









-
Clinacanthus nutans, a medicinal plant in the Acanthaceae family[1], is found throughout tropical and subtropical regions. It has significant applications in indigenous healthcare systems in Southeast Asia and southwestern China. C. nutans has various medicinal properties, including anti-inflammatory, antiviral, antioxidant, and anticancer activities, which have been largely attributed to its abundant accumulation of bioactive secondary metabolites such as polyphenols, triterpenes, and flavonoids[2]. Notably, the amount of these compounds varies across different tissues and is strongly affected by environmental factors[3]. For example, C. nutans grown in cooler temperatures has higher cytotoxicity in leaves compared with plants grown in warmer conditions[4]. Furthermore, changes in environmental conditions can lead to different morphological traits and physiological responses in C. nutans, which affect polyphenol and flavonoid content[5]. Plant hormones, such as methyl jasmonate (MeJA), are also known to play a key role in regulating the biosynthesis of secondary metabolites, including flavonoids, in medicinal plants[6]. Identifying candidate genes involved in the biosynthetic of active compounds like flavonoids and polyphenols in C. nutans is crucial for advancing molecular breeding and synthetic biology research in this species.
In plants, transcription factors (TFs) from several families, including MYB, bHLH, ERF, WRKY, NAC, bZIP, GRAS, and SPL, regulate gene expression related to stress tolerance and secondary metabolism. These TFs help plants adapt to environmental stressors and support the synthesis and accumulation of secondary metabolites that improve plant survival[7]. Among these, the bHLH family is one of the largest and plays crucial roles in plant growth and development, stress responses, and secondary metabolite biosynthesis. The basic helix-loop-helix (bHLH) is characterized by a conserved domain of about 60 amino acids[8]. This domain includes two regions: a basic region and a helix-loop-helix (HLH) region[9]. The basic region, located at the N-terminus, consists of 13–17 amino acids, typically containing around six basic residues necessary for DNA binding[10]. The HLH region, located at the C-terminus, contains approximately 40–50 amino acids, including two amphipathic alpha helices connected by a hydrophobic loop, which facilitates protein dimerization and promotes protein–protein interactions[11]. In plants, bHLHs regulate the transcription of target genes by binding to specific promoter sites, influencing processes like photomorphogenesis, flowering, and secondary metabolite biosynthesis, all of which are essential for plant adaptation and stress tolerance[12]. In Arabidopsis thaliana[13], AtICE1/AtbHLH116 binds to the CBF promoter region under cold stress, enhancing cold tolerance in plants overexpressing AtICE1/AtbHLH116. In Vitis vinifera[14], overexpression of VvbHLH1 increases flavonoid accumulation and Abscisic Acid (ABA) signaling in transgenic Arabidopsis, conferring salt and drought resistance. Accumulating evidence shows that bHLHs are key regulators that connect jasmonic acid (JA) signaling with downstream metabolic pathways. They play an important role in the specialized metabolism of medicinal plants and crops. In Artemisia annua[15], the AabHLH1 binds to E-box cis-elements in the promoters of ADS and CYP71AV1, regulating artemisinin biosynthesis. Furthermore, AabHLH113 integrates JA and ABA signaling, positively affecting artemisinin synthesis[16]. In Catharanthus roseus, CrMYC2, CrBIS1, and CrRMT1 regulate different branches of the monoterpenoid indole alkaloid (MIA) pathway, influencing MIA biosynthesis and accumulation[17]. In Cucumis melo, CmeBrp has been shown to regulate the biosynthesis of triterpenoid cucurbitacin compounds in roots[18]. These studies provide valuable insights into the functional roles of bHLHs in plants.
In recent years, bHLHs have been identified and analyzed on a genome-wide scale in medicinal plants such as Salvia miltiorrhiza[19], Andrographis paniculata[20], and Gynostemma pentaphyllum[21]. However, most research has primarily focused on model plants and crops[22]. To further the study of the bHLHs in medicinal plants, the first genome-wide identification of bHLH gene family members (CnbHLHs) in the genome of C. nutans was conducted. Using bioinformatics tools, the CnbHLHs were analyzed and visualized, examining their phylogenetic relationships, sequence features, conserved motifs, gene structure, cis-regulatory elements, and evolutionary history. Additionally, transcriptome data were used to study the expression profiles of CnbHLHs in different tissues and under MeJA treatment to understand their functional differentiation and regulatory patterns across developmental stages. Finally, qRT-PCR was performed to investigate the expression profiles of 20 CnbHLHs, providing insights into their roles under abiotic stress. This study offers a thorough analysis of CnbHLHs, presenting candidate genes for exploring the molecular mechanisms of abiotic stress responses and developmental processes in this species.
-
The complete genome sequence of C. nutans was generated by the current research group. Two approaches were used to identify all potential bHLH genes. First, the hidden Markov model (PF00010) of the bHLH domain was downloaded from the Pfam database[23] (
http://pfam.xfam.org ) and the HMMER web server[24] was employed to search candidate sequences within the C. nutans protein database, setting an E-value threshold of 10−5. Second, the AtbHLH protein sequences of A. thaliana were retrieved from TAIR (www.arabidopsis.org ) to construct a BLAST comparison database, and a second round of candidate sequence screening conducted using the BLASTP program[25], setting the E-value at 10−5. All candidate protein sequences were subsequently merged, and redundancies were removed. To confirm that these candidate genes belong to the bHLH family, all candidate sequences were further analyzed using the NCBI Conserved Domain Database[26] (www.ncbi.nlm.nih.gov/cdd ), and the SMART database[27] (http://smart.embl-heidelberg.de ), ultimately identifying 182 non-redundant sequences as putative bHLH proteins of C. nutans (Supplementary Table S1).Classification and phylogenetic analysis
-
All identified CnbHLHs were renamed sequentially according to their chromosomal distribution in C. nutans. Protein physicochemical properties, including molecular weight, isoelectric point, and grand average of hydropathicity (GRAVY), were predicted using the Peptides package[28]. Subfamily classification of CnbHLHs followed the methodology proposed by Pires[29], which organizes 544 representative bHLH sequences from major plant evolutionary lineages into 26 subfamilies. The combined dataset of 182 CnbHLHs and 162 AtbHLHs was used for phylogenetic inference. Sequence alignment of the combined protein dataset was performed using MUSCLE[30], and phylogenetic tree construction was conducted using IQ-TREE[31] with the maximum likelihood (ML) method under default parameters. Visualization and annotation of the phylogenetic tree were completed using the Evolview v3 online platform[32].
Gene structure, conserved motif, and cis-regulatory elements (CREs) analysis
-
The conserved motifs of CnbHLHs were analyzed using the MEME online suite[33] (
http://meme-suite.org ), with optimized parameters allowing any number of repeats, a maximum of 10 motifs, and an optimal motif width ranging from six to 200 amino acid residues. The intron-exon structural organization of C. nutans bHLH genes was annotated based on the GFF file of the C. nutans genome. The 2000 bp upstream sequences of each CnbHLH were extracted from the full-length genomic DNA using SeqKit[34], and subsequently uploaded to the PlantCARE database for cis-regulatory element prediction[35]. The prediction results were categorized in Excel, and visualized using R[36].Chromosomal location and gene duplication analysis
-
Based on the physical location information from the C. nutans genome database, the anchoring relationships between all CnbHLHs and chromosomes were visualized using the ChromoMap package. To illustrate synteny both within C. nutans and across species, homologous bHLHs were identified from genomic data of C. nutans and other selected species. The analysis included four species: A. thaliana (
www.arabidopsis.org ), V. vinifera (http://plants.ensembl.org ), A. paniculata (www.ncbi.nlm.nih.gov ), and Strobilanthes cusia (http://indigo-plant.iflora.cn ), with genomic data obtained from the corresponding database websites. Gene duplication events were analyzed using MCScanX with default parameters[37], and the results were visualized in the Advanced circos module of TBtools[38]. For Ka/Ks analysis, nonsynonymous substitution rate (Ka), synonymous substitution rate (Ks), and Ka/Ks ratios between homologous gene pairs in C. nutans and other species were calculated using the KaKs calculator[39].Prediction of protein–protein interaction network and homology modeling analysis
-
All submitted CnbHLH protein sequences were uploaded to the STRING website[40] (
http://string-db.org ), and Arabidopsis orthologs were chosen as reference sequences. After the BLAST analysis, the highest-scoring gene (bitscore) was used to construct the network, and genes without interactions with others were eliminated. Protein models homologous to the CnbHLHs were retrieved from the PDB database[41] (www.rcsb.org ). Protein secondary structures were then predicted through NetSurfP-3.0[42] (https://services.healthtech.dtu.dk/services/NetSurfP-3.0 ), and SWISS-MODEL (www.swissmodel.expasy.org ) was used for homology modeling to predict the tertiary structure[43]. Additionally, ConSurf (https://consurf.tau.ac.il ) was employed to examine conserved regions. Finally, the predicted model structures were visualized and refined using PyMOL[44].Plant culture and stress treatments
-
The C. nutans utilized in this study were cultivated at Fujian Agriculture and Forestry University (FAFU) (Fujian, China). Approximately 30 cm-long stem cuttings were obtained from tall and vigorous mother plants and propagated through stem cuttings. Once rooted and showing vigorous growth, each plant was individually transplanted into pots filled with a soil and peat mixture at a ratio of 2:1. Plants were grown under greenhouse conditions for 60 d (28 °C, 75% relative humidity, and a photoperiod of 16 h light/8 h dark). At the beginning of the treatments, plants of uniform growth status were randomly grouped for stress and hormone treatments, with three biological replicates per group. The experimental design included five treatment conditions: drought, salinity, cold, heat, and MeJA. Drought stress was simulated using 150 mmol/L mannitol solution, and salinity stress was induced by 150 mmol/L NaCl solution. For temperature stress, plants were exposed to 8 °C (cold) and 45 °C (heat). MeJA treatment was performed by foliar spraying with 200 μmol/L MeJA solution until leaf surfaces were fully covered and dripping. Samples were collected from roots and leaves at four time points (0, 24, 48, and 72 h) post-treatment. Additionally, organ-specific samples (root, stem, young leaf, mature leaf, and flower) were collected from flowering plants under normal growth conditions. All samples were collected in three biological replicates, immediately frozen in liquid nitrogen, and stored at −80 °C for further analysis.
Transcriptomic and qRT-PCR analyses
-
Transcriptome sequencing was performed on organ-specific samples (root, stem, young leaf, mature leaf, and flower) as well as MeJA-treated root and leaf samples collected at 0, 24, 48, and 72 h. All RNA-seq experiments, including RNA extraction, library construction, and sequencing, were conducted by Biomarker Technologies Co., Ltd (Beijing, China). Approximately 6.7 Gb of raw sequencing data were generated per sample and subsequently assessed using FastQC. Low-quality reads and adapter sequences were removed using fastp[45], resulting in high-quality clean reads. Clean reads were aligned to the C. nutans reference genome using HISAT2[46]. Transcript abundance was quantified using the featureCounts program in the Subread package[47]. Expression levels of CnbHLHs were normalized as TPM (Transcripts Per Kilobase Million), and a heatmap based on log2(TPM + 1) was generated using R. All RNA-seq data have been deposited in the China National GeneBank DataBase (CNGBdb) under accession number: CNP0006103.
To assess the expression of CnbHLHs under abiotic stress, qRT-PCR was performed using samples treated with cold (4 °C), heat (45 °C), drought (150 mmol/L mannitol), and salinity (150 mmol/L NaCl). Untreated plants served as the control group. Samples were collected from roots and leaves at 0, 24, 48, and 72 h post-treatment. Total RNA was extracted using the Trizol method (Invitrogen, Carlsbad, CA, USA), and first-strand cDNA synthesis was carried out using the ThermoScript RT-PCR Kit (Thermo Fisher Scientific, USA). Gene-specific primers were designed using Primer-BLAST, and primer sequences are listed in Supplementary Table S2. The qRT-PCR program was: 95 °C for 30 s; followed by 40 cycles of 95 °C for 5 s, 60 °C for 34 s, and 95 °C for 15 s. The CnRPL was used as an internal control to normalize expression levels[48]. The fold change in gene expression was calculated using the 2−ΔΔCᴛ method.
-
A total of 182 putative bHLH family members were identified in the C. nutans genome, which were sequentially named from CnbHLH001 to CnbHLH182 based on their chromosomal positions. To gain a comprehensive understanding of the physicochemical properties of the CnbHLH proteins, the predicted characteristics of the proteins encoded by each CnbHLH gene were analyzed (Fig. 1, Supplementary Table S3). The results indicated that the predicted protein lengths of all CnbHLHs ranged from 82 (CnbHLH118) to 1,074 (CnbHLH078) amino acids, with molecular weights spanning 9.39 to 118.90 kDa. The isoelectric points (pI) varied from 4.47 (CnbHLH020) to 12.29 (CnbHLH100); of these, 88 members predominantly contained basic amino acids (pI > 7), while 93 members were primarily composed of acidic amino acids (pI < 7), with CnbHLH175 exhibiting a pI of 7, representing a neutral protein. The GRAVY values ranged from −0.94 to 0.1, with the majority being hydrophilic proteins; only CnbHLH088 (0.02) and CnbHLH136 (0.1) exhibited a tendency toward hydrophobicity. The Instability Index values ranged from 27.12 (CnbHLH108) to 98.91 (CnbHLH087), with 166 (91.21%) members exceeding a value of 40, indicating that most proteins are relatively unstable and may be more susceptible to structural changes or degradation in vitro.
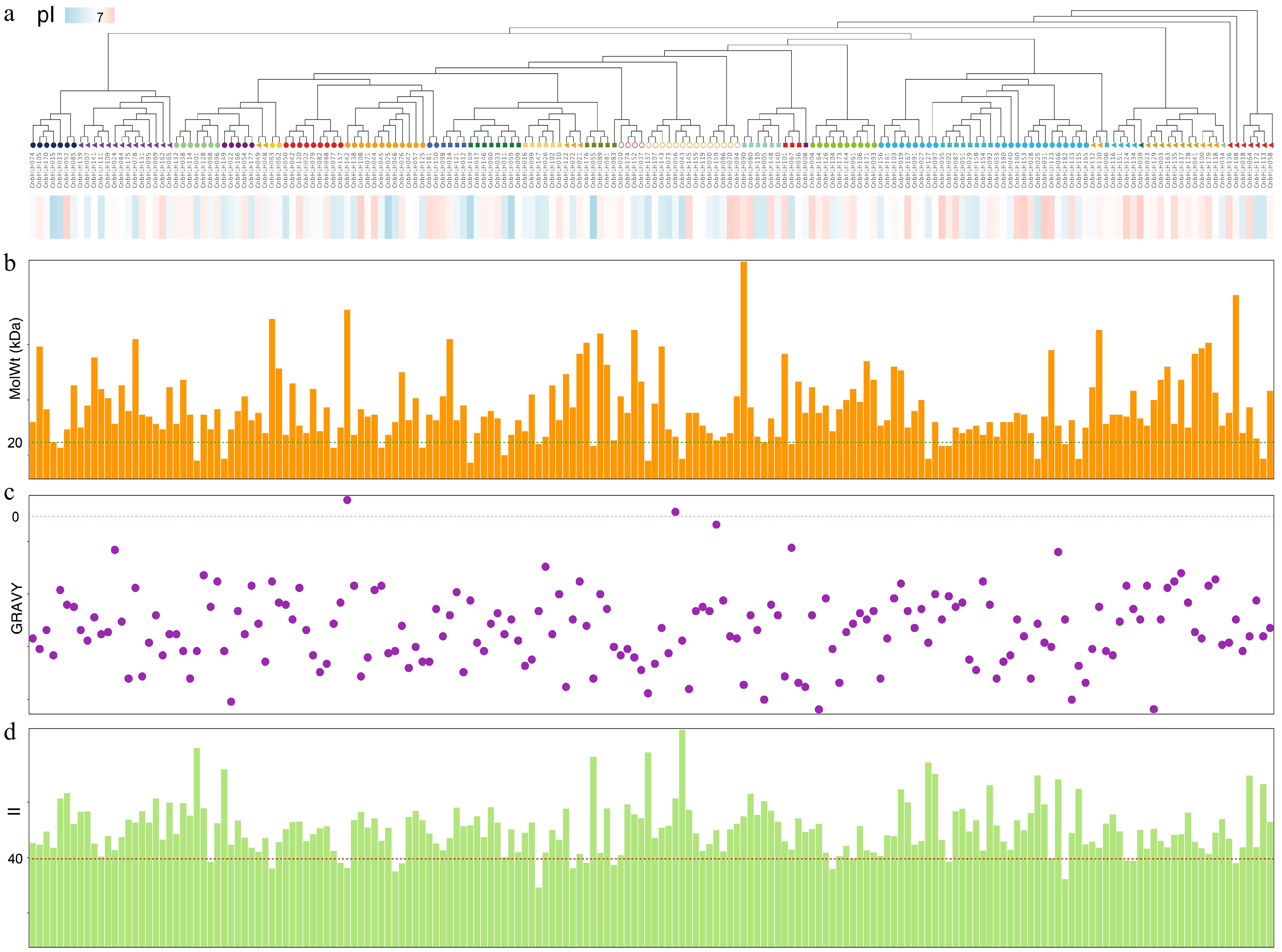
Figure 1.
Physicochemical properties of CnbHLHs. (a) Phylogenetic tree of the CnbHLHs constructed based on full-length amino acid sequences, with subfamily clades color-coded. The bar below indicates the isoelectric point (pI) values of each protein, ranging from acidic (blue) to basic (red). (b) Bar plot showing the molecular weight (MolWt) distribution of CnbHLHs in kilodaltons (kDa). (c) GRAVY (Grand Average of Hydropathy) index of each CnbHLH. Most proteins exhibited negative GRAVY values, indicating hydrophilicity. (d) Bar chart of instability index (II) values. The dashed line at II = 40 denotes the threshold above which proteins are considered potentially unstable.
An unrooted maximum likelihood (ML) phylogenetic tree, based on the complete protein sequences of 182 CnbHLHs from C. nutans and 162 AtbHLHs from A. thaliana, clearly depicted the evolutionary relationships among the bHLH proteins. A total of 172 CnbHLHs from C. nutans were classified into 26 subfamilies, and the remaining 10 were grouped as Orphans, showing no significant similarity or evolutionary relationship with other family members (Fig. 2). The tree's topology revealed considerable diversity and variation in the distribution of CnbHLHs across subfamilies. Subfamily XII with 23 genes, accounting for 12.64% of all CnbHLHs. Subfamilies Ib(2) and X had 10 or more members, while subfamilies VIIIa, VIIIc(1), and XIV each had only one member. Most subfamilies (20, comprising 76.92% of all subfamilies) had between two and 10 members.
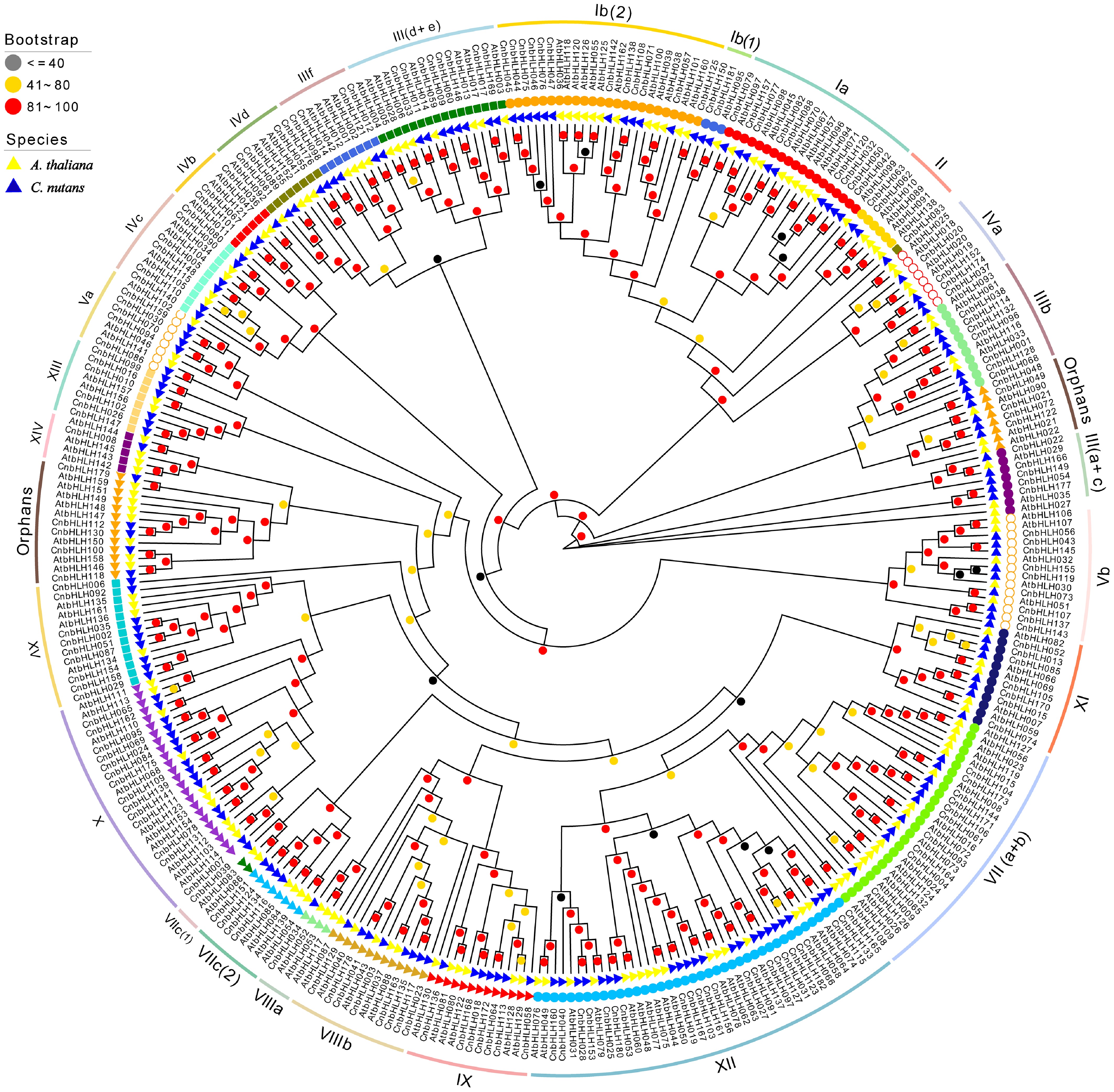
Figure 2.
Phylogenetic relationship of bHLHs between C. nutans and A. thaliana. The unrooted evolutionary tree was constructed using IQTree with the maximum likelihood (ML) method. The 182 CnbHLHs are classified into 26 subfamilies and one Orphan group, following the classification method used of AtbHLHs, represented by different colored subgroups within the tree. The bootstrap value is displayed a circle on each branch, with each color corresponding to a specific bootstrap value range.
Chromosomal localization analysis showed that CnbHLHs were distributed across all 18 chromosomes (Chr) of C. nutans. Visualization showed an uneven distribution of CnbHLHs on these chromosomes, with a tendency for clustering at the ends (Supplementary Fig. S1a). Ten chromosomes contained 10 or more CnbHLHs, and the remaining eight chromosomes had fewer than 10 members. Chr7 had the highest number, with 16 CnbHLHs, whereas Chr8 and Chr12 each contained only four members. Also, certain chromosome pairs shared the same number of CnbHLHs, including Chr1 and Chr18, Chr2 and Chr16, Chr3 and Chr14, Chr4 and Chr6, Chr8 and Chr12, Chr9 and Chr15, and Chr10 and Chr17 (Supplementary Table S3). Considering the subfamily classifications of CnbHLHs, no specific subfamily members were found to cluster on a single chromosome (Supplementary Fig. S1b), indicating a random distribution that may reflect their roles across different biological functions and regulatory networks.
Conserved domains and motifs, gene structure of CnbHLHs
-
To further explore gene structure in different subgroups, CnbHLHs gene structures were analyzed and visualized, including conserved domains, motifs, and exon–intron compositions. The bHLH domains in C. nutans ranged from 37 to 88 residues, with gaps inserted to optimize alignment across conserved regions. Multiple sequence alignment identified 113 conserved positions in the bHLH domain (Fig. 3a, Supplementary Table S4). Positions 1–13 corresponded to the basic amino acid region, 14–55 to the helix1 region, 56–71 to the loop region, and 72–113 to the helix2 region. Conserved residues, such as H5/Q5, E9/A9, and 13R, essential for E-box binding, matched previous reports[49] (Fig. 3b). Residues 10R and 12R showed significant conservation, with alignment rates of 75.80% and 86.30%, respectively. In helix1, P55—a critical residue for terminating the first helix and linking to the variable-length loop—was conserved (86.30%). Several hydrophobic residues, including I31, R34, L38, L41, and V54, were also conserved, contributing to the stability of the bHLH dimeric structure (Fig. 3c). In the loop region, two conserved residues, K69 and D71, were found. In helix2, several hydrophobic residues (A73, L76, A79, I86, L92, V96, and L113) were conserved, along with D71, S74, E78, and Y88, highlighting the structural stability of these domains.
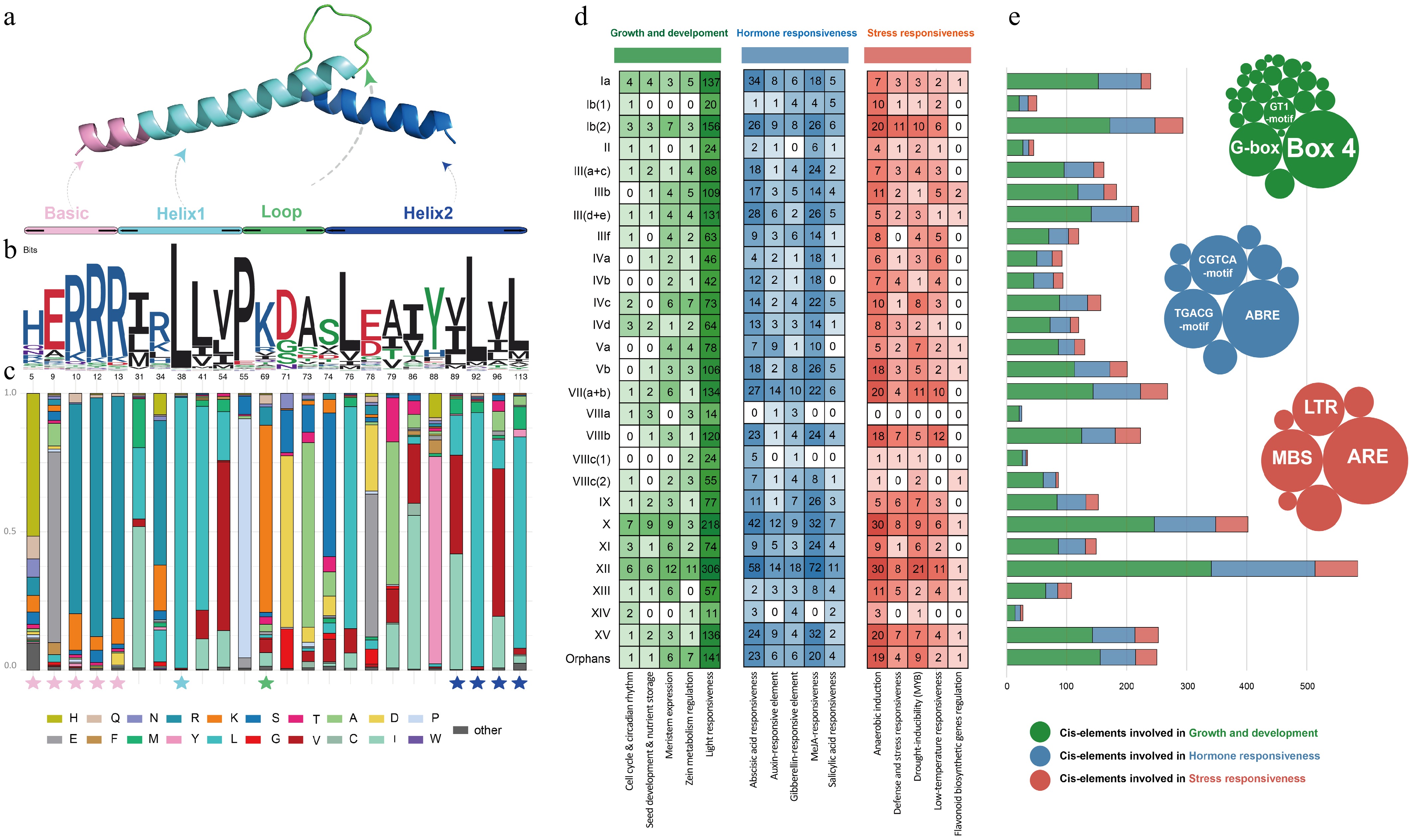
Figure 3.
Structural characteristics and CRE feature of CnbHLHs. (a) Three-dimensional protein structure of the bHLH domain. (b) Sequence logo of the CnbHLHs domain. The numbers at the bottom represent the positions of residues in the alignment. (c) Distribution of amino acids in the CnbHLHs conserved sites. The pentagrams below the bar chart indicate the positions of highly conserved residues. (d) The identified CREs were grouped into three functional categories, shown in green, blue, and red shades. The numbers show the quantity of corresponding CREs in the promoter regions of each subfamily of CnbHLHs. (e) Statistics for the total number of CREs. The bubble chart displays the number of all elements under each functional category, with the top three most abundant elements labeled.
Motifs are important for mediating interactions and signal transduction between different modules during gene transcription[50]. To investigate these motifs, MEME was used to analyze the conserved motifs in CnbHLHs. The composition patterns of the 10 identified motifs were largely consistent with the results of the present phylogenetic tree and gene structure analysis, showing high similarity within the same subfamily but significant differences between subfamilies (Supplementary Fig. S2b, Supplementary Table S5). The number of motifs in the 182 CnbHLHs ranged from one (CnbHLH100, CnbHLH112, CnbHLH125) to seven (CnbHLH060, CnbHLH098, CnbHLH121). For motif distribution, motif 1 and motif 2 were present in 97.25% and 98.90% of CnbHLHs, respectively. Except for five orphan genes (CnbHLH130, CnbHLH048, CnbHLH049, CnbHLH100, CnbHLH112), all other CnbHLHs contained motif 1. CnbHLH006 and CnbHLH125 were the only two genes lacking motif 2. Additionally, all members of subfamily XII contained motif 3, and all members of subfamily XIII contained motif 6. Notably, certain motifs were exclusive to specific groups; for example, motif 10 was found only in group VIIIb, while the combination of motif 7 and motif 8 was combined only in group III. This pattern reflects the conservation and specificity of motifs within particular subfamilies.
The diversity of exon-intron structures is critical to the evolution of multigene families[51]. The number of introns and exons in each gene were analyzed and subsequently visualized the gene structural characteristics of the CnbHLHs (Supplementary Fig. S2c, Supplementary Table S5). Among the 182 CnbHLHs, the number of exons ranges from one to 22, with 167 CnbHLHs (91.76%) containing multiple exons, and 30 genes (16.48%) with three exons, representing the highest proportion. Additionally, 15 CnbHLHs (8.24%) were intron-less, predominantly found in subfamilies III(d + e) and VIIIb, such as CnbHLH059, CnbHLH017, and CnbHLH100. Structural diversity was evident among members of different subfamilies, including variations in intron and exon counts and their relative positions. Conversely, genes within the same subfamily exhibit highly similar intron/exon patterns; for instance, members of subfamily Ia contain 2–3 exons, while those in subfamily III(a + c) contain 4–5 exons, and nearly all members (22/23) of subfamily XII have 5–9 exons. These findings align with the general structural characteristics observed in most plant bHLH family genes.
Cis-regulatory elements (CREs) analysis of CnbHLHs
-
Conserved motifs located in gene promoter regions act as recognition and binding sites for proteins. Using PlantCARE, the putative promoter sequences of 182 CnbHLHs were analyzed, identifying 4,672 CREs belonging to 42 types (Supplementary Table S6). These CREs were divided into three main groups based on function: plant growth and development (26 types, 2,765 CREs), plant hormone response (nine types, 1,282 CREs), and stress response (seven types, 625 CREs) (Fig. 3d). Among growth-related elements, light-responsive elements were the most common (2,504, accounting for 90.56%), widely distributed across nearly all CnbHLHs' promoter region (e.g., CAG-motif, GT1-motif, GATA-motif, G-box, ACE, and Box I). For hormone response elements, MeJA-responsive elements (508, 39.63%) and abscisic acid (ABA)-responsive elements (437, 34.09%) were the most abundant, showing enrichment in subfamily XII. In stress response elements, anaerobic induction elements (293, 46.88%) were the most frequent, with ARE elements being the most prominent in all subfamilies (Fig. 3e). Overall, the composition and number of CREs of different CnbHLHs vary significantly between and within subfamilies, suggesting that the expression of CnbHLHs is regulated by a diverse set of CREs associated with hormones, growth, development, and stress responses.
Gene duplication and synteny analysis of CnbHLHs
-
Gene duplication events are important for gene family evolution, with family expansion usually driven by tandem and segmental duplications. To understand the amplification mechanisms of CnbHLHs, gene duplication events in C. nutans were analyzed, revealing 97 gene duplication pairs (Fig. 4a, Supplementary Table S7). Among them, five pairs were tandem duplications. CnbHLH044–CnbHLH047 formed three consecutive tandem duplication pairs on Chr4, with one additional pair on Chr6 and Chr7. All these duplicated genes belong to the same subfamily. The other 92 pairs were segmental duplications, suggesting that segmental duplication plays a key role in CnbHLHs expansion and evolution. Notably, only three segmental duplication pairs are on the same chromosome: CnbHLH139/CnbHLH141 (Chr15), CnbHLH154/CnbHLH158 (Chr16), and CnbHLH156/CnbHLH161 (Chr16). These inter-chromosomal segmental duplications suggest recent chromosomal rearrangements in this species, defining potential hotspots for further rearrangements.
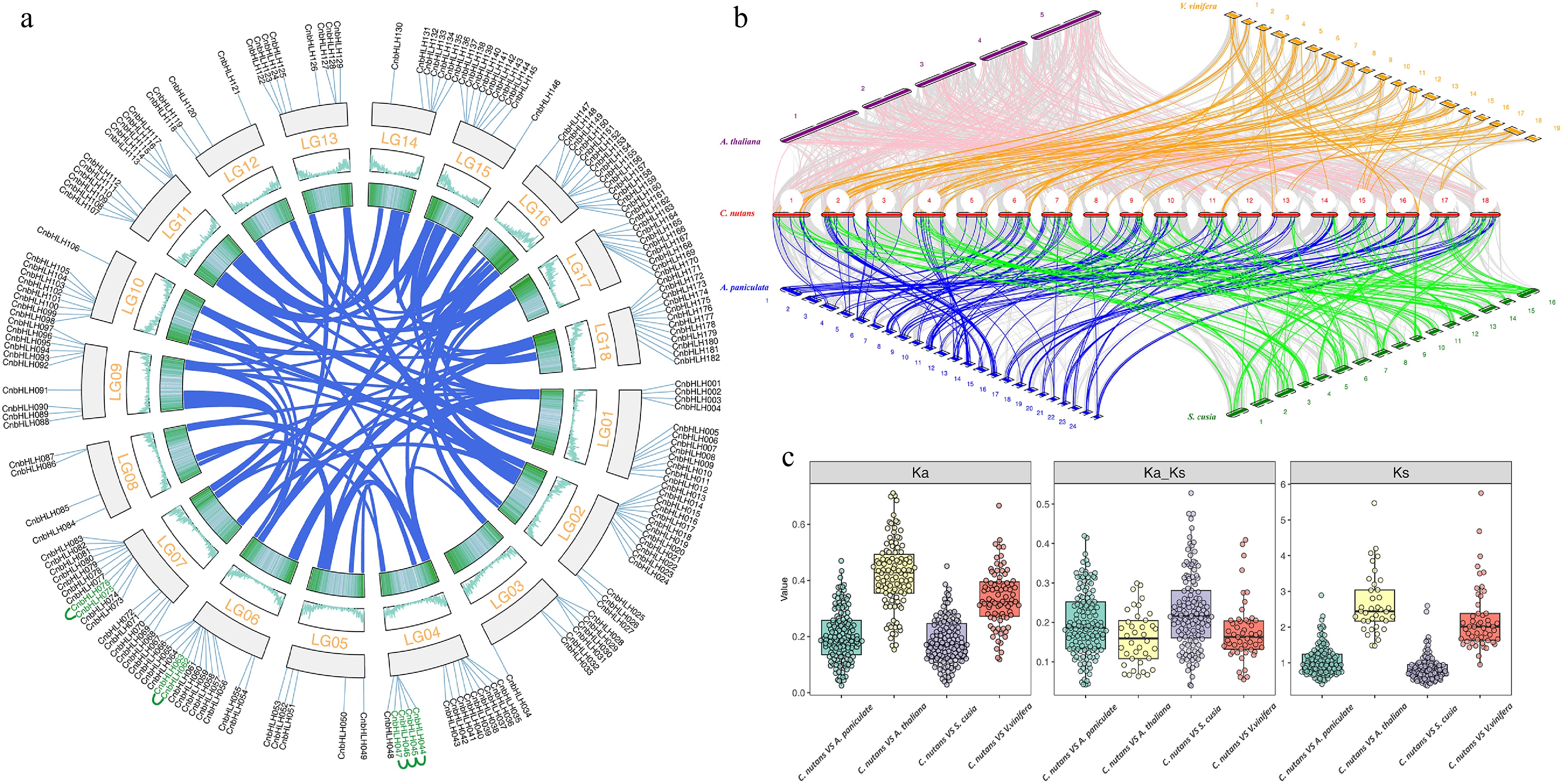
Figure 4.
Evolutionary analysis of the bHLH gene family between C. nutans and other species. (a) Collinearity analysis of CbHLHs. (b) bHLHs collinearity between C. nutans and other species (A. thaliana, V. vinifera, A. paniculata, S. cusia) genomes. (c) Ka/Ks analysis of bHLHs between C. nutans and closely related species (A. paniculata, S. cusia).
To investigate the evolutionary trajectory of family members, four representative species were selected for synteny analysis with C. nutans (Fig. 4b, Supplementary Table S8): A. thaliana (Brassicacea), V. vinifera (Vitaceae), and two closely related species, A. paniculata and S. cusia. A total of 138 CnbHLHs exhibited synteny with A. paniculata, followed by 129 CnbHLHs with S. cusia, 106 CnbHLHs with A. thaliana, and 82 CnbHLHs with V. vinifera. Notably, 37 CnbHLHs displayed syntenic relationships with all four species. Among these, 102 syntenic genes were shared across the three Acanthaceae species, while only 53 and 37 genes showed synteny with V. vinifera and A. thaliana, respectively. This suggests possible lineage-specific expansions within Acanthaceae, potentially linked to specific evolutionary or adaptive needs of the family. Furthermore, to explore the forces driving gene evolution, a Ka/Ks analysis of homologous genes between C. nutans and the four species was performed (Fig. 4c, Supplementary Table S9). The frequency of nonsynonymous mutations was lower than that of synonymous mutations, with Ka/Ks ratios below 1, indicating that these CnbHLHs underwent strong purifying selection during evolution.
Protein–protein interaction networks analysis
-
bHLH proteins typically function by forming homodimers or heterodimers, either among bHLH proteins or between bHLH and non-bHLH proteins. A protein interaction network was constructed for CnbHLHs based on collinearity, phylogenetic analysis, and alignment scores, using homologous proteins from A. thaliana as references (Fig. 5a, Supplementary Table S10). This network includes 408 interactions, with 90 showing high reliability (combined score > 0.7). These 408 interactions form 59 nodes. Among these nodes, only one protein pair shows a single interaction mode (CnbHLH053 and CnbHLH093), while other nodes have numerous connections (> 2) to various nodes, indicating complex multi-gene interactions. CnbHLH038 has the most interaction partners (16), though only three interactions have high binding scores: CnbHLH038–CnbHLH058 (0.852), CnbHLH038–CnbHLH079 (0.761), and CnbHLH038–CnbHLH153 (0.852). The CnbHLH168 node shows the highest interaction reliability, with five of eight interactions having high reliability: CnbHLH168–CnbHLH059 (0.72), CnbHLH168–CnbHLH175 (0.758), CnbHLH168–CnbHLH139 (0.881), CnbHLH168–CnbHLH001 (0.719), and CnbHLH168–CnbHLH121 (0.777). CnbHLH075, CnbHLH155, CnbHLH176, CnbHLH139, CnbHLH141, CnbHLH175, CnbHLH008, CnbHLH036, CnbHLH090, and CnbHLH140 each has more than 10 interactions, suggesting they may play key roles in specific functions or pathways. These findings further highlight the functional diversity of CnbHLHs.
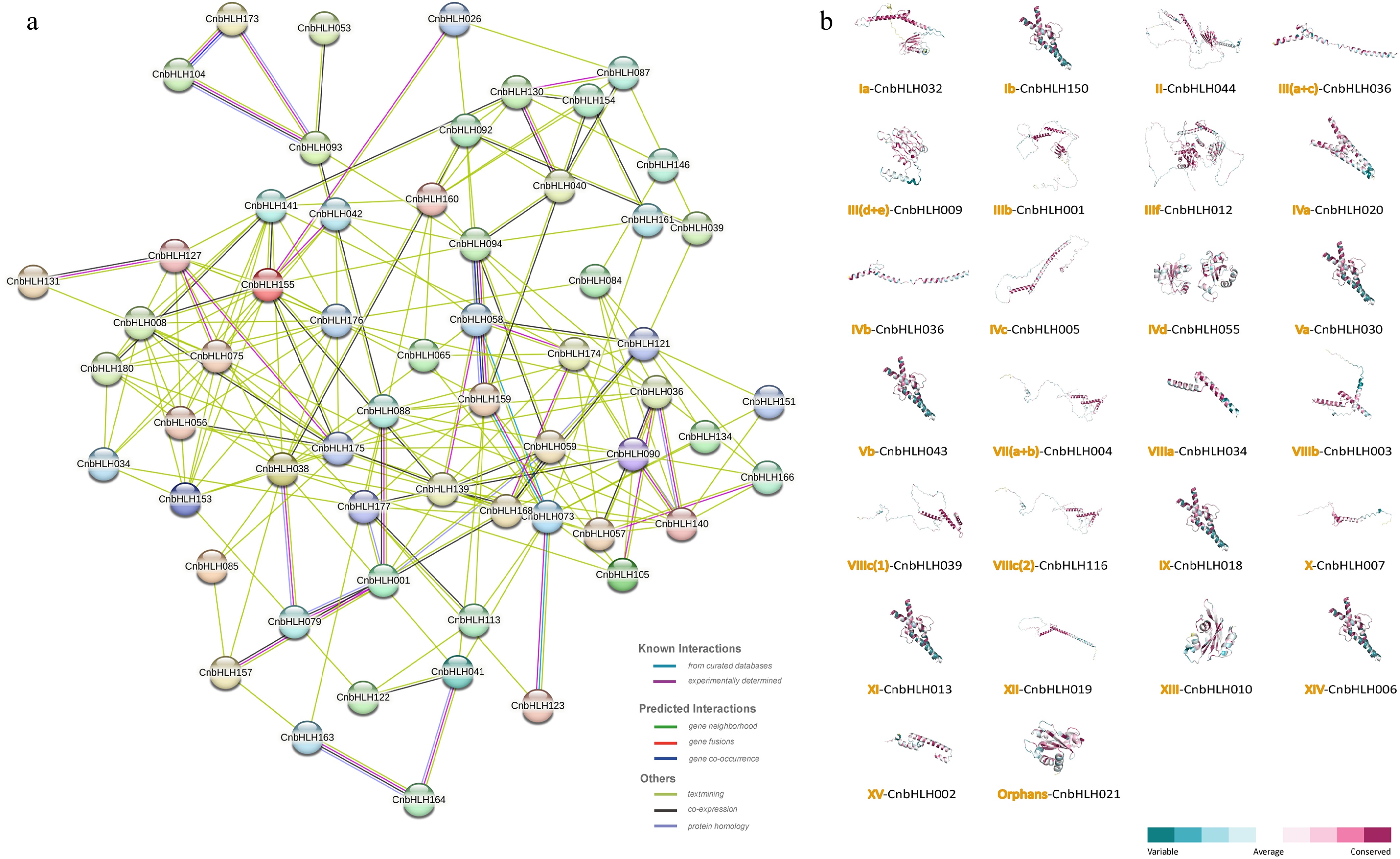
Figure 5.
Protein interaction network and predicted 3D structure of CnbHLHs. (a) Protein–protein interaction network of CnbHLHs based on homologous proteins in A. thaliana. (b) Predicted 3D structures of CnbHLHs, showing only the member with the highest GMQE value from each subfamily.
Secondary structure prediction results show that random coil structures dominate in CnbHLHs (35.19%–91.90%), while α-helix (7.48%–56.64%), and β-sheet (0.13%–20.24%) contents are relatively lower (Supplementary Table S11). The disorder scores of residues in these proteins range from 11.52 to 86.57, suggesting that CnbHLHs have structural flexibility while maintaining stability. Moderate Relative Solvent Accessibility (RSA) values (36.46–64.68) suggest that these proteins are involved in functional interactions. Subsequently. Homology modeling of CnbHLHs was performed using the SWISS-MODEL database, predicting three-dimensional structures for 182 CnbHLHs. CnbHLHs in different subfamilies showed diverse three-dimensional structures, while proteins in closely related subfamilies had similar 3D structures. The structure with the highest GMQE and QMean scores in each subfamily was chosen as the representative model for CnbHLHs (Fig. 5b, Supplementary Table S12). Subfamily XV showed the highest GMQE value (0.8), indicating high model reliability. Subfamilies Ib(1) and III(a + c) had multiple genes with GMQE values below 0.1, suggesting lower model reliability and the need for further optimization and validation. Overall, these homology models provide a basis for understanding the molecular functions of CnbHLHs.
Expression analysis
-
To investigate the functions of CnbHLHs, the expression profiles of 182 CnbHLHs across different tissues of C. nutans, including root, stem, young leaf, mature leaf, and flower, were analyzed using RNA-seq data (Fig. 6a, Supplementary Table S13). After filtering out genes with low expression levels (TPM < 30), 89 CnbHLHs were retained for further analysis. Many of these genes showed tissue-specific expression, suggesting that CnbHLHs are involved in tissue- or developmental stage-specific functions. Hierarchical clustering grouped the CnbHLHs into five blocks (1–5), with genes in each block showing unique expression patterns: (1) CnbHLHs in block a were mainly expressed in the root, with CnbHLH015, CnbHLH074, CnbHLH030, and CnbHLH017 also showing high expression in other tissues. (2) Most CnbHLHs in block b were highly expressed in mature leaves, while CnbHLH011, CnbHLH172, CnbHLH061, and CnbHLH008 exhibited high expression in all tissues. (3) CnbHLHs in block c were mainly expressed in young leaves, with CnbHLH101 and CnbHLH059 also showing higher expression in other tissues. (4) CnbHLHs in block d were mainly expressed in flowers, with CnbHLH148, CnbHLH110, CnbHLH126, CnbHLH130, and CnbHLH140 also showing high expression in other tissues. (5) CnbHLHs in block e were highly expressed in the stem, with more than half also showing abundant expression in the leaf. These results also highlight that different CnbHLHs have diverse expression profiles within and between subfamilies.
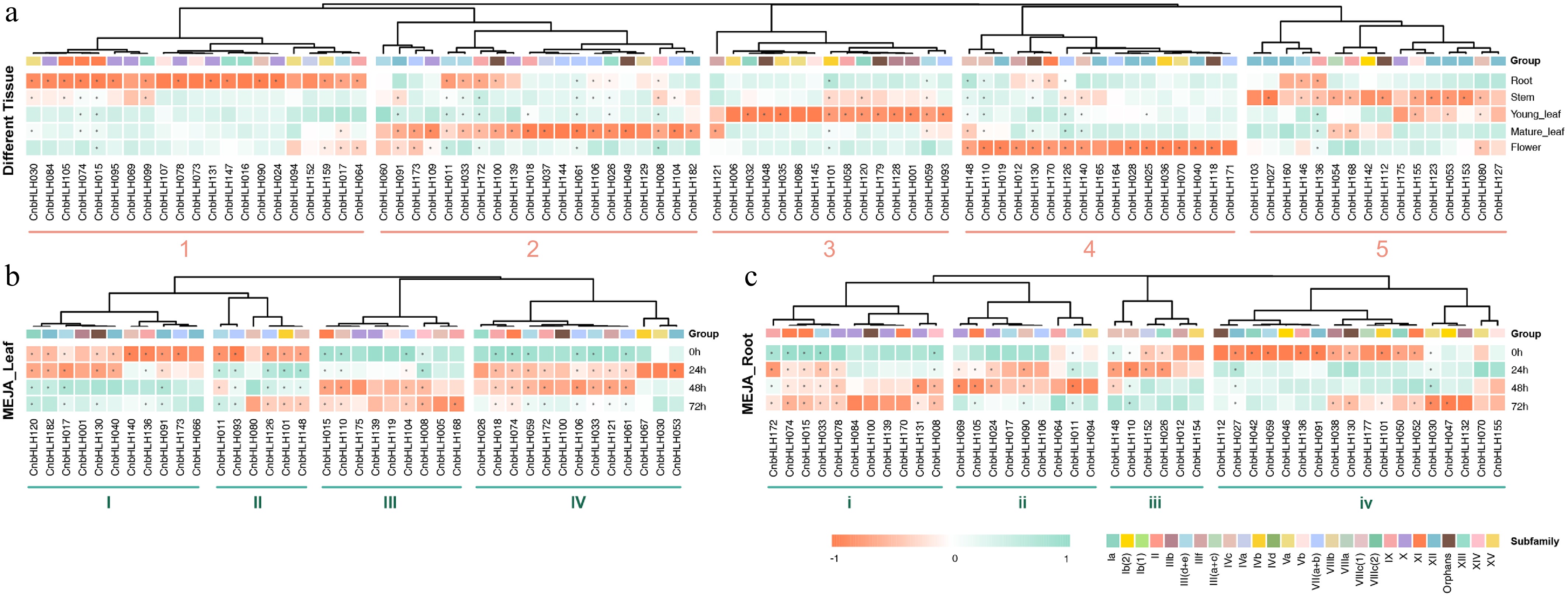
Figure 6.
The heat maps of gene expression of CnbHLHs in different tissues and the MeJA treatment. (a) The heat map of CnbHLHs expressionin different tissues. (b) The heat map of CnbHLHs expression in leaves after MeJA treatment. (c) The heat map of CnbHLHs expression in roots after MeJA treatment.
Studies show that MeJA treatment can modulate plant metabolic processes, influencing the synthesis and accumulation of bioactive compounds like flavonoids and triterpenes in medicinal plants[52]. Previous studies show that bHLHs play important roles in jasmonic acid (JA) signaling pathways[53]. To explore the roles of CnbHLHs in C. nutans in response to MeJA, the expression patterns of these genes in the leaves and roots were analyzed at different time points after MeJA treatment using transcriptome data. After filtering out genes with low expression, 39 genes remained in the leaf group (Fig. 6b), and 44 genes remained in the root group (Fig. 6c). Notably, the gene composition in the two groups differed, with only 19 genes in both tissues, suggesting distinct CnbHLHs responses to MeJA treatment in each tissue. The remaining CnbHLHs that were differentially expressed in the leaves under MeJA treatment were grouped into four groups (blocks I–IV) based on hierarchical clustering (Fig. 6b). Genes in block I were highly expressed at the pre-treatment (0 h) and early treatment (24 h) stages, with expression decreasing over time. In contrast, most genes in block II showed high expression across all time points, with CnbHLH011 and CnbHLH093 mainly expressed before treatment, while CnbHLH126, CnbHLH101, and CnbHLH148 showed high expression at both the pre-treatment and final time points (72 h), following a valley-shaped pattern. Genes in block III were mainly expressed during the later stages of treatment (48 and 72 h), while genes in block IV peaked at the mid-treatment stages (24 and 48 h). In the roots, the remaining CnbHLHs were grouped into four clusters (blocks i–iv) (Fig. 6c). Genes in block i were upregulated by MeJA and showed higher expression at the later time points (48 and 72 h). Genes in block ii were mainly expressed during the mid-treatment period (24 and 48 h), showing a peak-shaped expression pattern. Genes in block iii showed high expression at the pre-treatment (0 h), and early treatment (24 h) time points, with expression decreasing later. Most genes in block iv were mainly expressed before treatment (0 h), with their expression suppressed after MeJA induction; however, genes like CnbHLH038, CnbHLH130, CnbHLH177, CnbHLH101, CnbHLH050, and CnbHLH052 also showed higher expression later. A small subset of genes in this block, including CnbHLH030, CnbHLH047, and CnbHLH132, were mainly expressed at the later time points. These findings show that the CnbHLHs in the leaves and roots of C. nutans have diverse expression patterns in response to MeJA treatment, suggesting that different CnbHLHs may play distinct roles at various time points during treatment.
Expression patterns of CnbHLHs under abiotic stress
-
Numerous studies have shown that bHLHs play a critical role in abiotic stress response pathways, influencing the accumulation of secondary metabolites[12,54]. To explore the roles of CnbHLHs in response to various stress conditions in C. nutans, qRT-PCR was used to analyze the expression patterns of 20 representative CnbHLHs at different time points after exposure to cold (8 °C), heat (45 °C), salt (150 mM NaCl), and drought (150 mM mannitol) (Fig. 7, Supplementary Table S14). The results showed gene members from the same subfamily exhibited different response patterns to the same stress. Additionally, significant differences were observed in the expression patterns of the same genes between leaves and roots under identical treatment conditions, suggesting distinct responses in aboveground and underground tissues. Under cold stress, 15 out of the 20 CnbHLHs were significantly upregulated in the leaves, with expression initially increasing, followed by a decrease, and then an eventual increase. Several CnbHLHs showed significant upregulation at specific time points during cold stress, including CnbHLH148 (15-fold), CnbHLH18 (14-fold), CnbHLH104 (28-fold), CnbHLH126 (8-fold), CnbHLH173 (12-fold), and CnbHLH8 (14-fold). Under the same cold stress, 12 out of the 20 CnbHLHs showed decreased expression in the roots, while CnbHLH126 and CnbHLH173 were upregulated, showing an increasing expression trend. These genes were also upregulated in the leaves under cold stress, suggesting that they may play important roles in C. nutans response to cold stress. Under heat stress treatment, 14 out of the 20 CnbHLHs exhibited decreased expression in the leaves, while 15 CnbHLHs showed reduced expression in the roots, with variations between the two tissues. Notably, CnbHLH126 was initially repressed under heat stress but was upregulated over time in the root. Under salt stress treatment, 12 out of the 20 CnbHLHs showed decreased expression in the leaves, with only CnbHLH78 gradually upregulated. Interestingly, in the roots, only five CnbHLHs were downregulated post salt stress treatment, while most genes were upregulated at different time points. Under drought stress, 14 out of the 20 CnbHLHs showed decreased expression in the leaves, while 14 CnbHLHs were upregulated in the roots. Notably, the expression of CnbHLH11, CnbHLH17, CnbHLH30, CnbHLH110, CnbHLH140, CnbHLH148, CnbHLH130, CnbHLH61, CnbHLH104, and CnbHLH173 increased at all time points post-treatment, with CnbHLH11, CnbHLH140, and CnbHLH104 showing sustained upregulation throughout the treatment. This analysis shows that CnbHLHs in C. nutans exhibit tissue-specific and dynamic responses to various abiotic stresses, suggesting their role in regulating stress adaptation mechanisms.
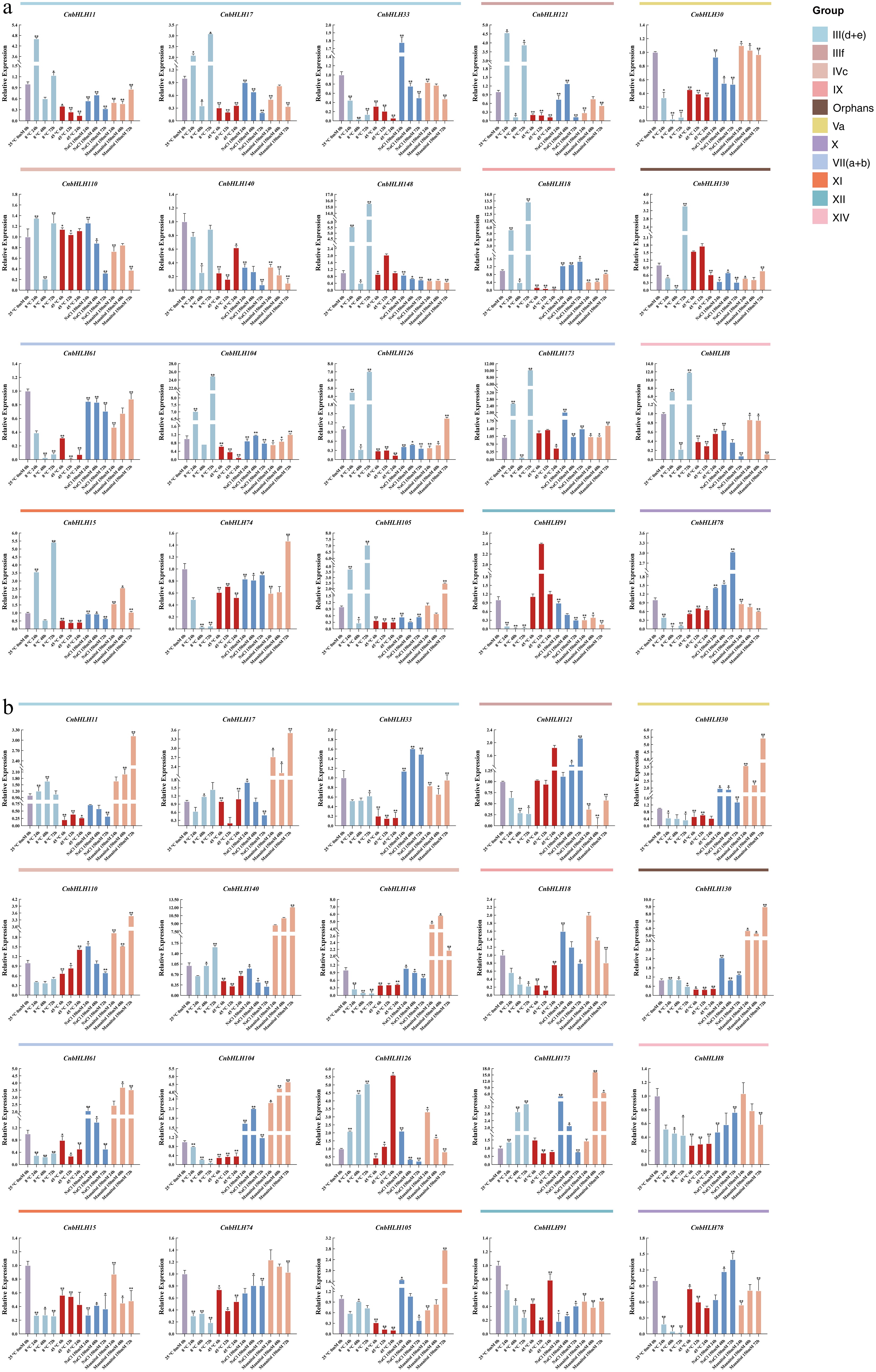
Figure 7.
qRT-PCR analysis of 20 CnbHLHs under various stress conditions, including cold (4 °C), heat (45 °C), NaCl (150 mM), and drought (mannitol 150 mM). All experiments were conducted independently three times. Error bars denote the standard deviation of triplicate samples. Asterisks denote statistically significant differences in transcript levels when compared with the blank control (25 °C, 0 h, 0 mM). (* p < 0.05, ** p < 0.01, *** p < 0.001). (a) Relative expression levels of selected CnbHLHs in leaf tissues. (b) Relative expression levels of selected CnbHLHs in root tissues.
A diagram was made to show the expression patterns of CnbHLHs in different tissues and after MeJA or stress treatments (Fig. 8). The expression patterns of CnbHLHs was different between tissues, which may mean they help control various growth and development processes. Hormones like MeJA help control the accumulation of secondary metabolites in plants[6]. The present analysis showed big differences in how CnbHLHs in the leaves and roots reacted to MeJA treatment, with only a few showing similar patterns in both tissues. This suggests that CnbHLHs may regulate metabolic pathways in different tissues. Stress conditions cause production and accumulation of secondary metabolites in plants[55]. qRT-PCR was used to check how 20 representative CnbHLHs were expressed under different stress conditions. The results showed that, for the same stress, the same genes were expressed differently in the roots and leaves, meaning stress may affect the metabolism of the above- and below-ground parts of the plant differently. The medicinal properties of different parts of C. nutans are related to differences in the accumulation of active compounds[56]. These results show that different CnbHLHs may regulate different metabolic processes in the above- and below-ground tissues. The results provide valuable insights for further functional studies on CnbHLHs and suggest potential gene targets for molecular breeding to enhance the accumulation of active compounds in C. nutans.
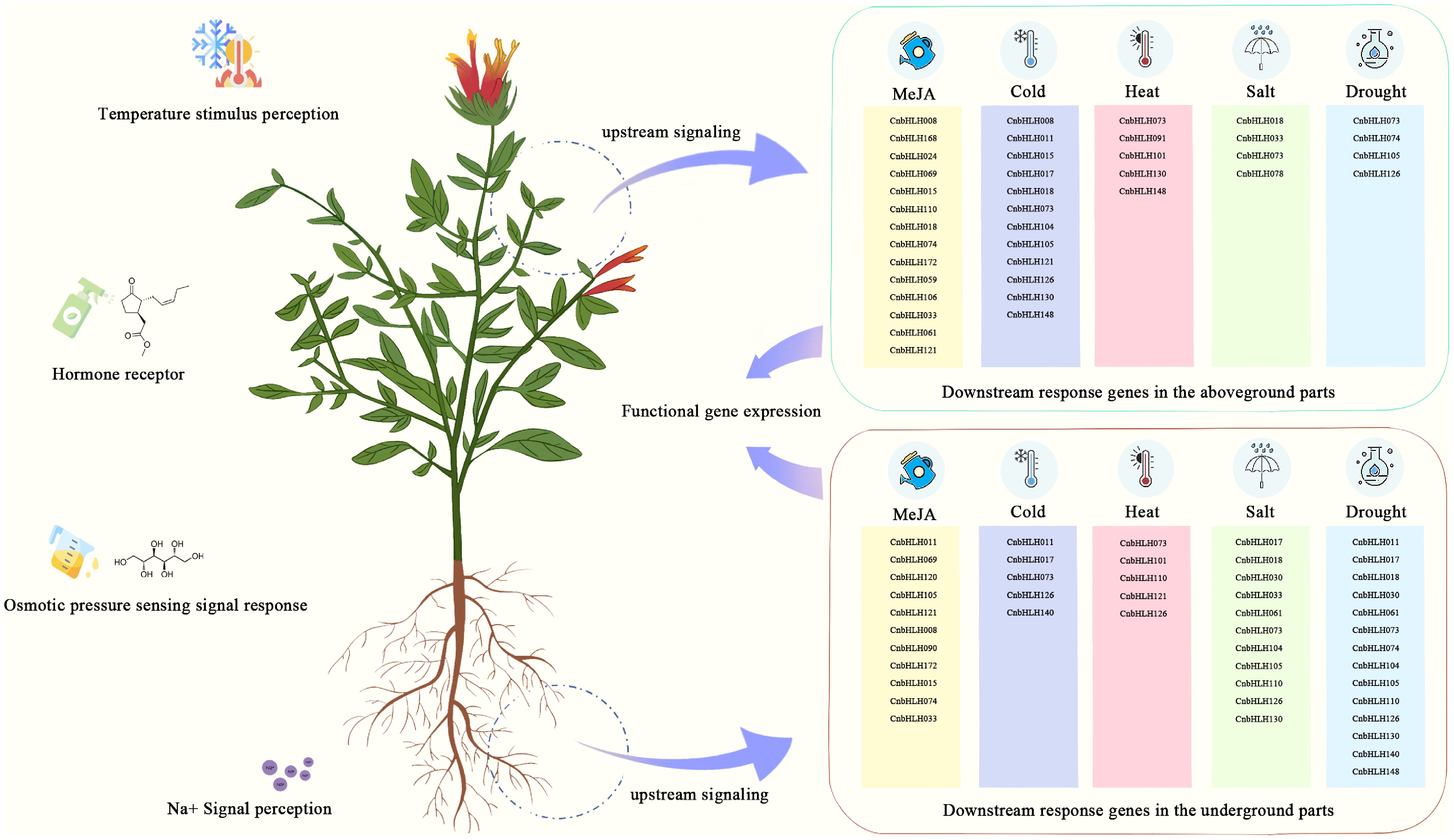
Figure 8.
Diagram showing the response pathways to different abiotic stresses mediated by various bHLHs in C. nutans.
-
C. nutans has gained attention for its potential medicinal compounds, such as flavonoids, polyphenols, and other secondary metabolites[57]. The accumulation of these metabolites vary in different tissues and are influenced by environmental factors and hormones like MeJA[6]. bHLHs regulate key processes, including plant growth and development, stress and hormone responses, and secondary metabolism[58]. However, the role of bHLHs in C. nutans remains unexplored. In this study, 182 bHLH genes in C. nutans were identified and named CnbHLH001–CnbHLH182 based on their chromosomal positions. These genes were unevenly distributed across the 18 chromosomes of C. nutans, with noticeable clustering at the chromosome ends (Supplementary Fig. S1). The number of CnbHLHs (n = 182) was similar to that in rice (n = 183)[22], but higher than in A. thaliana (n = 162)[59], Prunus mume (n = 100)[60], and Passiflora edulis (n = 138)[61]. The differences in bHLH gene numbers across species may be linked to genome size or gene duplication events[62]. The phylogenetic tree of C. nutans and Arabidopsis bHLH proteins further classified the 182 CnbHLHs into 26 subfamilies, with 10 additional CnbHLHs classified as orphans (Fig. 2). Conserved motifs and exon/intron analyses showed structural diversity among CnbHLHs from different subfamilies, while members within the same subfamily were more similar (Supplementary Fig. S2), confirming the accuracy of subfamily classification. Motif 1 and motif 2 were conserved in almost all CnbHLHs and were major components of the bHLH conserved domain, indicating their key role in the functional specificity of these TFs. Other motifs showed varying distributions across CnbHLHs, but many proteins in the same subfamily shared similar conserved motifs. For example, motif 7 and motif 8 were found only in subfamily III, and motif 10 was specific to group VIIIb. The number of exons in the CnbHLHs ranged from 1 to 22, reflecting the structural complexity of the bHLH family. Similarly, genes within the same subfamily had similar exon/intron patterns, such as subfamily Ia members having 2–3 exons, and subfamily III(a + c) members having 4–5 exons. Notably, some CnbHLHs were intron-less, mainly in subfamilies III(d + e) and VIIIb, including CnbHLH059, CnbHLH017, and CnbHLH100 (Supplementary Fig. S2, Supplementary Table S5). This feature might give these genes an advantage in rapidly responding to biological or environmental signals[63]. Transcriptome data showed that these genes were rapidly induced in response to MeJA treatment in leaves, with CnbHLH017 also showing a rapid response in roots (Fig. 7). Studies in Arabidopsis and maize have shown that intron-less genes exhibit rapid expression responses to light and temperature stimuli[64], suggesting that such CnbHLHs might help C. nutans's adapt quickly to changing environments. The 3D structural predictions reveal varied features across subfamilies (Fig. 5b), providing a basis for the functional diversity of family members while maintaining core roles. Analysis of physicochemical properties also showed significant differences among CnbHLHs (Fig. 1, Supplementary Table S3), suggesting that these proteins may have distinct roles in their microenvironments[65].
Gene duplication is prevalent in plant genomes and contributes significantly to evolution and adaptation in response to environmental changes[62]. In C. nutans, 97 gene duplication pairs were found(Fig. 4a, Supplementary Table S7), which include five tandem duplicates and 92 segmental duplicates. These results suggested that CnbHLHs expansion mainly occurred through segmental duplication, similar to other species[66]. Some duplicated genes with similar structures and expression patterns may have functional redundancy. For instance, the segmental duplicated gene pair CnbHLH061 and CnbHLH106, which were clustered in subfamily VII(a + b), both have motif 1, motif 2, and six exons. Tissue-specific expression analysis showed that both genes were mainly expressed in mature leaves. Their expression in leaves both increased initially and then decreased after MeJA treatment (Fig. 6). However, many duplicated gene pairs showed different expression patterns. For example, CnbHLH026 and CnbHLH147, which are in subfamily XIII, are segmentally duplicated genes. CnbHLH026 was specifically highly expressed in mature leaves and showed a transient increase in expression in both leaves and roots following short-term MeJA induction. In contrast, CnbHLH147 was specifically highly expressed in roots, but its expression in MeJA-treated leaves and roots was relatively low (Fig. 6). In addition, the segmentally duplicated gene pair CnbHLH140 and CnbHLH005, both from subfamily IVc, were preferentially expressed in leaves before MeJA treatment (0 h), and at the final time point (72 h), respectively. These gene duplication events in C. nutans may have provided additional genetic material with diversified functions for adapting to changing environments. In addition, a bHLH gene cluster consisting of the tandem duplicated gene members including CnbHLH044, CnbHLH045, CnbHLH046, and CnbHLH047 was identified, with all genes belonging to the same subfamily Ib(2). All of these genes exhibited higher expression in the roots compared with the leaves and responded to MeJA treatment. In Catharanthus roseus, three clustered CrbHLHs (named BIS1, BIS2, and BIS3) collectively regulate the biosynthesis of phenylethylamine in the terpene alkaloid biosynthesis pathway[18]. These clustered CnbHLHs might also have overlapping or synergistic regulatory functions in root development and/or MeJA response in C. nutans. Synteny analysis showed that 138 CnbHLHs had syntenic relationships with A. paniculata, followed by 129 with S. cusia, 106 with A. thaliana, and 82 with V. vinifera (Fig. 4b, Supplementary Table S8), suggesting large-scale gene expansion occurred before these species diverged. Thirty-seven CnbHLHs exhibited synteny across all five species, indicating that these genes within the bHLH family played a crucial role in evolutionary processes. The extent of synteny among Acanthaceae species (C. nutans, A. paniculata, and S. cusia) was much higher than with other species, suggesting that CnbHLHs may have expanded specifically in Acanthaceae to meet the unique evolutionary or adaptive needs of this plant family. The low Ka/Ks ratios of CnbHLHs compared with their homologs in other species suggest that these gene family members are under strong purifying selection (Fig. 4c, Supplementary Table S9), with harmful mutations being eliminated to maintain functional stability. This high level of functional conservation shows the importance of these genes in environmental adaptation and maintaining physiological stability across species. The CnbHLHs interaction network, consisting of 408 interactions and 59 nodes (Fig. 5a), showed that interactions occurred among different CnbHLHs. Notably, the CnbHLH168 node had the highest interaction reliability with several CnbHLHs, such as CnbHLH059, CnbHLH175, CnbHLH139, and CnbHLH121. The gene expression patterns of these interacting proteins were similar to CnbHLH168. CnbHLH168 was mainly expressed in leaves and stems, while its interacting partners, CnbHLH059 and CnbHLH175, were mainly expressed in young leaves (Fig. 6a, block b), and CnbHLH139 and CnbHLH121 were mainly expressed in mature leaves (Fig. 6a, block c). Additionally, CnbHLH168 and these highly reliable interacting CnbHLHs were all preferentially expressed in the leaves after MeJA treatment (Fig. 6b, block I), suggesting these gene pairs may jointly regulate leaf development or the response to MeJA. However, many other interacting CnbHLH pairs with low interaction reliability showed different expression patterns. For example, CnbHLH038 was highly expressed in the roots at both 0 and 72 h after MeJA treatment. The interacting genes showed different expression patterns: CnbHLH058 was highly expressed in young leaves, while CnbHLH153 had the highest expression in the stem. Whether these gene pairs interact to regulate certain biological processes together needs further validation.
Increasing evidence shows that bHLHs are crucial in plant growth, development, stress and hormone responses[58]. Transcriptomic analysis showed that most CnbHLHs had tissue-specific expression patterns (Fig. 6a), suggesting their broad involvement in growth and developmental processes of C. nutans. MeJA plays a crucial role in regulating plant secondary metabolism, with many TFs, including bHLH and MYB, involved in this process[53]. The present results showed that after filtering out low-expressed genes, 39 CnbHLHs were identified in the leaves (Fig. 6b), and 44 in the roots (Fig. 6c) in response to MeJA treatment. Only 19 genes responded to MeJA in both tissues, indicating notable differences in the CnbHLHs responding to MeJA in the root and leaves. In both the roots and leaves, CnbHLHs expression in response to MeJA showed varied patterns during treatment, with different CnbHLHs involved at various time points after MeJA treatment. This suggests that CnbHLHs help fine-tuning the plant's response to MeJA, with different members active at various stages of signal transduction or cellular processes. In many medicinal plants, the accumulation of bioactive compounds such as flavonoids and terpenoids is a result of the plant's response to environmental stresses[67]. qRT-PCR was employed to analyze the expression patterns of 20 representative CnbHLH genes under different stress conditions, including cold, heat, salt, and drought (Fig. 7). The results showed that the expression patterns of the same gene in response to the same stress were markedly different between the roots and leaves, suggesting that different stress conditions may affect the metabolism of the aerial and underground parts differently. Specifically, most CnbHLHs in the leaves showed an initial increase in expression followed by a decrease, and then another increase during cold stress treatment. This phenomenon suggests that the leaves may initially adjust the expression of these genes to cope with low temperatures, and then gradually recover or enhance expression to adapt to prolonged stress. For example, CnbHLH148 showed a more than 15-fold upregulation in the leaves after 72 h of cold stress treatment. Its homolog, AtILR3, has been reported to improve the activities of superoxide dismutase, peroxidase, and catalase, thereby enhancing stress tolerance[68]. In contrast, more than half of the CnbHLHs in the roots showed reduced expression under cold stress, suggesting that the roots may use a different mechanism to cope with cold stress. Moreover, most CnbHLHs in the roots showed increased expression after salt and drought stress. Some CnbHLHs were highly expressed in both salt and drought stress, such as CnbHLH30, which showed increased expression at various time points under both treatments in the roots. The homolog of CnbHLH30, AtbHLH102 (BIM2), works with TFs like HDA15 to regulate anthocyanin accumulation and growth responses under salt stress[69]. However, most CnbHLHs were more sensitive to one of these stresses. For instance, CnbHLH011 and CnbHLH017 showed consistently increased expression at all time points during drought stress in the roots. Their homolog, AtMYC2 (AtbHLH006), expressed in the roots, has been identified as a transcriptional activator induced by ABA during drought stress[70]. CnbHLH121 showed upregulated expression at various time points after salt stress in the roots. Its homologs, AtbHLH002 and AtbHLH042 from subfamily IIIf interacts with MYBs to regulate the biosynthesis of flavonoids and anthocyanins, enhancing stress tolerance[71]. Additionally, CnbHLH105 showed significantly increased expression after prolonged salt stress in the roots. Its homolog, AtbHLH059, is induced by salt stress and contributes to physiological processes like Na+/K+ balance, redox homeostasis, and transpiration, thus improving salt tolerance[72]. Conversely, in the leaves, most CnbHLHs were reduced by salt and drought stress, which may indicate different adaptation strategies of the root and leaf tissues to these stresses. Only a few CnbHLH genes exhibited similar expression patterns in both roots and leaves under stress conditions. For example, CnbHLH078 was induced to express higher levels after prolonged salt stress in both the roots and leaves. Its homolog, AtbHLH112, from subfamily X, binds to E-box and GCG-box elements in target gene promoters to regulate stress-responsive genes, enhancing stress resistance. Overexpression of AtbHLH112 increases tolerance, while its functional loss renders plants more susceptible to stress[73]. Collectively, these findings highlight the tissue-specific and dynamic expression patterns of CnbHLHs under various stress conditions. CnbHLHs might function similarly to their homologs by regulating key processes such as antioxidant enzyme activity and secondary metabolism in response to environmental stressors.
-
In this study, 182 bHLHs were identified and analyzed in C. nutans, revealing their potential roles in regulating important processes such as growth, development, and responses to hormones and stress. The expansion of CnbHLHs mainly happened through segmental duplication, with many genes showing functional redundancy, while others exhibited different expression patterns, indicating functional diversification. A group of tandemly duplicated genes, including CnbHLH044–CnbHLH047, which shared similar expression patterns, was identified, suggesting their possible cooperative functions in root development and MeJA responses. Synteny analysis offered insight into the evolutionary history of CnbHLHs, revealing higher synteny within Acanthaceae species, suggesting species–specific expansions. Tissue-specific expression patterns and varied responses to MeJA and environmental stress further emphasized the dynamic roles of CnbHLHs in regulating metabolic processes in different tissues. These findings provide new insights into the functional roles of CnbHLHs, offering a valuable resource for advancing molecular breeding and enhancing stress resilience in medicinal plants. However, this study has certain limitations. The current analysis was conducted based on a limited sample scope, and key regulatory mechanisms inferred from in silico predictions lack direct experimental validation. In addition, the integration of multi-omics data remains relatively shallow, which may limit the depth of functional interpretation. Future work should aim to perform molecular assays to verify predicted interactions, and incorporate epigenetic or metabolomic data. These efforts will help to achieve a more precise and holistic understanding of the roles of bHLHs in C. nutans.
We would like to express our sincere gratitude to the reviewers for their valuable comments and suggestions, which greatly improved the quality of our manuscript. This work was supported by the Fund of Fujian Key Laboratory of Island Monitoring and Ecological Development (Island Research Center, MNR) (2022ZD06), the Science and Technology Innovation Project of Fujian Agriculture and Forestry University (CXZX2019144G, CXZX2020086A), the National Natural Science Foundation of China (31970333 and 32100168), the Basic Discipline Research Enhancement Program of Fujian University of Traditional Chinese Medicine (XJC2023008), Construction Funds for Cross-disciplinary Synthetic Biology at the Institute of Future Technology, Fujian Agriculture and Forestry University (133-712023010), and the Scientific Research Start-up Fund of Fujian Agriculture and Forestry University (Yuan Qin).
-
The authors confirm their contributions to the paper as follows: resources, data curation, formal analysis, writing—original draft, writing—review and editing: An C; software: Wu D; software, methodology: Li N; conceptualization, supervision: Yao Y; software: Lu L; supervision: Zhang Z; supervision: Cheng Y; supervision, project administration: Zheng P; Funding acquisition, conceptualization: Qin Y. All authors reviewed the results and approved the final version of the manuscript.
-
The raw transcriptome sequencing data used in this study has been deposited in CNGBdb under the accession number CNP0006103. The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 List of CDS and protein sequences of 182 CnbHLHs
- Supplementary Table S2 Specific primers for 20 CnbHLHs for qRT-PCR.
- Supplementary Table S3 Chromosomal Locations and Physicochemical Properties of CnbHLHs.
- Supplementary Table S4 The Nucleotide Proportions of Conserved Sites in the Domains of CnbHLHs.
- Supplementary Table S5 Motif Distribution and Gene Structure of CnbHLHs.
- Supplementary Table S6 Identification of Cis-Regulatory Elements in CnbHLHs.
- Supplementary Table S7 Gene Duplication of CnbHLHs.
- Supplementary Table S8 Collinearity Relationships of bHLHs in Clinacanthus nutans and Related Species.
- Supplementary Table S9 Ka/Ks Analysis Results of CnbHLHs Across Different Species.
- Supplementary Table S10 Predicted Interaction Scores of the CnbHLHs Protein Interaction Network.
- Supplementary Table S11 Predicted Secondary Structure and Related Parameters of CnbHLHs.
- Supplementary Table S12 Predicted Three-Dimensional Structure and Related Parameters of CnbHLHs.
- Supplementary Table S13 Expression matrix of PebHLHs in different tissues and organs at corresponding stages.
- Supplementary Table S14 Relative Expression Analysis of Target Gene (CnbHLH) Based on One Housekeeping Genes (Abiotic stress Leaf).
- Supplementary Fig. S1 The chromosomal location of 182 CnbHLHs.
- Supplementary Fig. S2 Analysis of motif pattern and gene structure of the CnbHLHs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
An C, Wu D, Li N, Yao Y, Lu L, et al. 2025. The basic helix-loop-helix (bHLH) gene family in Clinacanthus nutans: insights into tissue-specific expression, stress responses, and methyl jasmonate (MeJA) responsiveness. Medicinal Plant Biology 4: e032 doi: 10.48130/mpb-0025-0030 

The basic helix-loop-helix (bHLH) gene family in Clinacanthus nutans: insights into tissue-specific expression, stress responses, and methyl jasmonate (MeJA) responsiveness
- Received: 23 April 2025
- Revised: 08 August 2025
- Accepted: 25 August 2025
- Published online: 29 September 2025
Abstract: Clinacanthus nutans is valued for its medicinal properties, largely attributed to bioactive compounds such as flavonoids, polyphenols, and other secondary metabolites. These compounds accumulate differentially across tissues and are influenced by environmental cues and phytohormones like methyl jasmonate (MeJA). Transcription factors play pivotal roles in mediating these responses, with the basic helix-loop-helix (bHLH) gene family emerging as key regulators of hormone- and stress-responsive gene expression involved in secondary metabolism. In this study, 182 bHLHs were identified in C. nutans, labeled CnbHLH001 to CnbHLH182 according to their chromosomal locations. Phylogenetic analysis grouped them into 26 subfamilies, with 10 classified as orphans. Structural characterization revealed subfamily-specific motif architectures and exon–intron structures. Gene duplication analysis indicated that segmental duplication was a major driver of CnbHLH family expansion, and expression analysis suggested both conserved and divergent expression among duplicated gene pairs. Tandemly duplicated genes such as CnbHLH044 to CnbHLH047 showed similar expression patterns. Synteny analysis showed that CnbHLHs in Acanthaceae species are more similar to each other than to genes in other species, suggesting a possible Acanthaceae-specific expansion. Furthermore, this study provides the first genome-wide analysis of CnbHLHs expression in response to abiotic stresses. Many CnbHLHs showed tissue-specific expression and responded differentially to MeJA and stress treatments. These findings provide a foundation for future research to explore the regulatory mechanisms of CnbHLHs and their potential to improve the medicinal properties of C. nutans.
-
Key words:
- Clinacanthus nutans /
- Basic helix-loop-helix /
- Genome evolution /
- Expression patterns /
- Abiotic stress