









-
Flower color is one of the most important characteristics of ornamental plants, and is primarily attributed to the presence of three pigments: flavonoids, carotenoids and betalains. Flavonoids are responsible for a wide range of colors, carotenoids are mainly responsible for yellow to orange, whereas betalains seem to be confined to caryophyllales though they produce yellow to orange and red to violet coloration[1−3]. By comparison, more than
9000 flavonoids have been found in vegetables, fruits, flowers, leaves and other plant tissues[4, 5]. The flavonoids bear a common diphenylpropane skeleton (C6-C3-C6) in which two benzene rings are linked via a three-carbon chain[6]. According to the different oxygenation patterns of the heterocyclic C ring, flavonoids can be further classified into flavones, flavanones, flavonols, isoflavones and anthocyanins[6, 7]. Among them, anthocyanins are one of the most abundant and widely distributed flavonoids in land plants, conferring red, blue and purple pigmentations to flowers, seeds and fruits[8, 9]. In addition, anthocyanins selectively attract pollinators and protect plants against biotic or abiotic stresses such as high UV intensity, low temperature, and pathogen infection[10−12].Although there are thousands of flavonoids with great structural and functional diversity, they share relatively conservative biosynthetic pathways with a C6-C3-C6 backbone from the pioneering land plants to higher vascular plants (Fig. 1)[6, 9, 13]. Generally, the core flavonoid biosynthetic pathway begins with the condensation of a molecule of p-coumarolyl-CoA and three molecules of malonyl-CoA by chalcone synthase (CHS) to generate naringenin chalcone (NC)[14]. NC is, to some extent, spontaneously isomerized into naringenin, which can also be rapidly promoted by chalcone isomerase (CHI)[15]. Subsequently, flavanone 3-hydroxylase (F3H) catalyzes naringenin to produce dihydrokaempferol which can be further hydroxylated by flavonoid 3´-hydroxylase (F3´H) or flavonoid 3´5´-hydroxylase (F3´5´H) on ring B to form dihydroquercetin and dihydromyricetin, respectively[16−18]. Alternatively, naringenin can also be catalyzed by F3´H or F3´5´H to produce eriodictyol or pentahydroxy flavone, respectively. Dihydroquercetin and dihydromyricetin can also be formed from hydroxylation of eriodictyol and pentahydroxy flavone by F3H[16, 17]. The dihydroflavonols (dihydrokaempferol, dihydroquercetin and dihydromyricetin) are further catalyzed by dihydroflavonol reductase (DFR) and anthocyanin synthase (ANS) to produce leucoanthocyanidins and anthocyanins successively[19, 20]. Specific flavonoid flux can be branched from the core pathway. For example, naringenin can also be exploited by isoflavone synthase (IFS), flavone synthase (FNS) or flavanone 2-hydroxylase (F2H) to produce isoflavones or other flavones such as apigenin or luteolin[21−23]. Flavonols can be formed from catalyzation of dihydroflavonols by specific flavonol synthase (FLS)[24]. Proanthocyanidins can be branched from leucoanthocyanidin by leucoanthocyanidin reductase (LAR) or from anthocyanidin by anthocyanidin reductase (ANR), while anthocyanidins can also be further glycosylated by flavonoid 3-O-glucosyltransferase (3GT) to become stable anthocyanins[13, 25−27]. Therefore, naringenin is a general precursor for flavonols, anthocyanins, proanthocyanidins, flavones, and isoflavones.
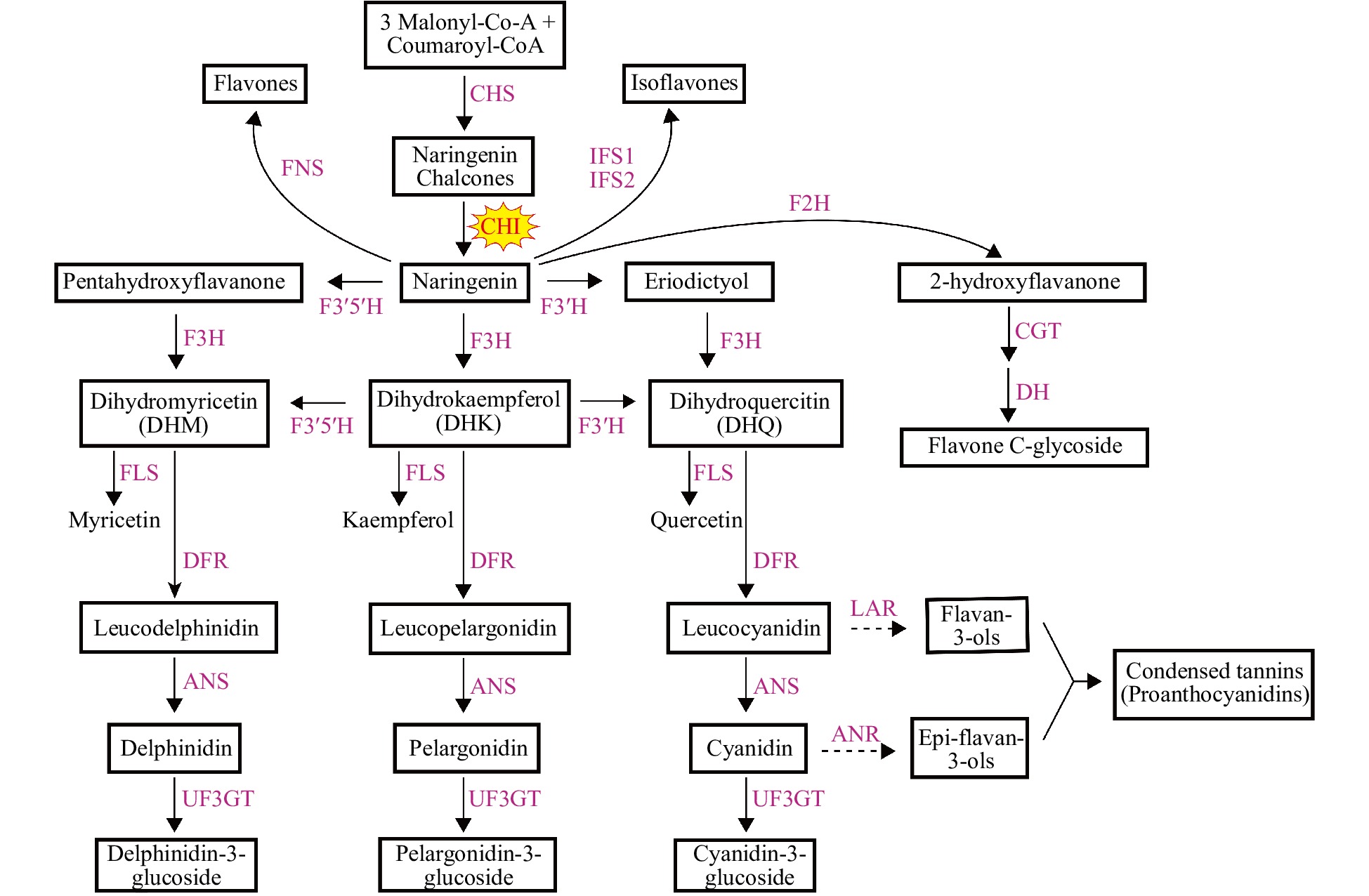
Figure 1.
Flavonoid biosynthetic pathway. The two dotted lines represent the flavan-3,4-diols (including leucodelphinidin, leucopelargonidin and leucocyanidin) which are catalyzed by LAR to generate flavan-3-ols and the 3-OH-anthocyanidins (including delphinidin, pelargonidin and cyanidin) and also catalyzed by ANR to generate epi-flavan-3-ols, respectively. Enzymes are indicated as capitals in red. The names of the enzymes are abbreviated as follows: CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone 3-hydroxylase; FLS, flavonol synthase; F3'H, flavonoid 3'-hydroxylase; DFR, dihydroflavonol 4-reductase; ANS, anthocyanidin synthase; LAR, leucoanthocyanidin reductase; ANR, anthocyanidin reductase; FNS, flavone synthase; F3'5'H, flavonoid 3',5'-hydroxylase; IFS, isoflavone synthase; F2H, flavanone 2-hydroxylase; UF3GT, UDP glucose: flavonoid 3-O-glucosyltransferase; CGT, C-glucosyl transferase; DH, dehydratase.
Chalcone isomerase is the key branch point enzyme in the early flavonoid metabolic pathway, controlling catalytic efficiency to produce flavonoid intermediate substrates which are the skeletons of downstream metabolites. Based on previous studies, the CHI superfamily comprises four types of CHI-fold proteins. Type I proteins are ubiquitous in vascular plants and functional in flavonoid biosynthesis, whereas type II proteins are specifically involved in isoflavonoid production in legumes[28]. Type III proteins tend to be fatty acid-binding proteins (FAPs) localized in plastids[29], whereas type IV CHI proteins lack conventional activities and might act as an enhancer of flavonoid biosynthesis[15, 30−32]. Therefore, the classical type I proteins and legume specific type II CHIs should be the real chalcone isomerases responsible for efficient naringenin production.
As one of the largest monocotyledonous families, Amaryllidaceae includes many economically important plants with ornamental, nutritional, and medicinal values and has been the focus of classification or phylogeny and cultivation studies for centuries[33]. Clivia miniata, a perennial herbaceous ornamental plant in the family Amaryllidaceae, is native to South Africa and best-known for its ornamental value. However, compared with other ornamental flowers, its monotonous flower color has significantly affected its ornamental and economic value[34−36]. The traditional line selection and cross breeding method has not generated novel flower colors. Alternatively, genetic engineering technology might be a potential way to modify the flower colors of C. miniata. The prerequisite for successfully engineering desirable flower colors is to isolate and identify functional anthocyanin biosynthetic genes and decode the underlying mechanism restraining the flower colors of C. miniata. Recently, there have been a few studies regarding flavonoid biosynthesis in this genus[35, 36]. However, the small amount of available information is more than enough to understand the mechanisms. In this study, three CmCHI genes encoding two type I and one type IV proteins were isolated and functionally characterized from C. miniata, which provides new insights into flavonoid biosynthesis in C. miniata.
-
An in situ TBLASTN search with AtCHI as bait probe against the transcriptome database of Clivia miniata and manual NCBI-BLASTX search were performed. Three unigenes encoding orthologs of AtCHI were mined and predicted as potential CHI genes in C. miniata, thus tentatively designated as CmCHI1, CmCHI2 and CmCHI3, respectively. Specific primers were designed and employed to clone the complete ORFs and results confirmed that CmCHI1, CmCHI2 and CmCHI3 have 714 bp, 657 bp, and 633 bp ORFs and encoded 237, 218, and 210 amino acids, respectively (Supplementary Table 1). Amino acid sequence analysis of CmCHI1 revealed a 59% similarity to AtCHI1 (Arabidopsis thaliana), and a highest value of 87% similarity to CHI1 from Lycoris radiata, respectively. The same comparison for CmCHI2 is 51% against AtCHI and a much lower index of 59% against CHI from Canarium album, respectively. Additionally, CmCHI3 yielded the highest similarities, with 83% against AtCHI3 and 88% against CHI2 from Narcissus tazetta subsp. chinensis, respectively.
To further study the homology of amino acid sequences, sequence alignment revealed that CmCHI1 shared all the conserved active sites, which were either in groups or singlet at the following eight sites (CmCHI1 numbering): R(42) G(43)L(44), F(53)T(54), I (56), L (107), Y (112), K(115)V(116), N(119) and S(196)I(197). CmCHI2, however, shared most of the eight sites except for three substitutions F(52)→E, I(56)→H and Y(112)→F. It is important to note that CmCHI2, as well as CmCHI1, shared the type I active sites SI. On the contrary, CmCHI3 did not share any of these active sites (Fig. 2a). In order to define the CmCHIs, the phylogenetic analysis of CHIs from C. miniata and other plant species was performed using the neighbor-joining method, and the CHI proteins in our analysis were sorted into four main groups (I, II, III, IV) (Fig. 2b). CmCHI1 was most closely related to the CHIs from monocotyledonous plants, belonging to the nonleguminous type I CHI family. CmCHI2, also placed into group I, was shown to be the root to the largest clade in the tree of our analysis. Our group II contained a clade including exclusively leguminous CHIs: MsCHI (Medicago sativa), PvCHI (Phaseolus vulgaris), GmCHI1A (Glycine max) and PlCHI (Pueraria lobate). CmCHI3 fell into type IV CHI family, which included CHIL from several species. Lastly, the type III CHI family included CHI from three species in our tree.
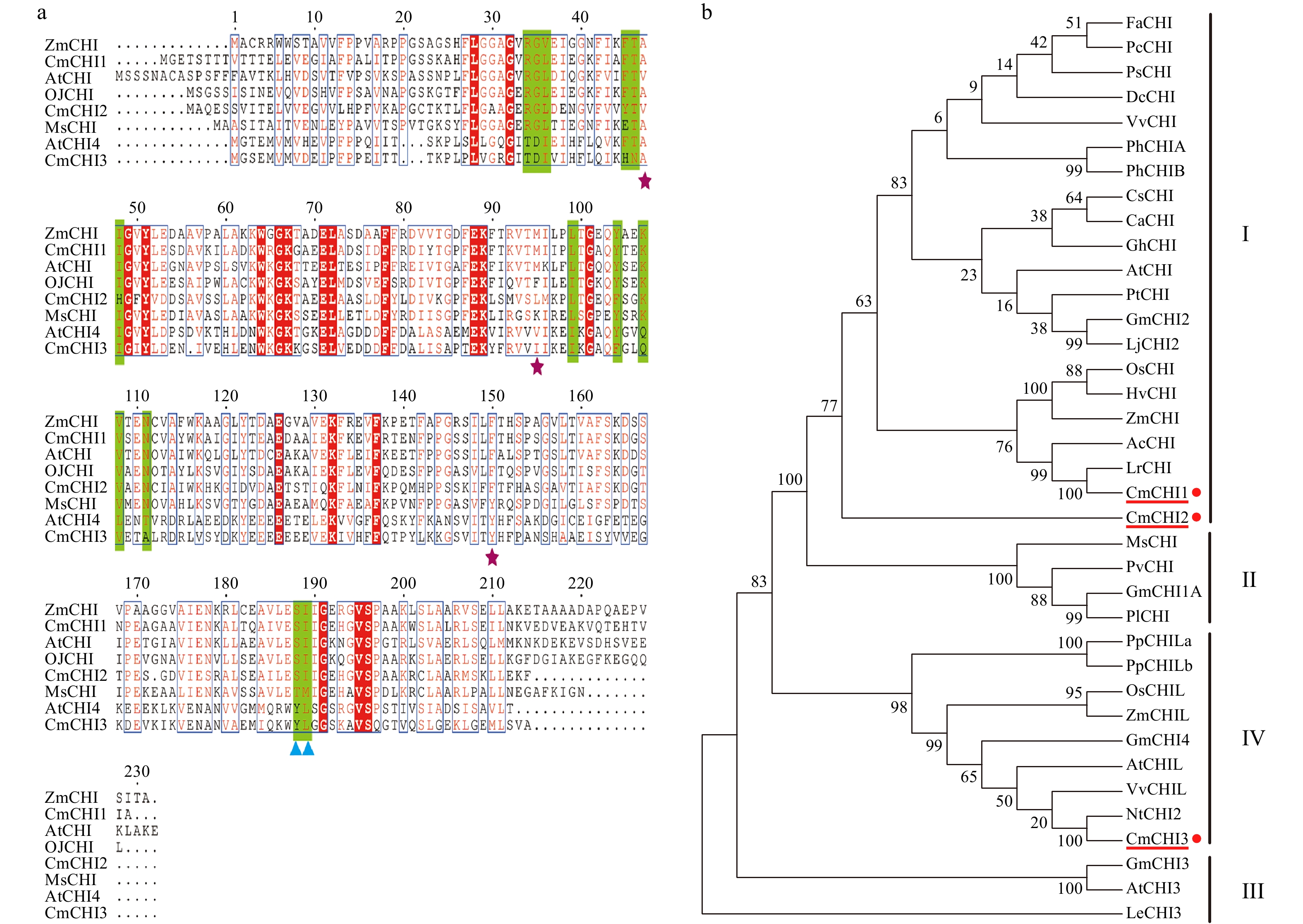
Figure 2.
Sequence and phylogenetic analysis of CmCHIs. (a) Sequence alignment of CHIs for C. miniata (Cm) and other species (Arabidopsis thaliana: At; Medicago sativa: Ms; Ophiorrhiza japonica: Oj). The green highlights indicate residues that bind (2S)-naringenin, the purple stars indicate the hydrogen bonding network residues of the active site. The blue triangles identify residues postulated to determine substrate preference for naringenin chalcone and isoliquiritigenin. (b) Phylogenetic analysis of CHI proteins from C. miniata and other plant species. Numbers indicate bootstrap values for 1,000 replicates. GenBank accession numbers are as follows: AtCHI (Arabidopsis thaliana, P41088), MsCHI (Medicago sativa, P28012), PsCHI (Paeonia suffruticosa, ADK55061), ZmCHI (Zea mays, CAA80441), VvCHI (Vitis vinifera, P51117), CsCHI (Citrus sinensis, BAA36552), GmCHI2 (Glycine max, AY595415), LjCHI2 (Lotus japonicus, Q8H0G1), GmCHI1B1 (Glycine max, AY595414), LjCHI (Lotus japonicus, AJ548840), PvCHI (Phaseolus vulgaris, P14298), GmCHI1A (Glycine max, AY595413), PlCHI (Pueraria lobata, Q43056), PpCHILa (Physcomitrella patens, XP_001773128), PpCHILb (Physcomitrella patens, XP_001769093), VvCHIL (Vitis vinifera, XP_002280158), AtCHIL (Arabidopsis thaliana, NP_568154), GmCHI4 (Glycine max, AY595417), OsCHIL (Oryza sativa, NP_001065587), ZmCHIL (Zea mays, NP_001151452), GmCHI3 (Glycine max, AY595416), PhCHIA (Petunia hybrida, AAF60296), PhCHIB (Petunia hybrida, CAA32730.1), AcCHI (Allium cepa, AY700850), ChCHI (Gossypium hirsutum, ABM64798), DcCHI (Dianthus caryophyllus, Q43754), AtCHI3 (Arabidopsis thaliana, AY084729), FaCHI (Fragaria ananassa, Q4AE11), PtCHI (Populus trichocarpa, XP_002315258), PcCHI (Pyrus communis, A5HBK6), and LeCHI3 (Lycopersicon esculentum, AY348871).
To further define the CmCHIs, we used the Arabidopsis protoplast transient transfection system to detect the subcellular localizations of the CmCHIs. Meanwhile, the GFP controlled by 35S promoter was used as a control. Consequently, CmCHI1, CmCHI2, and CmCHI3 were observed to have diffuse distributions similar to GFP, indicating their cytoplasmic localizations (Fig. 3). The aforementioned analysis indicated that CmCHI1 and CmCHI2 tended to be real CHI proteins while CmCHI3 might be their ancestral sequence without isomerase activity.
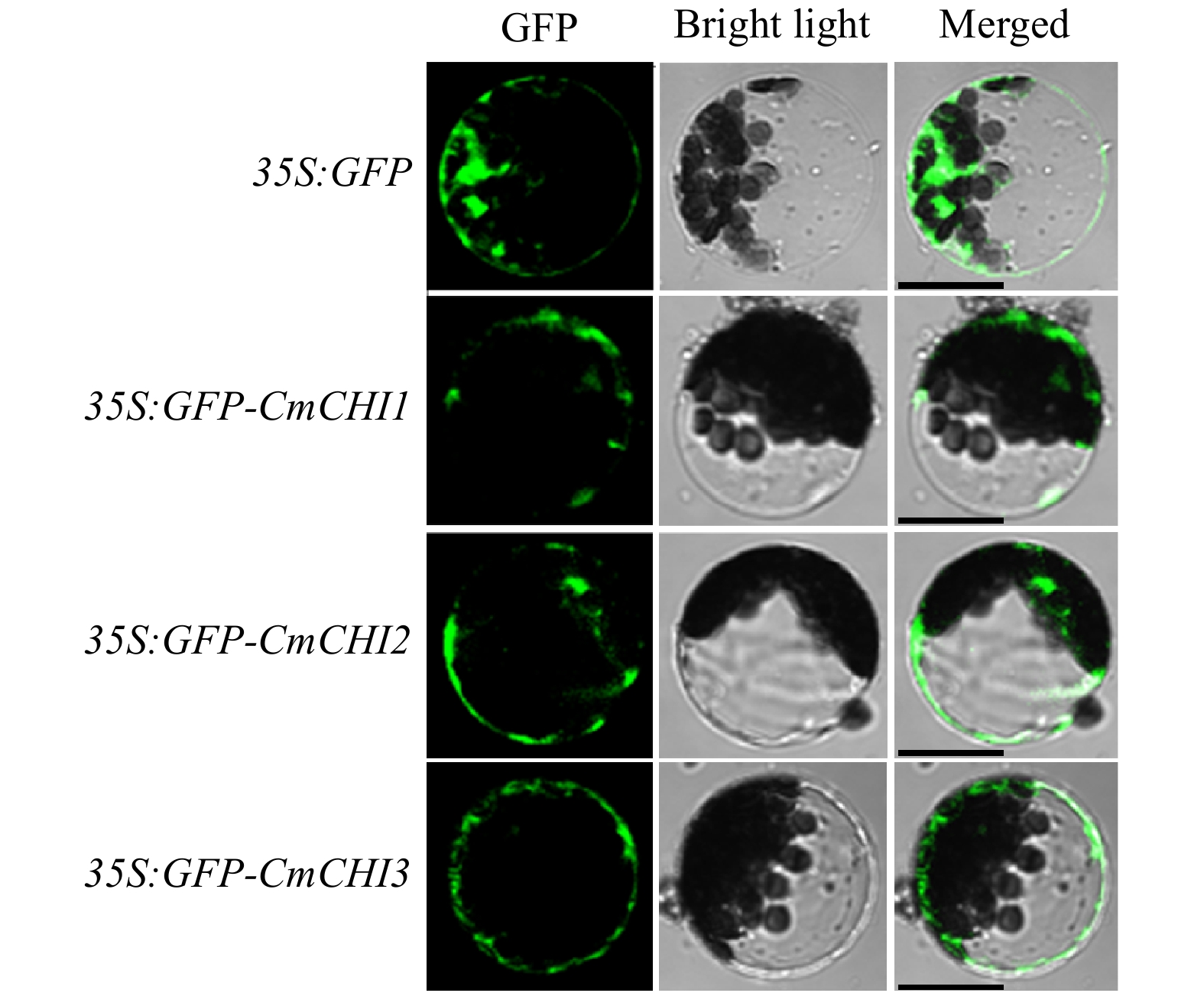
Figure 3.
Intracellular localizations of CmCHI1, CmCHI2 and CmCHI3 in Arabidopsis protoplasts. Different constructs were transiently transfected into wild-type Arabidopsis protoplasts and incubated for 21 h at room temperature in the dark. Laser scanning confocal microscopy captured the fluorescent proteins. Bars: 25 µm.
Anthocyanin accumulation and CmCHIs expression in C. miniata
-
To verify the relationship between CmCHIs transcripts and anthocyanin accumulation, both CmCHIs expression levels and the corresponding anthocyanins in flowers, leaves and fruit peels were measured. The results showed that anthocyanins accumulated less in green or white buds, peaked at the early opening period, and dropped when flowers were fully opened. Anthocyanins also mainly accumulated in the red fruit peels and red leaf bases (Fig. 4). qRT-qPCR analysis indicated that the expression of CmCHI1 had an overall upward trend, whereas CmCHI2 and CmCHI3 had an opposite trend with highest transcripts in green buds and reducing during flower development (Fig. 4). Considering their expression patterns in leaf bases and fruit peels, all the CmCHIs showed higher expression levels in red leaf bases and red peels compared to green tissues, consistent with anthocyanin profile data. Overall, CmCHI1 had an overwhelming expression level in all colored tissues detected when compared with CmCHI2 and CmCHI3, whereas expression of CmCHI2 was the lowest among all the three CmCHIs. The previous results indicated that CmCHI1 may encode the most pivotal chalcone isomerase in C. miniata.
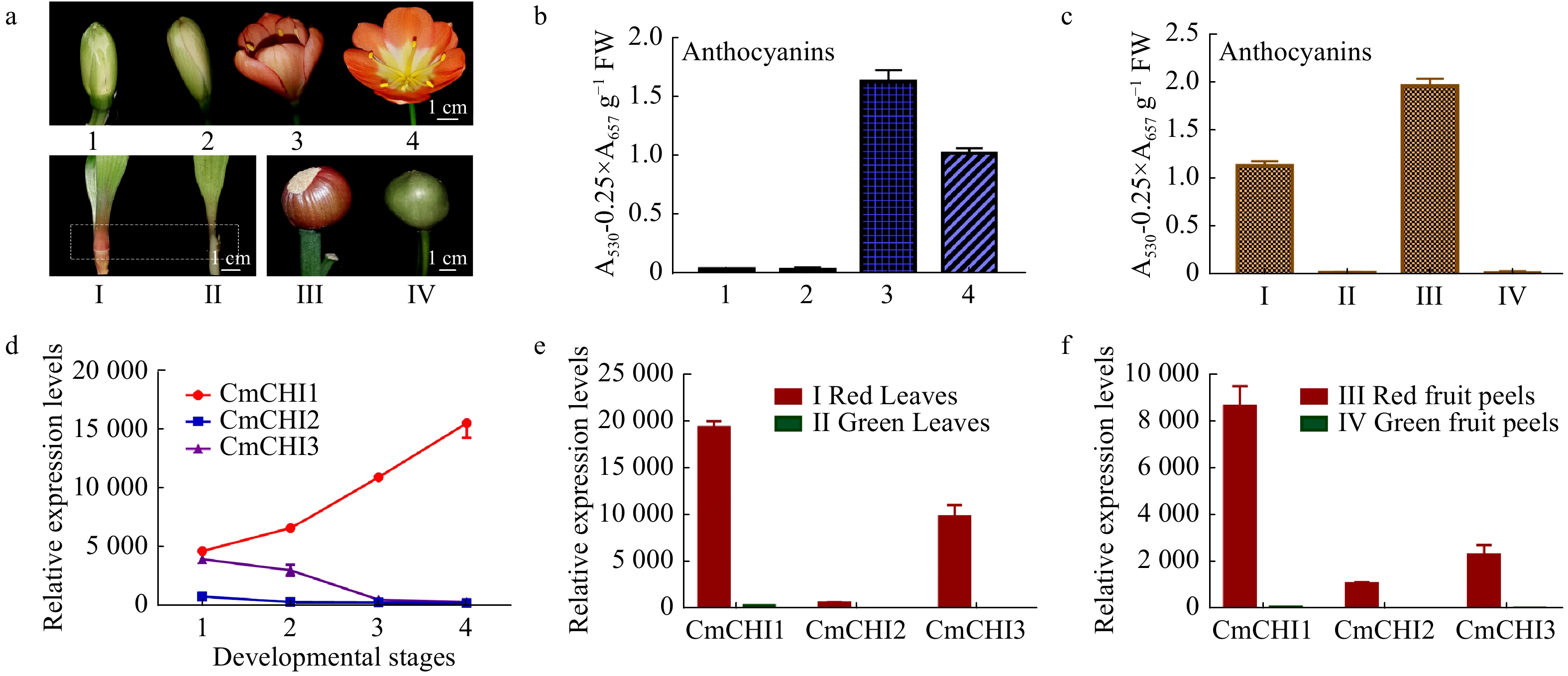
Figure 4.
The total anthocyanin content and CmCHI expression levels in different colored tissues of C. miniata. (a) The materials of C. miniata used in total anthocyanin analysis. 1−4 represent flowers at different developmental stages (flowers with green buds, flowers with white buds, fully pigmented flowers before complete blooming, fully opened flowers). I−IV represent different tissues of C. miniata. I and II represent young seedlings with differently colored leaf bases. The rectangle with dotted lines indicates the sample position. III and IV indicate fruit peels with different color. (b) The total anthocyanins detected in C. miniata flowers at different developmental stages. (c) The total anthocyanins detected in different colored leaf bases and fruit peels of C. miniata. (d) Transcripts of CmCHIs in C. miniata flowers at different developmental stages. 1−4 represent flowers at different developmental stages. (e) The relative expression levels of CmCHIs in different colored leaf bases of C. miniata. (f) The relative expression levels of CmCHIs in different pigmented fruit peels of C. miniata. Data represented means ± S.E. of three biological replicates.
Enzymatic activity of recombinant CmCHIs
-
To validate the isomerase activities of the CmCHIs, recombinant proteins were prepared and purified from Escherichia coli BL21 (Fig. 5a). As described in the introduction, the isomerization of naringenin chalcone to naringenin is somewhat spontaneous and CHI accelerates the isomerization (Fig. 5b). Using naringin chalcone as the substrate, the catalytic activity of CmCHIs was measured. As shown in Fig. 5c, CmCHI1 and CmCHI2 could completely convert all substrate NC into naringin after only 5 min reaction time, indicating their high catalytic properties. In contrast, no obvious increment of naringin was yielded in the reaction system using CmCHI3 as the enzymatic protein, implying that it might not be a functional chalcone isomerase.
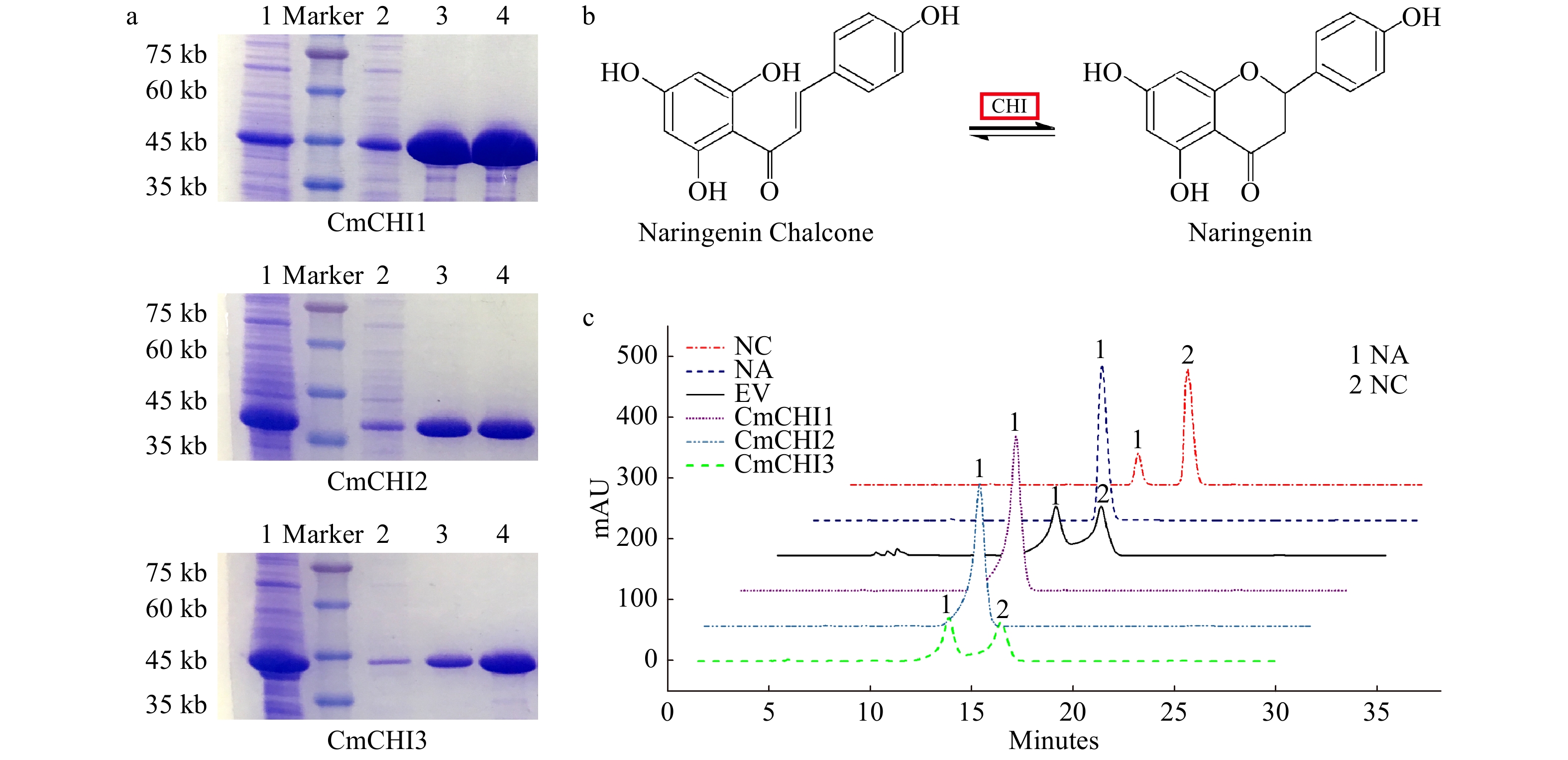
Figure 5.
In vitro enzymatic analysis of CmCHI1, CmCHI2 and CmCHI3. (a) The sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) of recombinant CmCHIs extracted from Escherichia coli BL21. 1, the induced CmCHI proteins without purification. 2, the uncombined CmCHI proteins after being processed by Ni column. 3−4, the purified CmCHI proteins eluted from Ni column with 20 mM or 100 mM imidazole in PBS. (b) The catalytic mechanism of chalcone isomerase on naringenin chalcone to produce naringenin. (c) HPLC analysis of the reaction products of CmCHIs and naringenin chalcone. Assay mixtures contained CmCHI protein extracts derived from E. coli harboring pET32-CmCHI1, pET32-CmCHI2 and pET32-CmCHI1 with NC as a substrate. Chromatograms were recorded at the UV absorbance wavelength of 304 nm. The identification of the naringenin product was confirmed based on relative retention time, UV spectra and the standard naringenin. NC, naringenin chalcone; NA, naringenin; and EV, empty vector.
CmCHIs can complement the deficient phenotypes of Arabidopsis tt5-1 mutant
-
To further demonstrate the role of CmCHIs in planta, we explored the heterologous expression of CmCHIs in Arabidopsis to determine their endogenous roles in plants. As shown in Fig. 6a, the CmCHI1 and CmCHI2 transformants exhibited purple color in cotyledons and hypocotyls similar to the wild type Arabidopsis seedlings (Ler), while the CmCHI3 transformants showed no coloration, similar to what is observed in tt5-1 mutants (chi). The brown color in Arabidopsis seed coats mainly as a result of proanthocyanidin accumulation[37]. To check the proanthocyanidins in seeds, DMACA was used to stain the seeds and results showed only tt5-1 mutant seeds over-expressing CmCHI1 and CmCHI2 recovered proanthocyanidin accumulation (Fig. 6a). To further confirm whether the phenotypic changes of the transgenic lines was caused by the foreign CmCHIs, we verified the ectopic gene expression by RT-PCR (Fig. 6b), and CmCHIs occurred in respective transgenic lines but not in wild-type (Ler) and mutant tt5-1 Arabidopsis. In addition, we also detected total anthocyanin content in wild-type, tt5-1 mutant and transgenic lines. Consequently, tt5-1 mutants with CmCHI1 and CmCHI2 resulted in the recovery of anthocyanins, and the transformation of CmCHI3 did not lead to obvious anthocyanin recovery (Fig. 6c). The results indicated that CmCHI1 and CmCHI2 were real chalcone isomerases in C. miniata.
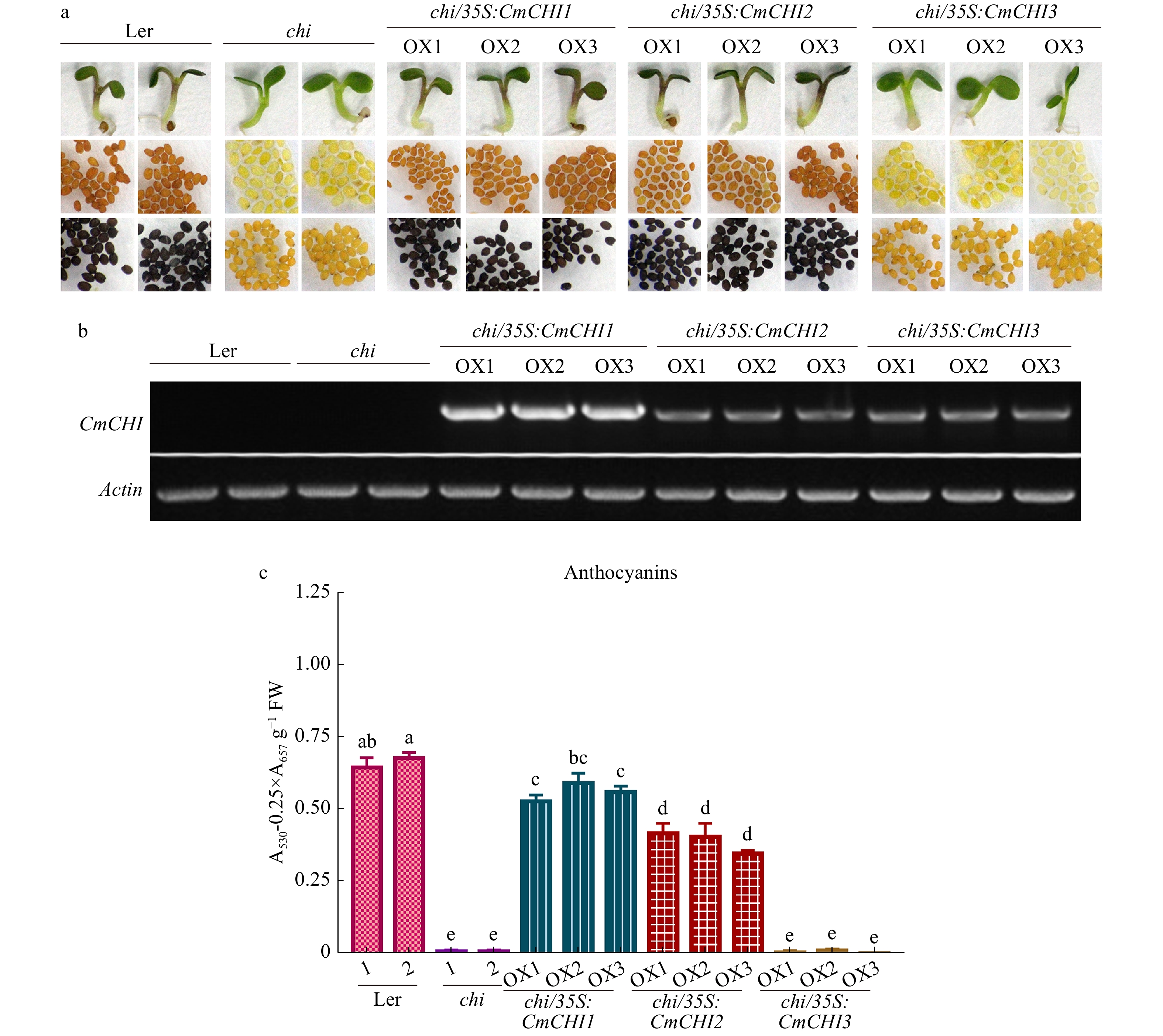
Figure 6.
Complementation of Arabidopsis tt5-1 mutant (chi) overexpressing CmCHI1, CmCHI2 and CmCHI3. (a) The seed coats, cotyledons and hypocotyls of wild-type (Ler-0), tt5-1 mutant (chi) and T2 transgenic lines. DMACA was used to stain and indicate proanthocyanidins in seeds. (b) Expression analysis of CmCHIs by reverse transcription polymerase chain reaction in the wild-type (Ler-0), tt5-1 mutant (chi) and transgenic lines. (c) Total anthocyanins in the wild-type, tt5-1 mutant (chi) and transgenic lines. Data represented means ± S.E. of three biological replicates. One-way ANOVA was carried out to compare statistical differences (Duncan's test, p < 0.05).
-
Diverse flavonoids are branched from the general phenylpropanoid pathway via the catalysis of CHS and CHI (Fig. 1)[9, 13]. Though isomerization of chalcones to flavanones can occur spontaneously in vivo, CHI-catalyzed intramolecular cyclization of chalcones is still a committed step in the production of flavonoids considering the abnormal phenotypes of chi-deficient mutants[38, 39]. As mentioned in the Introduction, there are four types of CHI proteins in CHI superfamily and the emergence of CHI proteins during evolution has attracted much attention as it is extremely rare for CHI to emerge from a non-catalytic protein to catalytic protein[29, 30].
CHI genes usually exist as a family in plants ranging from the basal bryophyte species to higher vascular plants[15, 32, 40, 41]. Only CHIs in type I and II are bona fide chalcone isomerases; however, in this study, we successfully cloned three CHI genes from C. miniata. Amino acid sequence analysis indicated several key residues in CmCHI proteins, indicating that CmCHIs were members of the CHI family (Fig. 2). Residues 190 and 191 were key to substrate specificity and composed one of the important indicators in determining type I or II proteins as revealed by earlier CHI's high-resolution three-dimensional structural analysis[42]. Particularly, residues 190 and 191 in type II CHI proteins were Thr and Met, while in type I were replaced by Ser and Ile[42, 43]. The alignment in Fig. 2a strongly suggests that CmCHI1 and CmCHI2 may be type I CHIs and are essential for the biosynthesis of anthocyanins and other flavonoids in plants, while CmCHI3 was only partially conserved in these sites and showed high similarity to type IV CHI (AtCHIL). The assumption was further validated by phylogenetic analysis (Fig. 2b). Evolution analysis of chalcone isomerase indicated the real type I and II CHIs evolved from type IV[29, 30]. Presumably, CmCHI3 may be the ancestor of CmCHI1 and CmCHI2, whereas CmCHI2 appeared to have evolved earlier than CmCHI1 as CmCHI2 rooted type I clade (Fig. 2b). Consequently, CmCHI2 may serve as a platform or bridge for future investigation of the key residue mutations determining CHI classification or how type I CHIs evolved, step by step, from type IV.
Consistent with the results of amino acid sequence comparison and phylogenetic analysis, the following prokaryotic analysis showed that almost all the substrate naringenin chalcone was completely catalyzed into naringenin by either CmCHI1 or CmCHI2 (Fig. 5). As expected, CmCHI3 had no catalytic activity in naringenin formation. The function of CmCHIs was also validated by genetic transformation of Arabidopsis tt5-1 mutant (Fig. 6). Generally, Type I CHIs found in most vascular plants, can only take 6'-hydroxychalcone (NA) as its substrate, whereas type II CHIs found in legumes, could use both 6'-hydroxychalcone (NA) and 6’-deoxychalcone as its substrates to synthesize isoflavones 5-deoxyflavanone and 5-hydroxyflavanone, respectively[28, 29, 32]. Whether CmCHI1 and CmCHI2 follow this would require further study.
The intracellular localization of flavonoids has been reported to exist in different cell compartments, such as chloroplasts, vacuoles, and nuclei[44−46]. Presumably, most of the enzymes acting in flavonoid biosynthesis were deemed to localize in the cytoplasm or membrane such as endoplasmic reticulum (ER). For example, flavonoid related type II CHIs were located in ER while the more ancient type III CHIs were located in the chloroplast[38, 41]. In the present study, CmCHIs had diffuse distributions in the cytoplasm (Fig. 3). However, more detailed analysis, such as organelle markers or use of microscopes with higher resolution, are still needed to accurately assess their localizations.
As the isomerization of chalcone is somehow spontaneous, and CHIs have been shown to be expressed spatially. OsCHI was expressed in the internode and leaf sheath of Oryza sativa and high expression was found in the leaves and panicles[47]. OjCHI in Ophiorrhiza japonica was expressed in all organs examined, including the roots[48]. In this study, the expression of CmCHI1 correlated well with anthocyanin accumulations in colored tissues (Fig. 4). Though the functional CmCHI2 was expressed in parallel with anthocyanin accumulations in leaf bases and fruit peels, it had negative correlations with anthocyanin accumulations in flower development. It is possible that CmCHI1 and CmCHI2 were functionally redundant in these tissues, and the most recently evolving CmCHI1 could be on its way to replace the older CmCHI2. Another possibility is that more precise functional differentiations have emerged between CmCHI1 and CmCHI2, such as response to different biotic or abiotic stresses, which need to be further investigated. As to the type IV CmCHI3, no isomerase activity was detected either in the enzymatic assay or in the genetic transformation of Arabidopsis tt5-1 mutant. Several previous studies however indicated that type IV CHI protein was characterized to be an enhancer of flavonoid production, and its function may be conserved among diverse land plant species[15, 31, 32, 40]. The high expression levels of CmCHI3 in red leaf bases or fruit peels also may indicate its potential role as an enhancer (Fig. 4). Further more detailed studies are needed to completely uncover the roles of CmCHIs.
-
In this study, three CmCHI genes were cloned from C. miniata via transcriptomic sequencing. The encoded proteins have cytoplasmic localizations and functionally cluster into two groups: CmCHI1 and CmCHI2 are type I proteins which are reported to be typical isomerases in vascular plants; whereas CmCHI3 belongs to type IV CHI proteins with undefined functions. Gene expression analysis indicates CmCHIs correlate with anthocyanin biosynthesis in C. miniata. Further enzymatic assays in vivo and the genetic transformation of Arabidopsis tt5-1 mutant, validate that CmCHI1 and CmCHI2 are real chalcone isomerases, while CmCHI3 has no isomerase catalytic properties, which requires further investigation. The results presented in this paper provide new insight into CHIs in C. miniata, which could be important for the modification of flower colors through genetic engineering.
-
Clivia miniata and Arabidopsis thaliana were grown in a growth chamber at 22 ℃ in the day and 15 ℃ at night under a 16 h light/8 h dark regime. Philips conventional T5 lamps were employed to provide a light intensity of 38 μmol m−2 s−1. The development of C. miniata flowers were arbitrarily divided into four stages: stage 1, flowers with green buds; stage 2, flower buds with slight pigmentation; stage 3, partially opened flowers; stage 4, open flowers. The flowers at different developmental stages were randomly harvested from different perennial C. miniata individuals. Meanwhile, the fruit peels of C. miniata also changed its color from green to red when fully ripened. The differently colored fruit peels were collected from the perennial C. miniata individuals. The seeds were germinated, and seedlings were grown in a growth chamber. The color of the young seedlings’ leaf bases could be changed when exposed to high light with a photon flux density at 115 μmol m−2 s−1. The leaf bases were gathered based on the different colors observed. All samples were immediately frozen in liquid nitrogen and stored at −80 ℃.
Nucleic acid extraction and cDNA synthesis
-
The Omini Plant RNA kit (both from CWBIO, Beijing, China) was used to extract total RNA from C. miniata or Arabidopsis according to the manufacturers instructions. The nucleic acid quality and concentration were measured by agarose gel electrophoresis and a Nanodrop
1000 spectrophotometry (ThermoFisher Scientific, Waltham, MA, USA). 500 ng of total RNA was reversely transcribed into cDNA by the UEIris II RT-PCR System for First-Strand cDNA Synthesis Kit (US EverbrightVR Inc., Suzhou, China) following the manufacturers instructions. 10 μg RNA samples were extracted from each of the flowers, leaves and fruit peels of C. miniata and were mixed together for transcriptome analysis by next-generation sequencing method, to construct a C. miniata transcriptomic database (Novogene, Beijing, China).Gene cloning and sequence analysis
-
The Arabidopsis AtCHI (locus:AT3G55120) was used as bait sequence to BLAST search the potential CHI genes against the constructed C. miniata transcriptomic database using TBLASTN algorithm[49−51]. The obtained sequences were analyzed by a manual BLASTX search of the National Center for Biotechnology Information site (NCBI,
https://www.ncbi.nlm.nih.gov ). Candidate genes were amplified with specific primers (Supplementary Table 2) and cloned into pESI-T vector by Hieff Clone® Zero TOPO-TA Cloning Kit (Yeasen, Shanghai, China) for sequencing confirmation. The CHI sequences from other plants were gathered from Genbank (http://www.ncbi.nlm.nih.gov/Genbank ) and processed using Clustal Omega online with default parameters (http://www.ebi.ac.uk/Tools/msa/clustalo ). MEGA version X was employed to generate a neighbour-joining phylogenetic tree. A total of1000 replicates were set as the bootstrapping number and gaps were handled with pair wise deletion.Subcellular localization analysis of CmCHIs proteins
-
The Minerva Super Fusion Cloning Kit (US Everbright® Inc., Suzhou, China) was employed in the following vector construction. The complete open reading frame (ORF) sequences of CmCHI genes were seamlessly cloned into NdeⅠ and AflⅡ digested vector 35S: GFP-FhMYB27 to construct 35S: GFP-CmCHIs in the pUC19 backbone[51]. The plasmids were then extracted by GoldHi EndoFree Plasmid Maxi Kit (CWBIO, Beijing, China) and reduced to a final concentration of 3−4 mg μL−1 by isopropanol and NaCl[51, 52]. Arabidopsis protoplasts were isolated by Cellulase R-10 and Macerozyme R-10 (both from Yakult Pharmaceutical Ind. Co., Ltd., Tokyo, Japan) and transfected by different plasmids following the method described previously[52−54]. After incubation in darkness at room temperature for 20−22 hours, the protoplasts were further detected with a fluorescence microscope for intracellular localization observation[52, 54].
Quantative real time PCR (qRT-PCR) analysis
-
In order to study the gene expression profile, specific primers for qRT-PCR were designed (Supplementary Table 2). The StepOne Plus Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) and SYBR Master Mix (TOYOBO, Osaka, Japan) were used with a reaction volume of 10 μL (5 μL of SYBR Master Mix, 0.2 μM for each primer and 1 μL of cDNA template). The reaction conditions were 95 ℃ for 5 minutes followed by a total of 45 cycles of 95 ℃ for 10 s and 60 ℃ for 10 s, and then 72 ℃ for 20 s. Ct values of Actin gene were used to normalize the reactions. The gene expression levels were calculated using the formula 2−ΔΔCт[55].
Protein preparation and purification
-
The prokaryotic protein was induced and purified following the method elsewhere (with modification)[19, 24, 56]. Briefly, the complete ORF sequences of CmCHI genes were subcloned into the expression vector pET-32a and transformed into E. coli strain BL21 (DE3). The positive colonies were pre-cultured into 5 mL LB (Luria-Bertani) liquid medium at 37 ℃ overnight. 3 mL of the culture was then transferred to 300 mL of fresh LB medium. When an A600 of 0.6 was reached, 0.5 mM isopropyl-β-D-thiogalactopyranoside (IPTG) was added to induce the recombinant proteins at 16 ℃ for 24 h. Afterwards, the cells were collected by centrifugation at 8,000 g for 15 min at 4 ℃ and suspended with 20 mL phosphate-buffered saline (PBS, pH 7.4) followed by sonication. After centrifugation at 13,000 g for 20 min, the supernatant containing crude proteins was processed by Ni Sepharose column (Sangon, Shanghai, China). Subsequently, the CmCHI proteins were purified by gradient imidazole in PBS (pH 7.4), desalted in PBS (pH 7.4) and concentrated in the silica gel dryer (Sangon, Shanghai, China). The concentrated protein was detected by Nanodrop
1000 spectrophotometer and further used for enzymatic activity analysis.In vitro enzymatic activity assay
-
The enzymatic assay was performed following Cheng et al (with some modification)[32]. In short, 30 μg of each purified recombinant protein was added to a reaction volume of 50 μL containing 5 μL of 1 mg mL−1 NC as substrate, 15 μL of 50 mM potassium phosphate buffer (pH 7.5) at 30 ℃ for 5 min. Protein extracts from BL21 (DE3) cells harboring empty pET-32a (+) vector were used as negative control. 100 μL of ethyl acetate was then added and mixed before being centrifuged at 12,000 g for 5 min. The aqueous layer containing the products was subjected to further high-performance liquid chromatography (HPLC) analysis equipped with an ACCHROM XUnion C18 column (250 mm × 4.6 mm, 5 µm). The column was eluted by the mobile phase containing 45% methanol, 3% acetic acid and 52% ultrapure water with a flow rate of 0.8 mL min−1. Detection was monitored at 304 nm.
Plant transformation
-
The complete ORF sequences of CmCHI genes were seamlessly subcloned into pBI121 vector and transformed into Agrobacterium tumefaciens strain GV3101. A 5 to 6-week-old Arabidopsis mutant (ABRC stock number: CS86, tt5-1) with a few mature flowers was used for plant transformation by the classical floral dip method[57]. The seeds were harvested, surface-sterilized by sodium hypochlorite and germinated on 1/2 MS medium supplemented with 50 ng mL−1 kanamycin and 25 ng mL−1 carbenicillin. 50 ng mL−1 kanamycin was further employed to screen T2 transgenic lines. Finally, seeds of landsberg-0 (Ler-0) ecotypic, tt5-1 mutant and T2 transgenic lines were either stained with 0.3% DMACA (4-dimethyla-minocinnamaldehyde) solution in ethanol or germinated on 1/2 MS medium supplemented with 3% w/v sucrose. About 6-day-old seedlings were collected from the medium for further anthocyanin content and gene expression analysis.
Total anthocyanin analysis
-
Total anthocyanin contents were measured following the method described in previous studies[50]. 0.1−0.15 g of samples were ground into powder in liquid nitrogen and soaked in 1 mL of acidic methanol solution (containing 1% hydrochloric acid) at 4 ℃ overnight in the dark. The mixture was then centrifuged at 12,000 g for 5 min to collect the supernatant. 400 μL of supernatant was mixed with 600 μL of acidic methanol as prior. The sample absorbance was detected at 530 nm (A530) and 657 nm (A657) by a spectrophotometer (BioTEK, USA). Total anthocyanin content was represented by the formula (A530−0.25×A657) g−1 fresh weight (FW).
Statistical analysis
-
The materials collected from C. miniata were sampled from different individuals to form one replicate. For the overexpression assays, the phenotypes were deemed as convincing when they independently appeared in at least 6 lines. Three transgenic lines were subjected to further analysis. One-way ANOVA was carried out by SPSS (v. 17.0) to compare statistical differences among groups with Duncan’s test. Significant differences were marked with the highest value at p < 0.05. The subcellular localization and in vitro enzymatic analysis were repeated at least three times.
This work was supported by the National Natural Science Foundation of China [31900252, 31972445]; the Department of Science and Technology of Jilin Province [20190201299JC, 20190303095SF, 20130604037TC]; the Jilin Province Development and Reform Commission [2020C024-5]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
-
The authors declare that they have no conflict of interest.
- Supplementary Table 1 Information of CmCHI1, CmCHI2 and CmCHI3.
- Supplementary Table 2 Primers used in the study.
- Copyright: © 2021 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Xue X, Zhao C, Zhang J, Liu M, et al. 2021. Cloning and Functional Characterization of Chalcone Isomerase Genes Involved in Anthocyanin Biosynthesis in Clivia miniata. Ornamental Plant Research 1: 2 doi: 10.48130/OPR-2021-0002 

Cloning and Functional Characterization of Chalcone Isomerase Genes Involved in Anthocyanin Biosynthesis in Clivia miniata
- Received: 21 December 2020
- Accepted: 25 January 2021
- Published online: 12 April 2021
Author Correction to this article was published on 13 April 2026, https://doi.org/10.48130/opr-0026-0009.
Abstract: Chalcone isomerase (CHI), catalyzing isomerization of chalcones, is a crucial enzyme in flavonoid biosynthesis. Three CHI genes were isolated from Clivia miniata and designated as CmCHI1, CmCHI2 and CmCHI3, respectively. Multiple sequence alignments and phylogenetic analysis showed that CmCHI1 and CmCHI2 were members of type I CHI proteins, whereas CmCHI3 belonged to type IV CHI proteins. Subcellular localization analysis found that all three CmCHIs had diffused distribution in the cytoplasm similar to green fluorescent protein (GFP). Anthocyanin biosynthesis and gene expression analysis demonstrated that CmCHIs were highly expressed in anthocyanin accumulated tissues. To further functionally characterize the role of CmCHIs, an in vitro enzymatic activity assay was carried out using the purified recombinant proteins. Results showed that CmCHI1 and CmCHI2 could completely convert the substrate naringenin chalcone (NC) into the product naringenin (NA), whereas CmCHI3 seemed nonfunctional as no increment of NA was detected. Further genetic transformation of Arabidopsis tt5-1 mutant validated that CmCHI1 and CmCHI2 rather than CmCHI3 could complement the chi deficient phenotypes. In summary, CmCHI1 and CmCHI2 are the real active CHI genes in Clivia miniata. The results not only broaden our knowledge on flavonoid biosynthesis in C. miniata but also lay a new foundation for further flavonoid modification in C. miniata.