









-
Fruit ripening and softening are key traits for many fleshy fruit[1,2]. During ripening, fruit will gradually soften and this is largely the result of fruit cell wall degradation[3−5]. Softening can improve the edible quality of fruit but also significantly reduces fruit resistance to pathogenic microorganisms and mechanical damage during storage and transportation, which can reduce the storage and shelf life[2] and lead to fruit deterioration and loss. For these reasons, research on the mechanism of fruit softening and control technology is a hot scientific topic in postharvest biology of fruit and vegetables. Although great progress in fruit softening has been made over the last 30 years, there are still many unsolved problems that need to be addressed.
Cell wall remodeling plays a major role in the texture changes in fleshy fruit and involves the coordinated expression of a large number of genes[6]. Tomato is a model material to study fruit softening[5, 7]. Fruit softening involves disassembly of polysaccharide-rich cell walls, a reduction in cell-to-cell adhesion and changes in cuticle properties that affect water loss[8, 9]. The precise mechanism of softening has been the subject of decades of research but has remained elusive. Enzymes such as polygalacturonase (PG)[10], pectin methyl esterase (PME)[11], β-galactanase (β-Gase)[12−14], expansin (EXP1)[15,16] and PL (Pectate lyase)[17] are highly expressed during the ripening process and all have been investigated as candidates for promoting changes in texture[5]. However, silencing of the gene encoding the major ripening-associated PG isozyme yielded only minimal improvements in slowing the rate of fruit softening[18−20]. Subsequent research targeted silencing of other ripening-associated pectin metabolic enzymes, including PME[20−22] and galactanase (β-Gase)[23], also only a minor effect on softening was demonstrated[4, 7]. The role of expansins in the control of over ripening and deterioration has also been investigated. In tomato, expression of expansin1 (EXP1) was shown to contribute to fruit softening[3]. Tomato pectate lyase (PL) gene is also believed to make a major contribution to fruit softening and silencing expression of tomato PL alters texture without affecting other aspects of ripening[7, 14, 24, 25]. Silencing of a strawberry gene encoding a PL also was shown to reduce fruit softening[26, 27].
The overall fruit softening process occurs in the final stages of fruit development and ripening, and is regulated by a series of transcription factors (TFs)[28, 29]. Softening, together with other aspects of ripening, is inhibit in several naturally-occurring tomato mutants including rin (ripening inhibitor)[30−33], nor (non-ripening)[34] and cnr (colorless non-ripening)[35−38], and have been considered to be ideal material for studying the transcriptional regulation of fruit softening. The knockout (KO) RIN mutant fruit generated by CRISPR/Cas9 technology showed accelerated cell wall degradation, suggesting that, contrary to the conventional view, RIN represses over-ripening in addition to facilitating ripening[39]. The softening of tomato cnr fruit was also inhibited[37]. Softening of CR-NOR mutant fruit was inhibited by down-regulating cell wall degrading enzymes gene including SlPG2a, SlPME2, SlTBG4, SlPL, SlCEL2, SlCEL8 and SlEXP1[40]. Recently, however, the role of these TFs has been re-evaluated. Although they are still regarded as important ripening TFs, their phenotypes have been found to result from gain-of-function mutations[31,32, 40−43]. Additional NAC TFs have also been shown to be involved in ripening regulation. NOR-like1 was highly expressed in tomato fruit, its mutation delayed fruit ripening, and reduced the expression levels of cell wall metabolic genes such as SlPG2a, SlPL, SlCEL2, and SlEXP1[44]. The overexpression of the SlNAC1 gene in tomato fruit resulted in the decrease in fruit firmness and peel thickness, which affected fruit softening[45] and the firmness of RNAi-SlNAC1 fruit was significantly higher than that of wild-type fruit[46]. SlAP2a belongs to AP2/ERF family transcription factor. In RNAi-SlAP2a tomato fruit, the expression of PG, CEL1, MAN3, MANS2 and other genes encoding cell wall degrading enzymes were up-regulated[47]. There is still a long way to go to excavate other transcription factors involved in regulating fruit softening and construct the transcriptional regulation network of fruit softening.
Another NAC gene, SlNAC4, has been reported to be a positive regulator of fruit ripening and carotenoid accumulation in tomato fruit[48], and participates in plant stress response[49]. Here, we use CRISPR/Cas9 technology to obtain SlNAC4-KO plants, and also transgenic plants overexpressing SlNAC4 under the control of the CaMV35S constitutive promoter. The results showed that SlNAC4 was not only a positive regulator of fruit ripening but is also involved in the regulation of fruit softening. RNA-seq, EMSA and DLR techniques were used to show that SlNAC4 can directly regulate the expression of cell wall metabolic genes SlEXP1 and SlCEL2, which enriched our understanding of the function of the SlNAC4 gene and provided a new perspective on the regulation of tomato fruit softening.
-
Wild type (WT) (Solanum lycopersicum cv. Ailsa Craig, abbreviated as AC) and mutant CR-SlNAC4 and OE-SlNAC4 lines were produced in the AC background and were grown in a standard greenhouse with natural light and regular feeding with supplementary fertilizer. Fertilization time was recorded at anthesis in order to determine subsequent fruit ripening stages, in combination with fruit color, fruit size and seed development. The onset of fruit ripening was determined as the breaker stage (first sign of color change). Fruit samples of WT and the CR-SlNAC4 and OE-SlNAC4 were harvested at different ripening stages (IMG: immature green, MG: mature green, BR: breaker, B+3: three days after breaker, B+6, B+9, B+12) for different experiments. The pericarp tissues were collected and frozen in liquid nitrogen immediately, then stored at −80°C until use.
Primers
-
Primers were designed using the online program Primer3 (v. 0.4.0), the Primer Premier 5 software or the CE Design v1.03 software. All primers used in qRT-PCR are shown in Supplemental Table S1, primers used in vector construction are listed in Supplemental Table S2.
Subcellular localization
-
The coding sequence of SlNAC4 (without the stop codon) was fused to GFP driven by the CaMV 35S promoter to generate SlNAC4-GFP fusion protein, at the same time, a nuclear marker protein MADS-RIN was fused to mCherry to generate RIN-mCherry. The two fusion proteins were cloned into the pEAQ vector[50] and the resulting constructs were transformed into A. tumefaciens GV3101 separately. Agrobacteria containing each construct were grown in Luria-Bertani medium at 28 °C for 16 h, and then 20% of each culture was transferred into a new Luria-Bertani medium containing 10 mM MES[2-(N-Morpholino) ethanesulfonic Acid] pH 5.6, 10 µM AS(Acetyl eugenone) for another 8 h. Subsequently, each agrobacterium was pelleted and diluted to OD600 = 0.5 with infiltration medium (double distilled water containing 10 mM MES pH 5.6, 10 mM MgCl2 and 100 µM AS). The same volume of SlNAC4:GFP and RIN:mCherry were mixed and infiltrated into four-week-old tobacco (Nicotiana benthamiana) leaves for transient co-expression. The free GFP and mCherry were co-expressed as the controls. Two days after infiltration, tobacco leaves were directly observed and images captured under a laser confocal microscope (Leica, Germany).
RNA isolation and quantitative real-time PCR
-
Total RNA from tomato vegetative organs (root, stem, leaf) and reproductive organs (flowers and fruit from different ripening stages) were isolated using the RNeasy Mini Kit (Qiagen, Germany) according to the manufacturer's protocol. DNaseI (Qiagen, Germany) digestion was added to remove genomic DNA. 1.5% (m/v) agar gel electrophoresis and NanoDrop 1000 spectrophotometry (Thermo, USA) were used to assess RNA quality and final concentration. cDNA was synthesized from 2 μg total RNA using TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen, China) with Oligo(dT)18 primer. Quantitative real-time PCR (qRT-PCR) was performed with SYBR Green PCR Master Mix (TransGen, China) in a 20 μl total sample volume using the CFX96 Real-Time PCR System (Bio-Rad, USA). Three independent biological replicates were analyzed per sample. Relative abundance of specific mRNA levels was confirmed using the
$2^{-\Delta\Delta} $ Cᴛ method and normalized by the housekeeping gene SlActin (Solyc03g078400).CRISPR/Cas9 and overexpression vectors construction and transformation
-
Four target sequences that were located specifically in the SlNAC4 sequence were selected using the CRISPR-P (http://cbi.hzau.edu.cn/crispr) online program[51]. The four sgRNAs with target sequences (target-sgRNAs) were amplified and inserted into the pYLCRISPR/Cas9Pubi-H binary vector by the Golden Gate ligation method as described previously[52]. Oligonucleotide primers used in pYLCRISPR-SlNAC4 vector construction are listed in Supplemental Table S2. For the SlNAC4 overexpression construct, the complete CDS (Coding DNA Sequence) fragment of SlNAC4 (without the stop codon) was amplified with cDNA from tomato fruit at the breaker stage. The 3×HA-tag was amplified and cloned into the pCAMBIA-1300-221 destination vector with the SlNAC4 CDS fragment using Mut Express II Fast Mutagenesis Kit (Vazyme, China). The resulting two constructs were confirmed by sequencing and then transformed into Solanum lycopersicum cv. Ailsa Craig using the stable Agrobacterium tumefactions (GV3101)-mediated transformation method[40]. The transgenic tomato lines were selected by their hygromycin resistance.
DNA extraction and transgenic lines identification
-
Total genomic DNA was extracted from fresh tomato leaves of WT, CR-SlNAC4 and OE-SlNAC4 transgenic lines frozen in liquid nitrogen using a DNA secure Plant Kit (Tiangen, China). For CRISPR/Cas9 mutation analysis, DNA from CR-SlNAC4 lines was used as a template to amplify the desired gene fragments using primers flanking the target sites. The PCR products were sequenced to identify mutations. Primers used in this section are listed in Supplemental Table S3. For the SlNAC4 gene editing lines, the two most likely off-target sites of each target (Supplemental Table S4) were tested; the primers used for off-target analysis are listed in Supplemental Table S5. For overexpression analysis, the presence of the transgene was verified by amplifying the hygromycin resistance gene using DNA from OE-SlNAC4 lines as templates. DNA from WT was used as the negative control. Hygromycin-positive plants were used to detect the expression levels of SlNAC4 by qRT-PCR.
Firmness measurement
-
Fruit firmness examination was performed as described previously with minor revision[53]. Freshly harvested WT, CR-SlNAC4 and OE-SlNAC4 transgenic fruit at six ripening stages (MG, Br, B+3, B+6, B+9 and B+12) were analyzed using a TA.XT Plus texture analyzer (Stable Micro Systems, England) with a 5 cm diameter cylindrical stainless probe. An equatorial slice about 1 cm thick was removed from each fruit and placed on the test bench. The junction of outer and radial pericarp were compressed 2 mm at a test speed of 1 mm/s, visible vascular bundles, fissures and locular tissue were avoided. Each fruit was measured at two or three sites, and the average of the maximum force developed during the test was used as one biological replicate. At least 6 and up to 12 fruit per genotype and stage were used for each measurement.
TEM (Transmission electron microscope)
-
The pericarp samples from WT, slnac4#13 and OE-SlNAC4#2 fruit at the breaker stage were excised, fixed and performed as previously described, respectively[54]. The transmission electron microscope (TEM) observation was performed using a H-7650 transmission electron microscope (Hitachi, Japan), and the micrographs were taken with an 832 camera (Gatan, USA).
RNA-sequencing
-
The RNeasy Mini Kit (Qiagen, Germany) was used to extract total RNA from the fruit of WT, slnac4#13 and OE-SlNAC4#2 at 38 DPA. The genomic DNA was removed by DNase I (Qiagen, Germany) digestion, three biological replicates were made for each sample. Then the RNA samples were sent to Novogene (Novogene, China) for RNA quality inspection, RNA-sequencing libraries construction, and sequencing. At least 6 G of raw data were generated for each pair-end sequencing. TopHat software (version 2.0.14) was used to map the clean data to the tomato reference genome (version SL2.50, ftp://ftp.sgn.cornell.edu/tomato_genome) and the fragments were assigned to genes by feature counts and count programs. Differential gene expression between slnac4#13 mutant and WT or OE-SlNAC4#2 and WT were identified by DESeq2 Library, respectively, and the fold change was calculated by RPKMslnam1/RPKMWT and RPKMOE-SlNAM1/RPKMWT. Genes of |fold change| ≥ 2 and modified p-value < 0.05 were considered as differentially expressed genes (DEGs). DEGs which were common in both CRISPR and overexpression lines and with opposite regulatory patterns were screened for further analysis.
Protein expression and EMSA
-
The coding sequence (CDS) of SlNAC4 gene was amplified from the cDNA of tomato fruit. Then the product was inserted into the linearized pGEX-GST vector which was digested with BamH I and Xho I to produce recombinant SlNAC4 protein with a glutathione S-transferases (GST) tag using ClonExpress II One Step Cloning Kit (Vazyme, China). The N-GST-SlNAC4 fusion protein was expressed in Escherichia coli Rosetta (DE3) and induced with 0.5 mM IPTG for 6 h at 25 °C, further purified with Glutathione Sepharose 4B (GE Healthcare, Sweden) following the manufacturer's protocol. The oligonucleotide probes (shown in Supplemental Table S5) containing the NACRS (NAC recognition sequence) [TGA][ACG]CGT[GA][TA][40] regions derived from the promoters of SlEXP1 and SlCEL2 were synthesized (Sangon Biotech, China) and biotin-labeled using the Biotin 3' End DNA Labeling Kit (Thermo Scientific, USA). EMSA was carried out as previously described[55]. Both the unlabeled probes and the mutant probes were used for competition. GST protein alone were used as negative control.
Dual-luciferase reporter assay
-
Dual-luciferase reporter assay was performed to analyze the transcriptional activity of SlNAC4 with the promoters of SlEXP1 and SlCEL2. The CDS fragment of SlNAC4 was amplified and cloned into the CaMV35S promoter-drove pEAQ vector[56] to construct the effector vector pEAQ-SlNAC4. The 1 kb promoter regions of SlEXP1 and SlCEL2, which were reported in our previous study[44], were amplified and inserted into the linearized double-reporter vector pGreenII 0800-LUC[57] to construct pGreenII 0800- SlEXP1-LUC/pGreenII 0800- SlCEL2-LUC using ClonExpress II One Step Cloning Kit (Vazyme, China). Subsequently, the recombinant effector was transferred into A. tumefaciens EHA105, and the reporters were transferred into EHA105 (pSoup). Transient transfection assay was performed in 4-week-old tobacco leaves as described previously[58]. After 48 h of infiltration, LUC and REN luciferase activities were measured respectively using the dual-luciferase assay kit (Promega, USA) with the Luminoskan Ascent Microplate Luminometer (Thermo Fisher Scientific, USA) and the results were calculated by the ratio of LUC to REN. At least six biological replicates were performed for each combination.
Data Analysis
-
Data were analyzed by the ANOVA procedure of Statistical Analysis Systems (SAS) software, version 5 (SAS Institute, Cary, NC).
-
Genome analysis revealed that the SlNAC4 gene (Solyc11g017470.1) is 1314 bp long, with three exons and two introns, encodes the SlNAC4 protein of 291 amino acids. SlNAC4 is a typical NAC transcription factor, containing a conserved DNA-binding NAC domain which includes five subdomains (A, B, C, D and E) at its N-terminus (Fig. 1a). Most plant transcription factors are translated in the cytoplasm, then enter the nucleus under the guidance of their nuclear localization signals, and activate or inhibit transcription of target genes such as SlNOR-like1[44], NAC-NOR[40], and RIN[31], among others. In addition, some of membrane-bound transcription factors (MTFs) are anchored in membranes in a dormant state. MTFs were activated by external or internal stimuli, released from parent membranes and are transported to the nucleus[59, 60]. The membrane-associated transcription factor NAC089 controls ER-stress-induced programmed cell death in plants[61]; OsNTL3 encodes a NAC transcription factor with a predicted C-terminal transmembrane domain. GFP-OsNTL3 relocates from the plasma membrane to the nucleus in response to heat stress and ER stress inducers[62]; A membrane-associated NAC domain transcription factor XVP interacts with TDIF co-receptor and regulates vascular meristem activity[63]; A membrane-associated NAC transcription factor regulates salt-responsive flowering via FLOWERING LOCUS T in Arabidopsis[64].

Figure 1. Gene expression pattern and subcellular localization of SlNAC4. (a) Structure of the SlNAC4 protein. A−E represents the 5 subdomains of the conserved NAC domain. (b) Gene expression of SlNAC4 in different organs and fruit ripening stages of tomato. IMG, immature green; MG, mature green; Br, breaker; RR, red ripe. Bars represent ± SD of three independent replicates. SlActin gene was used as the internal control. (c) SlNAC4 co-localized with RIN (a marker protein of the nucleus) in nuclei. CaMV35S::SlNAC4-GFP represents SlNAC4 and GFP fusion protein, CaMV35S::RIN-mCherry represents MADS-RIN and mCherry fusion protein. CaMV35S::GFP co-expressed with CaMV35S::mCherry represent the control. Bars = 20 μm.
To determine the subcellular localization of SlNAC4, the SlNAC4-GFP was co-expressed with a nucleus marker protein RIN-mCherry in tobacco leaves. The green fluorescence from SlNAC4-GFP protein was co-localized with the red fluorescence from RIN-mCherry in the nucleus, indicating that SlNAC4 is localized in the nucleus (Fig. 1b).
To understand possible SlNAC4 gene functions, we measured the expression levels of the SlNAC4 genes in different tomato tissues (root, stem, leaf, flower) and fruit at different ripening stages (immature green, mature green, breaker, red ripe) by quantitative RT-PCR (qRT-PCR). The results showed that SlNAC4 was expressed in all tested tissues at different levels, but its expression was up-regulated during tomato fruit ripening (Fig. 1c). Some of other ripening-related NAC TFs also have high expression level during tomato fruit ripening such as NAC-NOR[40], SlNOR-like1[44], SlNAC1[45, 46], SNAC4 and SNAC9[65].
These results indicated that SlNAC4 is located in the nucleus, consistent with its role as a transcription factor, and its expression pattern is consistent with it playing a role in the fruit ripening process.
CRISPR/Cas9 gene editing of SlNAC4 inhibits while overexpression of SlNAC4 promotes tomato fruit ripening
-
Previous studies have shown that the ripening process of RNAi-SlNAC4 (RNA interference) tomato fruit was delayed and their carotenoid accumulation and ethylene production were also inhibited compared to wild type tomato fruit[48]. Although RNAi has been a powerful tool for studying gene function in the past decades, it also has some disadvantage, such as inability to silence target genes completely and possible functional redundancy of target gene family. As a result, the phenotypes of RNAi (RNA interference) plant may only partially reflect the functions of the targeted gene. Recently, clustered regularly interspaced short palindromic repeats genomic targeting systems (CRISPR/Cas9) can specifically mutate, delete or inactivate target genes to achieve 100% gene silencing. This can also solve the problem of redundancy of gene family members, and is superior to RNAi for studies on specific gene function[66−70].
To evaluate the role of SlNAC4 in tomato fruit ripening, we generated SlNAC4 knock-out mutant (CR-SlNAC4) using a CRISPR/Cas9 genome targeting system as well as stable SlNAC4 overexpressing plant (OE-SlNAC4) lines, which are ideal materials to study the SlNAC4 function during fruit ripening. Four single guide RNA (sgRNA) target sequences driven by LacZ-AtU3d, AtU3b, AtU6-1 and AtU6-29 promoters were designed to specifically target SlNAC4 genomic DNA (Fig. 2a). The four target-sgRNAs were amplified and cloned into the binary CRISPR/Cas9 vectors, pYLCRISPR/Cas9Pubi-H, driven by the maize ubiquitin promoter (Pubi) with the Golden Gate ligation method. The pCRISPR/Cas9-SlNAC4 vector was transformed into tomatoes by Agrobacterium-mediated leaf disc-transformation. A total of 24 independent T0 CR-NAC4 lines were obtained and of these lines 24 plants were genome edited. We selected two representative CR-SlNAC4 T0 lines for further analysis and two homozygous mutants were screened from the T1 generation of CR-SlNAC4, which were named slnac4#7 and slnac4#13. Gene editing details of the two mutants are shown in Fig. 2b. Off-target analysis was performed to determine whether any gene other than SlNAC4 had been edited in CR-SlNAC4 line. Two of the most likely off-target genes of each targeted site were tested and the results suggested that no off-target occurred in either the slnac4#7 or slnac4#13 lines (Supplemental Table S4). Transgenic tomato lines overexpressing SlNAC4 under the control of the cauliflower mosaic virus (CaMV) 35S promoter were also generated. A total of 17 independent overexpression (OE) lines were obtained in the T0 generation and we also selected two representative OE lines (OE-SlNAC4#2, OE-SlNAC4#3) for further research, and in the T1 generation, SlNAC4 fruit gene transcript levels in both lines were substantially higher than in WT fruit (Fig. 2c).
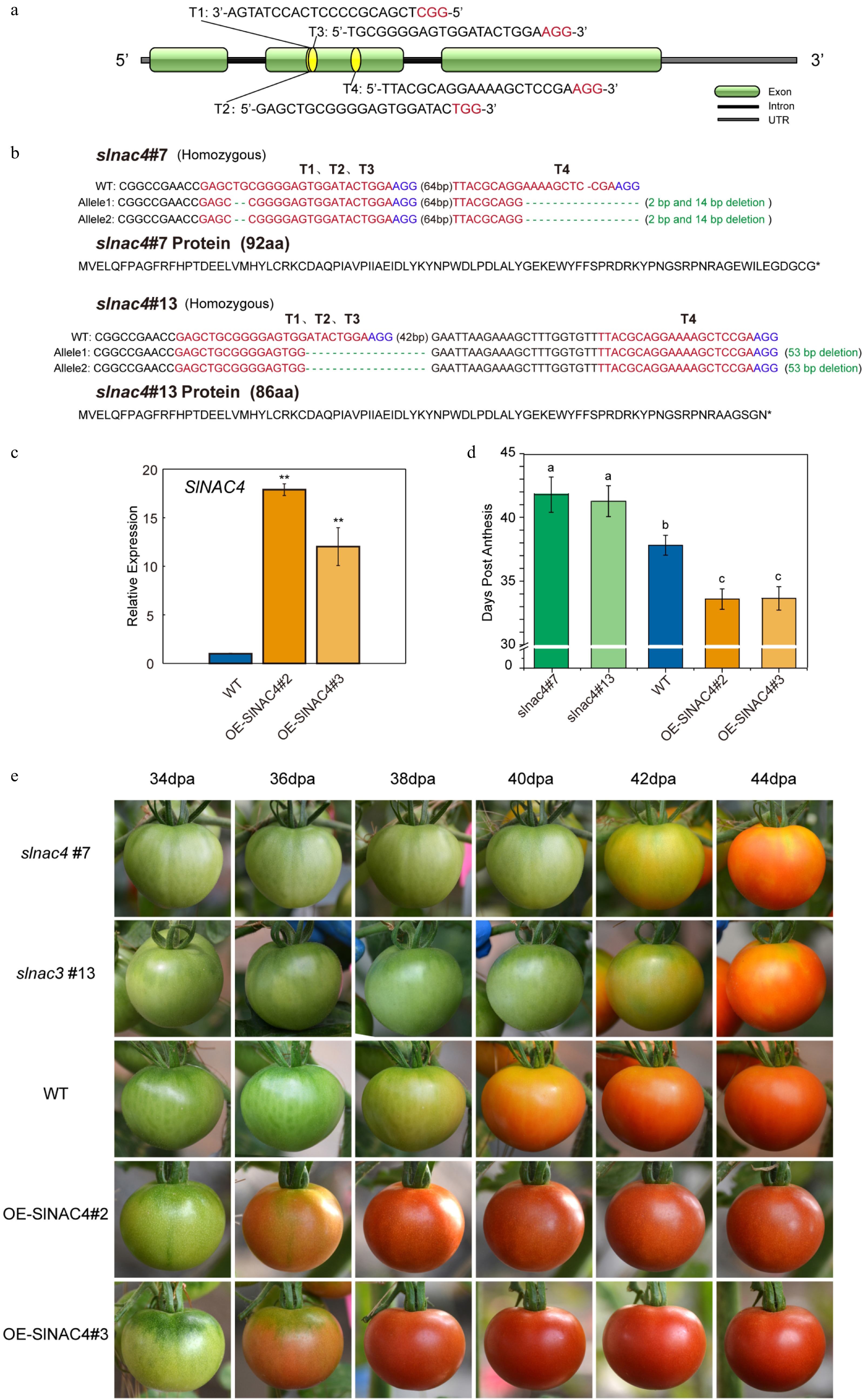
Figure 2. SlNAC4 CRISPR/Cas9 gene edited and overexpression lines, and their tomato fruit ripening phenotype. (a) Schematic illustration of four CRISPR/Cas9 target sites (T1, T2, T3, T4) in the SlNAC4 genomic sequence. (b) Gene editing details of slnac4#7 and slnac4#13 mutants. Blue letters represent the protospacer adjacent motif (PAM). (c) qRT-PCR analyses of SlNAC4 expression level in fruit of OE-SlNAC4#2 and OE-SlNAC4#3 overexpression lines compared with WT. Bars represent ± SD of three independent replicates. The SlActin gene was used as the internal control. (d) Days from anthesis to the fruit breaker stage of WT, slnac4#7, slnac4#13, OE-SlNAC4#2, and OE-SlNAC4#3. Lowercase letters represent significant differences (p < 0.05). (e) The tomato fruit ripening phenotype of WT, slnac4#7, slnac4#13, OE-SlNAC4#2, and OE-SlNAC4#3. dpa: days post anthesis.
To determine whether the CRISPR/Cas9-mediated mutagenesis and gene overexpression of SlNAC4 affected tomato fruit development or ripening, the flowering time was recorded and the fruit development and ripening stages, fruit size, color, and seed development were also recorded. Comparing the days from fertilization to fruit color break, it was found that this was 38 days for WT tomato, 34 days for OE lines (OE-SlNAC4#2, OE-SlNAC4#3) and 41 days for CR-SlNAC4 line (slnac4#7 and slnac4#13) (Fig. 3d). Although the time to the initiation of color change was different for the three types of fruit with different levels of SlNAC4 transcripts, all fruit eventually changed to be the same color as wild type fruit (Fig. 3e). It was concluded that SlNAC4 participated in the ripening process of tomato fruit as a positive TF, which is consistent with previous studies on the role of NAC4 in fruit ripening[48], and this transgenic material is suitable for investigating its role in fruit development and ripening. At present. NAC transcription factors have been found to be involved in fruit ripening in many species[71], such as tomato[44−46], strawberry[72], banana[73, 74], kiwi fruit[75], peach[76] and apple[77], and affect the formation of fruit quality, such as color, lycopene, chlorophyll degradation, anthocyanin synthesis, flavor, sugar and acid, and fruit softening[71−77], NAC regulate ethylene production to affect fruit ripening and quality[71−78].

Figure 3. Tomato fruit softening is affected by different levels of SlNAC4. (a) The firmness of WT, slnac4#7, slnac4#13, OE-SlNAC4#2, and OE-SlNAC4#3 tomato fruit at different ripening stages. SD (p < 0.05) are indicated with lowercase letters. MG, mature green; Br, breaker; B+n, n days after breaker. Error bars represent ± SD of at least six independent replicates. (b) Fruit cell wall structure of WT, slnac4#7, and OE-SlNAC4#2 at the breaker stage created by transmission electron microscopy. Bars = 0.5 μm.
SlNAC4 regulates tomato fruit softening
-
During the ripening of tomato fruit, we found that the fruit of OE-SlNAC4 lines were softer than WT fruit, while fruit of CR-SlNAC4 lines were firmer than that of WT at the same ripening stages. To confirm whether SlNAC4 may regulate tomato fruit softening, the firmness of freshly harvested fruit from WT, slnac4#7, slnac4#13, OE-SlNAC4#2 and OE-SlNAC4#3 at six ripening stages (MG, Br, B+3, B+6, B+9 and B+12) were harvested and analyzed using a texture analyser. As shown in Fig. 3a, the fruit firmness of OE-SlNAC4 lines was significantly lower than that of WT during all ripening stages from MG to B+12, while fruit of CR-SLNAC4 lines was harder than WT at the MG stage. In the early stages this may not be statistically significant but CR-SlNAC4 lines were harder than WT from the Br+3 stage to B+12 (Fig. 3a). Transmission electron microscopy (TEM) was carried out in order to get a more detailed picture of the cell walls of slnac4, OE-SlNAC4 lines and WT fruit. The results indicated that the microfilaments of the slnac4#7 mutant cell walls were arranged more closely compared with those in the WT cell walls, while more loose microfilaments were observed in cell walls of OE-SlNAC4#2 fruit (Fig. 3b), suggesting that cell wall degradation was inhibited in the fruit of slnac4 mutant, whereas in contrast it was promoted in OE-SlNAC4 fruit. Our previous results show that NAC-NOR[40], NOR-like1[44] also regulate fruit softening. However, the interaction between these NAC members regulating fruit softening has not been studied.
SlNAC4 directly targets the cell wall genes SlCEL2 and SlEXP1 to regulate tomato fruit softening
-
To analyze the roles of SlNAC4 in regulating fruit softening at the molecular level, strand-specific mRNA sequencing was applied to analyze the differential expression of genes in the pericarp of slnac4#7, OE-SlNAC4#2 and WT fruit at 38 dpa (Supplemental Data Set S1). Using a cutoff threshold of | Log2 (fold change) | > 1 and the modified p-value < 0.05 between transgenic and control samples, analysis of differentially expressed genes (DEGs) in slac4#7/WT and OE-SlNAC4#2/WT revealed that 600 genes were up-regulated while 821 genes were down-regulated in the slnac4#7 mutant compared to WT; 1,127 genes were up-regulated while 1,678 genes were down-regulated in OE-SlNAC4#2 compared to WT (Fig. 4a, 4b). When we overlapped the DEGs, which were differentially expressed in opposite patterns in slnac4#7 and OE-SlNAC4#2 lines, 215 DEGs were obtained (Fig. 4c), including two key softening-associated genes SlCEL2(Solyc09g010210) and SlEXP1(Solyc06g051800) (Fig. 4d).
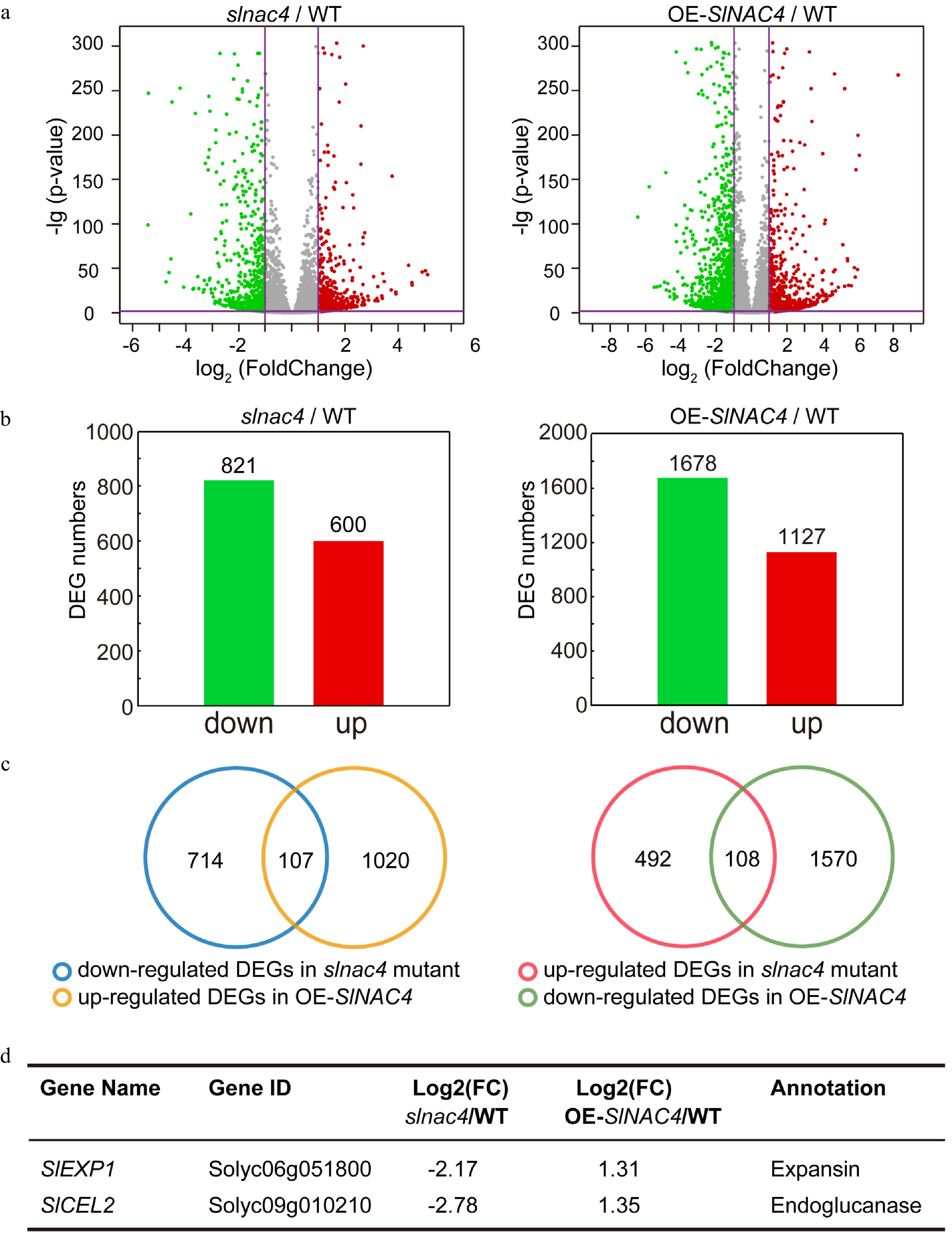
Figure 4. RNA-Sequence analyses of WT, slnac4#7, and OE-SlNAC4#2 fruit. (a) RNA-sequencing data visualized by volcano plots. Each point corresponds to a DEG. Red dots represent up-regulated genes, green dots represent down-regulating genes in slnac4#7 compared with WT or OE-SlNAC4#2 compared with WT. | Log2 (Fold change) | = 1 and p-value = 0.05 are marked with purple lines. (b) The number of up-regulated and down-regulated genes in slnac4#7/WT and OE-SlNAC4#2/WT fruit. (c) DEGs opposite regulated in slnac4#7 and OE-SlNAC4#2. (d) Two key tomato fruit softening related genes SlEXP1 and SlCEL2 are significantly down-regulated in slnac4 mutant and up-regulated in OE-SlNAC4 fruit.
In order to confirm the RNA-Seq results, we analyzed the expression level of SlCEL2 and SlEXP1 by qRT-PCR in slnac4#7 and slnac4#13, OE-SlNAC4#2, OE-SlNAC4#3 and WT fruit. The results were consistent with RNA-Seq data (Fig. 5a). Base on the results that SlCEL2 and SlEXP1 were down-regulated in slnac4 mutant, while upregulated in OE-SlNAC4 lines compared with WT, we asked whether the two genes were direct targets of SlNAC4. To test this hypothesis, we first checked whether the 1 Kb promoter regions of SlCEL2 and SlEXP1 harbor the characteristic NAC recognition sequences. The results show that NACRS sits were found in the 5' regulatory regions of SlCEL2 and SlEXP1, suggesting that they may both be direct targets of SlNAC4. In order to investigate whether SlNAC4 can directly bind to the promoter regions of SlCEL2 and SlEXP1 in vitro, an electrophoretic mobility shift assay (EMSA) was performed using the purified recombinant GST-SlNAC4 protein, with purified GST protein used as the control. As shown in Fig. 5b, strong SlNAC4 binding activity to the target gene promoter fragments was reduced greatly by incubating with excess concentrations of cold probes, but not with excess mutant probes. In the slnac4 mutant, the expression of SlCEL2 and SlEXP1 was significantly inhibited, suggesting that SlNAC4 positively regulated the transcription of both genes. To test this assumption, DLR assay was performed. The relative LUC/REN ratio in tobacco leaves co-transformed with CaMV35S-SlNAC4 and CaMV35S-REN/pSlCEL2-LUC or CaMV35S-REN/p SlEXP1-LUC was significantly higher than in leaves co-transformed with CaMV35S-Empty and CaMV35S-REN/p SlCEL2-LUC or CaMV35S-REN/pSlEXP1-LUC (Fig. 5c), indicating that SlNAC4 could activate the promoter activity of SlCEL2 and SlEXP1 in tobacco. Taken together, the results demonstrated that SlNAC4 is a transcriptional activator that positively regulates fruit softening in tomato fruit by directly targeting the promoter of SlCEL2 and SlEXP1. The model of SlNAC4 regulating tomato fruit softening is shown in Fig. 6.
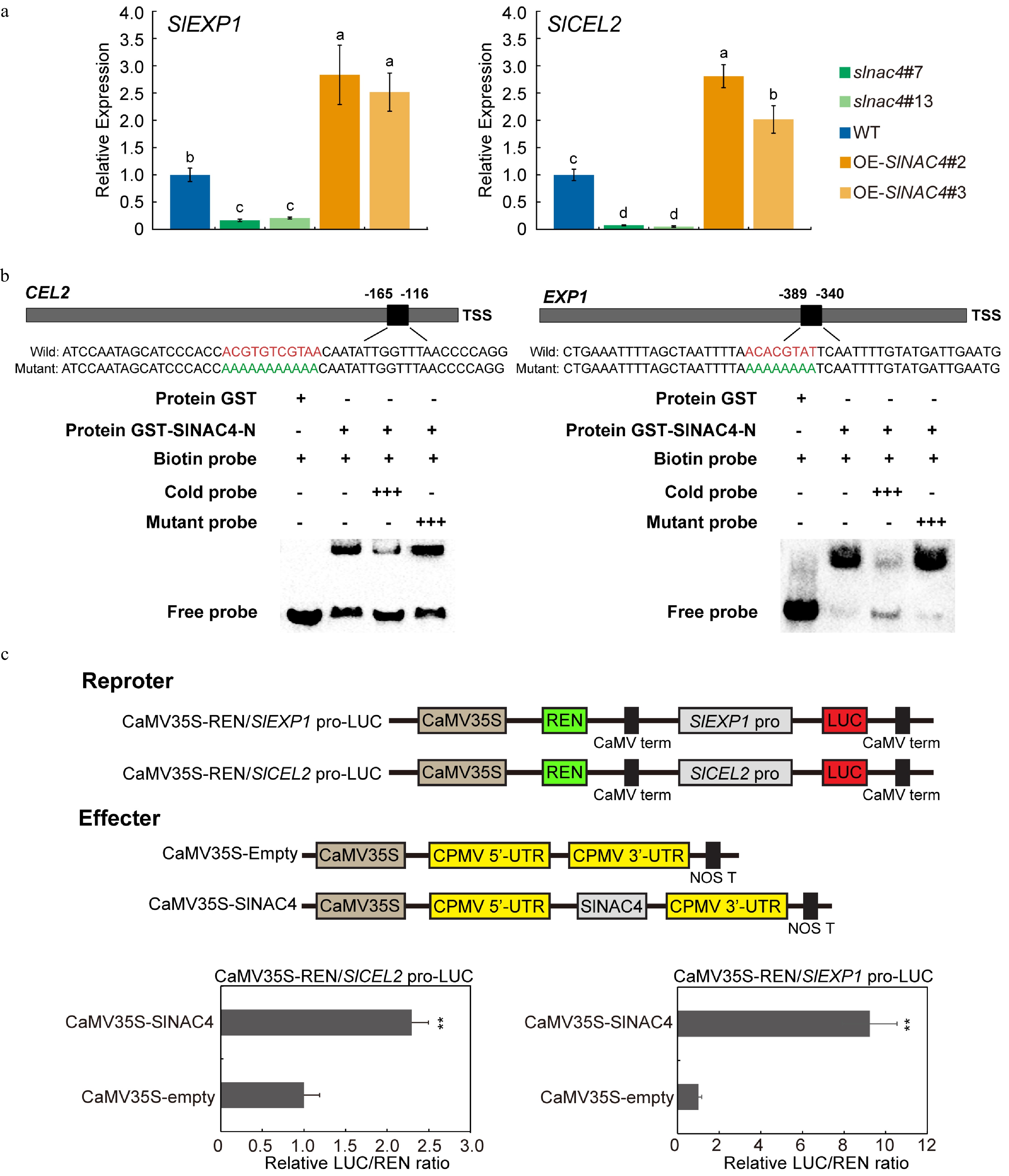
Figure 5. SlEXP1 and SlCEL2 are direct targets of SlNAC4. (a) qRT-PCR analysis of SlEXP1 and SlCEL2 in WT, slnac4#7, slnac4#13, OE-SlNAC4#2, and OE-SlNAC4#3 at the breaker stage. Bars represent ± SD of three independent replicates. SlActin was used as the internal control. (b) SlNAC4 directly binds to the promoters of SlEXP1 and SlCEL2. The wild-type probes containing the NACRS ([TA][TG][AGC]CGT[GA][TA]) were biotin-labeled. 500×cold probes containing the wild-type NACRS or mutated NACRS were performed for competition. + represents presence, and − represents absence, +++ indicates increasing amounts. (c) The promoters of SlEXP1 and SlCEL2 were significantly activated by SlNAC4. Error bars represent ± SD of six replicates. ** indicates p < 0.01 (Student's t-test).

Figure 6. Regulatory model of SlNAC4 in tomato fruit softening. The expression of CEL2 and EXP1 genes related to cell wall metabolism was actived in SlNAC4 over-expressing tomato fruit. The CEL2 and EXP1 enzymes can degrade fruit cell wall components including pectin, cellulose and hemicellulose, which result in the adhesion between normal cells is destroyed and fruit softening is accelerated. On the contrary, the transcription of CEL2 and EXP1 genes is not activated in slnac4 mutant fruit, and the degradation of fruit cell wall component such as pectin, cellulose and hemicellulose are inhibited, and fruit softening was significantly slowed.
-
Our study is the first to compare SlNAC4 mutants obtained by gene editing with OE-SlNAC4 tomato fruit driven by CaMV35S promotor. The expression of SlNAC4 was shown to be enhanced during fruit ripening, and its encoded protein was located in the nucleus. Compared to wild type fruit, the initiation of ripening of slnac4 fruit was delayed and the ripening of OE-SlNAC4 occurred earlier. Fruit with increased or decreased levels of SlNAC4 transcripts showed either increased or decreased softening. SlCEL2 and SlEXP1 involved in cell wall metabolism were shown by RNA-seq to be down-regulated in slnac4 fruit compared to WT fruit. EMSA and DLR confirmed that these genes are the direct targets of SlNAC4.
-
We thank Yaoguang Liu (South China Agricultural University) for providing the binary vector pYLCRISPR/Cas9 system.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers used for qRT-PCR.
- Supplemental Table S2 Primers used for vector construction.
- Supplemental Table S3 Primers used for target site mutation analysis.
- Supplemental Table S4 Detection of mutations on putative off-target sites.
- Supplemental Table S5 Probes containing NACRS used in EMSA.
- Supplemental Data Set S1 Supplemental Data Set S1.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao Y, Zhang Y, Fan Z, Jing Y, Chen J, et al. 2021. Mutagenesis of SlNAC4 by CRISPR/Cas9 alters gene expression and softening of ripening tomato fruit. Vegetable Research 1: 8 doi: 10.48130/VR-2021-0008

Mutagenesis of SlNAC4 by CRISPR/Cas9 alters gene expression and softening of ripening tomato fruit
- Received Date: 14 July 2021
- Accepted Date: 18 September 2021
- Published Online: 28 September 2021
Abstract: Softening is one of the key fruit quality traits, which results from the selective expression of cell wall metabolism genes during ripening. The identification of transcription factors (TFs) that regulate fruit softening is an important field in order to understand and control fruit softening. In tomato, NAC (NAM, ATAF, and CUC) TFs members have been demonstrated to be involved in fruit ripening regulation, including NAC-NOR (non-ripening), NOR-like1, SlNAC4, SlNAC1. Here, we generated slnac4 mutant knockout (CR-SlNAC4) tomato plant by a clustered regularly interspaced short palindromic repeats genomic targeting system (CRISPR/Cas9) and SlNAC4 overexpressing (OE-SlNAC4) plant. In addition to confirming the previously reported results that SlNAC4 positively regulates fruit ripening, we found that SlNAC4 has a strong effect on tomato fruit softening. Compared with the control fruit, fruit softening was inhibited in slnac4 fruit and conversely was accelerated in OE-SlNAC4 tomato fruit. Through RNA-sequencing (RNA-seq) analysis, we found that expression levels of SlEXP1 (expansin) and SlCEL2 (endo-β-1,4 glucanase) genes involved in cell wall metabolism were significantly different in WT (wild type)/slnac4 and WT/OE-SlNAC4 fruit. Further study showed that these genes contained a NAC TF binding domain in their promoter regions. In vitro electrophoretic mobility shift assays (EMSA) and dual-luciferase reporter assays (DLR) demonstrated that these two genes were the direct targets of SlNAC4 binding and transactivation. The results enriched the function of SlNAC4 and provided a new dimension in understanding the regulation of tomato fruit softening.
-
Key words:
- Fruit softening /
- SlNAC4 /
- Transcription factors /
- Cell wall metabolism