








-
With changes in climate and precipitation patterns caused by global warming, drought conditions continue to be of serious concern. Lack of water inhibits the growth and development of plants, which ultimately leads to severe yield reduction and plant death[1]. Drought has seriously affected agricultural development and ecological sustainable development, and has gradually become a restricting factor in large-scale cultivation of chrysanthemums. The problem of drought resistance of plants can be solved through physiological regulation, population adaptation and genetic improvement. Among them, the use of genetic manipulation to cultivate drought-tolerant varieties is one of the effective methods to combat drought stress.
Phospholipid as an integral part of the membrane skeleton is an important eukaryotic cell component, and its metabolites are involved in regulating a variety of cellular functions and signal transduction. Phospholipases are important enzymes in the phospholipid signaling pathway. According to the hydrolytic parts of phospholipids, phospholipases can be divided into five categories including phospholipase A1 (PLA1), phospholipase A2 (PLA2), phospholipase C (PLC) and phospholipase D (PLD), of which PLD is the most abundant phospholipase occuring in plants. The hydrolysis products of PLD are phosphatidic acid (PA) and hydroxy compound. Recently, research on PLD and its hydrolysis products in cellular signal transduction has deepened. PLD in castor was the first to be isolated by reverse transcription[2], then PLD genes have been cloned from many plants such as Arabidopsis[3], maize[4], cowpea[5], tomato[6], tobacco[7], poppy[8], peanut[9], peach[10], sunflower[11] and soybean[12].
According to sequence characteristics and biochemical properties, 12 Arabidopsis PLD genes are divided into: PLDα (1, 2, 3), PLDβ (1, 2), PLDγ (1, 2, 3), PLDδ, PLDε and PLDζ (1, 2)[13]. PLDs cloned from plants belong mainly to the PLDα subgroup. AtPLDα encodes 809 amino acids. The sequence similarity of PLDα from different plants ranges from 75% to 90%. All members of PLD have two HKD motifs (HXKXXXXD) at intervals of 320 amino acids, where H represents His (histidine), K represents Lys (lysine) and D represents Asp (aspartic acid)[14].
PLD plays a role in the regulation of plant growth and development, plant hormones, abiotic stresses tolerance, disease resistance through lipid degradation, reconstruction of the membrane and cytoskeleton and vesicle trafficking. Drought enhanced expression of PLD and activity in drought sensitive cowpea varieties, while PLD expression is not affected in the drought-resistant varieties[5]. PLDα-depleted plants are more sensitive to moisture loss, and the leaves of PLDα-overexpressing plants are more sensitive to ABA compared to wild type. Under drought stress, PLDα generates PA to regulate stomata opening and closing in ABA signaling[15]. PA binds to ABI1 (protein phosphatases 2C), and anchored ABI1 from the cytoplasm to the cell membrane, consequently inhibiting the negative regulatory role of ABI1 on stomata closure[16]. In addition, the interaction of G protein and PLDα1 enhances the ABA inhibitory effect on stomata opening[17].
Drought hampers the yield and quality of cut chrysanthemum, a leading ornamental plant globally. A number of genes and transcription factors involved in stress tolerance regulation have been explored[18−21]. However, how phospholipase participates in drought tolerance regulation has not been reported in chrysanthemum. Here we cloned CmPLDα from chrysanthemum 'Jinba', and generated CmPLDα transgenic chrysanthemum. Drought tolerance assays showed that CmPLDα enhanced the drought tolerance of chrysanthemum by maintaining water balance and membrane integrity.
-
Chrysanthemum 'Jinba' was obtained from the Chrysanthemum Germplasm Resource Preserving Centre, Nanjing Agricultural University, China. Total RNA was isolated from chrysanthemum 'Jinba' leaves using the RNAiso reagent (TaKaRa, Japan). The degenerate PCR primers (DPF/R in Table 1) were designed based on a peptide alignment of the PLD sequences of Helianthus annuus (GenBank accession number ABU54776.1), Ricinus communis (AAB37305.1), Solanum lycopersicum (AAF17557.1), Litchi chinensis (ADP23922.1), Citrus sinensis (ACA49723.1), Dimocarpus longan (ADY75750.1). RACE technology was used to isolate the full length CmPLDα cDNA sequence following the description of Song et al.[20].
Table 1. Primer sequences used in this study.
Primer Sequence (5'–3') Usage DPF TGCATGCTGGTGTGGGAYGAYMGNAC Degenerated PCR DPR CGTCCATGGACCGCTGRTTDATRTT / Oligo (dT) prime GACTCGAGTCGACATCGATTTTTTTTTTTTTTTTT / dT-AP AAGCAGTGGTATCAACGCAGAGTACTTTTTTTTTTTTTTTTT 3'-RACE AP-R AAGCAGTGGTATCAACGCAGAGTAC / GSP1 TCACGCTATAAGAAGAGCAA / GSP2 GGACTGGCAAAGGAGGACTA / RGSP1 TAAACCGACCATCCCGTG 5'-RACE RGSP2 CGCACTAGTAATGGCATCGAATATA / RGSP3 ACATCTGTGGGGTTCGTAATACTTC / AAP GGCCACGCGTCGACTAGTACGGGIIGGGIIGGGIIG / AUAP GGCCACGCGTCGACTAGTAC / PLDSP ATGGCTCAGATACTGCTCCATGGTA ORF amplifications PLDAP CAATGCAAACATGGCTTATTACATC / qGSP-F CACTTGCTTCGGTACCCTATTG qRT-PCR qGSP-R ACACAACCACCAAACATGACCT / EF1α-F TGTAACAAGATGGATGCCACAA / EF1α-R TCGCCCTCAAACCCAGAAAT / SmaI-SF TCCCCCGGGGGAATTCGATTCCAAGATTATGGCTCAG Vector construction SacI-SR CCGAGCTCGGCACACAACCACCAAACATGACCTTA / SmaI-AF TCCCCCGGGGGACACACAACCACCAAACATGACCTTA / SacI-AR CCGAGCTCGGATTCGATTCCAAGATTATGGCTCAG / NPTII-F TCTGATGCCGCCGTGTTC Transgenic detection NPTII-R GATGTTTCGCTTGGTGGTCG / Protein structure and phylogenetic analysis of CmPLDα
-
The GenBank BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was applied on sequence homology analysis. The open reading frame (ORF) of the gene was found by DNAMAN v9.0 software. The homologous amino acid sequence was aligned using ESPript3.0 (https://espript.ibcp.fr/ESPript/ESPript/index.php). The tertiary structure was constructed using SWISS-MODEL (https://swissmodel.expasy.org) and a phylogenetic tree was constructed using Clustal X and MEGA X software.
Expression vector constructs and generation of transgenic chrysanthemum
-
The pBIG vector driven by CaMV 35S promoter was employed. Two pairs of primer SacI-SF, SmaI-SR and SacI-AF, SmaI-AR (Table 1) were used to amplify the ORF of CmPLDα, pBIG- (+/−) -CmPLDα was constructed using the restriction enzyme of SacI and SmaI. pBIG- (+/−) -CmPLDα was transformed into Agrobacterium tumefaciens EHA105 using the heat shock method[22]. Leaf discs measuring 0.5 cm × 0.5 cm were prepared from 25−30 day old in vitro chrysanthemum 'Jinba' seedlings, then were cultivated on MS medium with 1.0 mg·L−1 6-BA, 0.5 mg·L−1 NAA. The Agrobacterium infected discs were transferred to MS medium with 1.0 mg·L−1 6-BA, 0.5 mg·L−1 NAA, 10 mg·L−1 kanamycin and 300 mg·L−1 carbenicillin. Kanamycin-resistant shoots were rooted on MS medium with 7 mg·L−1 kanamycin and 200 mg·L−1 carbenicillin[23,24]. Kanamycin-resistant plants were identified using a PCR assay with the primer pair NPTII-F/R (Table 1). The PCR programs were 95 °C/3 min, 35 cycles of 94 °C/30 s, 55 °C/30 s, 72 °C/30 s, 72 °C/10 min. Quantitative Real-Time PCR was used to determine the expression of CmPLDα in the transgenic lines with the primer pair qGSP-F/R and the reference primer pair EF1α-F/R (Table 1). 20 μl RT-qPCR reaction consisted of 10 ng cDNA, 0.2 mmol·L−1 of each primer and 10 μl SYBR Green PCR master mix. The PCR regime contained a denaturation step of 95 °C/60 s, followed by 40 cycles of 95 °C/15 s, 60 °C/15 s, 72 °C/45 s. Relative expression levels were calculated using the 2−ΔΔCT method[25].
Drought stress treatment
-
Cuttings of chrysanthemum 'Jinba' were rooted in a substrate comprising of perlite and vermiculite in a 1:1 ratio based on volume in a greenhouse with a photoperiod of 16 h/8 h (light/dark) and a relative humidity of 70%. The day/night temperature was 23 °C/18 °C and the light intensity 100 μmol·m−2S−1, respectively. Rooted plants at the 6−8 leaf stage were subjected to a mimicked drought treatment using 20% w/v polyethylene glycol (PEG) 6000 for 24 h. The leaves were harvested after 0, 1, 4, 8, 12 and 24 h following the PEG treatment. Harvested leaves were frozen in liquid nitrogen and stored at −80 °C. Survival rates were monitored after a week of recovery, where the roots of the PEG treated plant were washed in tap water for 5 min, then plants were placed in fresh water for one week for recovery. The experiment includes three replicates
Determination of drought tolerance and physiological changes
-
Morphological appearance was documented by photographic recording. The wilting index was divided into 7 levels (0−6) according to the degree of leaf wilting[26]. Leaf relative water content (RWC) = (fresh weight − dry weight)/fresh weight × 100%. Relative electrical conductivity (REC) = (initial conductivity/electrical conductivity after heating and cooling) × 100%. Malondialdehyde (MDA) assay was conducted using thiobarbituric acid colorimetry[27]. Leaf samples in triplicate were extracted with 80% methanol, and Abscisic acid (ABA) contents were analyzed using HPLC based on the method described by Chen and Yang[28].
Data analysis
-
Data were presented as mean ± standard deviation (S.D.). All data were subjected to analysis of variance (ANOVA) using SPSS 20.0 (IBM Corp, Somers, NY, USA). The least significant difference (LSD) multiple range test was used to analyze the results after one-way analysis of variance. Data mapping were performed using GraphPad Prism 5 software (San Diego, CA, USA).
-
Based on the conserved sequences of PLDα from other plants, a putative PLDα homologous gene designated as CmPLDα was isolated from chrysanthemum by RT-PCR and RACE-PCR methods. The CmPLDα cDNA consists of 2,697 bp with a 2,427 bp ORF encoding for 809-amino acid proteins. A homology blast showed that CmPLDα contains conserved PLDα domains, two HKD motifs and C2 domain (Fig. 1b). CmPLDα is substantially homologous to other PLDα from other species with a sequence identity between 71% and 88%, and CmPLDα showed the highest similarity to that of Helianthus annuus (Fig. 1a). The secondary structure of CmPLDα contains 30 β-strands, 24 β-turns, 15 α-helices and 8 η helices (Fig. 1a). These structures constitute the tertiary structure of CmPLDα (PDB: 6kz9, sequence identity: 78.99%, Fig. 1c). Phylogenetic analysis showed that CmPLDα clustered with PLDα1 members from Compositae such as Cynara cardunculus, Helianthus annuus, Lactuca sativa and Artemisia annua (Fig. 2).

Figure 1. Sequence analysis of CmPLDα. (a) Alignment of CmPLDα with other known PLD homolog proteins. Secondary structure elements are shown at the top (helix with wavy lines, β-strand with arrows, and turns with TT letters). (b) C2 domain (existed between 9−125aa) and two HKD motifs (existed between 332−339aa and 660−667aa). (c) The tertiary structure of CmPLDα (PDB: 6kz9) with sequence identity 78.99% using homolog modeling.
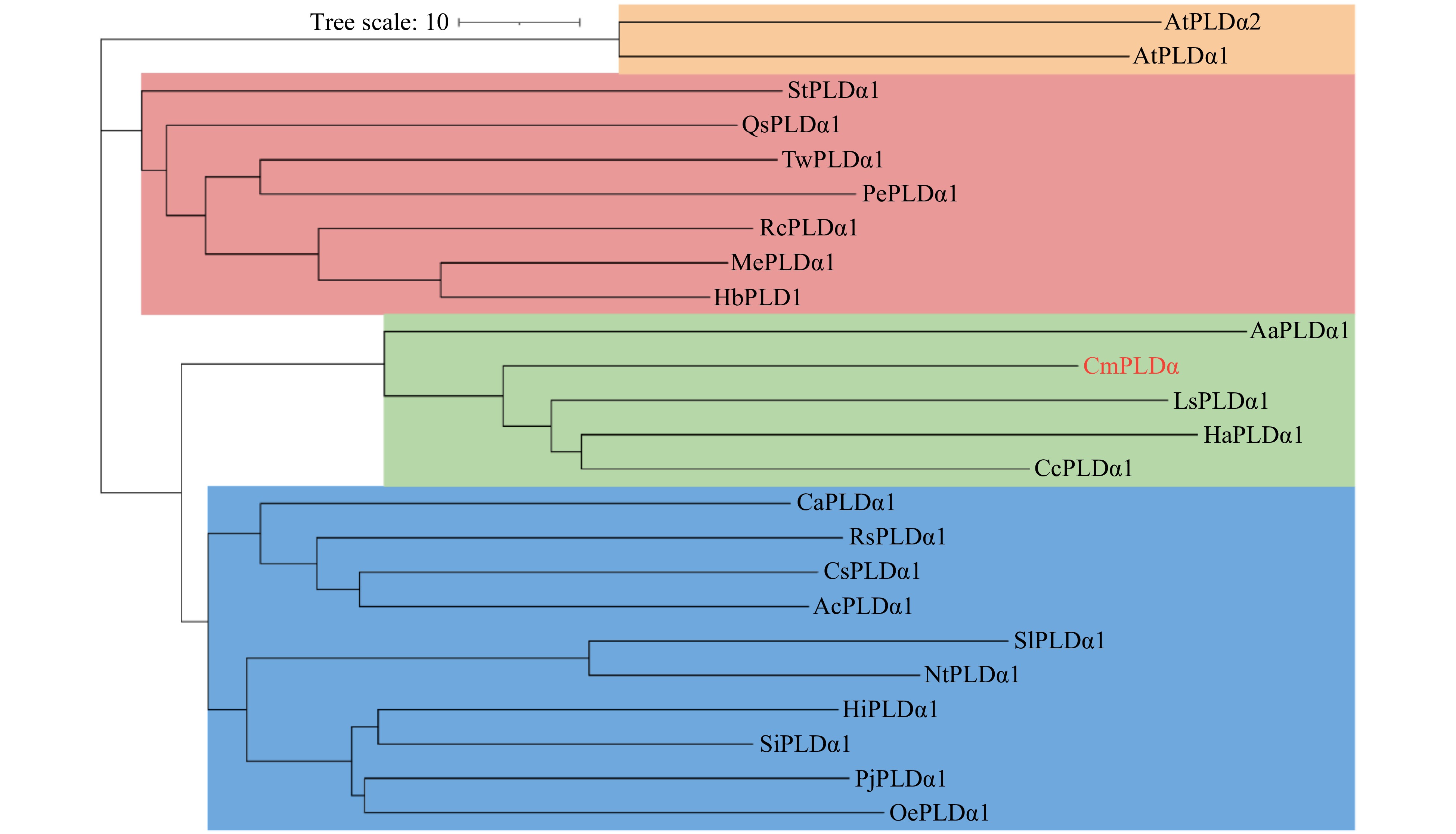
Figure 2. Phylogenetic tree analysis of CmPLDα. The phylogenetic tree revealed four branches and the red font indicates CmPLDα. The source and accession number of the amino acids are as follows: AtPLDα1 (Arabidopsis Thaliana NP_188194.1) , AtPLDα2 (Arabidopsis Thaliana NP_175666.1), StPLDα1 (Senna tora KAF7814366.1), QsPLDα1 (Quercus suber XP_023925867.1), TwPLDα1 (Tripterygium wilfordii XP_038717853.1), PePLDα1 (Populus euphratica XP_011008452.1), RcPLDα1 (Ricinus communis NP_001310687.1), MePLDα1 (Manihot esculenta XP_021629145.1), HbPLDα (Hevea brasiliensis XP_021672614.1), AaPLDα1 (Artemisia annua PWA90284.1), LsPLDα1 (Lactuca sativa XP_023761181.1), HaPLDα1 (Helianthus annuus ABU54776.1), CcPLDα1 (Cynara cardunculus XP_024972255.1), CaPLDα1 (Coffea arabica XP_027126646.1), RsPLDα1 (Rhododendron simsii KAF7135150.1), CsPLDα1 (Camellia sinensis XP_028072531.1), AcPLDα1 (Actinidia chinensis PSR96034.1), SlPLDα1 (Solanum lycopersicum AAG45485.1), NtPLDα1 (Nicotiana tabacum XP_016458333.1), HiPLDα1 (Handroanthus impetiginosus PIN16148.1), SiPLDα1 (Sesamum indicum XP_011073436.1), PjPLDα1 (Phtheirospermum japonicum GFP88976.1) , OePLDα1 (Olea europaea CAA3025459.1).
CmPLDα transgenics
-
In total, three sense and four antisense CmPLDα transgenic lines were detected from the kanamycin-resistant lines using PCR analysis (Fig. 3a). The overexpression transgenic lines Z1 and Z2 lines showing 2.4 and 2.5 times up-regulated expression levels compared with the control, and the silencing transgenic lines F1 and F2 with 0.46 and 0.56 times down-regulated expression of the control were selected for subsequent assay (Fig. 3b).
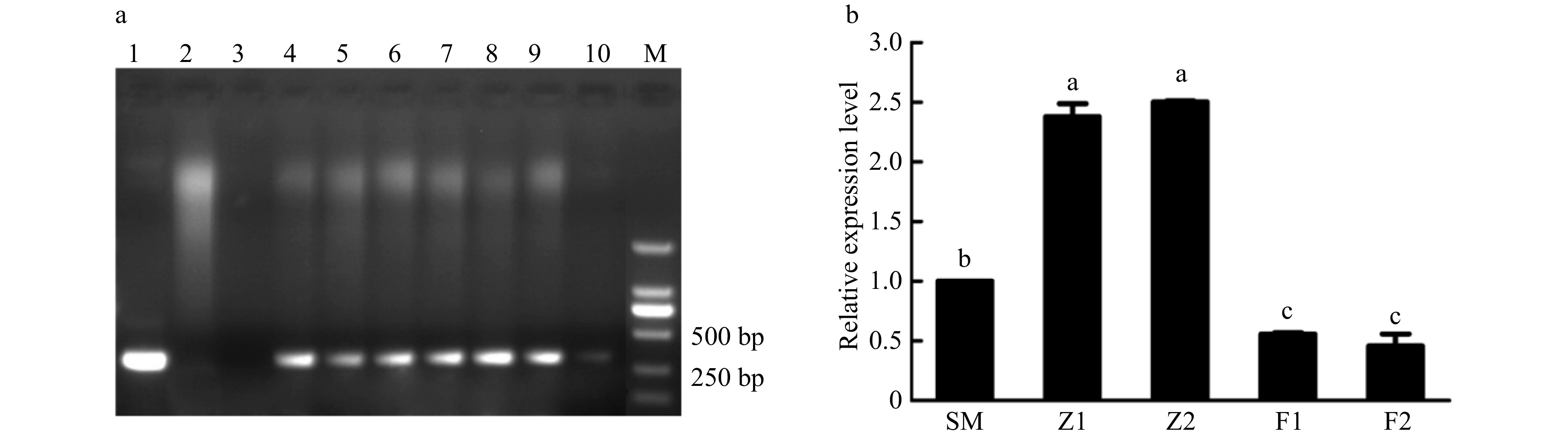
Figure 3. Detection of CmPLDα transgenic chrysanthemum plants. (a) PCR analysis of putative CmPLDα transgenic plants in the kanamycin resistant lines. M: DL2000, 1: Positive control (Plasmid pBIG-CmPLDα DNA), 2: Non-transformed plant, 3: Blank control (H2O), 4−6: CmPLDα overexpressing lines, 7−10: CmPLDα antisense transgenic lines. (b) The relative expressions level of CmPLDα in the wild type and transgenic plants. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines. Representative results from three biological replicates are shown. Values are mean ± S.D., and different letters indicate significant differences at p < 0.05 (Fisher's LSD).
Drought tolerance assay of transgenic plants
-
Following drought stress treatment, most plants of transgenic lines Z1, Z2 remained green, only the basal leaves wrinkled and wilted. Wilting index of Z1, Z2 were 3 (Fig. 4), and the survival rate was 85.4%−90.6% after the recovery growth (Fig. 5b), however, only a few leaves of the wild type plants remained green, and the wilting index of these was 4 (Fig. 4) with a survival rate of 52.4% (Fig. 5b). Of note for antisense lines F1, F2, most plants were heavily wilted, and the wilting indexes were 5 and 6 (Fig. 4), the survival rates were 33.7% and 41.3%, respectively (Fig. 5b).

Figure 4. The morphological response of the wild type and CmPLDα transgenic plants to PEG-induced drought stress. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines.
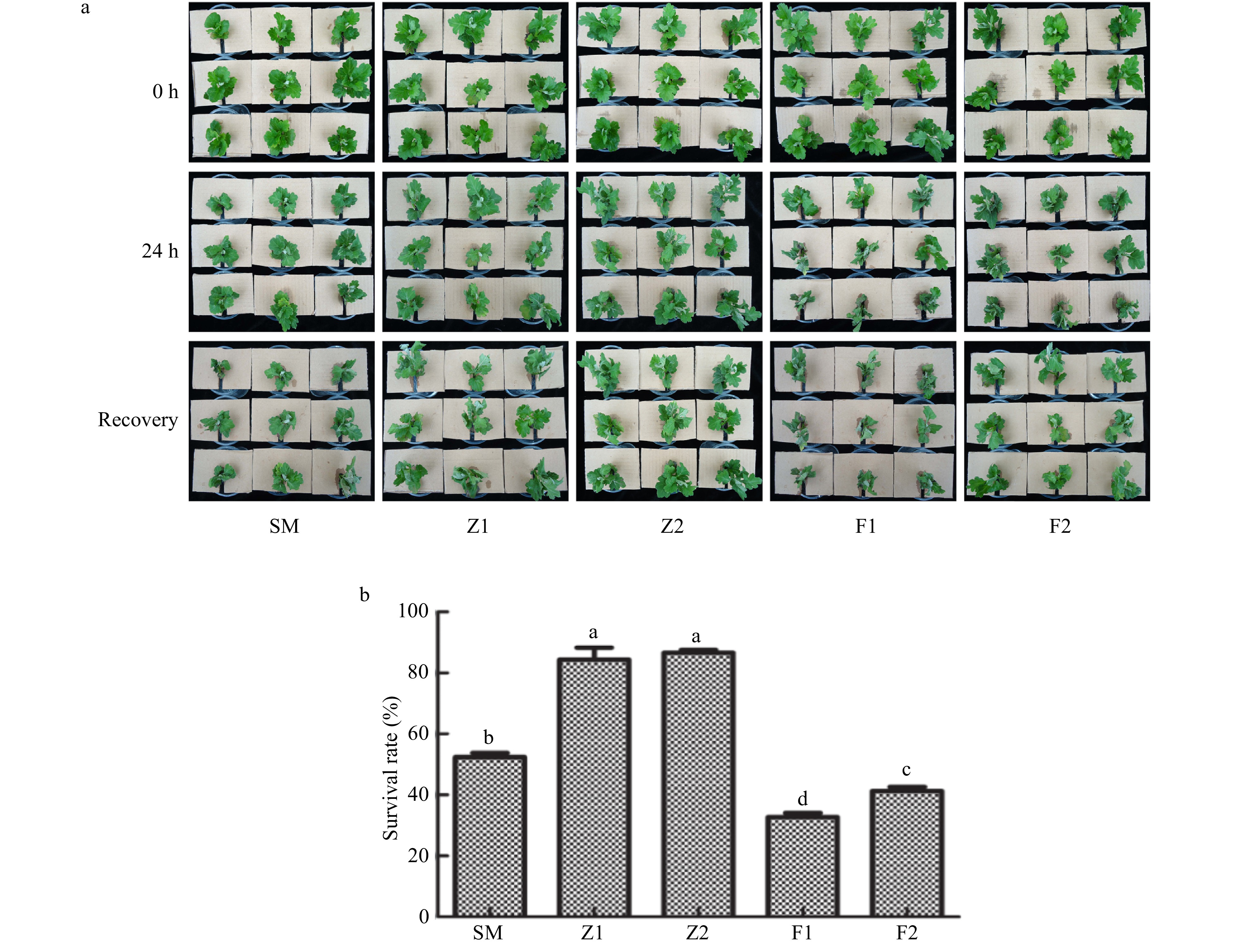
Figure 5. Drought tolerance assay of the wild type and CmPLDα transgenic plants. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines. (a) Survival rate of recovery growth. (b) Each value is the mean ± S.D. of nine biological determinations, different letters indicate significant differences at p < 0.05 (Fisher's LSD) when comparing values under the same treatment conditions.
The water content of the CmPLDα overexpression lines was significantly higher than that of the WT plants and antisense transgenic lines after 4 h of PEG treatment. After 12 h, the water content of the overexpression lines Z1, Z2 were 10%, 11.5% higher than that of the WT plants (Fig. 6). The water content of antisense lines F1, F2 were 3%, 5% lower than that of the control (Fig. 6), indicating that the CmPLDα gene can increase the water retention capacity of the plant.
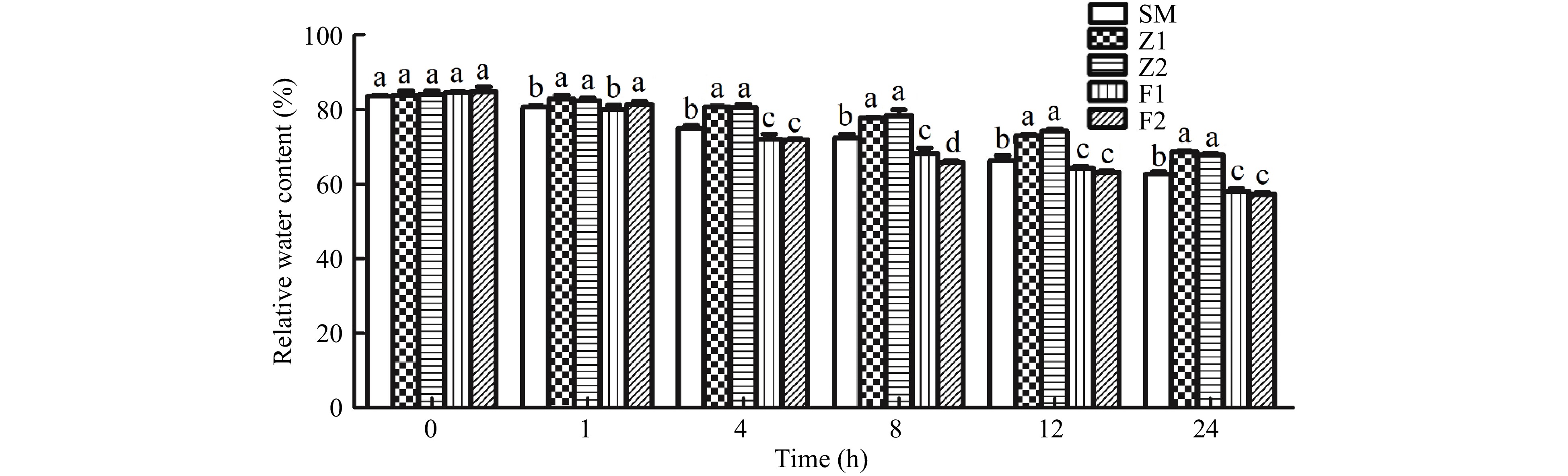
Figure 6. The leaf RWC in the wild type and CmPLDα transgenic plants under drought stress. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines. Each value is the mean ± S.D. of nine biological determinations, and different letters indicate significant differences at p < 0.05 (Fisher's LSD) when comparing values under the same time conditions.
After 12 h of PEG treatment, the relative electrical conductivity of the CmPLDα overexpression lines were 18.4%, 17.5% lower than that of WT plants, the relative electrical conductivity of the CmPLDα antisense lines were 24.8%, 21.1% higher than that of WT plants (Fig. 7a). MDA content in WT plants and the rate of increase of MDA was significantly greater than those of the CmPLDα overexpression lines Z1, Z2. After 24 h of PEG treatment, MDA content of the CmPLDα overexpression lines was 14.4%, 17.5% lower than that of WT plants (Fig. 7b). MDA content of CmPLDα antisense lines were significantly higher than that of the CmPLDα overexpression lines and WT plants (Fig. 7b), suggesting that CmPLDα provided a better membrane permeability in chrysanthemum in response to drought stress.
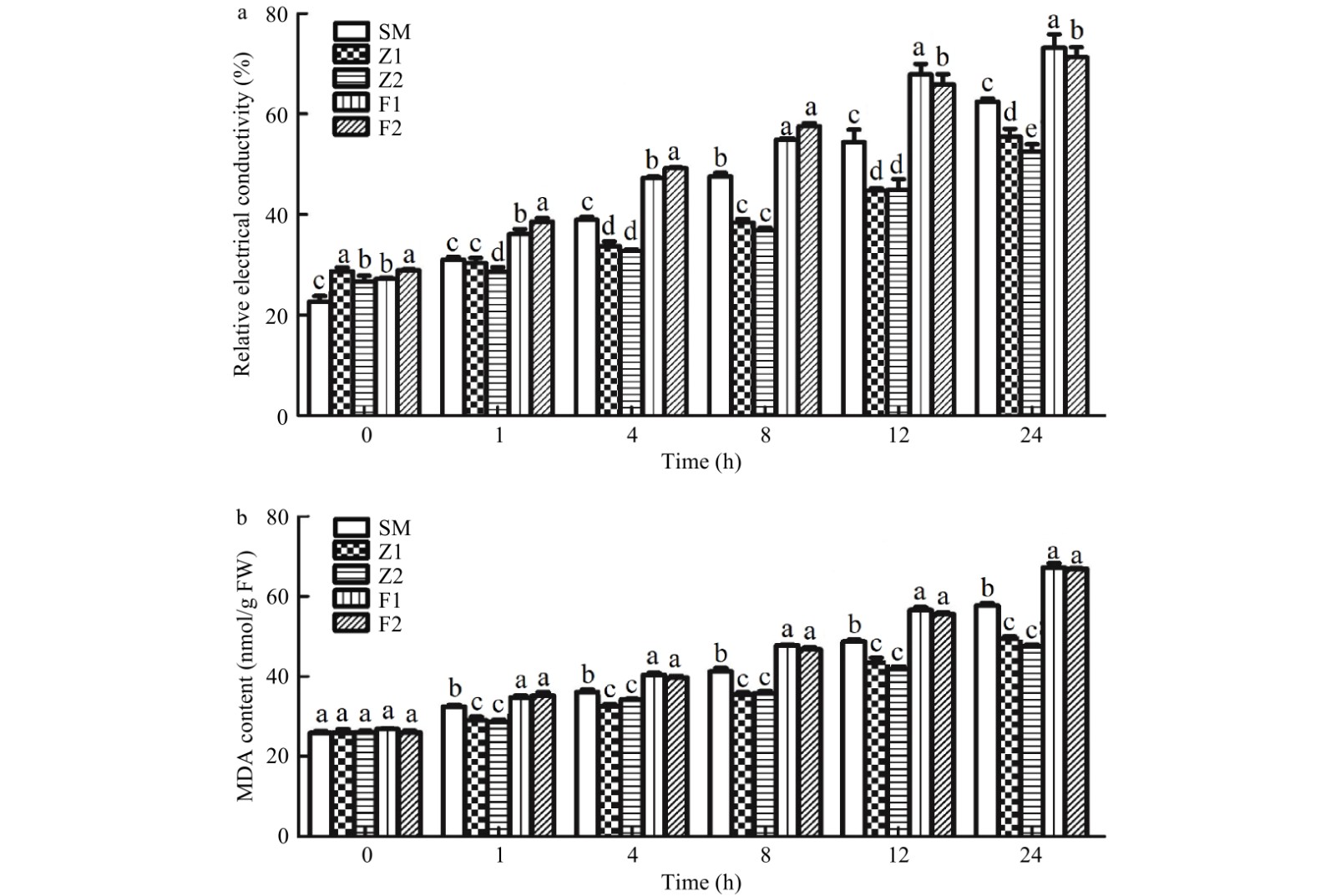
Figure 7. Relative electrical conductivity (a) and MDA content (b) in drought stressed leaves of wild type and CmPLDα transgenic plants. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines. Each value is the mean ± S.D. of three biological replicates, different letters indicate significant differences at p < 0.05 (Fisher's LSD) when comparing values under the same time conditions.
Changes in ABA content in different lines was detected (Fig. 8). After 1 h of drought stress, the ABA content of CmPLDα overexpression lines were significantly higher than that of WT plants. The ABA content was highest 24 h after drought stress treatment, and ABA content in the overexpression lines Z1, Z2 were 1.3, 1.22 times that of the WT (Fig. 8a). The ABA content of antisense transgenic lines F1, F2 were 0.83, 0.81 times that of WT plants at 8h drought treatment (Fig. 8a). The expression level of ABA synthesis key gene CmNCED was consistent with the changes in ABA content. At 4h of drought stress, gene expression levels of the overexpression lines were 10.9, 12.0 times that of the WT plants (Fig. 8c). The expression of the downstream gene CmRD29B were 4.1, 4.2 times that of WT plants at 1 h drought stress (Fig. 8b). The expression levels of CmNCED and CmRD29B of the CmPLDα antisense lines were lower overall than those of CmPLDα overexpression lines and WT plants (Fig. 8b, c).
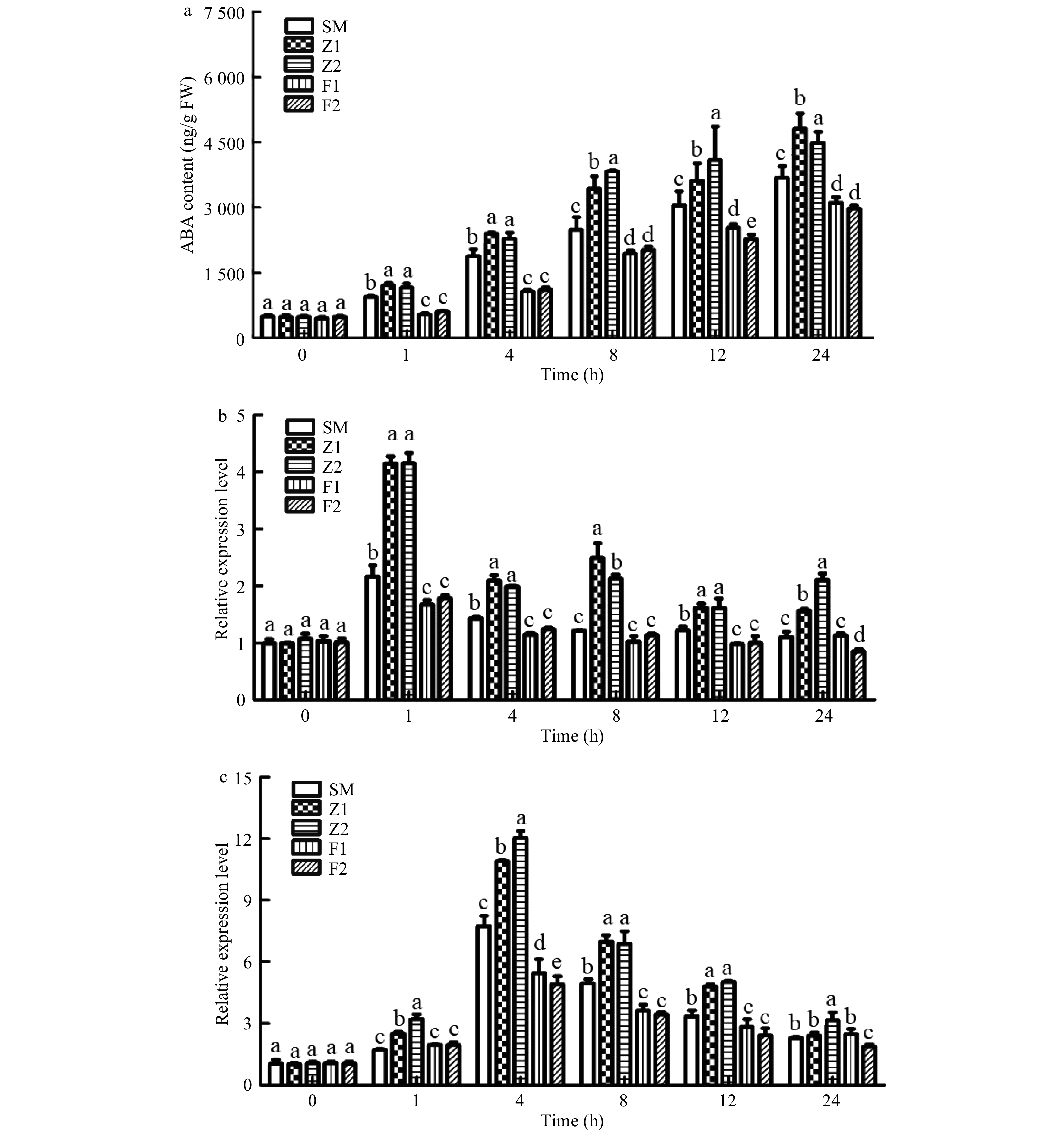
Figure 8. ABA content (a) and the relative expressions level of ABA-responsive gene CmRD29B (b) and ABA biosynthesis gene CmNCED3 (c) in wild type and CmPLDα transgenic plants under drought stress. SM: Wild type, Z1, Z2: CmPLDα overexpressing lines, F1, F2: CmPLDα antisense transgenic lines. Each value is the mean ± S.D. of three biological replicates, different letters indicate significant differences at p < 0.05 (Fisher's LSD) when comparing values under the same time conditions.
-
PLD from Arabidopsis[3], tobacco[7], tomato[6], sunflower[11] and other plants have been cloned, however, its homologue has not been isolated in chrysanthemum. Here we cloned a PLD from chrysanthemum, and the sequence characteristics and phylogenetic analysis showed that the cloned CmPLDα belongs to C2-PLDs of the PLDα family.
Previous studies have shown that PLD functions as a phospholipid transfer protein involved in the hydrolysis of phospholipids, the hydrolysis products such as phosphatidic acid (PA), diacylglycerol (DAG), free fatty acid (FFA) participate in stress responses or developmental regulation. In the present study, we showed that drought resistance of CmPLDα overexpressing chrysanthemum had been significantly improved. Similarly, poplar PLDα1 overexpressing plants had higher drought resistance than the wild type lines, but RNAi plant had lower drought resistance than wild type. Most of the CmPLDα overexpressing lines remained green, whereas most of the CmPLDα antisense transgenic plants wilted under PEG treatment. As an important indicator of plant water status, the higher relative water content (RWC) in the overexpression lines suggested that CmPLDα could maintain better water status to alleviate drought stress. Relative electrical conductivity and MDA content in CmPLDα overexpressing lines were lower than those in wild type and antisense lines, indicating that CmPLDα improved plant drought tolerance through maintaining membrane integrity of the plant, which is in line with previous studies[29,30].
It has been found that PLDα played an important role in stomate movement. PLDα1 improves plant drought tolerance by maintaining the plant cell membrane stability, and inducing stomate closure to reduce the loss of moisture when subjected to water stress[29,30]. ABA is an important plant hormone in the regulation of stomate opening and closing under stress. PLDα regulates stomate opening and closing through ABA. Leaves of PLDα overexpression were more sensitive to ABA[15]. PLDα1 participates in the ABA regulation of stomata movement mainly through two processes: PA anchored negative regulatory protein ABI1 from the cytoplasm to the cell membrane through interaction with ABI1, thereby inhibiting the negative regulatory role stomata closure of ABI1[16], PLDα1 and G protein both positively regulate ABA inhibition of stomata opening[17]. Hong et al. found that PLDα3 overexpressing and knockout lines had a small, yet significant, effect on ABA content, ABA content of overexpression lines were higher than knockout lines, RD29B expression levels of overexpression lines increased after drought stress[31]. In our study, expression of the ABA synthesis gene CmNCED and ABA content increased in CmPLDα overexpression lines compared to the WT plants, which may promote ABA regulated stomate closure. As one of Arabidopsis dehydration-induced genes, RD29B encoding hydrophilic proteins[32] may regulate genes participating in downstream resilience of ABA signal transduction, thereby improving the drought tolerance of transgenic plants. Here we also observed an increase in the expression of CmRD29B in CmPLDα overexpression lines, which again supports the hypothesis that CmPLDα enhanced drought tolerance is related to the ABA signaling pathway. In summary, CmPLDα could enhance drought tolerance in preventing partial water loss via ABA, stabilizing the membrane under drought stress.
- The work is funded by the National Key Research and Development Program of China (2018YFD1000402), the National Natural Science Foundation of China (32030098), Jiangsu Agriculture Science and Technology Innovation Fund (CX(18)2020).
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhai L, Zhu X, Yang S, Gu C, Liu P, et al. 2021. Constitutive expression of a chrysanthemum phospholipase Dα gene in Chrysanthemum morifolium enhances drought tolerance. Ornamental Plant Research 1: 8 doi: 10.48130/OPR-2021-0008

Constitutive expression of a chrysanthemum phospholipase Dα gene in Chrysanthemum morifolium enhances drought tolerance
- Received Date: 24 May 2021
- Accepted Date: 30 July 2021
- Published Online: 17 August 2021
Abstract: Drought causes water shortage and consequent retardation of plants growth and development. Therefore, improving the drought tolerance of plants is necessary for expanding cultivation and resource promotion. Increasing evidence indicates that phospholipase is involved in the response of plants to drought stress. The objective of this study was to create new drought-tolerant chrysanthemum germplasm, which lays a foundation for the study of the molecular mechanism of phospholipase mediated stress response in chrysanthemum. CmPLDα has the closest relationship with sunflower HaPLDα, and belongs to the PLDα family. CmPLDα over-expressing plants showed a slight shrinking under 20% PEG6000 treatment. The survival rate increased significantly by 1.7−1.8 times that of the wild type. Relative water content (RWC) of CmPLDα over-expressing plants were nearly 10% higher than that of the wild type. Relative electrical conductivity and MDA content were significantly lower than those of the wild type. ABA content of the over-expression lines Z1, Z2 were 1.3 and 1.22 times that of wild type, but ABA content of antisense lines F1, F2 was approximately 0.83 and 0.81 of those of wild type. Most plants of antisense transgenic lines F1, F2 were wrinkled, with a wilting index of 5 and 6, and the survival rate was also lower than that of the wild type after recovery growth. RWC of antisense lines were lower than over-expression lines, relative electrical conductivity and MDA content were significantly higher than those of the wild type. In summary, CmPLDα could enhance tolerance of chrysanthemum to drought conditions.
-
Key words:
- Chrysanthemum /
- CmPLDα /
- ABA content /
- Drought stress