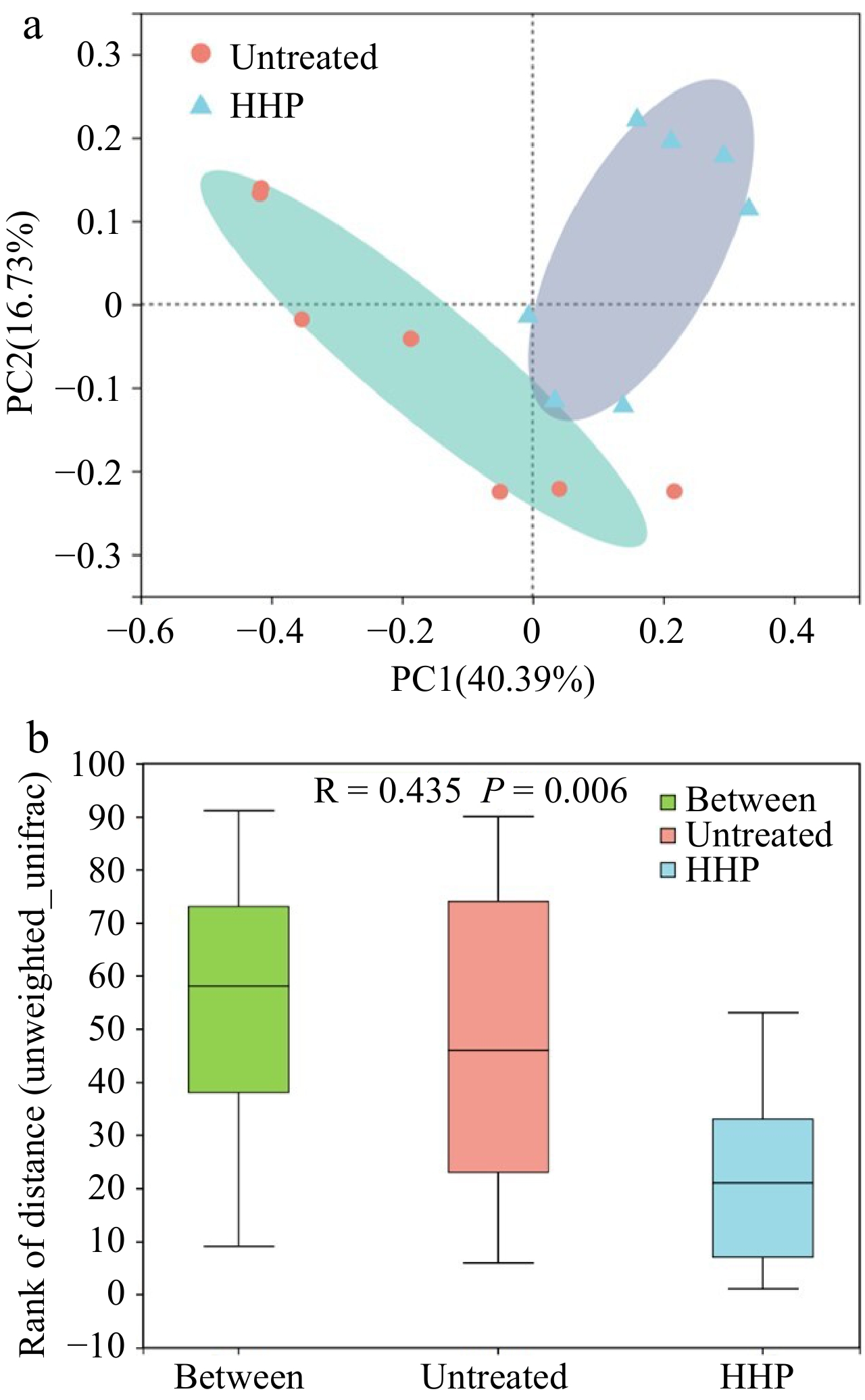
Figure 3.
(a) Principle Coordinate Analysis (PCoA) based on genus level in untreated (red) and HHP treated (blue) donkey milk, using unweighted UniFrac distance for separation. ANOISM analysis was assessed with 999 permutations and showed the difference in the microbial community between untreated and HHP treated donkey milk. (b) The distance was calculated based on genus level.
Microbial community in the untreated and HHP treated donkey milk
-
To further clarify the effect of HHP treatment on the microbial composition in donkey milk, microbial composition in the untreated and HHP treated donkey milk are shown in Fig. 4 and Table 1. The total sequencing reads were assigned to seven phyla with 43 genera in the untreated donkey milk, whereas six phyla with 65 genera were observed in HHP treated donkey milk.

Figure 4.
Relative abundance of operational taxonomic units (OTUs) at (a) family and (b) genus level in untreated and HHP treated donkey milk.
The major bacteria in the untreated and HHP treated donkey milk at family and genus levels are shown in Fig, 4 (abundance of family or genera < 0.1% were clustered as others). In the untreated and HHP treated donkey milk, the vast majority of OTUs were clustered to Proteobacteria (98.79% vs 97.33%) at phylum level in both groups (data was not shown). As shown in Fig. 4a, similar communities at family level were present in donkey milk before and after treatment with HHP. Pseudomonadaceae, Moraxellaceae, Enterobacteriaceae, and Erwiniaceae were the dominant members of the microbiota in the both groups (total relative abundance > 0.1%). However, after treatment with HHP, the abundance of Pseudomonadaceae and Moraxellaceae (23.88% vs 19.42%) was decreased by 4.92% and 4.46%, respectively, whereas the proportions of Enterobacteriaceae and Erwiniaceae was increased by 6.77% and 0.83% at family level, respectively (Fig. 4a).
Notably, the relative percentage of Pseudomonas and Acinetobacter relating to milk spoilage were both reduced in donkey milk after the treatment of HHP (Fig. 4b, Table 1). Compared with the untreated samples, a significant decrease of 4.92% was observed for the relative percentage of Pseudomonas in the HHP treated donkey milk (P < 0.05), meanwhile, the relative abundance of Acinetobacter was also decreased by 4.82% after treatment with HHP (P < 0.05). However, at the level of genus, the relative abundance of Enterobacteriaceae, Erwinia, and Enterobacter was increased in donkey milk after the treatment of HHP (Table 1).
The common and difference of bacterial proportions in the untreated and HHP treated donkey milk
-
The common bacteria in the untreated and HHP treated donkey milk were evaluated consequently through the performing of Venn diagrams (Fig. 5). It was found that six common phyla and 60 genera were shared between the untreated and HHP treated donkey milk (Fig. 5a & b). Specifically, the number of phyla observed in the HHP treated donkey milk was less than that in the untreated donkey milk, but the number of genera was 1.25 times of the observation in the untreated donkey milk (Fig. 5a & b). Among the shared genera, Pseudomonas and unclassified Enterobacteriaceae were the two dominant genera with high percentages of 40.50% and 28.72%, respectively (Fig. 5c).
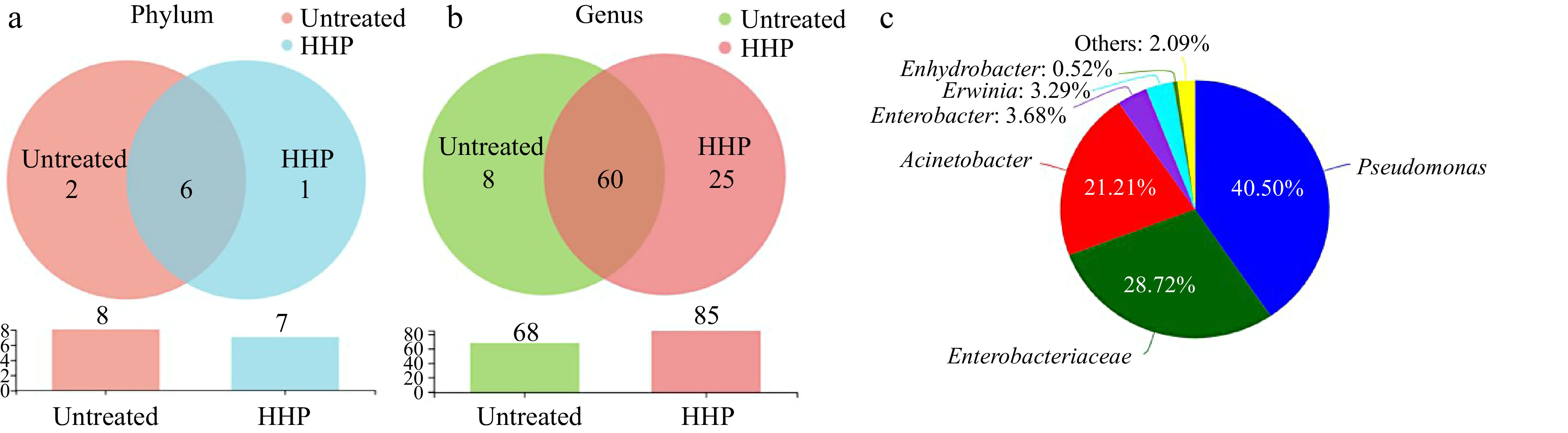
Figure 5.
Venn diagram of operational taxonomic units (OTUs) at (a) phylum and (b) genus level. (c) Genera of microbiota communities shared in two groups (abundance of genera < 0.5% were clustered as others).
To identify the difference clade in bacteria structure between the untreated and HHP treated donkey milk, LEfSe analysis was performed using all-against-all analysis from phylum to genus level. As shown in Fig. 6, with an LDA score threshold of > 2.0, three clades within the untreated group and 47 clades within the HHP group were identified. Compared to that in the untreated donkey milk, LEfSe analysis revealed an increased representation of bacterial clade after the treatment of HHP (Fig. 6a). The bacterial taxa with differential abundances were evaluated by LDA effect size analysis between the untreated and HHP treated donkey milk (Fig. 6b). The phylum Firmicutes was significantly enriched in the untreated donkey milk (LDA = 2.62, P < 0.05), mainly consisting of the genus Bacillus. Compared with the untreated donkey milk, more taxa differences were enriched in donkey milk after the treatment of HHP, including the class of Clostridia (LDA = 3.36, P = 0.012), Bacteroidia (LDA = 3.21, P < 0.05) and Alphaproteobacteria (including the family Rhizobiaceae) (LDA = 3.21, P < 0.05).

Figure 6.
(a) Differences of bacterial composition between untreated and HHP treated donkey milk based on LEfSe Analysis. (b) Linear discriminant analysis (LDA) scores calculated for differentially abundant bacteria intwo groups, only LDA score above 2 are shown.
LAB isolation based on culture-depended methods from donkey milk
-
To further improve the taxonomic resolution and allow physiological studies, culture-dependent methods combined with full-length 16S rRNA genes sequencing were used for strain isolation and identification. A total of 26 strains were isolated and purified from the untreated and HHP treated donkey milk, respectively. Isolated strains comprised of pathogens, spoilage bacteria, and LAB are shown in Supplemental Table S3. A total of 26 bacterial strains were identified into six different genera including Enterobacter spp. (12 strains), Klebsiella spp. (six strains), Escherichia spp. (three strains), Enterococcus spp. (two strains), Pseudomonas spp. (two strains), and Acinetobacter spp. (one strain), respectively. Except for the genus Klebsiella, the other five genera were captured by HTS (Enterococcus was clustered as others, data was not shown) (Table 1 & Supplemental Table S3). To evaluate the effects of isolated strains on physicochemical properties of donkey milk, gas- and acid-producing abilities of each isolate were determined consequently. As shown in Supplemental Table S3, Enterobacter asburiae was found to be the most abundant species (five strains), following by Klebsiella oxytoca (three strains) and Escherichia coli (three strains). Only Pseudomonas spp. was recovered from HHP treated donkey milk, indicating that Pseudomonas spp. was the most pressure resistant genus among all of the isolates. Pseudomonas spp. was found to survive after the treatment of 500 MPa for 10 min, which was one of the important genera that caused milk corruption during the storage of donkey milk. A total of 18 strains were considered to have gas-producing ability, including Escherichia spp. (all three strains), Enterobacter spp. (all 12 strains), and Klebsiella spp. (three out of the six strains). Notably, the other three strains belonging to Klebsiella spp. did not have the gas-producing ability, including Klebsiella oxytoca MR7, Klebsiella oxytoca N7 and Klebsiella sp. SR55. Besides, the strains belonging to both Acinetobacter spp. (one strain), Enterococcus casseliflavus (both of the two strains), and Pseudomonas spp. (both of the two strains) did not have gas production. The bacteria isolated from donkey milk was considered as acid-producing positive when the pH value of its broth cultures was recorded as less than 6.5[29]. Almost all isolated bacteria have the acid-producing ability, except for Acinetobacter strain B5. Among them, Klebsiella spp., Escherichia coli, and Enterococcus casseliflavus had the best acid-producing ability.
-
To date, only a few studies have focused on the contrast of microbial communities and diversity in donkey milk before and after the treatment of HHP based on culture-dependent methods[30−34]. This study mainly investigated the bacterial communities in the untreated and HHP treated donkey milk through HTS based on 16S rRNA genes (V3-V4) instead of traditional methods, accordingly revealing the effect of the HHP treatment on the microbial composition and its inactivation of pathogens.
Common bacterial composition in the untreated and HHP treated donkey milk
-
The dominant phyla found in the untreated donkey milk in this study, being mostly composed of Proteobacteria, showed the same prevalence as that found in our previous research, which also observed Proteobacteria as the highest member in donkey milk from Xinjiang. China (Fig. 3)[16]. However, compared with the untreated donkey milk, the number of genera was 1.25 times in HHP treated donkey milk. It is speculated that the increase in the number of genera is associated with the decrease in bacterial load after the HHP treatment in donkey milk. Because of the constraint of the total number sequences, some certain OTUs at low abundance might fall under the detection limit (> 0.1%) in the untreated donkey milk. After the HHP treatment, some genera with a low relative abundance initially (< 0.1% in the untreated samples) might be detected with the decrease of the dominant number of OTUs[35]. It is speculated that the dry climate in Xinjiang is an important reason for the dominant relative abundance of Proteobacteria as a previous study found a consistent correlation between the high abundance of Proteobacteria and the dry climate[36]. Consistent with the present study, Proteobacteria was the dominant presence phyla in goat milk and camel milk with about 71.31% or 48.73%, respectively[30−32]. However in sheep milk and buffalo milk, Firmicutes became the dominant presence in the microbial composition with about 64.44% or 42%, respectively[33,34]. Although many of the microorganisms detected in this study were previously reported, the genus Enhydrobacter observed in the untreated group was not previously detected in donkey milk (Fig. 3 & Table 1)[14,15]. Compared to the previous studies on bacterial composition in donkey milk, the relative abundances of Actinobacteriota (0.71%), Firmicutes (0.35%), and Bacteroidota (0.09%) were observed in lower proportion in the untreated donkey milk[14,16].
The different bacterial composition in the untreated and HHP treated donkey milk
-
In the present study, the total relative abundance of the G− bacteria was much higher than that of the gram-positive (G+) bacteria (98.47% vs 0.88%) (Table 1). Compared to the G+ bacteria that detected in the untreated donkey milk, a higher relative abundance of G− bacteria was observed in donkey milk, resulting from the presence of a high concentration of lysozyme[4]. Lysozyme can hydrolyze bacteria by degrading peptidoglycan (PG)[37]. In G− bacteria, the lipopolysaccharide (LPS) could form an effective barrier to block the hydrolysis of cell walls by lysozyme[37]. The HHP treatment showed more effective inactivation on G− bacteria compared to G+ bacteria in donkey milk (Table 1). It was speculated that HHP treatment enhanced the permeability of the outer membrane and made lysozyme easily pass through the outer membrane barrier to inactivate G− bacteria[38]. Instead, G+ bacteria are surrounded by a thick peptidoglycan layer, which can protect cells from the attacks of HHP compared to G− bacteria[39].
In our previous study, there was not such a big difference between the total relative abundance of the G+ and G− bacteria (52.76% vs 46.03%)[16]. Compared to our previous study, the untreated donkey milk in this study has a higher relative abundance of Pseudomonas (42.82% in this study vs 1.26% in our previous study) and a lower relative abundance of Macrococcus (G+) (0.05% in this study vs 32.96% in our previous study), which was speculated to be the principal cause of this big difference between G+ and G− bacteria[16] (Table 1). The geographic variations were the reasonable assumption for the difference in G+ and G− composition between this study and our previous one, including climatic conditions, water, and fodder between Urumqi (this study) and Yupurga (previous study)[16,40]. The high relative abundance of Macrococcus was frequently observed in dairy products from southern Xinjiang[16,41]. Pseudomonas was a typical class of spoilage psychrotrophic bacteria that can grow and metabolize at low temperatures (range of 4−42 °C)[14,42]. Compared to the climatic conditions in Yupurga (southern Xinjiang), lower temperatures in Urumqi (northern Xinjiang) might encourage the increase of Pseudomonas in donkey milk[40]. The condition of −80 °C can minimize the efficiency of temperature on the microbial composition when the donkey milk was transferred before the further process in the laboratory. However, some psychrotolerant bacterium were able to grow, which was likely a factor of the dominant presence of Pseudomonas[30]. Cold shock proteins (CSPs) and cold acclimation proteins (CAPs) could regulate the metabolic function, which confers the ability to survive at low temperatures for Pseudomonas[43]. CAPs and CSPs maintain the single-stranded state of the target DNA or RNA by destroying the secondary structure at low temperature[43]. The single-stranded state of DNA or RNA structure makes efficient transcription and translation possible[44]. Consequently, more attentions should be paid to the effects of cold environment on the bacteria communities in donkey milk, which may lead to the increase of Pseudomonas.
Besides, the family Rhizobiaceae observed in the untreated and HHP treated donkey milk was usually from the fodder ingested by donkeys[45]. Additionally, in our study, pathogens including Salmonella, Listeria, or Campylobacter were not detected but the genus Bacillus was observed in the untreated donkey milk, showing the same relative abundance as the previous investigated samples of donkey milk produced from Shandong, China[16].
Bacterial isolation from the untreated and HHP treated donkey milk
-
In the present study, two different strains belonging to Enterococcus casseliflavus were isolated from the untreated donkey milk (Supplemental Table S3), which usually have notable viability in a broad range of pH, pasteurization temperatures, and 6.5% sodium chloride[46]. Besides, their use in probiotics has been described in recent studies as high proteolytic and lipolytic activities and citrate metabolism was considered to be an important cause of cheese ripening[47]. As natural inhabitants of the gastrointestinal tract of animals, E. casseliflavus has usually been observed in untreated cow milk and cheeses, but very few have been described in donkey milk[18]. Enterococcus faecalis isolated from Italian cheeses could decrease pH of 10% (w/v) reconstituted skim milk powder to values around 4.95 after 24 h, in contrast, E. casseliflavus isolated in our untreated donkey milk exhibited better acidifying ability which can produce enough acid to lower pH in a liquid medium below 4.6 during incubation of 24 h at 37 °C[48]. Therefore, the further exploration can be performed to investigate the potential fermentation characteristics of Enterococcus strains in donkey milk.
The decrease of Pseudomonas in the HHP treated samples revealed that HHP inhibited their growth and reduced the risk of spoilage during storage. Significantly, although previous studies have proved that the treatment of HHP can inactivate Pseudomonas in PBS buffer (more than 6 log cfu/mL) efficiently under the pressure of 300 MPa for 15 min, Pseudomonas monteilii was still found to be recovered from our HHP treated donkey milk[49]. Previous study has shown that the population of Pseudomonas was lack of growth in pasteurized buffalo milk stored for up to 21 d[34]. It was speculated that Pseudomonas was more sensitive to pasteurization comparing to HHP treatment[34]. Casein and lactose in donkey milk are speculated to be the major contributors for the surviving of Pseudomonas monteilii during HHP treatment[50]. Besides, refrigerated storage after HHP treatment could promote the recovery and growth of Pseudomonas[51]. Most Pseudomonas do not pose a serious risk to public health, but Pseudomonas monteilii isolated from our samples was considered as an opportunistic pathogen that leads to acquired infections in hospitals[52]. Pseudomonas monteilii was frequently observed in soils, water drains, and vertebrate skins rather than in dairy products, accordingly we postulate that the presence of this species in donkey milk is due to environmental contamination[53].
In this study, we compared the change of microbial composition in donkey milk before and after the treatment of HHP, and also revealed the existence of bacteria that was not previously observed in the untreated donkey milk. Although the relative percentage of Acinetobacter and Pseudomonas was significantly decreased in donkey milk after HHP treatment, previous studies showed that Acinetobacter was commonly resistant to heat and various antimicrobials[54,55]. Among them, A. baumannii was an important pathogen causing infection in immunocompromised patients[56]. Besides, Pseudomonas was considered as a spoilage microorganism and opportunistic human pathogen, including the high pressure treated donkey milk isolate P. monteilii as mentioned above[52,53]. The occurrence and survival of these bacteria after processing may still pose a potential health risk to consumers. Moreover, a transmissible locus of stress tolerance conferring pressure/heat resistance has been recently discovered to exist in Pseudomonas, two strains of which have been isolated from the HHP treated donkey milk in this study[57,58]. The acquisition of such resistance genes or genomic island through lateral gene transfer might promote the pressure resistance of microbiota, thereby posing a possible challenge to the single application of HHP treatment in donkey milk[59].
-
In this study, effects of HHP treatment on bacterial composition in donkey milk was conducted by HTS. Bacterial evenness was similar between the untreated and HHP treated groups. The treatment with HHP changed the microbial population and bacterial diversity but not the domination of Pseudomonas spp. in donkey milk. After treatment with HHP, the relative percentage of Pseudomonas and Acinetobacter in donkey milk was significantly decreased. A rare genus Enhydrobacter was observed in the untreated donkey milk. Enterococcus with good acid-producing capacity isolated from the untreated donkey milk had potential fermentation performance, and Pseudomonas which was usually considered to be high pressure sensitive could survive after treatment with HHP. Whereas the reductions of living cells have been significantly observed, the presence of pressure resistant bacteria in HHP treated donkey milk needs closer tracking and attention.
This work was supported by National Natural Science Foundation of China (Grant No. 31901798) and the Open Project Program of the Beijing Laboratory of Food Quality and Safety, Beijing Technology and Business University (FQS-202101).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 PCR conditions used for NGS.
- Supplemental Table S2 PCR conditions used for bacteria identification.
- Supplemental Table S3 Strain isolation from the untreated and HHP treated donkey milk.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Kong J, Luoyizha W, Zhao L, Fan C, Li H, et al. 2023. Effects of high hydrostatic pressure treatment on bacterial composition in donkey milk studied by high throughput sequencing. Food Innovation and Advances 2(2):85−94 doi: 10.48130/FIA-2023-0010 

Effects of high hydrostatic pressure treatment on bacterial composition in donkey milk studied by high throughput sequencing
- Received: 07 January 2023
- Accepted: 15 February 2023
- Published online: 17 April 2023
Abstract: High hydrostatic pressure has become a non-thermal alternative to thermal pasteurization in dairy product processing. In this study, we investigated the effects of the treatment of high hydrostatic pressure on the bacterial composition in donkey milk using high-throughput sequencing technology and culture-dependent methods. Compared with the microbial composition in the untreated donkey milk, the relative percentage of Pseudomonas and Acinetobacter in donkey milk after high hydrostatic pressure was significantly decreased by 4.92% and 4.82%, respectively. Beta diversity analysis demonstrated that the treatment of high hydrostatic pressure affected the microbial composition in donkey milk significantly. The potential probiotic Enterococcus casseliflavus isolated from the untreated donkey milk has a good acidifying ability. This study revealed the effects of high hydrostatic pressure treatment on the microbial composition in donkey milk, exhibiting its practical industrial application and the potential use of biological resources in the future.