









-
Steviol glycosides are a group of glycosylated diterpenes that can be isolated from the plant leaves of Stevia rebaudiana Bertoni ('sweet herb'), native to Paraguay and Brazil, where it is used to sweeten yerba mate tea[1−3]. Besides South America, the Stevia rebaudiana plant is now also grown commercially in countries including China, Japan, the USA, Korea, India, and Vietnam[4]. The mixture of extracted steviol glycosides possesses an overall 100−150-fold sweetness compared with sucrose[5] and its current market value of US
${\$} $ ${\$} $ www.imarcgroup.com/stevia-market ). The main constituents of Stevia extracts are steviol glycosides, which are steviol diterpenes with a series of mono-, di-, and trisaccharide moieties (Fig. 1).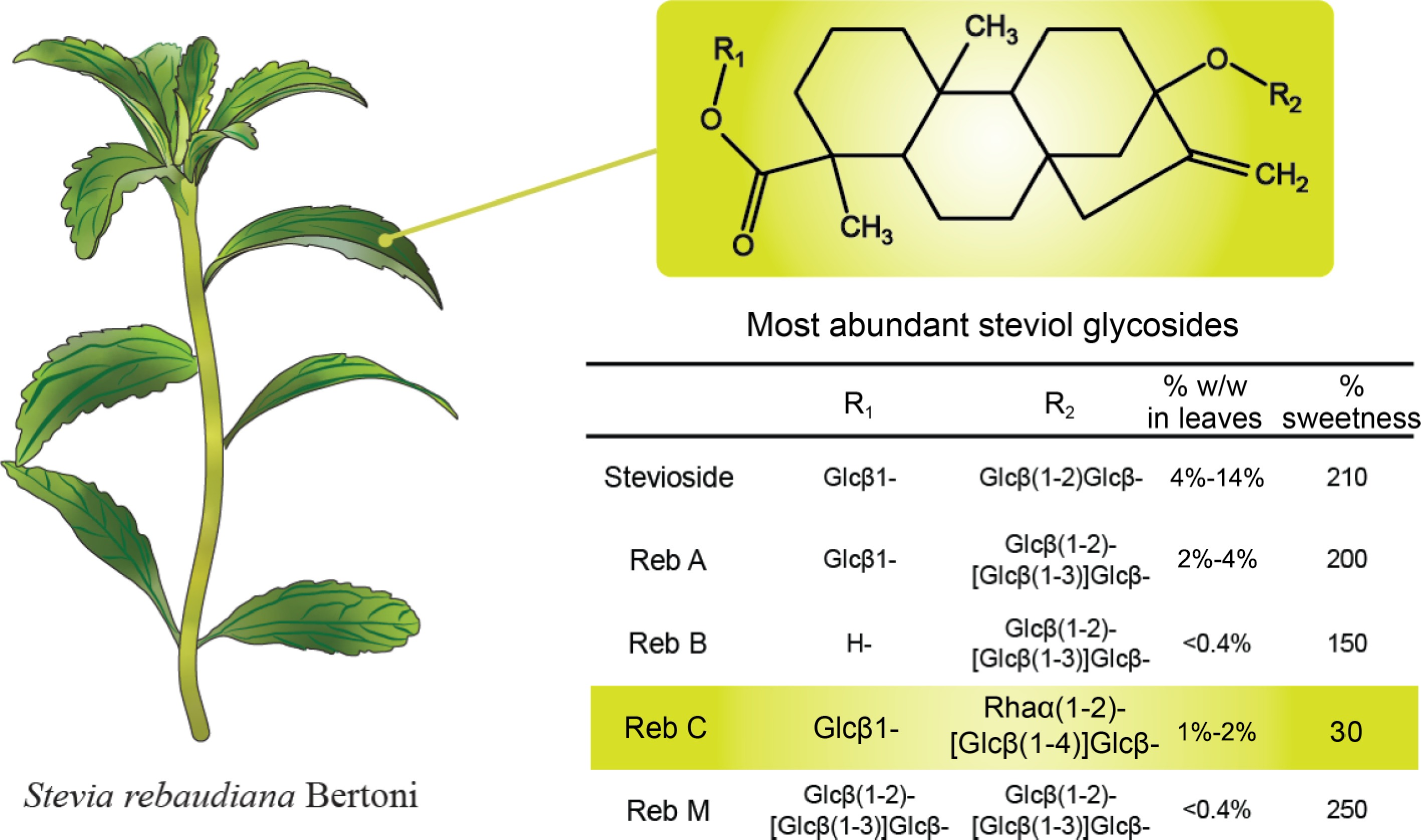
Figure 1.
Stevia rebaudiana Bertoni plant and the structure of relevant steviol glycoside (Reb) variants.
The carbohydrate heterogeneity of steviol glycosides isolated from Stevia leaves has also affected the overall sweetness of the extract, given that the sweetness of each type of glycoside varies significantly when compared to sucrose[6]. For example, the most abundant steviol glycoside present in stevia leaves is stevioside, which possesses 210-fold the sweetness of sucrose, whereas other glycosides such as Rebaudioside A or Rebaudioside B have a slightly lower sweetness of 200 and 150, respectively. Another main steviol glycoside present in stevia leaves is Rebaudioside C (Reb C). Reb C possesses a relatively low sweetness of 30 and contains a bitter aftertaste, as few studies have been conducted on it, we believe that changing the glycosylation of Reb C is a strategy to increase its sweetness, enhance its application, and extend the range of steviol glycosides analogues.
Several genetic and chemoenzymatic approaches have been developed to improve the sweetness of stevioside glycosides. The application of physical and chemical mutagenesis techniques as well as transgenic tools allowed the generation of Stevia plant variants with increased and/or altered levels of the individual types of steviol glycosides[7−11]. Modifications of steviol glycosides were also described using metabolically engineered yeast and bacteria in whole-cell biotransformation[12−16]. However, the majority of reported research on changing the glycosylation of steviol glycosides uses enzyme preparations was comprehensively reviewed[17] and can be divided into the use of recombinant glucosyltransferases[18−22], fructofuranosidases[23−26], cyclodextrin glucanotransferases[27−31], glucosidases[32−36], and galactosidases[37−39]. Interestingly, no attempts to perform biotransformation of steviol glycosides using glycosynthases have been reported yet.
Glycosynthases are mutant variants of glycosidases, in which a catalytically relevant nucleophile in the substrate binding site (usually aspartic acid or glutamic acid residues), is replaced with a small non-nucleophilic amino acid residue (commonly alanine or serine)[40]. While these mutant variants are inactive in hydrolyzing the glycosidases' native carbohydrate substrates in the presence of water. However, in the presence of alcohols which possess good leaving groups (i.e., 1-fluoryl or 1-aryl sugar donors), glycosynthases can catalyze glycosidic linkages at high yields but are void of hydrolase activity. In this work, we chemically synthesized α-glucosyl and α-galactosyl fluorides as donor substrates and applied two previously described glycosynthases derived from Streptomyces sp. (StspBGlcE383A, UniProt ID A0A0N6ZTF1)[41] from Agrobacterium sp. (AgtuBGlcE358S, UniProt ID P12614)[42] to synthesize Reb C bearing additional glucosylated and galactosylated moieties. Given that Reb C is one of the most abundant steviol glycosides present in Stevia extracts, but has a significantly lower sweetness compared to other Rebaudiosides, the addition of glucose or galactose moieties may be of use for increasing the overall sweetness of Stevia extracts.
-
Pyridine, acetic anhydride ((CH3CO)2O), sodium sulfate (Na2SO4), magnesium sulfate (MgSO4) and dichloromethane (CH2Cl2) were obtained from Sinopharm Chemical Reagent Co. (Shanghai, China). Ethyl acetate (EtOAc), petroleum ether and methanol (MeOH) were obtained from General-Reagent Co. (Shanghai, China). Stevia leaves extract, HF-pyridine and sodium methoxide were obtained from Macklin Co. (Shanghai, China). Thin-layer chromatography (TLC) was performed on Merck 60 F254 HPTLC silica gel-coated aluminum sheets. Flash chromatography was performed with silica gel with a 300-400 mesh size. Carbohydrates were visualized using an orcinol staining solution (consisting of 40 mg orcinol monohydrate dissolved in 20 mL of aqueous H2SO4 (3.6 M)). The StspBGlcE383A and AgtuBGlcE358S glycosynthase genes were synthesized by Tsingke (Nanjing, China), and E. coli BL21 (Invitrogen, Shanghai, China) was used as an expression system for the production of the recombinant glycosynthases. Kanamycin and isopropyl-β-D-1-thiogalactopyranoside (IPTG) were purchased from Sigma-Aldrich (Shanghai, China). LCMS-grade acetonitrile and formic acid were purchased from Merck (Nanjing, China) and Aladdin Ltd. (Shanghai, China).
Expression and purification of StspBGlcE383A and AgtuBGlcE358S
-
StspBGlcE383A and AgtuBGlcE358S (Supplemental File S1) were subcloned into a pET-30a expression vector and transformed into E. coli BL21 (DE3) competent cells, then plated on LB agar supplemented with 50 mg/L kanamycin. A 5 mL LB medium containing 50 mg/L kanamycin was inoculated with a single colony and cultivated at 37 °C overnight. One mL of the cell suspension was then transferred into 400 mL LB medium (37 °C, 200 rpm), and grown in log phase until an OD600 value of 0.6 was reached. The recombinant protein expression was then conducted for 16 h at 18 °C in the presence of 1 mM IPTG. The cells were then pelleted by centrifugation at 4,000 g for 15 min and resuspended in 10 mL lysis buffer (50 mM Tris, 100 mM NaCl, 1% Triton X-100 and 1 mM PMSF, pH = 8.0). The cell suspension was lysed by sonication for 20 min on ice and centrifuged at 12,000 g at 4 °C for 20 min. The cleared supernatant was purified by Ni2+-nitrilotriacetate (Ni2+NTA) agarose affinity chromatography using five column volumes of washing buffer (50 mM NaCl, 50 mM Tris, pH = 8.0) before the elution of the recombinant proteins with imidazole buffer (50 mM NaCl, 50 mM Tris, 500 mM imidazole, pH = 8.0). Fractions showing the high absorbance at 280 nm were collected and stored at −80 °C after the addition of 20% (v/v) glycerol. Samples of recombinant protein were analyzed by SDS-PAGE after Coomassie Brilliant G-250 staining.
Synthesis of glycosyl fluorides
-
1-F-glucose (4a) and 1-F-galactose (4b) were synthesized from glucose (1a) and galactose (1b) according to the acetylation/fluorination/deacetylation scheme shown in Fig. 2. In a first step, per-acetylated monosaccharides (2a, 2b) were generated by using acetic anhydride/pyridine. The resulting O-acetylated sugars were converted into the corresponding glycosyl 1-fluorides using 70% hydrogen fluoride-pyridine (HF-Py). After purification, the acetylated glycosyl fluorides (3a, 3b) were then deacetylated using sodium methoxide, yielding the desired products 1-F-glucose (4a) and 1-F-galactose (4b). Product formation was monitored by TLC and MALDI-ToF-MS analysis using a Bruker Autoflex Speed instrument (equipped with a 1000 Hz Smartbeam-II laser) using 6-aza-2-thiothymine as a crystallization matrix.
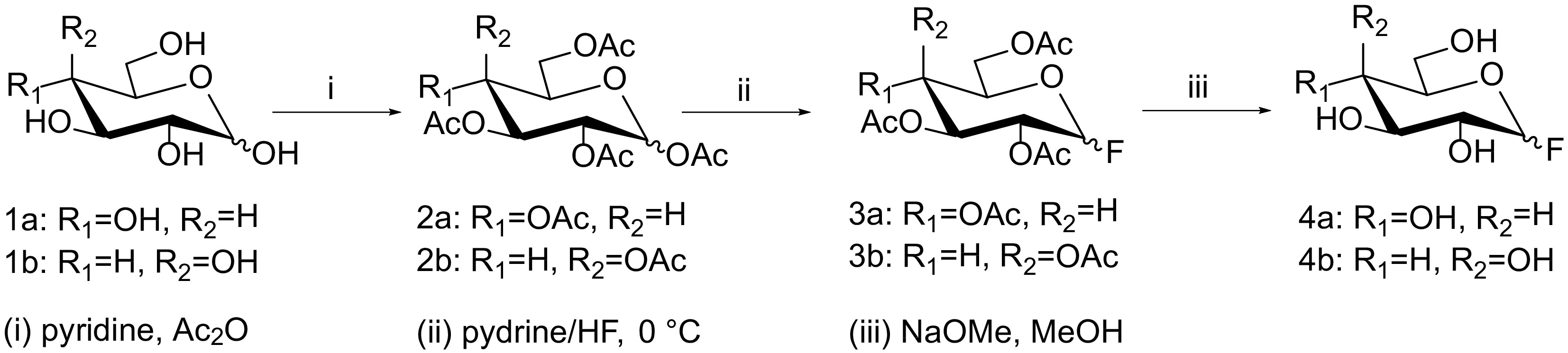
Figure 2.
Synthesis scheme for 1-F-glucose (4a) and 1-F-galactose (4b) from glucose (1a) and galactose (1b). Reagents and conditions: (i) pyridine, acetic anhydride; (ii) 70% hydrogen fluoride-pyridine, 0 °C; (iii) sodium methoxide, methanol.
Glycosylation of Reb C and product analysis
-
Enzymatic glycosylation reactions were performed in volumes of 200 μL which consisted of 5 mM of Reb C, 50 mM of glycosyl fluoride 4a or 4b, and recombinant glycosynthases StspBGlcE383A or AgtuBGlcE358S (5 mg/mL) sodium phosphate buffer (100 mM, pH 7.0). After 24 h at 37 °C, the reactions were terminated by heating at 95 °C for 10 min.
Reactants were analyzed using a LC-ESI-MS method which was based on an RP-HPLC method for steviol glycosides recommended by the Joint FAO/WHO Expert Committee on Food Additives[26]. The analytes were separated on a Shimadzu LCMS 8040 system (Shimadzu Corporation, Kyoto, Japan) using a Cosmosil reversed-phase C18 column (Nacalai Tesque Co., Kyoto, Japan, 250 mm × 4.6 mm; 5 μm pore size) with a flow rate at 0.5 mL/min. Water acidified with formic acid (0.1%v/v) and 100% acetonitrile were used as mobile phases for the gradual elution of the analytes (Supplemental Table S1). Analytes were detected by LC-ESI-MS analysis using scan mode detection in the m/z range between 500−2,000.
-
The expression and purification of StspBGlcE383A and AgtuBGlcE358S were evaluated by SDS-PAGE (Fig. 3). Protein bands corresponding to the expected molecular weight of the recombinant proteins were observed after induction by IPTG and could also be observed in the soluble elution fraction nickel-chelation affinity chromatography.
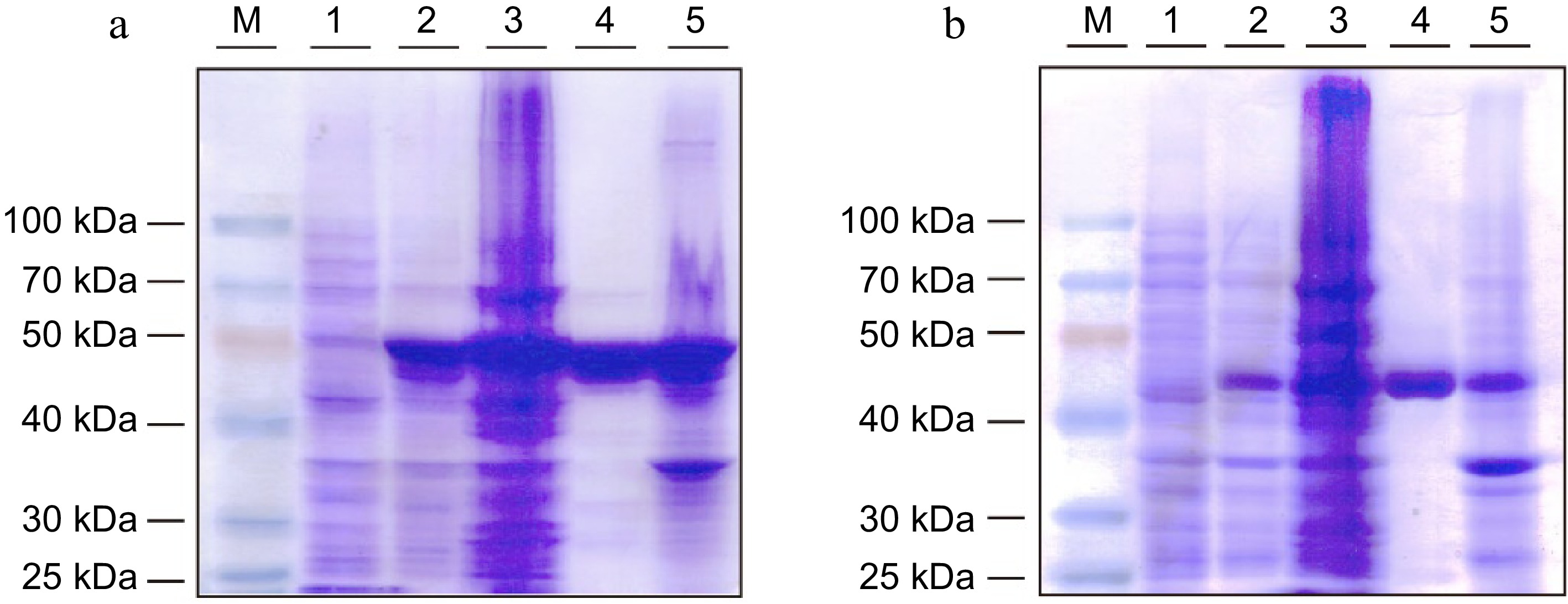
Figure 3.
SDS-PAGE analysis of recombinant (a) StspBGlcE383A and (b) AgtuBGlcE358S protein expressed in E. coli BL21 (DE3) cells. M: Protein marker, Lane 1: cell suspension before induction, Lane 2: cell suspension after induction, Lane 3: supernatant of cell lysis, Lane 4: purified recombinant protein, Lane 5: precipitate of cell lysis.
Synthesis of 1,2,3,4,6-penta-O-acetyl-glucose (2a) and 1,2,3,4,6-penta-O-acetyl-galactose (2b)
-
Two mmol glucose 1a or galactose 1b was added in 2 mL dry pyridine. After the addition of 1.5 mL acetic anhydride, the mixture was stirred in a round-bottom flask at room temperature for 4 h. TLC analysis (2:1 petroleum ether-ethyl acetate) and MALDI-ToF mass spectrometry were used to judge the completion of the per-acetylation reaction (Supplemental Fig. S1). After adding 80 mL of ice-cold water, compounds 2a or 2b were extracted in 60 mL dichloromethane. The organic layer was collected and washed with 80 mL water and 80 mL of saturated sodium hydrogen carbonate. After the addition of anhydrous sodium sulfate, the suspension was filtered and the resulting clear solution dried under reduced pressure (2a: 735 mg, 94%; 2b: 711 mg, 91%).
Synthesis of 1-F-2,3,4,6-tetra-O-acetyl-glucose (3a) and 1-F-2,3,4,6-tetra-O-acetyl-galactose (3b)
-
The per acylated monosaccharides 2a and 2b were dissolved in 6 mL 70% HF-Py in a 50 mL falcon tube at 0 °C, then gently stirred for 30 h. The completion of the reaction was judged by TLC analysis (2:1 petroleum ether-ethyl acetate) and MALDI-ToF mass spectrometry (Supplemental Fig. S2). The reaction was stopped by adding 20 mL of saturated aqueous sodium hydrogen carbonate. After adding 60 mL of dichloromethane, the organic phase was washed once with 80 mL saturated copper sulfate solution and washed twice with 80 mL saturated sodium chloride solution. After the addition of anhydrous magnesium sulfate, the suspension was filtered and the resulting clear solution dried under reduced pressure. The brown residue was purified by silica column chromatography (2:1 petroleum ether-ethyl acetate), and the acetylated glycosyl 1-fluorides 3a and 3b were obtained. Concentration under reduced pressure yielded a pale-yellow powder (3a: 154 mg, 21%; 3b: 132 mg, 18%). These compounds were stored in methanol at a concentration of 200 mM.
Synthesis of 1-F-α-glucose (4a) and 1-F-α-galactose (4b)
-
Due to the instability of components 4a and 4b, the deprotection reactions were only performed in the aliquoted amounts required for the subsequent enzymatic reaction. Typically, 10 μL of acetylated glycosyl fluoride 3a or 3b (200 mM in methanol) were mixed with 980 μL of methanol and 10 μL of methanolic 0.3 M sodium methoxide. The progress of the deacetylation reaction was completed after 4 h at room temperature as judged by TLC (7:2:1 ethyl acetate-methanol-water) and MALDI-ToF mass spectrometry (Supplemental Fig. S3). Methanol was removed under reduced pressure to yield a pale-yellow powder, which was then dissolved in 10 μL of distilled water and immediately used as an acceptor for the glycosynthase reactions of Reb C.
Enzymatic Reb C glycosylation
-
Reb C is one of the main steviol glycoside present in Stevia leaves. Compared to other Rebaudiosides, it possesses a relatively low sweetness, therefore glycosidic decorations with glucose and galactose may be a way to modulate its sweetness. The glycosynthases StspBGlcE383A and AgtuBGlcE358S, which were previously used to glycosylate para-nitrophenyl glucoside in high yield in the presence of 1-F-α-galactose or 1-F-α-glucose[41, 42], were also considered for this study. However, given that Reb C has three terminal glucose moieties, we anticipated that these biocatalysts could be also used for glycosylation reactions of this natural sweetener.
Initial enzymatic reaction trials using para-nitrophenyl-α- and β-glucosides as acceptor substrates and 1-F-α-glucose showed moderate glycosylation efficiency (between 8%−35% product formation, Supplemental Fig. S4) and encouraged us to apply Reb C as acceptor substrate. Using LC-ESI-MS in scan mode of Reb C (Supplemental Fig. S5) and the enzymatic reaction mixtures containing Reb C in the presence of 1-F-α-galactose or 1-F-α-glucose allowed the observation of additional mass signals corresponding to the addition of up to three extra galactose or glucose moieties on Reb C (Fig. 4a−e). Interestingly, StspBGlcE383A showed similar glycosylation abilities in reaction mixtures containing 1-F-α-galactose or 1-F-α-glucose, and allowed to add up to two hexose units to Reb C (Fig. 4b, c).
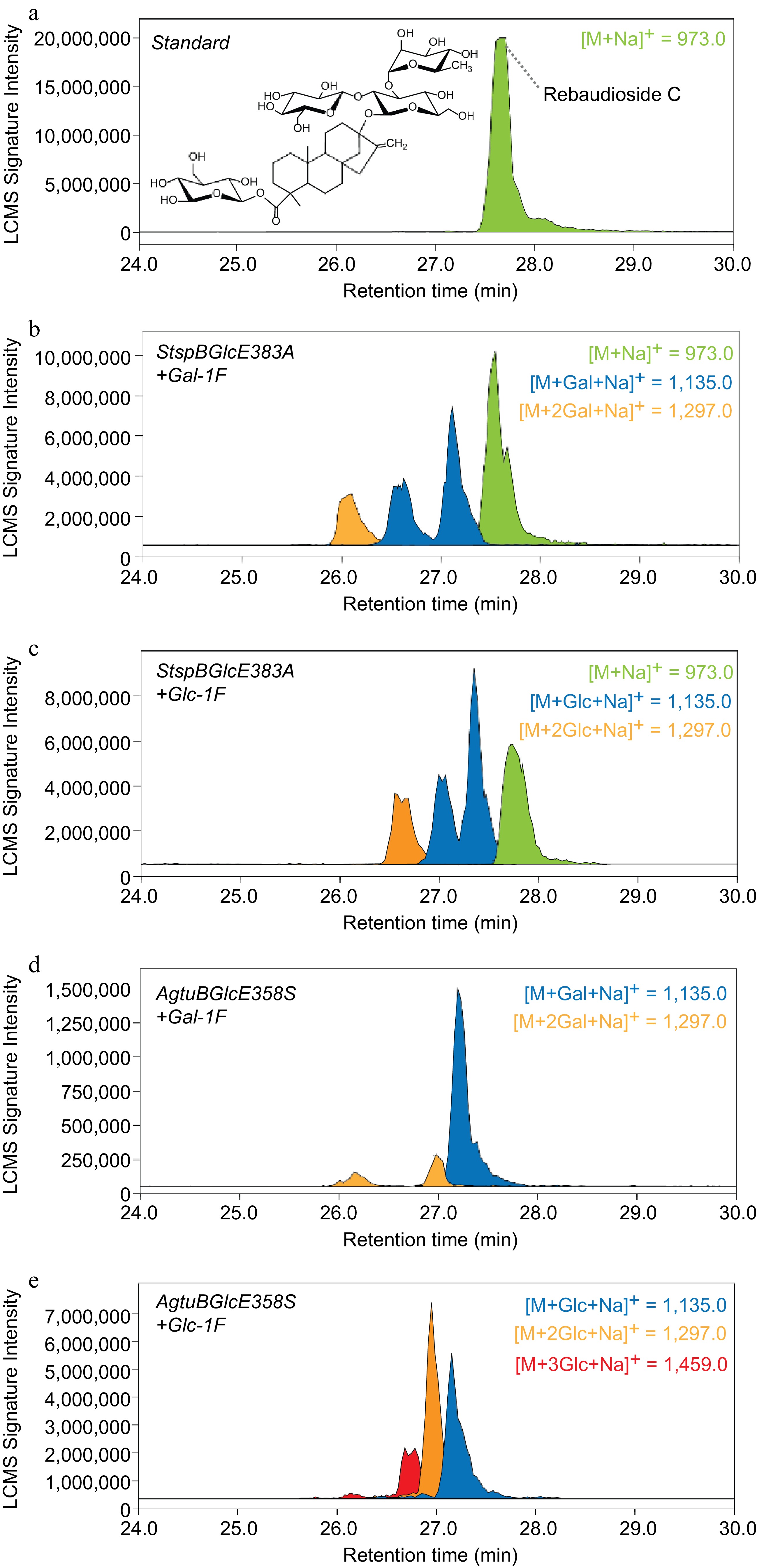
Figure 4.
Extracted ion masses of Reb C and its enzymatically glycosylated reaction products. The analytes were separated using a reversed-phase C18 column and detected using a m/z range between 500−2,000.
According to a previous study, StspBGlcE383A is able to form both β-1,3- and β-1,4-glycosylation, on the other hand, there are more than one hexose that can accept the glycosyl fluoride donor on Reb C, we believe this resulted in a diversity of glycosylated Reb C products. AgtuBGlcE358S was a more efficient biocatalyst and resulted in a quantitative glycosylation of the acceptor substrate (no observable Reb C mass signal present after the reaction) and allowed the addition of up to two glucose or up to three galactose units to Reb C (Fig. 4d, e). Although the feasibility of glycosylating Reb C with glycosynthases could be demonstrated, further optimization of this process may be required. By adding more enzyme to the reaction mixtures, higher glycosylation yields in shorter reaction times are expected as less glycoside fluoride donor is spontaneously degraded[41, 43].
-
We demonstrated that chemically synthesized 1-deoxy-1-fluoro-D-glucose and 1-deoxy-1-fluoro-D-galactose could be used for the enzymatic transglycosylation of Reb C. The two applied glycosynthases were able to further glucosylate or galactosylate Reb C, and therefore allow the expansion of the current repertoire of steviol glycoside analogues. We envisage that both StspBGlcE383A and AgtuBGlcE358S may be also useful glycosynthases for glycosylating a variety of other natural products bearing terminal glucose moieties.
This work was supported in part by the National Natural Science Foundation of China (Grant Nos 31471703, 31671854, 31871793, and 31871754 to JV and LL), and the 100 Foreign Talents Plan (Grant No. JSB2014012 to JV).
-
The authors confirm contribution to the paper as follows: study conception and design: Voglmeir J, Liu L; data collection: Yu YY, Zhang SY, Sun JH, Li YY, Zhang YY, Lu AM; analysis and interpretation of results: Voglmeir J, Liu L; draft manuscript preparation: Voglmeir J, Liu L. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest. Josef Voglmeir and Li Liu are the Editorial Board members of Food Materials Research who were blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and their research groups.
- Supplemental Table S1 HPLC gradient applied for the separation of the stevioside isomers.
- Supplemental File S1
- Supplemental Fig. S1 Thin layer chromatography and MALDI-ToF-MS of per acetylated glucose (2b) and per acetylated galactose (2b). Glc: glucose standard, i(a): glucose after peracylation, Gal: galactose standard, i(b): galactose after peracylation; 2a: 1,2,3,4,6-penta-O-acetyl-glucose, 2b: 1,2,3,4,6-penta-O-acetyl-galactose.
- Supplemental Fig. S2 Thin layer chromatography and MALDI-ToF-MS of 1-F-2,3,4,6-tetra-O-aceytl-glucose (3a) and 1-F-2,3,4,6-tetra-O-aceytl-galactose (3b). ii(a): fluorination of 1,2,3,4,6-penta-O-acetyl-glucose, ii(b): fluorination of 1,2,3,4,6-penta-O-acetyl-galactose, 3a: 1-F-2,3,4,6-tetra-O-acetyl-glucose, 3b: 1-F-2,3,4,6-tetra-O-acetyl-galactose.
- Supplemental Fig. S3 Thin layer chromatography and MS analysis of 1-F-glucose (4a) and 1-F-galactose (4b). Glc=glucose standard, iii(a): deprotected 1-F-2,3,4,6-tetra-acetyl-glucose, Gal: galactose standard, iii(b): deprotected 1-F-2,3,4,6-tetra-acetyl-galactose; 4a:1-F-glucose, 4b:1-F-galactose.
- Supplemental Fig. S4 Enzymatic glycosylation reactions using pNP-α-glucoside (top) and pNP-β-glucoside (bottom) as acceptor substrate were performed in volumes of 20 μl (consisting of 1 mM of the respective pNP-glucoside, 10 mM of glycosyl fluoride 4a or 4b, and recombinant glycosynthases StspBGlcE383A or AgtuBGlcE358S (5 mg/mL) in 100 Mm sodium phosphate buffer, pH 7.0). After 16 h at 37 °C, the reactants were analyzed using a HPLC system equipped with a reversed-phase column (Cosmosil 5C18 MS-II 4.6×250 mm, Nacalai Inc., Japan). The UV absorbance was measured at 300 nm. Solvent A was 50 mM aqueous ammonium formate buffer (pH 4.5), and solvent B was acetonitrile. The analytes were separated using a linear gradient between 25%–50% of solvent B from 0–10 min at a flow rate set to 0.5 mL/min. Peak integration and other chromatographic calculations were performed using the Shimadzu Labsolutions software package (version 5.91).
- Supplemental Fig. S5 LCMS-analysis of Reb C standard. The top panel shows the UV signal (λ = 210 nm) with the relevant m/z signal at t-27.3 min (m/z range 500−2,000) shown in the bottom panel.
- Supplemental File. S1 Gene and Protein sequences of two glycosynthases.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu YY, Zhang SY, Sun JH, Li YY, Zhang YY, et al. 2024. Biocatalytic β-glucosylation/β-galactosylation of Rebaudioside C by glycosynthases. Food Materials Research 4: e008 doi: 10.48130/fmr-0023-0043 

Biocatalytic β-glucosylation/β-galactosylation of Rebaudioside C by glycosynthases
- Received: 05 November 2023
- Revised: 17 December 2023
- Accepted: 27 December 2023
- Published online: 01 March 2024
Abstract: Extracts of Stevia rebaudiana Bertoni plant leaves contain a mixture of diterpene glycosides which possess high sweetness and are used as non-caloric sweeteners in foods and beverages. One of the most abundant of these steviol glycosides is Rebaudioside C (Reb C), which bears one esterified β-D-glucose and one glycosidically linked α-L-rhamnosyl-(1→2)-[β-D-glucosyl-(1→4)]-D-glucose trisaccharide. In this work, we isolated Reb C from commercial Stevia extracts using an orthogonal normal-phase × reversed-phase purification strategy. We then demonstrated that Reb C could be used as a donor substrate for enzymatic trans glycosylation reactions using chemically synthesized 1-deoxy-1-fluoro-D-glucose and 1-deoxy-1-fluoro-D-galactose donors and two engineered glycosynthases derived from Agrobacterium and Streptomyces β-glycosidases. This chemoenzymatic glucosylation/galactosylation strategy may be of use to extend the current repertoire of steviol glycosides analogues for analysis or the sensory evaluation of these novel steviol glycosides.
-
Key words:
- Rebaudioside C /
- Glycosynthase /
- Glycosyl fluoride /
- Glycosylation /
- Protein Engineering