








-
The family Stachybotriaceae in the order Hypocreales has been created by Crous et al.[1] to include the genera Myrothecium (Myr.), Peethambara (Pe.), and Stachybotrys (St.). These genera contain approximately 210 species (
www.mycobank.org ;www.indexfungorum.org ). The majority of these fungi are saprobic or toxic to plants and animals, and some are even detrimental to people. Asexual morphs with mononematous to sporodochial to synnematous conidiomata, frequently with phialidic conidiogenous cells that form 0-1-septate conidia in slimy or dry masses ranging from dark green to black, differentiate them[2].The ongoing study in the field of environmental mycology at the New Valley Governorate, Egypt is an expansion of mycological surveys carried out in the last three decades[3−8]. Despite the fact that the New Valley Governorate accounts for around 44% of Egypt's land area, mycological research in such settings are extremely rare and should be increased in the future to uncover more fascinating microorganisms. Numerous debate papers have been written throughout the years on the estimated numbers of fungi. The number of fungal species has long been a topic of discussion among mycologists since it has significant implications for conservation strategies. The number of fungi, which comprise visible mushrooms and unseen microorganisms, is unknown, and the estimates that have been generated so far are dubious. Despite their abundance and important role in the genetic diversity of life, fungi are many[9].
The 1.5 million figure is fully explained and justified by Hawksworth[10]. This was based on the estimate that there are six different taxa for each plant species. Numerous calculations have been made after the publication of this significant research, with the most current estimate being 11.7–13.2 million[11]. Mycologists, in particular, must concentrate on the finding of new or existing species in the many habitats in which they live[12]. As a result, the current work provides insight into the morphological categorization and phylogenetic investigation of four species isolated from reclaimed desert soil samples in the New Valley Governorate and published for the first time in Egypt.
-
New Valley Governorate is located in the Western Desert and is bounded on the east by the Governorates of Minya, Assiut, Sohag, Qena, and Aswan, on the west by the Egyptian border with Libya, on the north by the Governorates of Matrouh and the Coastal Oasis of the 6th October City, and on the south by the Egyptian border with Sudan. It covers 44,0098 Km2, or roughly 44% of Egypt's overall land and 66% of Egypt's southern territory[5].
Fungal isolation and maintenance
-
In this study, fungi were isolated from soil samples taken from El-Kharga city in Egypt's New Valley Governorate. The collected samples were maintained in sterile plastic bags and swiftly delivered to the laboratory of the Assiut University Mycological Centre (AUMC) for fungal isolation. The dilution plate method[13] was used to isolate the fungi. Suitable dilutions of the soil solution were added to Petri plates before being filled with Czapek's Dox agar (CzA)[14]. After that, the cultures were incubated at 25 °C for 2 weeks. To acquire pure cultures of the fungi, the developed colonies were purified on CzA using the single spore isolation method[15]. According to Al-Bedak et al.[16], the acquired pure cultures were subsequently maintained at 4 °C on Cz slants along with cotton balls. The Assiut University Mycological Centre's culture collection contains the four intriguing strains identified in this study as Paramyrothecium sp. AUMC 14059, Paramyrothecium sp. AUMC 14060, Striaticonidium brachysporum AUMC 13377, and Metarhizium pingshaense AUMC 14353. The four strains' internal transcribed spacer (ITS) sequences were uploaded to GenBank as MT940584, MT940585, MT940586, and MT940587, respectively.
Morphological studies
-
Spore suspension obtained from 7-day-old cultures was used for inoculation. Plates were inoculated in a three-point pattern using an inoculum size of 1 μl/spot. Microscopic features on potato dextrose agar (PDA)[17] were examined after 14 or 21 d of incubation at 25 °C.
Molecular identification
DNA isolation, PCR and sequencing
-
DNA extraction was carried out according to Moubasher et al.[18]. PCR reaction was performed according to protocol of Al-Bedak et al.[19] and Al-Bedak & Moubasher[20]. ITS1 and ITS4 universal primers[21] were used for amplification of ITS region, at SolGent Company using SolGent EF-Taq (SolGent Co. LTD, Yuseong-Gu, Daejeon, South Korea).
Phylogenetic analysis
-
The ITS dataset contained 22 species, four of which were obtained in the current study for Paramyrothecium (two sequences), Striaticonidium, and Metarhizium (one sequence each), and 17 sequences for the closest sequences of Metarhizium, Paramyrothecium, and Striaticonidium strains including the available type species. Sequence of Stachybotrys limonispora CBS 128809 was used as an outgroup. This study's DNA sequences were assembled using the DNASTAR computer software (version 5.05), and their assembled sequences were aligned with those acquired from GenBank using MAFFT[22]. BMGE was used to fix up alignment gaps and parsimony uninformative characters[23]. MEGA X (version 10.2.26)[24] was used to conduct maximum-likelihood (ML) and maximum-parsimony (MP) phylogenetic analyses, evaluating the robustness of the most parsimonious trees using 1000 bootstrap replications[25]. The best optimal model of nucleotide substitution for the ML analyses was determined using Smart Model Selection (SMS) version 1.8.1[26]. The phylogenetic tree was drawn and visualized using MEGA X version 10.2.26[24], and edited using Microsoft Power Point (2016).
-
Paramyrothecium sp. AUMC 14059: The megablast search of NCBIs GenBank nucleotide database using ITS sequence revealed that, the closest hits are Myrothecium sp. SWG2 [(GenBank accession number MW940740; identities = 592/594 (99.66%); Gaps = 1/594 (0%)] and Myrothecium roridum F04 [(GenBank accession number HQ839773; identities = 591/594 (99.49%); Gaps = 1/594 (0%)]. When compared to the type species, the closest similar are Paramyrothecium humicola CBS 127295 [(GenBank accession number MH864508; identities = 579/582 (99.48%); Gaps = 1/582 (0%)] and Paramyrothecium parvum CBS 257.35 [(GenBank accession number MH855673; identities = 585/592 (98.82%); Gaps = 3/592 (0%)].
Paramyrothecium sp. AUMC 14060: A megablast search of NCBIs GenBank nucleotide database using ITS sequence showed that, the closest matches are Myrothecium sp. SWG2 [(GenBank accession number MW940740; identities = 592/594 (99.66%); Gaps = 1/594 (0%)] and Myrothecium roridum F04 [(GenBank accession number HQ839773; identities = 591/594 (99.49%); Gaps = 0/581 (0%)]. When compared to the type species, Paramyrothecium humicola CBS 127295 [(GenBank accession number MH864508; identities = 579/581 (99.66%); Gaps = 1/582 (0%)] is the most similar along with Paramyrothecium parvum CBS 257.35 [(GenBank accession number MH855673; identities = 585/592 (98.82%); Gaps = 3/592 (0%)].
Striaticonidium sp. AUMC 13377: Using ITS sequence in a megablast search of NCBIs GenBank nucleotide database showed that, the closest hits are Myrothecium sp. CGMCC 3.3665 and Myrothecium cinctum strain ATCC 32918 [(GenBank accession number FJ235086 and DQ135998; identities = 558/560 (99.64%); Gaps = 1/560 (0%)]. When compared to the type species, Striaticonidium brachysporum CBS 513.71 [(GenBank accession number NR_154429; identities = 545/547 (99.63%); Gaps = 1/547 (0%)] is the most similar.
Metarhizium sp. AUMC 14353: Based on a megablast search of NCBIs GenBank nucleotide database using ITS sequence, the closest hits are Metarhizium anisopliae CBS 218.29 [(GenBank accession number MH855048; identities = 540/542 (99.63%); Gaps = 0/542 (0%)]. When compared to the type species, Metarhizium robertsii ARSEF 2575 [(GenBank accession number NR_132011; identities = 537/542 (99.08%); Gaps = 2/542 (0%)] and Metarhizium brunneum CBS 316.51 [(GenBank accession number MH856876; identities = 536/544 (98.53%); Gaps = 3/544 (0%)] are the closest hits.
Phylogenetic analysis
-
The final ITS analysis comprised 22 sequences. The maximum parsimony analysis produced 571 characters, of which 451 characters could be aligned correctly (no gaps, no N), 125 variable characters which were parsimony-uninformative, and 96 characters were counted as parsimony informative. Kimura 2-parameter using a discrete Gamma distribution (K2+G) was the best model for nucleotide substitution. The dataset for maximum parsimony yielded one tree with a tree length of 239 steps. The best scoring ML tree out of eight most parsimonious trees with the final ML optimization likelihood value of −1937.36, consistency index of 0.857895, retention index of 0.965823, and composite index of 0.828574 is presented in Fig. 1.
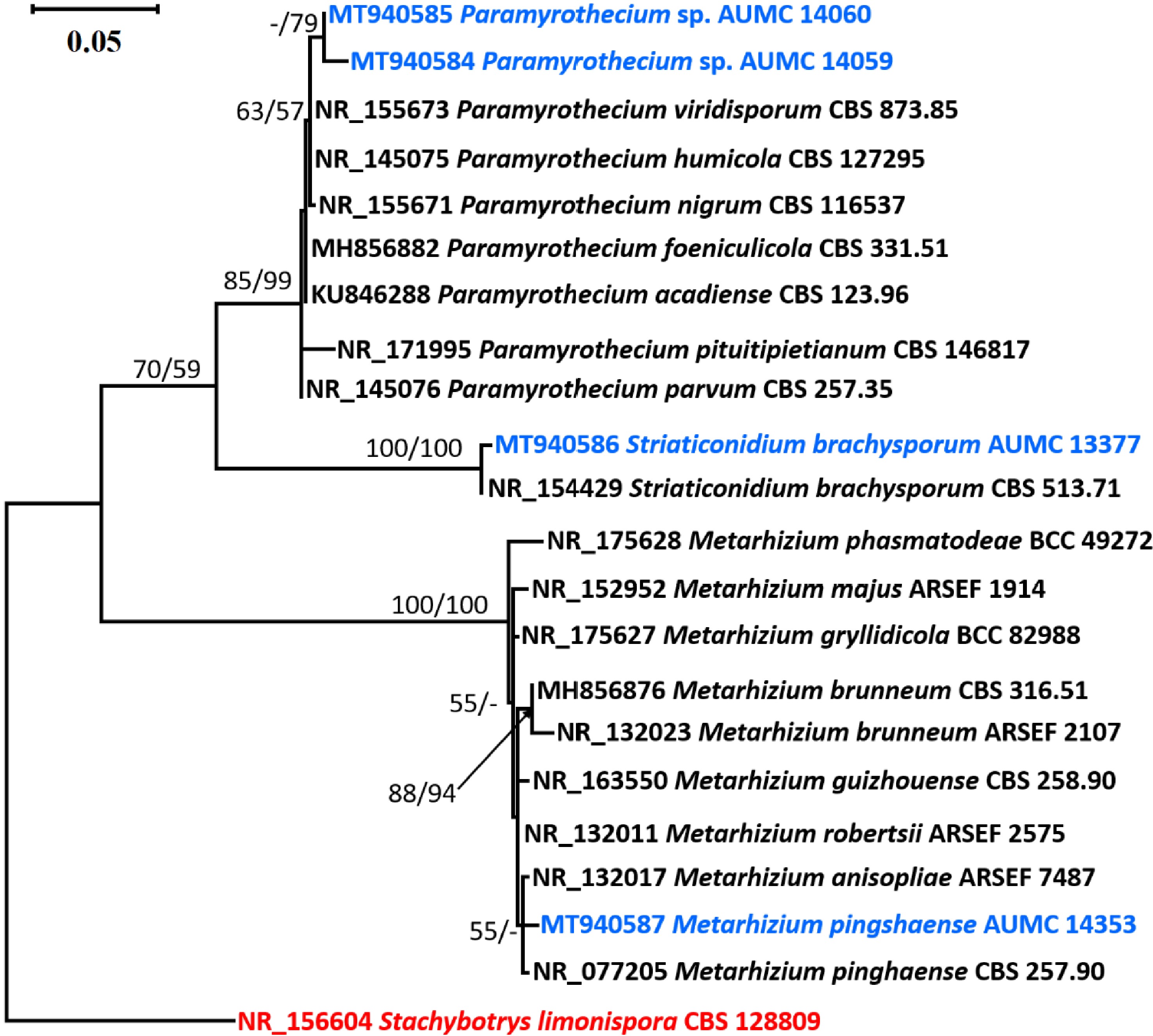
Figure 1.
Phylogenetic tree generated from MP analysis based on ITS sequence data of Paramyrothecium sp. AUMC 14059, Paramyrothecium sp. AUMC 14060, Striaticonidium brachysporum AUMC 13377 and Metarhizium pingshaense AUMC 14353 (in blue color) compared to other related ITS sequences in GenBank. Bootstraps (1000 replications) for ML/MP ≥ 50% are indicated above/below the respective nodes. The tree is rooted to Stachybotrys limonispora CBS 128809 as out group (in red color).
Phylogeny
-
The species clade of Paramyrothecium was supported by the ITS rDNA tree (85% ML/99% MP). The Paramyrothecium clade had one branch separating the Paramyrothecium spp. strains AUMC 14059 and AUMC 14060. However, they are distinct from one another morphologically. Since additional gene sequencing is required for a correct identification, they are thus presented here as potentially new species. Striaticonidium brachysporum AUMC 13377 established a clade with the type species that was highly well supported (100% ML/100% MP). It shared the same morphological features as the type species. The type strains of Metarhizium anisopoliae and Metarhizium pingshaense were found to be associated with Metarhizium pingshaense AUMC 14353 in a poorly supported clade (55% ML/< 50% MP). Metarhizium pingshaense was the classification given to it since it had the same morphological traits as the type species.
Brief descriptions of the obtained strains
Paramyrothecium sp.
AUMC 14059 -
Culture characteristics: Colonies on Cz attaining a diameter of 55−65 mm after 21 d at 25 °C. Conidiomata sporodochial, oval or irregular, 85−550 μm diam. Conidiophores arising from the basal stroma, consisting of a stipe and a penicillately branched conidiogenous phialides. Stipes unbranched, hyaline, septate, smooth, 12−22 × 2−3 μm. Conidiogenous cells phialidic, cylindrical, hyaline becoming darker at the apex, smooth, straight to slightly curved, 8–13 × 1−3 μm. Conidia aseptate, hyaline, smooth, ellipsoidal to cylindrical, 6−7 × 1−2 μm, rounded at both ends, forming an olivaceous green slimy mass (Fig. 2).
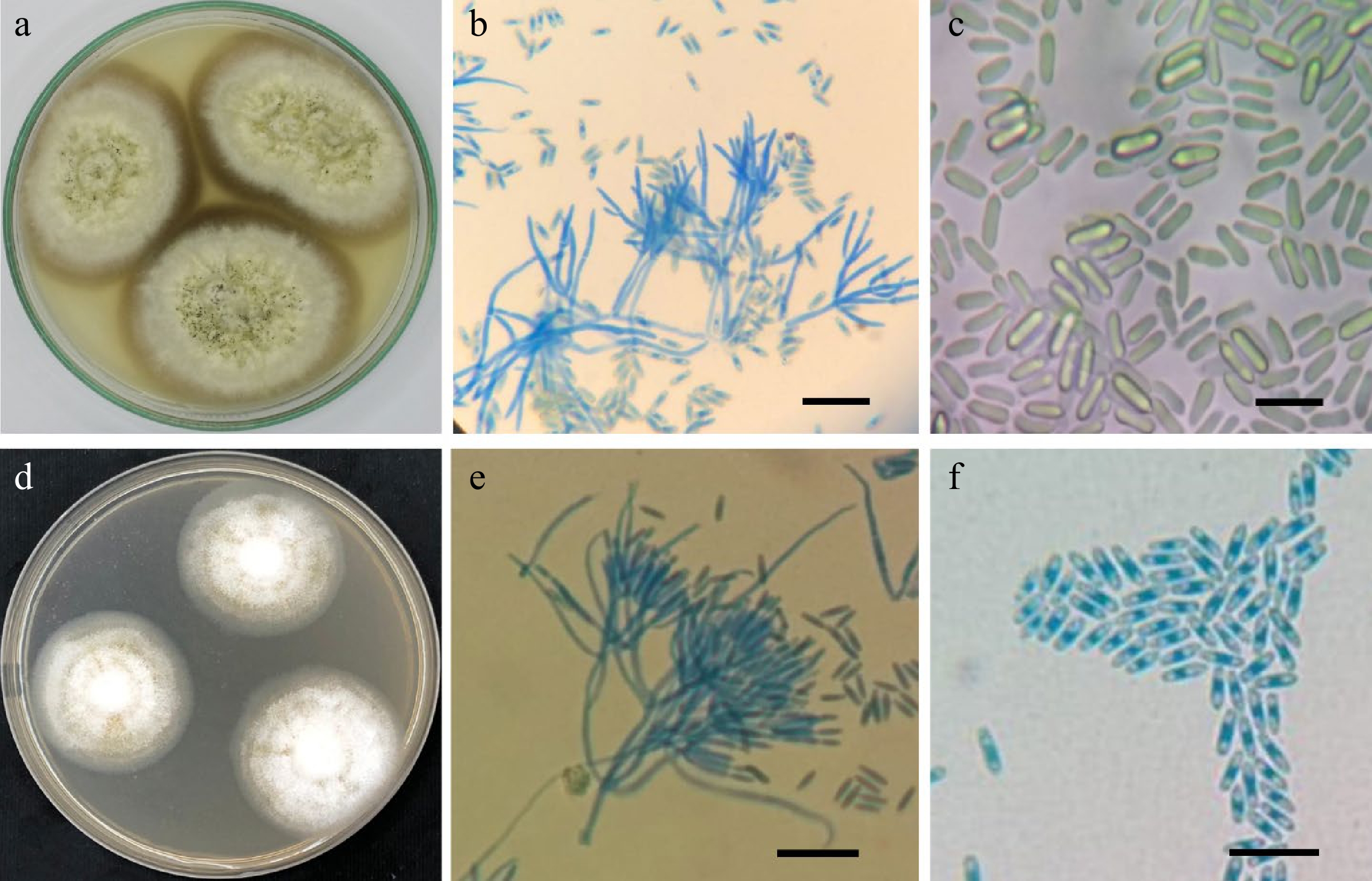
Figure 2.
(a−c), Paramyrothecium sp.
AUMC 14059: (a) 21-day-old colonies on Cz at 25 °C; (b) conidiophores bearing penicillate branched phialides; (c) cylindrical conidia rounded at both ends. (d−f) Paramyrothecium sp. AUMC 14060: (d) 14-day-old colonies on Cz at 25 °C with abundant white aerial mycelium; (e) conidiophores bearing penicillate branched phialides; (f) conidia (scale bars = 10 µm). Habitat and known distribution: USA, Kansas, from soil[2], and Egypt, New Valley, from soil (this study).
Paramyrothecium sp.
AUMC 14060 -
Culture characteristics: Colonies on Cz attaining 40−42 mm in diameter after 14 d at 25 °C, abundant white aerial mycelium with sporodochia forming on the surface of the medium in concentric rings, covered by slimy mouse grey to olivaceous green conidial masses. Conidiomata sporodochial, superficial, scattered or gregarious, oval or irregular. Conidiophores consisting of a stipe and a penicillately branched phialides. Stipes unbranched, hyaline, septate, smooth, 15−35 × 2−3 µm, terminating in a whorl of 3−6 conidiogenous cells. Conidiogenous cells phialidic, cylindrical to elongate doliiform, hyaline, smooth, straight to slightly curved, 6−12 × 3−5 µm. Conidia aseptate, hyaline, smooth, ellipsoidal to cylindrical, 3−5 × 2 µm, rounded at both ends (Fig. 2).
Habitat and known distribution: Turkey, from soil; Usak-Sükraniye, from soil; USA, Wyoming, Rock Springs, from soil in sagebrush grassland, from soil in bunchgrass rhizosphere, Grand Teton National Park, from soil in sagebrush grasslands, near Dubois, from soil in desert grassland, north and west of Hanna, from soil in strip mine area[2], and Egypt, New Valley, from soil (this study).
Striaticonidium brachysporum AUMC 13377
-
Culture characteristics: Colonies on Cz reaching 55−60 mm in diameter after 21 d at 25 °C. Conidiomata sporodochial, stromatic, superficial, scattered or gregarious, oval to elongate or irregular in outline, 55−220 μm diam. Conidiophores arising from the basal stroma, consisting of a stipe and a penicillately branched conidiogenous cells. Stipes unbranched, hyaline, septate becoming constricted at the septum, smooth, 18−33 × 2−4 μm. Conidiogenous cells phialidic, cylindrical, hyaline, smooth, slightly verrucose, straight, flexuous or bent at the upper third, 5−22 × 2−3 μm. Conidia aseptate, longitudinally striate, olivaceous green to brown, fusiform to ellipsoidal, (6–) 6.5−7.5 (–9.5) × (2–) 2.5−3.5 (–5) μm, with a distinct apical hilum (Fig. 3).
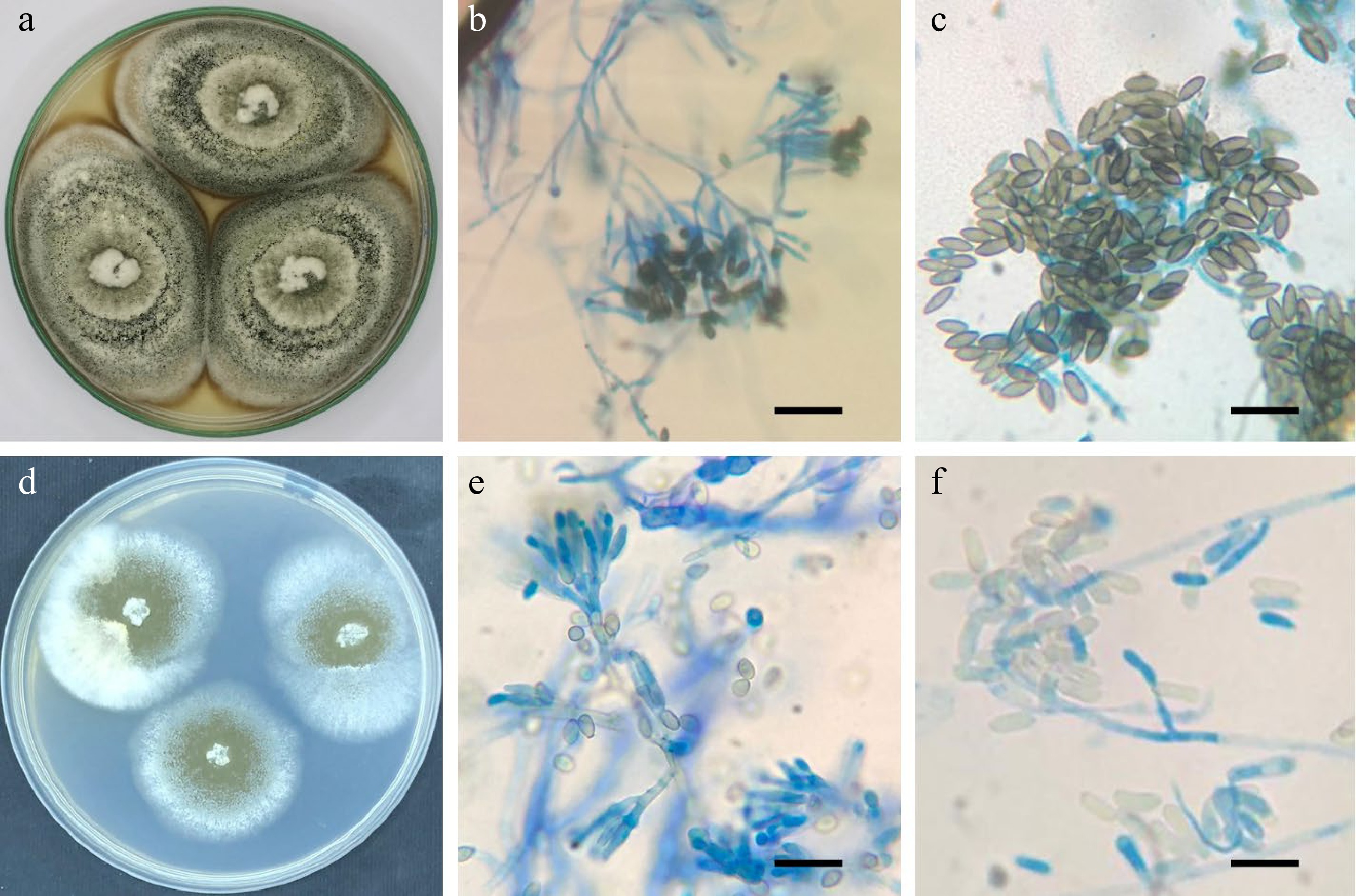
Figure 3.
(a−c), Striaticonidium brachysporum AUMC 13377: (a) 21-day-old colonies on Cz at 25 °C; (b) conidiophores bearing penicillate branched phialides; (c) longitudinally striate, fusiform to ellipsoidal conidia. (d−f) Metarhizium pingshaense AUMC 14353: (d) 14-day-old colonies on Cz at 25 °C with olive appearance; (e) conidiophores bearing penicillate, branched, cylindrical phialides; (f) ellipsoidal to cylindrical conidia (scale bars = 10 µm).
Habitat and known distribution: Iran, from dune sand; South Africa, from leaf litter of Acacia karroo; Ukraine, Kiev, from soil; USA, Kansas, from soil in tallgrass prairie, and Wisconsin, from soil in grassland prairie[2], and Egypt, New Valley from soil (this study).
Metarhizium pingshaense AUMC 14353
-
Culture characteristics: Colonies on Cz attaining a diameter of 36−48 mm after 14 d at 25 °C. Mycelium closely appressed, flat, white at the margins, turning to olive while sporulating. Conidiophores arising from aerial mycelium, erect, smooth-walled. Phialides smooth-walled, Phialides cylindrical, 7–17 × 2.5–3.5 μm. Conidia smooth-walled, ellipsoid, 6–8 × 2.5–3.5 μm (Fig. 3).
Habitat: Coleoptera: Cerambycidae, Chrysomelidae, Curculionidae, Scarabaeidae; Diptera: Stratiomyidae; Hemiptera: Cicadidae, Cydnidae, Delphacidae, Pentatomidae, Pseudococcidae; Isoptera: Kalotermitidae, Rhinotermitidae, Termitidae; Lepidoptera: Noctuidae; Orthoptera: Gryllidae[27,28] and from soil (this study).
Known distribution: Australia, Brazil, China, Egypt (this study), India, Indonesia, Japan, Papua New Guinea, Philippines, Solomon Islands, Thailand.
-
Four noteworthy fungal isolates from three different species have been reported in this study. Two of these strains were uncovered in Egypt for the first time, while the other two may be brand-new species. The four strains were identified as Paramyrothecium spp. (two strains), Striaticonidium brachysporum, and Metarhizium pingshaense after ITS region of the isolates was sequenced and matched to the closest match strains in GenBank. Fungi are so common and they have been found in a wide variety of environments. For instance, they can live inside plant tissues as endophytes[5,6,8,29,30], be pathogens of economically significant crops[31−33], perform the function of decomposers and play a significant role in the cycling of nutrients[34−36]. Due to the relevance of some fungi to humans, some communities have received much research, whilst others have received less attention[37].
There are many understudied ecosystems that support a wide variety of species, and if they are thoroughly investigated, new species might be found. Karst fungi, for instance, caves, forests (especially unpolluted rainforests), extreme environments, volcanoes, mountains, deserts, freshwater aquatic systems, lakes, grasslands, indoor environments, and many others are additional, less-studied habitats where fungi could be abundant and reveal a number of novel species discoveries[9,18−20,38−40]. Therefore, studying understudied areas is likely to lead to the discovery of novel fungi in unexpectedly large quantities.
The generic name Paramyrothecium was introduced as a new genus by Lombard et al.[2]. The sporodochial conidiomata are stromatic, superficial, cupulate, dispersed or gregarious, oval or irregular in appearance, with or without a white setose border enclosing an olivaceous green to dark green slimy mass of conidia. Conidiophores are penicillately branching, hyaline, and smooth. Phialides hyaline, occasionally darker at the apex, smooth to mildly verrucose, cylindrical to subcylindrical, narrowing at the tip, with prominent collarettes. Conidia are aseptate to 1-septate, cylindrical to ellipsoidal to obovoid, straight to bending, hyaline to pale green in colour, and smooth.
Striaticonidium was named for a group of Myrothecium-like fungi with striate conidia, which is supported by phylogenetic inference in this research. Tulloch[41] classified all Myrothecium species with striate conidia as Myrothecium cinctum due to similarities in conidial size and conidiomata marginal ornamentation. Later, Nag Raj[42] synonymized Myrothecium cinctum with Hymenopsis ellipsospora, a species in a genus in need of revision. Lombard et al.[2] kept the name 'cinctum' for Striaticonidium type species because it precedes 'ellipsospora'[43] and designated an epitype for this species.
Metarhizium is a common genus of entomopathogenic fungus with a variety of asexual reproductive morphologies and life-cycle stages[44−46]. It comprises soil saprophytes of the Hyphomycetes family, which are the most extensively isolated fungus from agricultural or natural soil and are found in both tropical and temperate climates[47,48]. Metarhizium spp. are well-recognized for their deadly arthropod pathogenic ability as agents of 'green muscardine' disease[49], affecting more than 200 insects and arthropod species, so that Metarhizium spp. are used as mycopesticides[28,48,50].
Tulloch[41] reviewed the genus and narrowed it to just two species, M. anisopliae and M. flavoviride, but with two varieties, M. anisopliae var. anisopliae with small conidia and M. anisopliae var. majus with large conidia. Rombach et al.[51] took into consideration the forms of conidia and phialides as well as the conidial structure of prismatic columns and the presence or lack of subhymenial zones as diagnostic criteria for species delimitation. They've even included M. flavoviride var. minus for Asian isolates with shorter conidia and revived M. album as a separate species. China and Japan have recorded M. pingshaense, M. cylindrosporum, M. guizhouense[27,28] and M. taii with its sexual morph Cordyceps taii[52].
Data availability
-
All data related to this manuscript is incorporated in the manuscript only.
Ethics statement
-
This article does not contain any studies with human participants or animals performed by any of the authors.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Mazen MBH, Moharram AM, Hassan SHA, Abdel-Latif SM, Al-Bedak OA. 2022. Contribution to the soil-based Egyptian mycobiota in Hypocreales (Sordariomycetes) from Egypt. Studies in Fungi 7:7 doi: 10.48130/SIF-2022-0007 

Contribution to the soil-based Egyptian mycobiota in Hypocreales (Sordariomycetes) from Egypt
- Received: 07 January 2022
- Accepted: 24 August 2022
- Published online: 09 September 2022
Abstract: During soil fungal surveys in Egypt's New Valley Governorate, four significant fungal isolates from three different species have been reported. Two of these strains were uncovered in Egypt for the first time, while the other two may be potential new species. The four strains were identified as being Paramyrothecium spp. (two strains), Striaticonidium brachysporum, and Metarhizium pingshaense after the internal transcribed spacer (ITS) region of the isolates was sequenced and matched to the closest match strains in GenBank. The strains in this investigation are being documented for the first time in Egypt. The four strains are briefly described and microscopically displayed in the article.
-
Key words:
- ITS /
- Metarhizium /
- New Valley /
- Paramyrothecium /
- Phylogeny /
- Soil /
- Striaticonidium.