









-
Sweet potato (Ipomoea batatas (L.) Lam.) is the third-largest starchy root crop in the world after potato and cassava, with an annual production of approximately 113 million tons[1]. Rich in nutrients, sweet potatoes contain carbohydrates, carotenoids, anthocyanins, and other bioactive compounds that provide multiple health benefits, including anti-aging, liver protection, and anti-inflammatory properties[2,3]. However, due to the large surface area and high water content of sweet potato tubers, they are highly susceptible to water loss and microbial contamination under the influence of temperature and humidity[4]. During postharvest storage, these factors often lead to excessive dehydration, wilting, and pithiness, accompanied by flesh softening, a decline in nutritional quality, and in severe cases, extensive spoilage and decay.
In China, which accounts for over 50% of the world's sweet potato production, sweet potatoes are typically harvested in late autumn to early winter and require long-term storage to meet year-round demand[5]. Traditional storage methods involve simple preservation techniques such as burying sweet potatoes in pits or wells covered with sand, ash, or rice husks. With advancements in technology, more modern methods such as 1-MCP[6], radiation treatment[7], and high voltage alternating electric[8] have been developed to preserve sweet potatoes.
Ozone (O3), as a green and pollution-free sterilizing agent, shows great potential in the field of postharvest preservation of fruits and vegetables[9]. Previous studies have demonstrated that O3 combined with precooling can effectively maintain the firmness and color of mulberry fruits[10]; O3 fumigation combined with nanofilm packaging can significantly delay the deterioration of sensory quality in button mushrooms, inhibit the outbreak of saprophytic microorganisms, and reduce the accumulation of reactive oxygen species[11]. Additionally, O3 has been shown to suppress the development of dry rot in potatoes and the accumulation of diacetoxyscirpenol (DAS) by inhibiting Fusarium sulfureum growth and downregulating genes related to ergosterol and DAS biosynthesis[12]. However, the effects of O3 on the postharvest quality and physiological changes of sweet potatoes, as well as its regulatory mechanisms, remain unclear.
Omics approaches, such as transcriptomics and metabolomics, have been widely applied to investigate molecular mechanisms. In this study, transcriptomics and metabolomics were used to analyze the changes in the internal metabolite composition of sweet potatoes and the expression of related regulatory genes following O3 treatment, aiming to elucidate the role of O3 in maintaining sweet potato quality.
-
The firmness of nine fruits was measured at each sampling time using a fruit firmness tester (Yueqing Handpi Instruments Co., Ltd., GY-4), with three points taken at the 'equatorial' position of each fruit and averaged.
For each replicate group, the central part of three sweet potatoes was selected, cut into small pieces, wrapped in gauze, and juiced with a mortar and pestle. The SSC of the extracted juice was measured using a Portable Brix Meter (PAL-1, ATAGO, Japan).
Five sweet potatoes of equal size were taken from each treatment to determine their DMC. The tubers were peeled and cut into slices about 0.5 cm thick, then the sliced tubers were mixed and equally divided into three samples of similar mass and baked at 70 °C for 4 d. The DMC was determined by determining the original weight of the substance before drying (W) and the weight of the substance after drying (D) and estimating the percentage of the dry weight: DMC = D/W × 100%[13].
TCC was determined using the acetone-hexane method. Carrot samples were homogenised with liquid nitrogen and repeated three times for each organism and 2.0 g of powder was taken for further analysis. The resulting powder was extracted using a mixture of acetone and hexane followed by cold incubation for 10 min. After decanting the separated solution and passing it through a 0.45 mm filter, the absorbance of the extract at A 450 was determined as an indicator of the TCC[14].
Sample preparation
-
The experimental materials used in this study were 'Xiguahong' (XGH) sweet potatoes sourced from Shixia Village, Gaoling Town, Miyun District, Beijing, China. The sweet potatoes were harvested 130 d after planting and transported to the underground warehouse at the Shixia Village Committee within 24 h of harvest for storage. Sweet potatoes with mechanical damage, pest infestations, or those that did not meet the average size (diameter: 4−5.5 cm, length: 15−20 cm), and weight (300−500 g) were excluded. The selected sweet potatoes were placed in plastic baskets and neatly stacked in the storage pit.
A total of 2,000 kg of sweet potatoes were placed in Pit 1, where an O3 generator (independently developed by the research unit) was installed. The O3 generator operated on a pre-set working mode, alternating between one hour of operation and one hour of rest, with a total of three hours of operation per day to maintain a stable O3 concentration of 15% in the storage pit. The sweet potatoes in Pit 1 were designated as the ozone-treated group (O3). Another 2,000 kg of sweet potatoes were placed in Pit 2, which received no treatment, and was designated as the control group (CK). Both Pit 1 and Pit 2 were kept ventilated, with a temperature maintained between 10−14 °C and humidity between 80%−95%. On day 0 (immediately after storage) and day 120 (4 months later), 15 sweet potatoes were randomly selected from each of the CK and O3 groups. Every five sweet potatoes were treated as one replicate, resulting in a total of three replicates for each group (designated as CK-0d, CK-4M, and O3-4M). After cleaning, approximately 5 cm of flesh (with a volume of about 80−100 cm3) from the middle section of each sweet potato was excised, chopped, flash-frozen in liquid nitrogen, and stored at −80 °C for further analysis.
Transcriptome analysis
-
Total RNA was extracted using the Trizol method and dissolved in 50 µL of DEPC-treated water. The concentration and quality of the RNA were assessed using a NanoDrop spectrophotometer and an Agilent 2100 Bioanalyzer (Thermo Fisher Scientific, MA, USA). Library preparation started with total RNA, with an input requirement of at least 1 µg. The library was constructed using the NEBNext® Ultra™ RNA Library Prep Kit for Illumina. After library construction, initial quantification was performed using a Qubit 2.0 Fluorometer. The library was then diluted to a concentration of 1.5 ng/µL, and the insert size was evaluated using an Agilent 2100 Bioanalyzer. Once the insert size met expectations, the library's effective concentration was accurately quantified using qRT-PCR, ensuring an effective concentration above 2 nM to guarantee library quality. After quality control, the qualified libraries were pooled based on their effective concentration and the required sequencing data output and sequenced on an Illumina platform, generating 150 bp paired-end reads. The raw sequencing data were filtered using fastp to obtain clean reads. The clean reads were then aligned to the reference genome using HISAT to construct the index. Gene alignment was quantified using feature counts, and the FPKM (Fragments Per Kilobase of transcript per Million mapped reads) value for each gene was calculated based on gene length. Differential expression analysis between groups was performed using DESeq2, with significance thresholds determined by the Benjamini & Hochberg method for p-value adjustment. Adjusted p-values and log2 fold changes were used to identify significantly differentially expressed genes. Enrichment analysis was conducted based on hypergeometric tests. For KEGG analysis, pathways were used as the enrichment unit, while for GO analysis, the enrichment was performed based on GO terms.
Metabolomics analysis
-
Three g samples were placed in a freeze-dryer (Scientz-100F) for vacuum freeze-drying. The dried sample was ground into a fine powder using a grinding mill (MM 400, Retsch) at 30 Hz for 1.5 min. A 50 mg portion of the powdered sample was weighed and mixed with 1,200 µL of pre-cooled (−20 °C) 70% methanol extraction solution containing internal standards. The mixture was vortexed every 30 min for 30 s each time, for a total of six vortexing cycles. After centrifugation at 12,000 rpm for 3 min, the supernatant was collected, filtered through a microporous membrane (0.22 µm pore size), and stored in vials for UPLC-MS/MS analysis (ExionLC™ AD, SCIEX). For two-group comparisons, differential metabolites were identified based on variable importance in projection (VIP > 1) and absolute Log2FC (|Log2FC| ≥ 1.0). For multi-group comparisons, differential metabolites (DAMs) were determined using VIP (VIP > 1) and statistical significance (p-value < 0.05, ANOVA). Identified metabolites were annotated using the KEGG Compound database (
www.kegg.jp/kegg/compound/ ). Annotated metabolites were subsequently mapped to the KEGG pathway database (www.kegg.jp/kegg/pathway.html ).Combined analysis of transcriptome and metabolomics
-
Principal component analysis (PCA) was conducted to elucidate the disparities among the samples. Genes and metabolites exhibiting differential expression or concentration relative to the control group were subsequently integrated into the KEGG pathway map. An analysis was conducted on the enriched metabolites and genes within the same pathway to discern their interrelationships, adhering to a stringent screening criterion of p-value less than 0.05. For the purpose of quantifying correlations among genes, the cor function in R was utilized, whereas the Pearson correlation coefficient was applied for the analysis of metabolite correlations. The criteria for screening were established as |PCC ≥ 0.80 and p < 0.05|. Furthermore, canonical correlation analysis (CCA) was implemented to investigate the interactions between sweet potato metabolites and gene molecules within individual pathways, facilitating the construction of a comprehensive network.
Statistical analysis
-
Data were processed using Excel 2019, histograms were plotted using Origin 2024 software.
-
O3 treatment had a significant effect on postharvest storage quality of sweetpotato (Supplementary Fig. S1). After 4 months of storage, the firmness of sweetpotato in the O3-treated group was significantly higher than that in the control group (Fig. 1a). Similarly, SSC and TCC contents showed the same trend, i.e., SSC and TCC were significantly higher in the treatment group than in the control group (Fig. 1b, d). However, the DMC showed an opposite trend, with the O3 treatment group having significantly lower DMC than the control group (Fig. 1c).
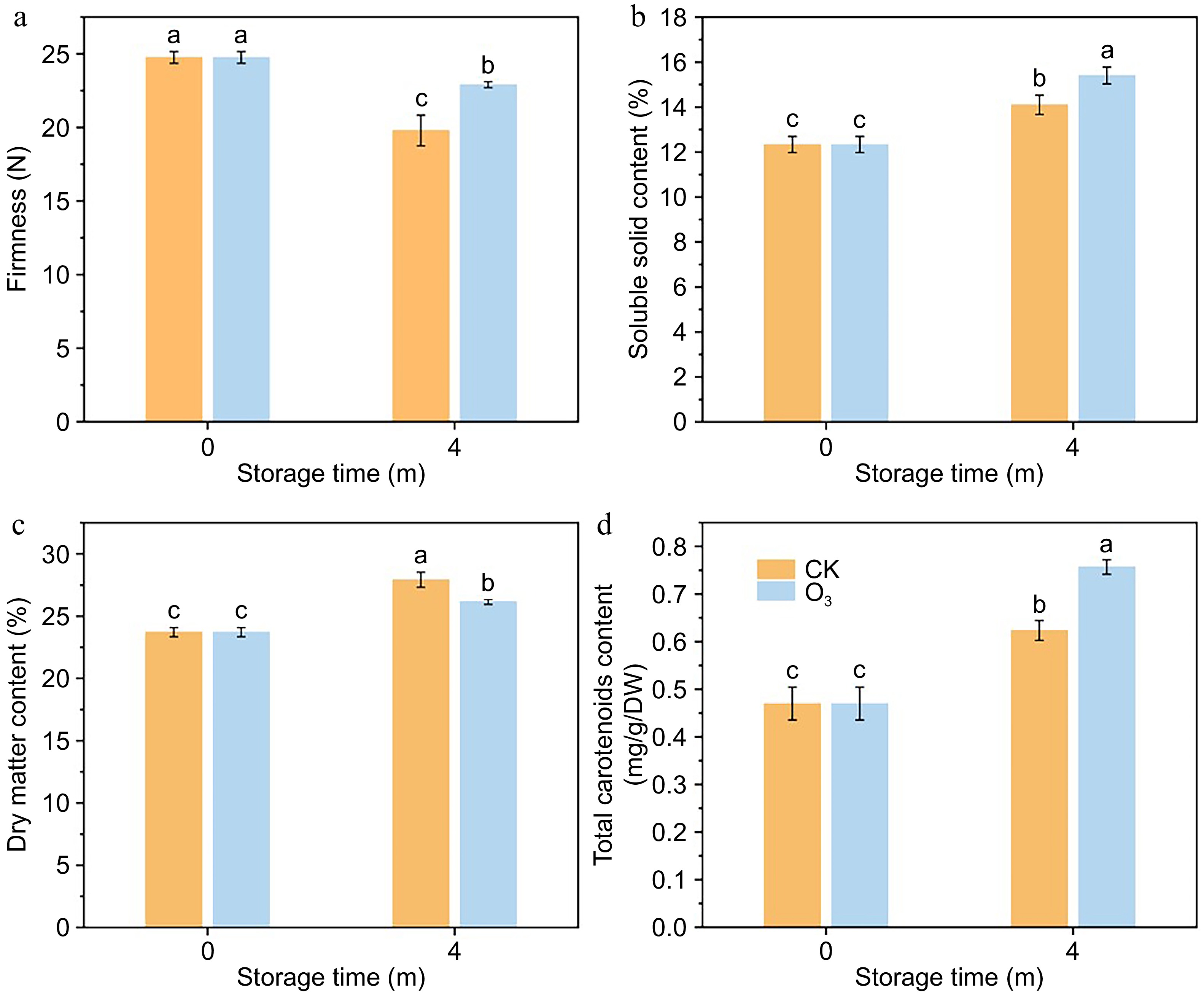
Figure 1.
Effect of O3 treatment on physiological quality of sweet potato. (a) Firmness. (b) SSC content. (c) DMC content. (d) TCC content.
Effects of O3 on metabolites in sweet potatoes
-
To investigate the effects of O3 on sweet potato metabolites, a comprehensive targeted metabolomics analysis was conducted. A total of 1,638 metabolites were identified in 'XGH' sweet potatoes, which were classified into 13 categories (Fig. 2a). These categories included: phenolic acids (303 metabolites), amino acids and derivatives (255 metabolites), others (253 metabolites), lipids (151 metabolites), alkaloids (139 metabolites), flavonoids (118 metabolites), terpenoids (117 metabolites), organic acids (116 metabolites), lignans and coumarins (96 metabolites), nucleotides and derivatives (67 metabolites), quinones (18 metabolites), steroids (four metabolites), and tannins (one metabolite). Among these, phenolic acids, amino acids, and derivatives were the most abundant, accounting for 18.4% and 15.5% of the total metabolites, respectively.
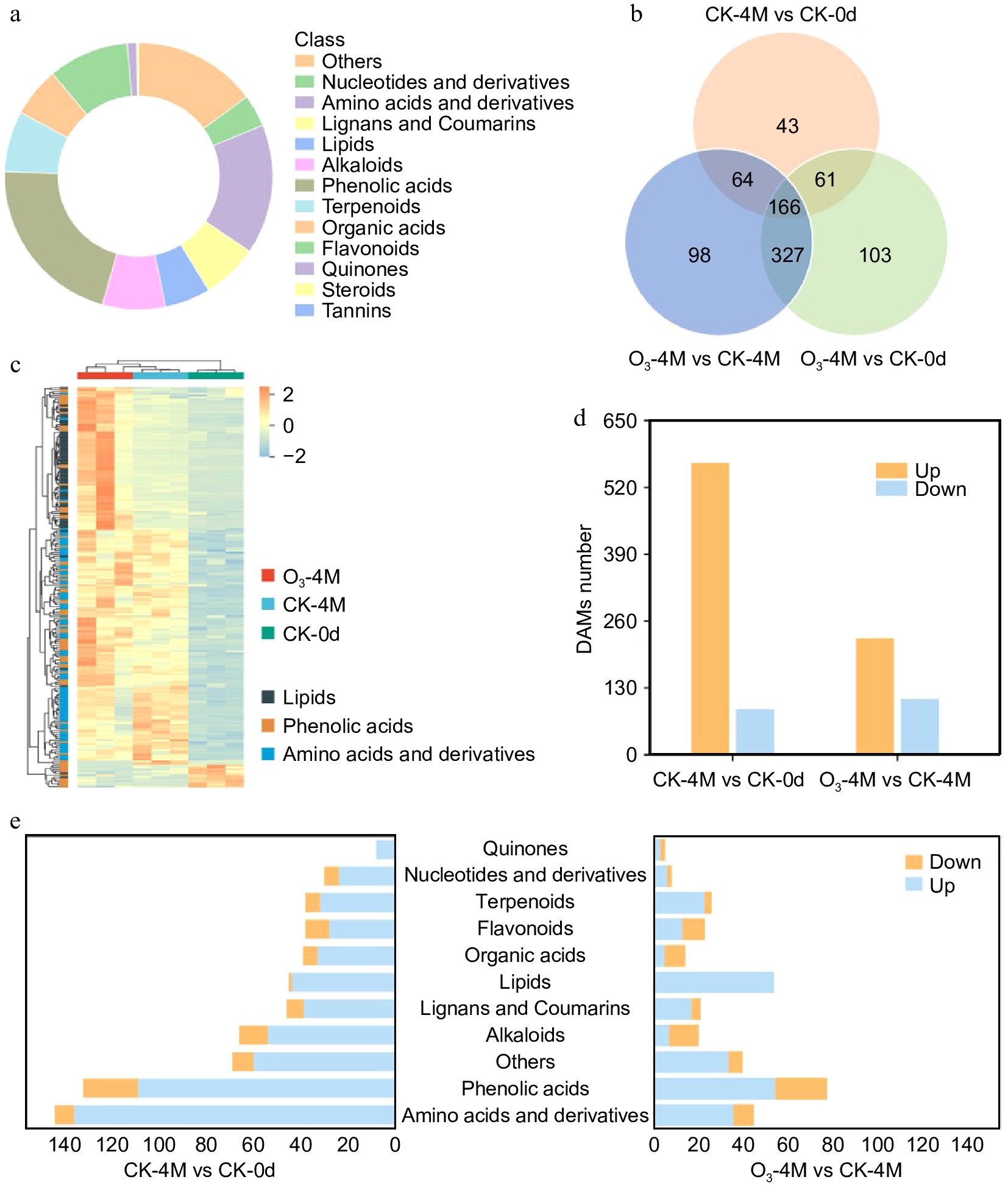
Figure 2.
Metabolome data analysis. (a) DAMs classification statistics. (b) The quantity of different metabolites in three comparative groups. (c) Heat map of clustering of lipids, phenolic acids, and amino acids and derivatives in the comparison of the three groups. (d) Number of upregulated and downregulated DAMs in two comparison groups. (e) Classification and quantity of DAMs in the two comparison groups.
Further analysis of the impact of O3 treatment on differential metabolites (DAMs) revealed that 166 DAMs were shared across the comparison groups, as shown by the Venn diagram (Fig 2b). These metabolites were divided into 11 categories: lipids (40 metabolites), phenolic acids (37 metabolites), terpenoids (16 metabolites), amino acids and derivatives (14 metabolites), others (13 metabolites), lignans and coumarins (12 metabolites), organic acids (10 metabolites), flavonoids (nine metabolites), alkaloids (eight metabolites), nucleotides and derivatives (four metabolites), and quinones (three metabolites). Notably, most lipids, phenolic acids, and amino acids and derivatives increased in CK-4M vs CK-0d and O3-4M vs CK-0d, with lipids and phenolic acids also increasing in O3-4M vs CK-4M. However, the relative abundance of amino acids and derivatives decreased in O3-4M vs CK-4M (Supplementary Table S1; Fig. 2c).
To further analyze metabolite changes during storage, differential expression analysis was performed. In CK-4M vs CK-0d, a total of 655 DAMs were identified, with 567 upregulated and 88 downregulated (Supplementary Table S2; Fig. 2d). The upregulated metabolites primarily included amino acids and derivatives (136 metabolites), phenolic acids (109 metabolites), others (60 metabolites), alkaloids (54 metabolites), and lipids (44 metabolites) (Fig. 2e). The most significantly upregulated metabolites were 2-O-caffeoyl-5-O-(3-hydroxy-4-phenoxy)-6-acetylglucose (15.8-fold), S-methyl-L-cysteine (14.4-fold), 4'-Hydroxy-3'-methoxyacetophenone (acetovanillone) (14.3-fold), and N-feruloyltyramine; moupinamide (13.5-fold). Downregulated metabolites mainly included phenolic acids (23 metabolites), alkaloids (12 metabolites), flavonoids (10 metabolites), others (nine metabolites), and amino acids and derivatives (eight metabolites). The most significantly downregulated metabolites were 4-O-acetyl-3-O-caffeoyl-2-C-methyl-D-erythronate (−16.6-fold), Asp-Pro-Phe (−14.2-fold), shomaside B (−10.5-fold), 3,4,5-trihydroxy-5-[4-(prop-2-enoyloxymethyl) phenoxy]oxane-2-carboxylic acid (−13.3-fold), and caffeoyl(p-Hydroxybenzoyl)artaric acid (−10.5-fold). These results suggest that the changes in these metabolites during sweet potato senescence could have a significant impact on sweet potato quality.
In O3-4M vs CK-4M, 334 DAMs were identified, with 226 upregulated and 108 downregulated (Supplementary Table S2; Fig. 2d). The upregulated metabolites mainly included phenolic acids (55 metabolites), lipids (54 metabolites), terpenoids (23 metabolites), lignans and coumarins (17 metabolites), and flavonoids (13 metabolites) (Fig. 2e). The most significantly upregulated metabolites were Asp-Pro-Phe (14.37-fold), 4-O-acetyl-3-O-caffeoyl-2-C-methyl-D-erythronate (17.42-fold), didymin (13.42-fold), 1-Hydroxypinoresinol-1-O-glucoside (13.18-fold), and 3,4,5-trihydroxy-5-[4-(prop-2-enoyloxymethyl)phenoxy]oxane-2-carboxylic acid (13.81-fold). Among these, 1-Hydroxypinoresinol-1-O-glucoside and 3,4,5-trihydroxy-5-[4-(prop-2-enoyloxymethyl)phenoxy]oxane-2-carboxylic acid belong to phenolic acids and flavonoids, which are known for their antioxidant properties. The downregulated metabolites mainly included amino acids and derivatives (36 metabolites), phenolic acids (23 metabolites), and alkaloids (13 metabolites). The most significantly downregulated metabolites were 5-Glucosyloxy-2-Hydroxybenzoic acid methyl ester (−13.98-fold), cannabisin F (−11.45-fold), feruloyltartaric acid (−11.29-fold), and perillyl alcohol (−11.10-fold). It is noteworthy that all lipids in the O3-4M vs CK-4M group were upregulated (Supplementary Table S1; Supplementary Fig. S2), including lysophosphatidylethanolamine (LPE) and lysophosphatidylcholine (LPC). Studies have shown that these lysophospholipids (LPLs) play important roles in delaying the senescence of fruits and vegetables and in interactions with the environment[15,16]. Based on these results, we propose that the ability of O3 treatment to delay sweet potato senescence may be closely related to the increased content of phenolic acids, flavonoids, and lipids.
Effects of O3 on storage-quality-related genes in sweet potatoes
-
To elucidate the potential transcriptional regulatory mechanisms of O3 treatment in delaying sweet potato senescence, we constructed nine cDNA libraries for subsequent analysis. The number of clean reads per library ranged from a minimum of 47,516,352 to a maximum of 93,913,796. The clean reads of the remaining libraries fell between these values, with an error rate of only 0.03%. The Q20 percentages for all libraries exceeded 97.29%, and the average GC content was 45.98% (Supplementary Table S3). Additionally, Pearson correlation coefficients among biological replicates were all greater than 0.9, indicating that the bioinformatics data obtained were highly reliable (Fig. 3a). The FPKM values of protein-coding genes ranged between 10−2 and 104, with most genes expressed at moderate levels and a smaller subset exhibiting low or high expression levels (Fig. 3b).
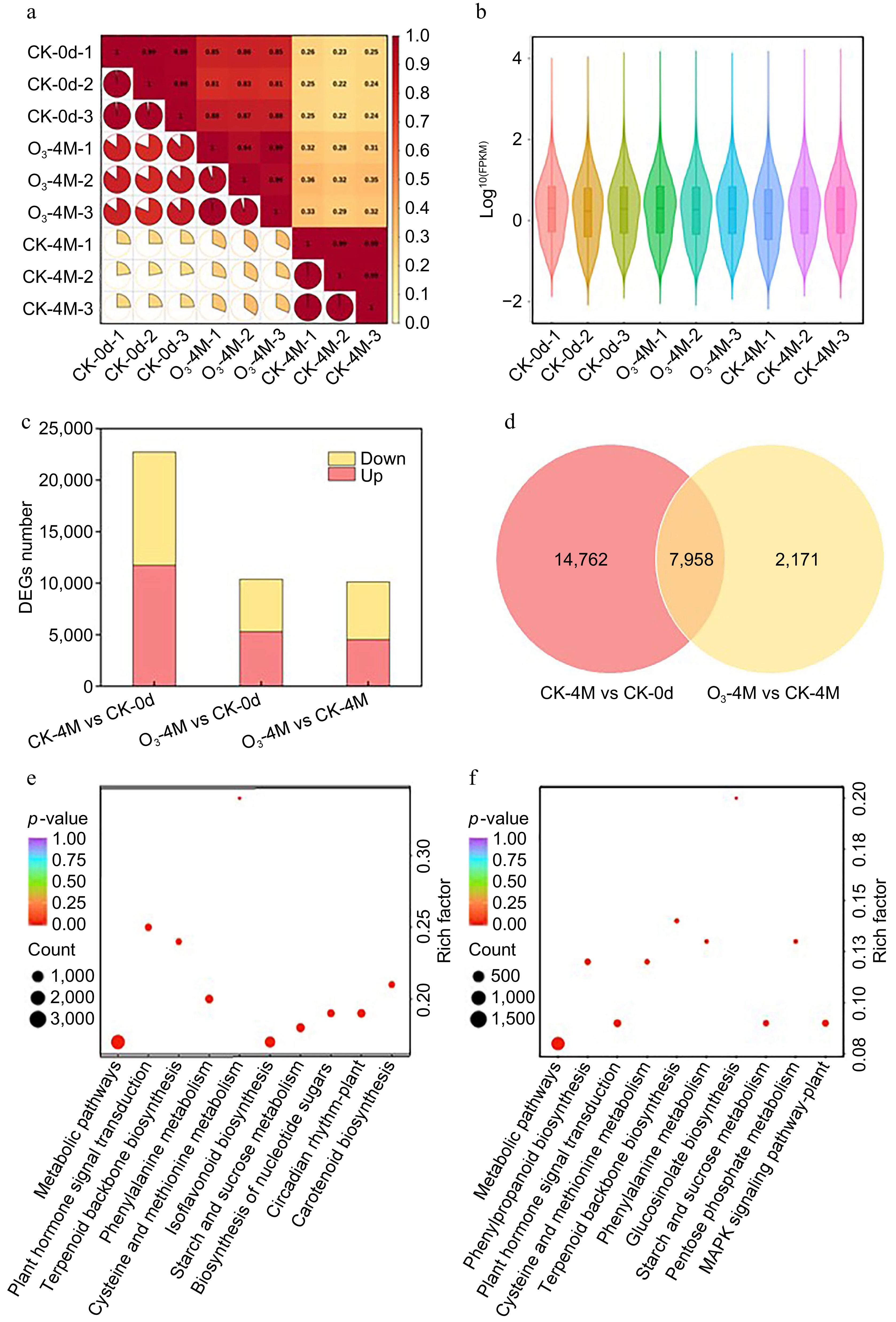
Figure 3.
Transcriptome data analysis. (a) Correlation diagram. (b) Gene expression of nine samples. (c) Number of DEGs up and downregulated in the three comparison groups. (d) Number of duplicate DEGs between the two groups. KEGG enrichment maps of DEGs in (e) CK-4M vs CK-0d, and (f) O3-4M vs CK-0d.
Transcriptome analyses of three sample groups (CK-0d, CK-4M, and O3-4M) were conducted to explore the potential mechanisms by which O3 delays sweet potato senescence. In the comparison of CK-4M vs CK-0d, a total of 22,720 differentially expressed genes (DEGs) were identified, of which 11,739 genes were upregulated and 10,981 genes were downregulated. In the comparison of O3-4M vs CK-0d, 10,383 DEGs were detected, including 5,297 upregulated and 5,086 downregulated genes. These results suggest that O3 treatment mitigated some of the gene expression changes associated with sweet potato senescence, potentially delaying aging during storage by modulating these genes. In the O3-4M vs CK-4M comparison, 10,129 DEGs were identified, with 4,521 upregulated and 5,608 downregulated genes (Fig. 3c). Additionally, 7,958 overlapping DEGs were identified between the CK-4M vs CK-0d and O3-4M vs CK-4M comparisons (Fig. 3d), and KEGG enrichment analysis was performed on these overlapping genes (Fig. 3e, f). The results revealed that the enriched pathways included phenylpropanoid biosynthesis, starch and sucrose metabolism, and plant hormone signal transduction. This suggests that O3 treatment may influence the color, texture, and sweetness of sweet potatoes by modulating genes in these pathways, thereby enhancing the storage quality and preservation of sweet potatoes.
DEGs related to the carotenoid biosynthesis pathway
-
Carotenoids play a critical role in sweet potato pigmentation[16]. The flesh of 'XGH' sweet potatoes is orange-red, which is attributed to its high carotenoid content. Our analysis revealed that O3 treatment influenced the expression of genes associated with the carotenoid biosynthesis pathway. The identified DEGs related to carotenoid biosynthesis included phytoene synthase (PSY), phytoene synthase 2 (PSY2), beta-carotene isomerase D27 (D27), zeta-carotene desaturase (ZDS), beta-carotene hydroxylase 2 (BCH2), 15-cis-zeta-carotene isomerase (Z-ISO), and zeaxanthin epoxidase (ZEP). In the CK-4M vs CK-0d comparison, all of these genes were downregulated, but the degree of downregulation was mitigated in the O3-4M vs CK-0d comparison. Among them, ZEP exhibited the greatest downregulation, with a fold change of −7.35 in CK-4M vs CK-0d, which was reduced to −4.93 in O3-4M vs CK-0d. Notably, in the O3-4M vs CK-4M comparison, the expression levels of PSY, PSY2, ZEP, and Z-ISO were upregulated, with PSY2 showing the highest upregulation (2.49-fold) (Supplementary Table S4; Fig. 4a). These results indicate that O3 treatment promoted the expression of key genes involved in the carotenoid biosynthesis pathway, which likely contributed to maintaining the orange-red coloration of sweet potato flesh during storage.
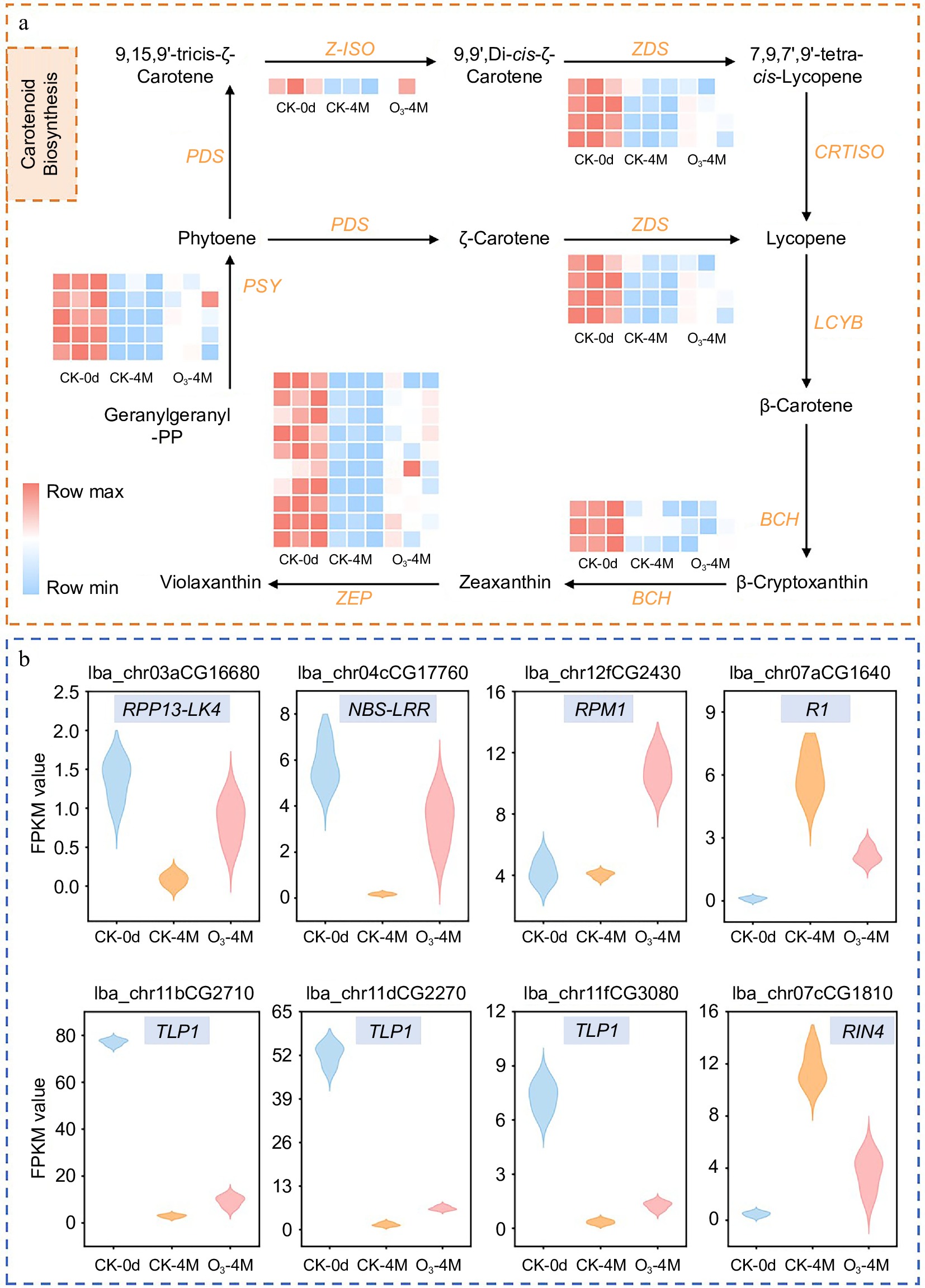
Figure 4.
(a) Carotenoid synthesis map of sweet potato. Heatmaps represent gene expression; heatmaps are plotted using FPKM values, with colors ranging from blue to red indicating a gradual increase in gene expression. (b) Relative expression of disease resistance-related genes in different groups, including disease resistance RPP13-like protein 4 (RPP13-LK4), NBS-NBS-LRR type disease resistance protein (NBS-LRR), disease resistance protein RPM1 (RPM1), late blight resistance protein R1 (R1), thaumatin-like protein 1 (TLP1), RPM1-interacting protein 4 (RIN4).
DEGs related to lignin synthesis
-
During postharvest storage, sweet potatoes are prone to lignification as storage time increases[17], which is primarily characterized by rapid textural deterioration, including an increase in lignin content[18]. We identified DEGs associated with lignin synthesis, including phenylalanine ammonia-lyase (PAL), caffeoyl-CoA O-methyltransferase (CCoAOMT), cinnamoyl-CoA reductase 1 (CCR1), cinnamate 4-hydroxylase (C4H), 4-coumarate--CoA ligase (4CL), 4-coumarate--CoA ligase 2 (4CL2), cinnamyl alcohol dehydrogenase (CAD), and trans-cinnamate 4-monooxygenase (CA4H). In both the CK-4M vs CK-0d and O3-4M vs CK-0d comparisons, all of these genes were upregulated, indicating an increase in lignin synthesis during the natural senescence of sweet potatoes. However, in the O3-4M vs CK-4M comparison, these genes were downregulated, suggesting that O3 treatment inhibited the expression of lignin synthesis-related genes (Supplementary Table S4; Fig. 5). These findings indicate that O3 treatment can suppress the expression of genes involved in lignin biosynthesis, potentially reducing lignification and helping to preserve the texture and quality of sweet potatoes during storage.
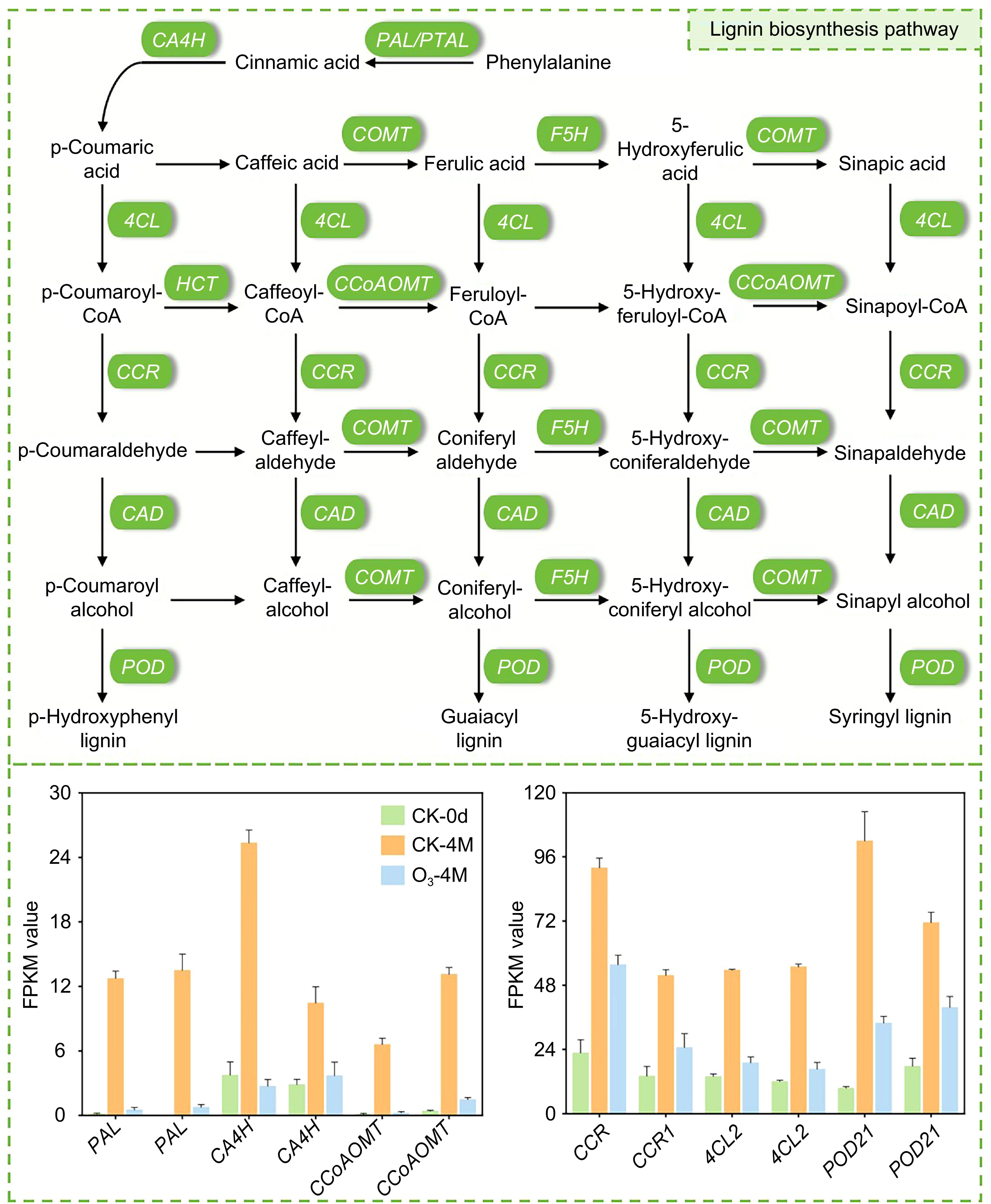
Figure 5.
Map of the lignin synthesis pathway in sweet potato. Bar graphs indicate the relative expression of DEGs, and data are expressed as FPKM values.
DEGs related to sweetness
-
The primary sugars responsible for the sweetness of sweet potatoes are sucrose, fructose, and glucose, with sucrose playing a dominant role in enhancing sweetness[19]. The DEGs identified as being involved in sucrose synthesis include sucrose-phosphate synthase 4 (SPS) and sucrose synthase (SUS). In the CK-4M vs CK-0d and O3-4M vs CK-0d comparisons, the expression of SUS was upregulated, while the expression of SPS was downregulated. In contrast, in the O3-4M vs CK-4M comparison, SUS expression was downregulated, while SPS expression was upregulated. DEGs related to sucrose degradation included acid beta-fructofuranosidase (INV) and soluble acid invertase (S-AI), which were downregulated in CK-4M vs CK-0d, O3-4M vs CK-0d, and O3-4M vs CK-4M (Supplementary Table S4; Fig. 6a). These results suggest that O3 treatment maintains the sweetness of sweet potatoes by upregulating the expression of the sucrose synthesis gene SPS and inhibiting the activity of sucrose-degrading enzymes S-AI and INV.
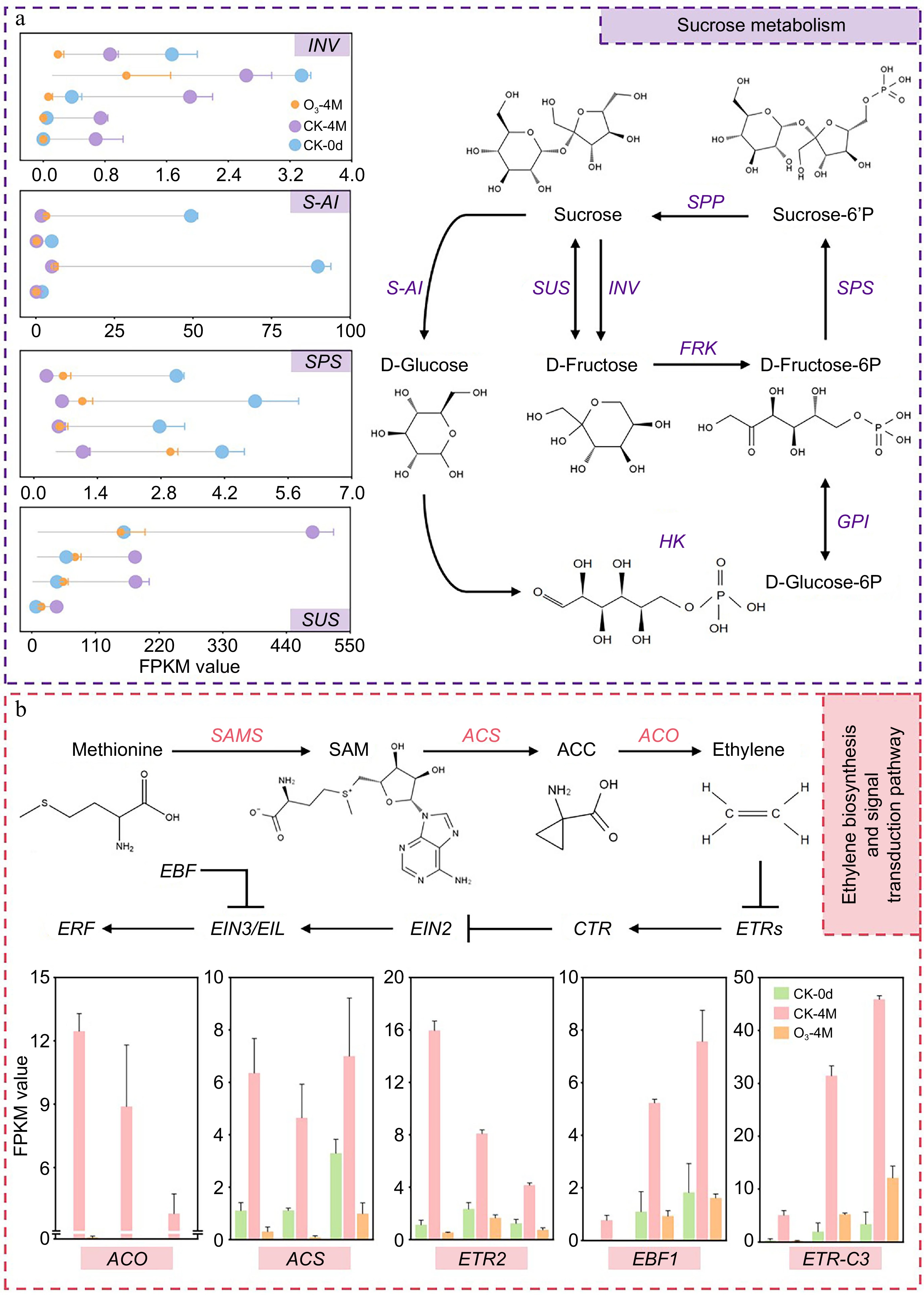
Figure 6.
(a) Diagram of the pathway of sucrose metabolism in sweet potato. Lollipop plots indicate the relative expression of DEGs, and data are expressed as FPKM values. (b) Map of sweet potato ethylene biosynthesis and signal transduction pathway. Bar graphs indicate the relative expression of DEGs, and data are expressed as FPKM values.
DEGs related to ethylene biosynthesis and signal transduction pathways
-
As a climacteric crop, ethylene is a key signaling molecule that regulates most aspects of sweet potato maturation[20]. The DEGs related to ethylene biosynthesis included S-adenosylmethionine synthase 3 (MATs3), 1-aminocyclopropane-1-carboxylate oxidase 1 (ACO1), 1-aminocyclopropane-1-carboxylate oxidase 3 (ACO3), mitogen-activated protein kinase homolog NTF4 (NTF4), and 1-aminocyclopropane-1-carboxylate synthase (ACS). These genes were upregulated in the CK-4M vs CK-0d and O3-4M vs CK-0d comparisons but downregulated in the O3-4M vs CK-4M comparison, with ACO3 showing the largest downregulation (−9.67-fold). DEGs related to ethylene signal transduction pathways included ethylene receptor 2 (ETR2), ethylene-response factor C3 (ERFC3), ethylene response factor 4 (ERF4), and EIN3-binding F-box protein 1 (EBF). Among them, ETR2, CTR1, ERFC3, and ERF4 were upregulated in CK-4M vs CK-0d and O3-4M vs CK-0d but downregulated in O3-4M vs CK-4M. Additionally, EIN3-binding F-box protein 1 (EBF) was upregulated in CK-4M vs CK-0d but downregulated in O3-4M vs CK-0d, and O3-4M vs CK-4M (Supplementary Table S4; Fig. 6b). These findings suggest that O3 treatment suppresses the expression of genes related to ethylene biosynthesis and signal transduction pathways, thereby delaying the senescence of sweet potatoes during storage.
DEGs related to disease resistance
-
Disease resistance refers to the ability of plants to prevent, block, or inhibit the invasion and spread of pathogens, thereby reducing the severity and losses caused by diseases[21]. Several DEGs related to disease resistance were identified, including disease resistance protein RPM1 (RPM1), disease resistance RPP13-like protein 4 (RPP13-LK4), late blight resistance protein R1-A (R1-A), RPM1-interacting protein 4 (RIN4), and thaumatin-like protein 1 (TLP). In the CK-4M vs CK-0d comparison, the expression of all these DEGs was downregulated except for RIN4, which showed upregulation. In the O3-4M vs CK-0d comparison, both RIN4 and RPM1 were upregulated, while the other DEGs were downregulated. Notably, in the O3-4M vs CK-4M comparison, the expression of all these DEGs was upregulated except for RIN4, which was downregulated (Supplementary Table S4; Fig. 4b). These results suggest that O3 treatment promotes the expression of key disease resistance genes, such as RPM1, RPP13-LK4, R1-A, and TLP, while modulating the expression of RIN4. This enhanced expression of disease resistance-related genes under O3 treatment may contribute to improving the ability of sweet potatoes to resist pathogen attacks during storage, thereby maintaining their quality and reducing losses.
Combined transcriptomic and metabolomic analysis
-
Combined KEGG enrichment analysis of transcriptomics and metabolomics revealed that DEGs and DAMs were enriched in pathways such as biosynthesis of secondary metabolites, phenylpropanoid biosynthesis, and starch and sucrose metabolism in both CK-4M vs CK-0d and O3-4M vs CK-4M comparisons (Fig. 7a, c).
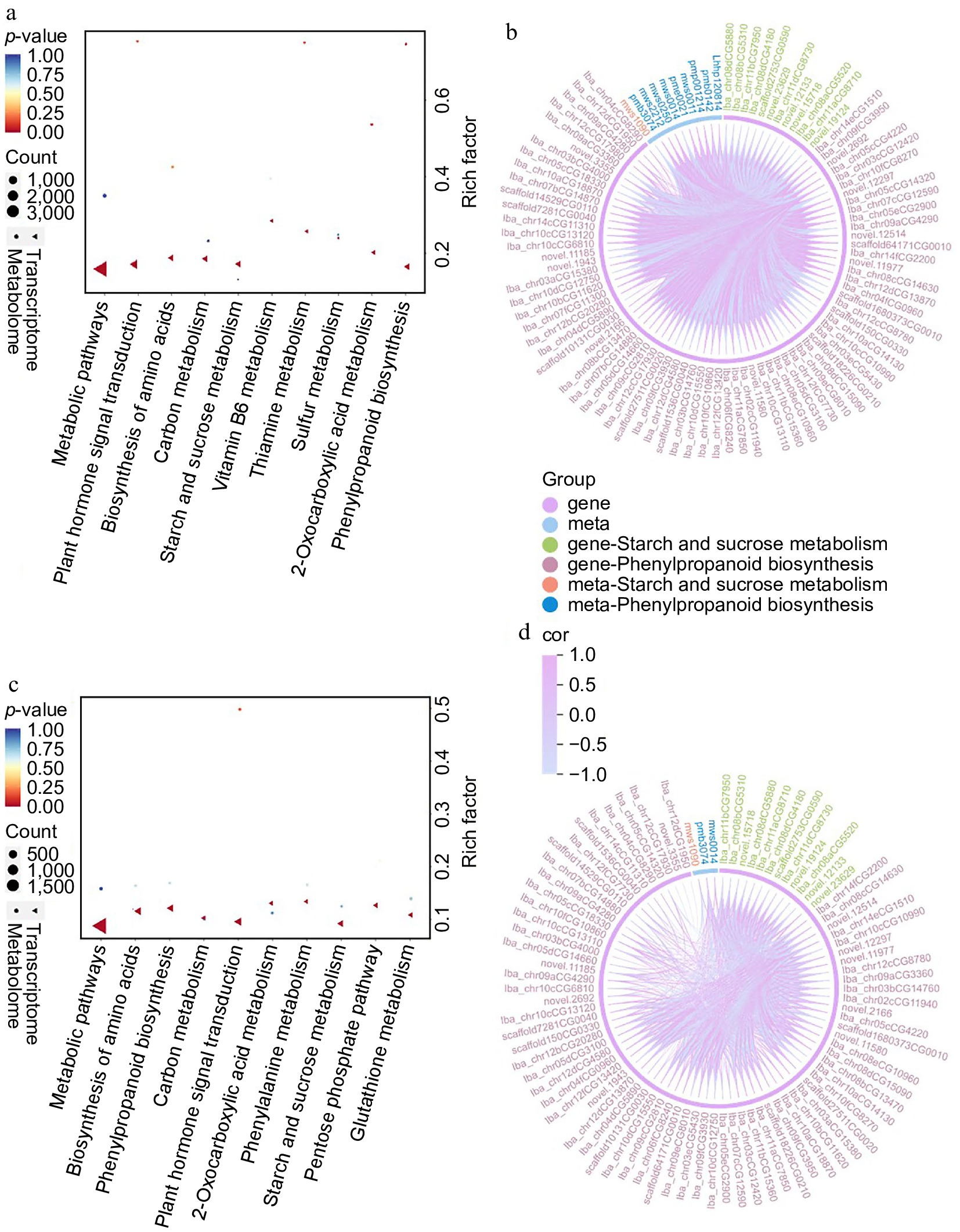
Figure 7.
KEGG enrichment pathways for DEGs and DAMs in (a) CK-4M vs CK-0d, and (c) O3-4M vs CK-4M. Correlation and chord plots of DAMs and DEGs enriched for phenylpropanoid biosynthesis and starch and sucrose metabolism in (b) CK-4M vs CK-0d, and (d) O3-4M vs CK-4M.
To further explore the interactions between DEGs and DAMs, the correlation networks in key enriched pathways were analyzed. There are 152 genes and one metabolite (D-glucose-1-phosphate) involved in the regulation of starch and sucrose metabolism in the CK-4M vs CK-0d comparison, and the regulatory relationships are complex. There were 108 up regulated genes, which were positively correlated with D-glucose-1-phosphate. There were also 44 down-regulated genes, which in turn were negatively correlated with this metabolite. A similar pattern of regulation was observed in the comparison of O3-4M vs CK-4M, with 105 genes and one metabolite (D-glucose-1-phosphate), of which 74 were up-regulated genes, which were also positively associated with this metabolite, and 31 were down regulated genes, which were negatively associated with this metabolite. (Supplementary Table S5; Fig. 7b, d). This suggests that D-glucose-1-phosphate plays a central role in this pathway, and O3 treatment may modulate its metabolism by regulating key genes, thereby maintaining sweet potato sweetness during storage.
Genes and metabolites involved in phenylpropanoid biosynthesis were also analyzed in detail. A total of 538 genes (including 445 upregulated and 93 downregulated genes) and nine metabolites are involved in the regulation of this pathway in CK-4M vs CK-0d. Among the downregulated genes: aldehyde dehydrogenase family 2 member (ALDH2), anionic peroxidase swpb2 (SWPB2), CCoAOMT 5, caffeoylshikimate esterase (CSE), CCR SNL6. peroxidase P7-like (POD P7), shikimate O-hydroxycinnamoyltransferase-like, stemmadenine O-acetyltransferase-like (SAT) with L-phenylalanine, syringin, L-tyrosine, 5-O-p-coumaroylquinic acid* were negatively correlated. Upregulated genes: 4-coumarate-CoA ligase-like 1 (4CL), CSE was positively correlated with eugenol, while the downregulated gene, salutaridinol 7-O-acetyltransferase-like (EC) was negatively correlated with eugenol. Upregulated genes:berberine bridge enzyme 15 (BBE 15), CSE, POD 47 was positively correlated with caffeic acid and sinapine; downregulated gene: aldehyde dehydrogenase family 2 member C4 (ALDH2 C4), SWPB 2, CCoAOMT 5, CCR SNL, shikimate O-hydroxycinnamoyltransferase-like were negatively correlated with caffeic acid and sinapine. The upregulated genes were negatively correlated with ferulic acid*, except for BBE 21, CCR 2, and tetraketide alpha-pyrone reductase 1 (TKPR1), which were positively correlated with ferulic acid*. A total of 229 genes and two metabolites were involved in the regulation of this pathway in O3-4M vs CK-4M. These include 153 upregulated genes and 76 downregulated genes among them. Specifically, the downregulated genes: 4CL2, anthocyanidin 3-O-glucosyltransferase 5 (UFGT 5), caffeic acid 3-O-methyltransferase-like (COMT), C4H, CCR1, POD 21, PAL, and CA4H were positively correlated with 5-O-p-coumaroylquinic acid*, a positive correlation, and ferulic acid*, a negative correlation (Supplementary Table S5; Fig. 7b, d). These findings suggest that O3 treatment significantly influences the phenylpropanoid biosynthesis pathway by regulating the expression of key genes and their interactions with metabolites. By enhancing or suppressing the accumulation of critical compounds such as D-glucose-1-phosphate, 5-O-p-coumaroylquinic acid*, and ferulic acid*, O3 treatment contributes to the preservation of sweet potato texture and sweetness.
-
To elucidate the preservation effect of O3 treatment on sweet potato, its firmness, SSC and DMC were studied. Decreased hardness is an indicator of postharvest quality deterioration of sweet potato. At the end of storage, the firmness of sweet potato in the O3 treatment group was 15.71% higher than that of sweet potato in the CK group. SSC was an important trait for measuring fruit quality[22]. At 4 months of storage, the SSC of sweet potato in the O3 treated group was 13.74% higher than that of sweet potato in the CK group. These results indicated that O3 treatment could inhibit the decrease in firmness and increase the SSC of sweet potato during storage. Previous studies have shown that there is a positive correlation between dry matter content and storability during storage of agricultural products and that agricultural products with a rapid increase in DMC have high tissue permeability, are less prone to anaerobic respiration, and are therefore more resistant to storage[23]. However, this study found that the DMC of the O3-treated group was relatively low, which was about 6.34% lower than that of the CK group. Comprehensively, other indicators showed that O3-treated sweet potato showed better storage tolerance, probably because O3-treated sweet potato had a more compact texture structure compared with the CK group, which was more conducive to the retention of moisture in the potato during the storage process with less exchange of moisture with the outside world, and the constancy of moisture in the late stage of storage could ensure the normal physiological activities of potato respiration and other physiological activities to ensure the supply of energy, which showed better storage tolerance.
Our study revealed that the major metabolites in sweet potatoes are phenolic acids, and amino acids and derivatives. During storage, the relative content of phenolic acids, amino acids and derivatives, and lipids increased. Previous studies have shown that the content of phenolic acids is closely related to the antioxidant activity of fruits and vegetables[24]. In our research, O3 treatment promoted the accumulation of phenolic acids in sweet potatoes during storage, including 4,5-O-dicaffeoylquinic acid methyl ester (4,5-CQME), elaidolinolenic acid, 6-O-caffeoyl-D-glucose, and O-caffeoyl maltotriose. This finding aligns with the study by Cheng et al., which demonstrated that hot air treatment increased the content of phenolic acids in sweet potatoes and maintained their quality[25]. Lipids are involved in various physiological activities and play a vital role in the plant's response to abiotic stresses. O3 treatment also promoted the accumulation of lipids in sweet potatoes, such as LPE and LPC. This result is consistent with Xu et al.'s findings, which showed that melatonin treatment increased the lipid levels in fruits compared to untreated samples[26]. Notably, exogenous LPE application has been reported to delay plant tissue senescence by counteracting membrane deterioration caused by membrane-bound enzymes[27]. Amino acids are associated with cell metabolism and growth in plants[28], reflecting fruit quality and influencing senescence[29]. An increase in amino acid content is often closely related to the postharvest senescence of agricultural products[30]. In this study, O3 treatment inhibited the synthesis of amino acids and derivatives in sweet potatoes, such as Pro-Asp, L-Lysine, L-Glutamine, and Ile-Pro-Thr. This result is consistent with Zhou et al.'s study, which found that LED irradiation maintained low amino acid levels in postharvest bok choy[31].
The color of sweet potato tubers is an important factor affecting quality traits and is mainly influenced by carotenoid content. The most common carotenoid in the 'XGH' variety is β-carotene[32]. In the present study, it was found that the carotenoid content of O3-treated sweetpotato was elevated by about 22.58% compared to that of sweet potato in the CK group. This study also found that O3 treatment affects the expression of genes related to carotenoid biosynthesis, including PSY, PSY2, Z-ISO, and ZEP.PSY catalyzes the initial stage of carotenoid biosynthesis and is a key enzyme that determines total carotenoid content[33,34]. Studies have shown that overexpression of PSY in carrots significantly increases carotenoid biosynthesis[35]. In our study, PSY2 showed the highest fold change in the O3-4M vs CK-4M comparison, suggesting that O3 treatment may inhibit the downregulation of PSY2 and promote the accumulation of carotenoid precursors, inducing carotenoid biosynthesis. ZEP is a crucial mediator in carotenoid accumulation in yellow-fleshed sweet potatoes, as it participates in the epoxidation of β-carotene and β-cryptoxanthin, influencing flesh color[36]. In this study, O3 treatment increased the expression of ZEP in sweet potatoes stored for four months compared to untreated samples. Additionally, Z-ISO expression levels were positively correlated with carotenoid content[37], and our study found that Z-ISO expression was also upregulated in O3-4M vs CK-4M. Overall, O3 treatment promoted carotenoid accumulation by increasing the expression of carotenoid biosynthesis-related genes, thereby maintaining the color of sweet potato tubers. This aligns with the findings of Tzortzakis et al., who reported that O3 treatment increased β-carotene, lutein, and lycopene content in tomato fruits[38].
Changes in hardness during sweet potato storage reflect texture and are an important indicator of storage effectiveness. One manifestation of quality decline during storage is the significant increase in tuber hardness[39], likely caused by water loss and lignification of the tissue[40]. In this study, O3 treatment inhibited the expression of lignin biosynthesis-related genes, including PAL, CCoAOMT, CCR1, C4H, 4CL, 4CL2, CAD, and CA4H. We hypothesize that O3 treatment may regulate lignin accumulation by suppressing the activity of enzymes encoded by these genes, thereby delaying the hardening of sweet potato tissue and maintaining better quality. Similar findings have been reported in bamboo shoots, where Luo & Zhang demonstrated that O3 treatment delayed the increase in hardness by inhibiting lignification during cold storage[41].
Changes in sweet potato tuber sweetness are primarily attributed to changes in sucrose content, followed by fructose and glucose[42]. In this study, we identified key enzymes related to sucrose synthesis, including SPS and SUS. SPS activity is positively correlated with sucrose accumulation in higher plants[43]. The expression of SPS in sweet potatoes decreased after O3 treatment, which may be related to variety or storage temperature. In contrast, SUS expression increased following O3 treatment[44], as SUS is more active in sucrose-rich tissues such as roots, tubers, and fruits. For sucrose degradation, key enzymes INV and S-AI showed reduced expression levels after O3 treatment. Overall, O3 treatment maintained the sweetness of sweet potatoes by upregulating sucrose synthase (SUS) and downregulating sucrose-hydrolyzing enzymes (INV and S-AI). This result aligns with studies on cyclic variable temperature control (CVTC), where CVTC induced SUS activity, promoted sucrose synthesis, and inhibited acid invertase activity, delaying sucrose degradation to enhance sweetness and preserve physiological quality[45]. Additionally, we observed that the content of D-Glucose-1-phosphate, a key metabolite involved in glucose synthesis and degradation, increased after O3 treatment. The gene phosphoglucomutase, which is responsible for D-glucose-1-phosphate metabolism, was also upregulated, consistent with findings by Chen et al., who reported that low-temperature treatment effectively suppressed the decrease in D-glucose-1-phosphate in blue honeysuckle[46].
As a climacteric crop, the ripening of sweet potato tubers is primarily regulated by ethylene. Ethylene biosynthesis involves two steps, the first of which is rate-limiting: S-adenosyl-L-methionine (SAM) is converted to ACC under the catalysis of ACC synthase (ACS), followed by ACC's conversion to ethylene via ACC oxidase (ACO). In this study, the expression of ethylene biosynthesis-related genes (ACS and ACO) was suppressed by O3 treatment. Ethylene signal transduction begins with ethylene recognition by receptors (e.g., ETR), followed by signal transduction through the transcription factor EIN3 to the nucleus, ultimately activating or repressing downstream genes via ERF transcription factors[47]. Our results showed that the expression of ETR2 and CTR1, negative regulators of ethylene signaling, was inhibited by O3 treatment, which may be a mechanism to re-activate downstream signaling pathways[48]. The expression of MPK3, a MAPK protein kinase that phosphorylates and stabilizes ACS proteins, was also suppressed by O3 treatment, reducing ethylene biosynthesis[49]. Additionally, EIN3 and ERFC3 expression was downregulated, suggesting that O3 treatment reduced ethylene production and signal transduction, thereby delaying sweet potato senescence and extending storage life.
The plant immune system responds to biotic and abiotic stresses by recognizing external signals and activating defensive pathways[50]. In this study, we observed that ozone treatment influenced the expression of key immune regulators. RIN4, a negative regulator of PAMP-triggered immunity (PTI) and effector-triggered immunity (ETI) was downregulated after O3 treatment[51]. Conversely, the expression of RPM1 (a key gene involved in various defense mechanisms)[52], RPP13-LK4 (which plays a role in heat tolerance)[53], R1 (a late blight resistance gene)[54], and TLP1 (which enhances resistance by regulating MAPK signaling through JA and ABA pathways)[55] was upregulated following O3 treatment. These findings suggest that O3 treatment modulated the expression of immune regulatory factors, enhancing the disease resistance of sweet potatoes and contributing to their quality preservation during storage.
-
Our study elucidated the effect of O3 treatment on the postharvest storage quality of 'XGH' sweet potato and its underlying mechanisms. The findings demonstrated that O3 treatment preserved the color and firmness of sweet potatoes by preventing the downregulation of carotenoid biosynthesis-related genes and inhibiting the upregulation of lignin synthesis-related genes. Additionally, the significant upregulation of disease resistance-related genes enhanced the sweet potato's resistance to pathogens. Moreover, differential regulation of genes in the sucrose metabolism and ethylene biosynthesis pathways delayed senescence and maintained sweetness. The increase in lipid and phenolic acid metabolite levels, coupled with the reduction in amino acid content, contributed to maintaining the nutritional quality of sweet potatoes during storage. Based on these results, we propose a regulatory network model illustrating how O3 treatment modulates the postharvest quality of sweet potatoes. This model highlights the multifaceted role of O3 in maintaining physiological and nutritional integrity, providing a framework for future applications of O3 in sweet potato storage and preservation (Fig. 8).
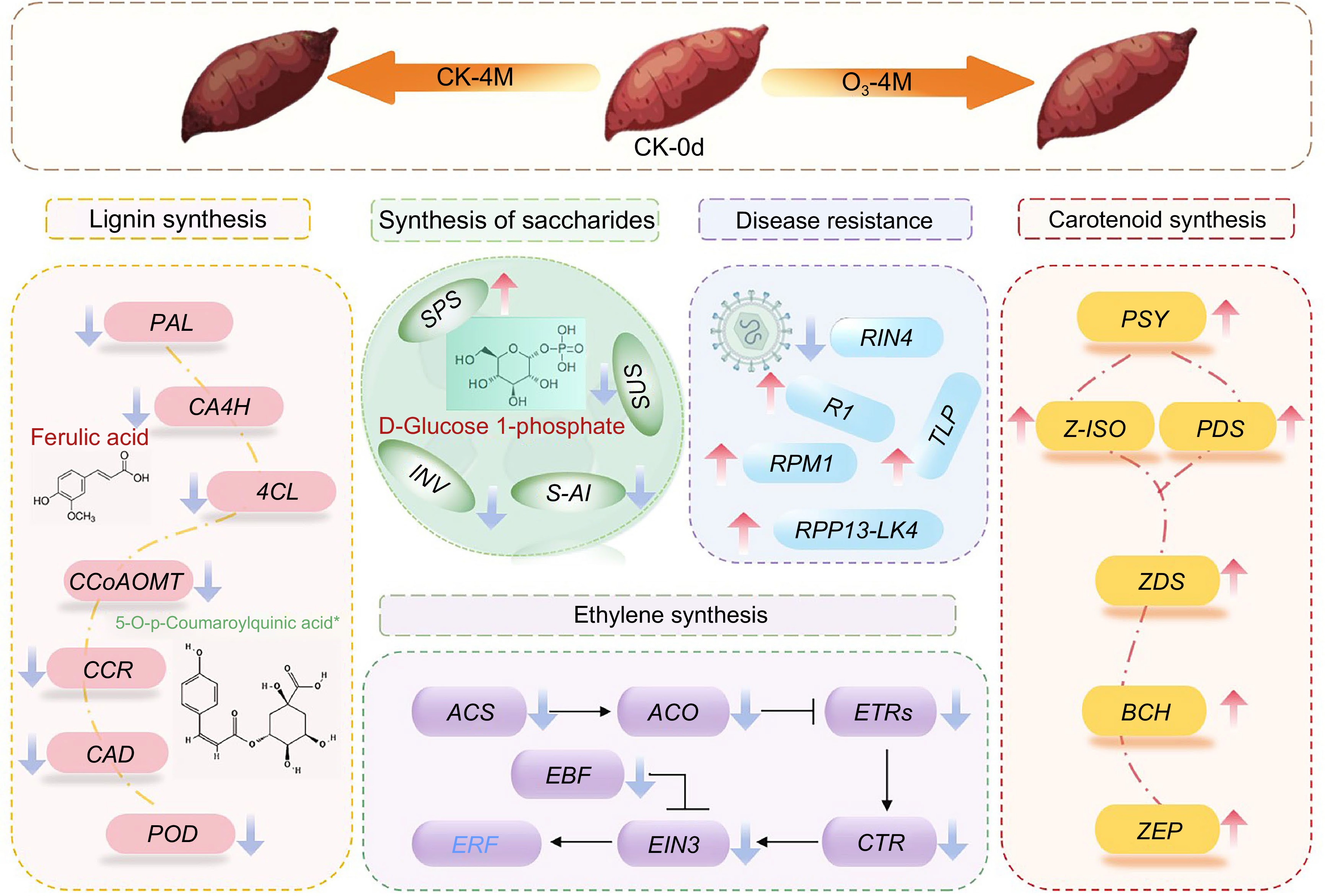
Figure 8.
Mechanistic model diagram of postharvest senescence mechanisms in O3 treated sweet potato. Genes are indicated in italics. Upward arrows indicate upregulation of genes and downward arrows indicate downregulation of genes. Metabolites are represented by red or green characters, with red representing increased levels, and green representing decreased levels.
This work was supported by Beijing Innovation Consortium of Agriculture Research System (BAIC02-2025), Special Innovation Ability Construction Fund of Beijing Academy of Agricultural and Forestry Sciences (20240405), and the Special Foundation for Reform and Development of Institute of Agri-food Processing and Nutrition, Beijing Academy of Agricultural and Forestry Sciences (GGFA2024).
-
The authors confirm contribution to the paper as follows: study conception and design: Gao Z, Zheng Y, Wang YX, Zuo J; data collection: He X and Zhou X; analysis and interpretation of results: Sang Z, Chen S; draft manuscript preparation: Liu Y, Wang YB, Liu S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during or analyzed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yuelan Liu, Yubin Wang, Shiyu Liu
- Supplementary Table S1 DAMs analysis of lipids, phenolic acids and amino acids and derivatives.
- Supplementary Table S2 Up and down regulation of DAMs in CK-4M vs CK-0d and O3-4M vs CK-4M.
- Supplementary Table S3 Statistical table of transcriptome sequencing outputs.
- Supplementary Table S4 Analysis of DEGs associated with sweet potato storage quality.
- Supplementary Table S5 Combined analysis of DAMs and DEGs for phenylketone biosynthesis and starch and sucrose metabolism in CK-4M vs CK-0d and O3-4M vs CK-4M.
- Supplementary Fig. S1 Comparative cross section of sweet potato after storage.
- Supplementary Fig. S2 Up and down regulation of lipids in O3-4M vs CK-4M.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Wang Y, Liu S, Zuo J, Sang Z, et al. 2025. Mechanisms of ozone regulation of sweet potato storage quality. Vegetable Research 5: e022 doi: 10.48130/vegres-0025-0017 

Mechanisms of ozone regulation of sweet potato storage quality
- Received: 21 January 2025
- Revised: 06 May 2025
- Accepted: 14 May 2025
- Published online: 02 July 2025
Abstract: Ozone, as a safe and environmentally friendly sterilization method, has been applied in the field of fruit and vegetable preservation. However, its preservation effects and underlying mechanisms on postharvest sweet potatoes remain unclear. This study revealed the mechanism of ozone treatment in preserving sweet potatoes through transcriptomics and metabolomics analysis. After ozone treatment, the relative content of lipids and phenolic acid metabolites in sweet potatoes increased, while the relative content of amino acids decreased. In addition, ozone treatment upregulated the expression of genes related to carotenoid biosynthesis (PSY, D27, ZEP, Z-ISO), sucrose metabolism (SPS, SUS, INV, S-AI), and disease resistance (RPM1, RPP13-LK4, R1-A, TLP). Conversely, it downregulated the expression of genes involved in lignin biosynthesis (PAL, CCoAOMT, CCR1, C4H, 4CL, CAD, CA4H), and ethylene biosynthesis and signal transduction (ETR2, CTR1, MATs3, ACO1, ACO3, NTF4, ACC, ERFC3, ERF4). These changes helped maintain the color, sweetness, and texture of sweet potatoes, effectively delaying their senescence.
-
Key words:
- Ozone /
- Sweet potatoes /
- Postharvest quality /
- Gene expression /
- Metabolites