









-
Fleshy fruit texture and whole-fruit eating experience are drivers of consumer acceptability, and removal of inedible elements such as seeds enhances marketability. For many fruits, seedless varieties have greater consumer appeal and often improved fruit quality, which has led to seedless varieties such as grapes, citrus, bananas, and watermelons outcompeting seeded cultivars for market share[1, 2]. In these crops, breeding programmes often focus on the seedless character as a primary attribute for new varieties[3]. In pipfruit (apple, pear), seedless varieties are available such as ‘Spencer Seedless’, ‘Rae Ime’, and ‘Wellington Bloomless’, but market domination is not achieved because while seeds are removed, the cartilaginous core remains, which is unpalatable and inedible for most consumers.
Fruit morphology and sub-structures of the fruit are determined by the morphology of the flower. Most fruit derive from carpel tissues, while others, such as apple and strawberry, derive from extra-carpellary tissues such as floral tube/hypanthium and receptacle, respectively. In turn, the four organ types of a perfect flower are determined by floral organ identity genes, as described by the ABCE model[4]. In this model, most genes are MADS-box transcription factors, including A-type APETALA1 (AP1), B-type PISTILLATA (PI) and APETALA3 (AP3), C-type AGAMOUS (AG) and E-type SEPALLATA1-4 (SEP). Specific combinations of ABCE type transcription factors interact in quaternary complexes to determine organ identity in each floral whorl[5]. Carpel identity is determined by E- and C-type transcription factors, while B-, C-, E-type combination determines stamen identity. C-type AG therefore contributes to both carpel and stamen identity, collectively known as C-function.
The function of E- and C-type transcription factors often varies between species. In dry-fruiting Arabidopsis, loss of C-type AG function resulted in conversion of stamens to petals, and carpels to indeterminate flowers with sepal-petal-petal patterning and owing to loss of carpel tissues, no siliques were produced[6]. In tomato, loss of C-type AG with precision gene editing techniques had weaker effects, generating flowers with altered stamen identity and carpels with loss of determinacy exhibited by seedless fruit and fruit-in-fruit phenotypes[7, 8]. In apple, suppression of C-type AG increased the number of petal whorls, like Arabidopsis, but did not abolish carpel tissues and fruit were produced[9]. In Arabidopsis, considerable efforts have sought to identify AG-regulated gene networks controlling floral organ identity[10, 11]. Partial redundancy in function is supplied by genes of a monophyletic clade with AG which are additionally regulated by AG, namely SHATTERPROOF (SHP1/2)[12] and SEEDSTICK (STK)[13], which were found to control AG-independent carpel and ovule identity determination[13]. In contrast to the conserved nature of the AG monophyletic clades across species, E-type transcription factors are more functionally diverse owing to extensive clade expansions in this family[14]. Arabidopsis has four E-type SEP genes, with SEP3 being an important network hub protein, while SEP1/2/4 are mostly redundant but with some specialised functions[15, 16]. In apple, suppression of E-type SEP1/2-like MADS8/9 modifies floral morphology and significantly reduces hypanthial-derived fruit flesh, but core tissues are mostly unaffected, which suggests flesh and core development are independently regulated[17]. In support of this, partial AG-suppressed apples reported full-sized fruit and thus no effect on hypanthial-derived flesh tissue[9].
Carpel tissues are an important site of integration of fruit initiation cues and in all available seedless fruit the carpel necessarily persists. Seedless fruits typically arise when fruits develop in the absence of fertilisation (parthenocarpy) or from post-fertilisation seed abortion (stenospermocarpy). Many lines of evidence demonstrate the inherent properties of carpel tissues to integrate fruiting signals and trigger fruit development, for instance, incomplete pollen tube growth restricted to the female style can set fruit; presence of additional carpelloid organs or loss of stamens correlates with parthenocarpy; and hormone-mediated pistil growth can last longer than ovule viability[18]. Treatment with phytohormones at flowering is a well-established method for generating seedless fruit. Auxin, gibberellin and cytokinin, alone or combined, have all been linked to parthenocarpic induction[19]. In apple, various combinations of all three hormones, singly or in combination, have been trialled with varying results, but predominantly success is achieved with a combination of gibberellins and cytokinins[20, 21]. Recently, gibberellins alone were sufficient to induce parthenocarpy in ‘Honeycrisp’ apples but it was noted that cytokinin was putatively required for proper ovary development[22]. In all mechanisms to induce fruit without fertilisation, carpel tissues persist to enable integration of fruit initiation cues; in apple, this also means the inedible core persists.
In this study, we aimed to investigate the possibility of creating coreless apples through suppression of C-type AG-like genes and induction of fruit set with hormone treatment. The purpose of the investigation was two-fold. One, to provide a proof-of-concept for a coreless apple from which the pipfruit industry could harness the economic benefits current seedless varieties cannot provide. Two, to address the biological question of whether fruit tissues can develop in the absence of all sexual tissues.
-
Consistent with the ancient genome duplication in apple[23], six apple genes occur within the AG clade. Phylogenetic analysis revealed the two closest apple homologues to AG were MADS15[24] and MADS115, to SHP1/2 were MADS14[24] and MADS114, and to STK were MADS10[25] and MADS110 (Fig. 1c, Supplemental Table S1). To eliminate carpel tissues, the most similar apple gene to AG, MADS15, was used as the basis for RNAi construct design (Supplemental Fig. S1). The MADS box region was included for broader targeting of the AG-like clade, as specific targeting of AG in tomato and apple did not remove carpel tissues[7−9]. Twelve independent RNAi lines containing the MADS15 hairpin construct (ag) were generated. Five showed a range of severity in loss of carpel tissue and an expected increase in repetitive whorls of petals commonly observed in ag mutants (Fig. 1a, b), as well as varying degrees of premature yellow patching on mature leaves (Supplemental Table S2). One line, ag-05, showed an increase in the number of petal whorls, and closer examination revealed remnant ovule- and stamen-like structures in central whorls, consistent with a partial knock-out phenotype (Fig. 1b). Despite the existence of these structures and repeated attempts to pollinate, fruit set was not successful. An extreme line, ag-21, showed no ovule-like tissues, but inner most petals were slightly stamenoid and contained pollen-like granules (Fig. 1b, d) suggesting residual C-function at the centre of the meristem; similarly to line ag-05, repeated pollination attempts were unable to set fruit. The phenotype of both ag-05 and ag-21 surpassed the AG-knock-down phenotype described previously[9] in achieving near or complete removal of carpel tissues, respectively, and an inability to set fruit with pollination.
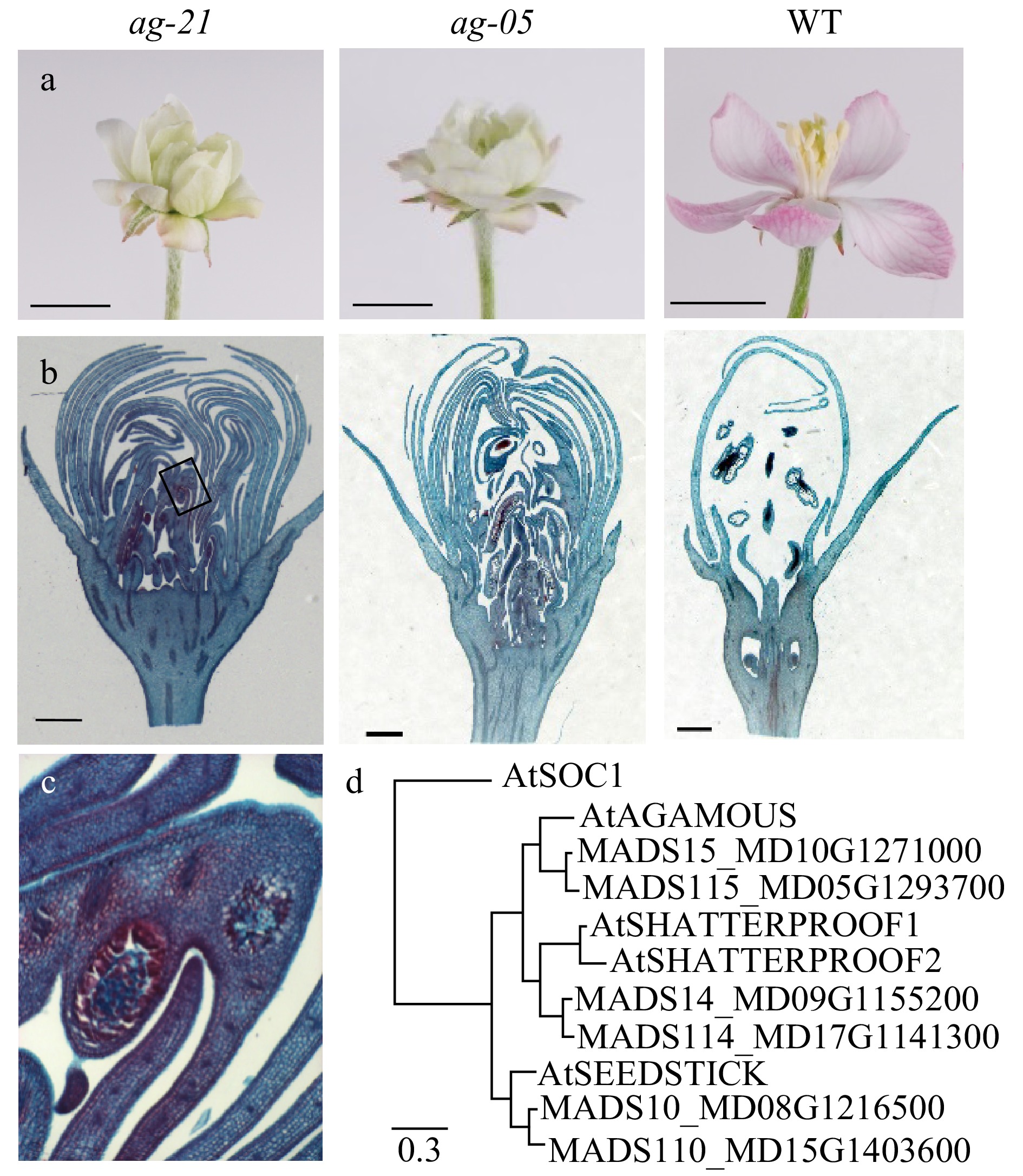
Figure 1. Removing apple carpel tissues through suppression of AG-like genes. (a, b, c) Floral phenotype of wild-type and RNAi lines. (a) Flowers at full bloom (open stage). Scale bar, 1 cm. (b) Longitudinal sections of flowers at balloon stage stained with safranin fast-green. Rectangle on ag-21 image borders tissues with pollen-like granules and demarcates boundary of magnified image in (c). Scale bar, 1 mm. (c) Magnification (10X) of pollen-like granules in ag-21. (d) Phylogenetic tree of AG-like genes in apple and Arabidopsis, model and accession numbers listed in Supplementa Table S1, shown with AtSOC1 (AT2G45660.1) as outgroup. Scale indicates 0.3 substitutions per site.
mRNA-seq analysis was used to assess the effects of RNAi suppression on the AG-like clade and other MADS-box genes. In Arabidopsis, AG is expressed early in flower development to determine carpel and stamen organ identity[26], but is also highly expressed in silique tissues at the beginning of fruit development. However, AG function at this development stage is not known[27]. For the latter, the same is true for AG orthologues in apple[17]. To determine the effects of AG-suppression we chose two late floral stages in which fruit-competent floral tissues are preparing for eventual fruit set: balloon (when the flower is immature and expanding) and open (when the flower is receptive to pollination). It was found that AG-like MADS15/115 were significantly down-regulated in both RNAi lines (p ≤ 0.05; Fig. 2a). AG clade members, SHP1/2-like MADS14/114 and STK-like MADS10/110 were also significantly down-regulated (p ≤ 0.05; Fig. 2a) and were overall more suppressed in the extreme line ag-21, likely accounting for the more extreme phenotype. Transcripts of the AG sister-clade SOC1-like genes, however, were not down-regulated (Supplemental Fig. S2) thus demonstrating constraint of RNAi specificity to the AG clade.
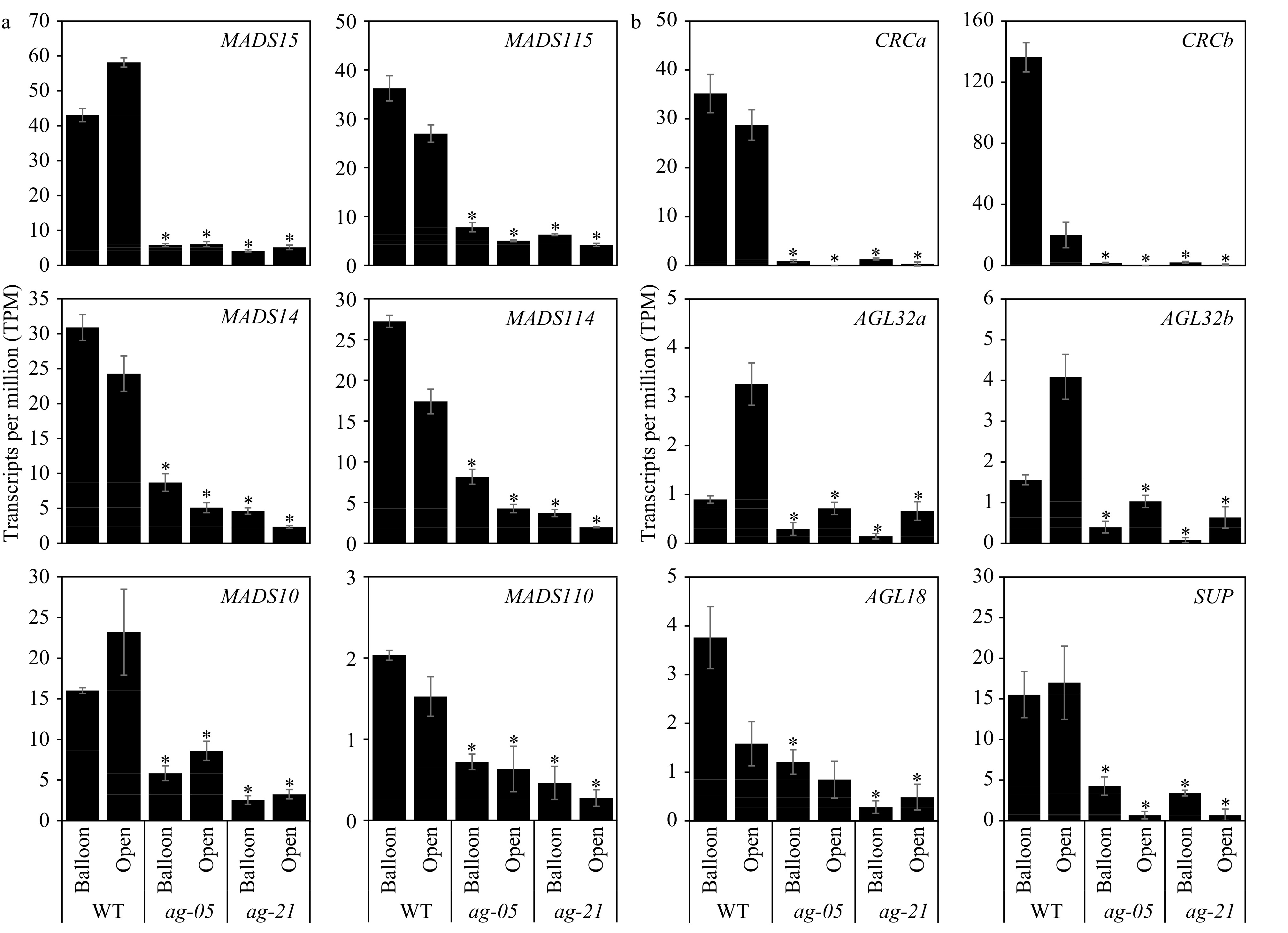
Figure 2. mRNA-seq analysis of apple AG-like gene models (a) and apple homologues to direct targets of AG (b) in wild-type (WT) and RNAi lines ag-05 and ag-21. Gene expression (transcripts per million, TPM) as found in whole flowers at two development stages (balloon and open flower). Values are mean ± SEM (n = 3). * p ≤ 0.05, −1 ≥ Log2FC ≥ 1 compared with WT stage. Apple AG clade accession and model numbers provided in Supplemental Table S1. HFTH1 gene model IDs for apple homologues are: CRCa, HF07613; CRCb, HF42325; AGL32a, HF11534; AGL32b, HF23066; AGL18, HF20484; SUP, HF14066.
Transcriptomic analysis of AG-suppressed lines shows conservation of AG-dependent gene networks
-
The mRNA-seq data were assessed for effects on apple homologues to known direct targets of AG in Arabidopsis. Analysis of the AG-dependent gene regulatory network in Arabidopsis found 225 direct targets of AG[10]. Apple gene models were identified by BLASTp for 177 of the 225 direct AG targets (48 returned low or no identity) resulting in 329 apple homologues consistent with whole genome duplicationin apple. Of these, only 10 (3%) were not expressed at all, 47% (156 genes) were significantly (p ≤ 0.05) differentially expressed with a 2-fold cut-off, and 66% were DE with a 1.5-fold cut-off (p ≤ 0.1). Apple homologues of AG direct targets significantly down-regulated (p ≤ 0.05, Log2FC ≤ −1) included downstream carpel-sub-structure identity genes CRABS CLAW (CRC), AGAMOUS-LIKE 18 (AGL18) and AGAMOUS-LIKE 32/ARABIDOPSIS B-SISTER/TRANSPARENT TESTA 16 (AGL32/ABS/TT16), and the organ boundary gene SUPERMAN (SUP) (Fig. 2b). These effects may be due to the absence of carpel tissues rather than direct regulation, but the trend of down-regulation of these genes mirrors the effects seen in Arabidopsis when AG is suppressed.
Analysis of all significantly differentially expressed genes (DEGs) (p ≤ 0.05 and −1 ≥ Log2FC ≥ 1) was conducted to identify transcriptomic trends related to removal of the carpels from fruit-competent floral tissues. Removal of carpel tissues was incomplete in the partial line (ag-05), and consistent with this, the extreme line (ag-21) exhibited more consistent and greater DEGs overall (Fig. 3a), and set analysis showed a greater number of DEGs in the ag-21-only DEG set (n = 2,051) compared with the ag-05-only DEG set (n = 1,331) (Fig. 3b). Based on homology to TAIR loci, and using keyword search for terms of interest (see Methods), the ag-21-only DEG set had proportionally more DEGs with carpel- and stamen-related terms (Supplemental Fig. S3a), consistent with more complete AG knock-out, while the ag-05-only DEG set had fewer DEGs with gibberellin-related terms, suggesting a more wild-type (WT) state of gibberellin-related transcription (Supplemental Fig. S3b).
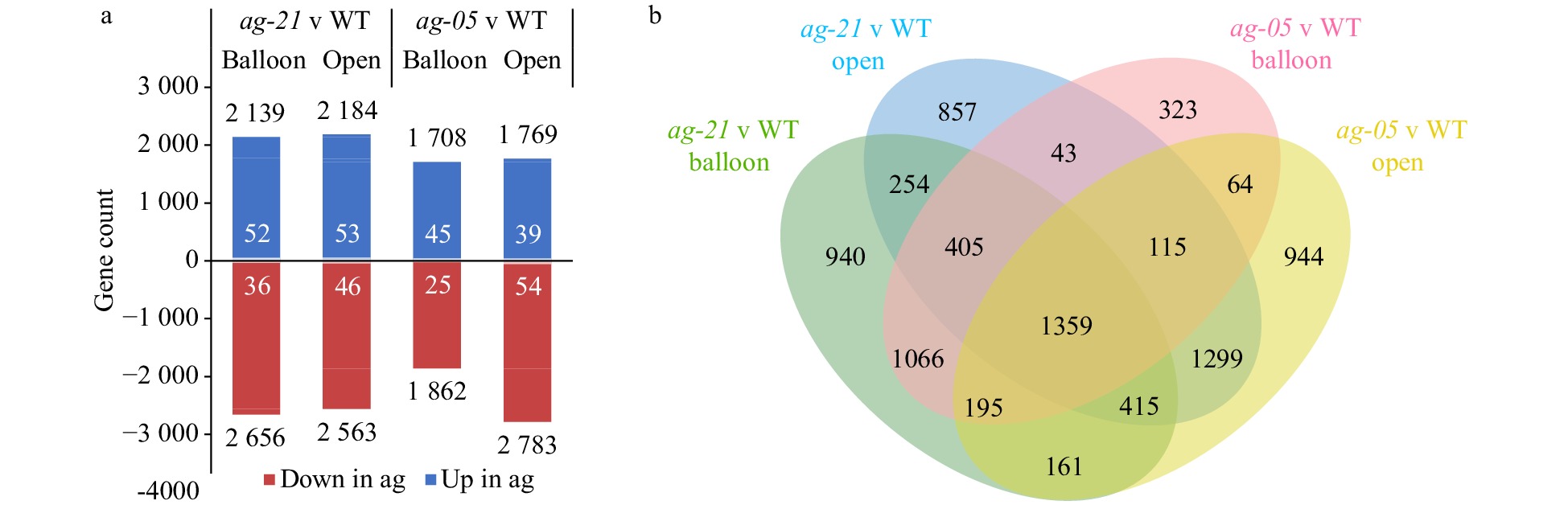
Figure 3. Comparison of transcriptomes of extreme and partial RNAi apple lines. (a) Significantly differentially expressed genes (p ≤ 0.05, −1 ≥ Log2FC ≥ 1, black data labels) in RNAi lines compared with wild-type (WT) at two floral stages (balloon and open flower) and corresponding proportion of AG direct target homologues (white data labels). (b) Venn diagram showing relationships of differentially expressed genes in terms of floral stage and genotype.
Ovary tissues are important sites of integration of fruit initiation signals and many key genetic regulators of fruit initiation have been identified in species with ovary-derived fruits[28]. Apple homologues to these genes were assessed for the effect of carpel removal on expression. Non-orthologous auxin-response factor (ARF) genes, AtARF8 in Arabidopsis[29] and SlARF7 in tomato[30], are negative regulators of fruit initiation. Apple ARFs with expression patterns consistent with negative fruit set regulation include MdARF107 and orthologue to SlARF7, MdARF105[31]. Of all 33 ARF genes in apple, only these two genes were significantly down-regulated in ag-21 open flowers compared with WT (p ≤ 0.05, Log2FC ≤ −1) (Supplemental Fig. S4a, b). Gibberellin-signalling DELLA transcription factors have also been implicated in fruit set regulation in ovary-derived tomato[32]. Apple has six DELLA transcription factors[33] and all were mostly unaffected by carpel removal (Supplemental Fig. S4c).
Reiterating petal whorls seen in RNAi lines were consistent with the known role of AG in controlling floral determinacy, and so mRNA-seq data were assessed for effects on meristem maintenance genes. AG repression of WUSCHEL (WUS) is central to the genetic network regulating floral meristem determinacy[34]. One of two WUS apple homologues (HF19331) was down-regulated in RNAi lines especially at the open flower stage (Supplemental Fig. S5a). Further meristem maintenance genes, SHOOT MERISTEMLESS (STM), POUNDFOOLISH (PNF/BLH8), and KNOTTED-LIKE FROM ARABIDOPSIS THALIANA 6 (KNAT6) were also down-regulated in RNAi lines and again particularly at the open flower stage (Supplemental Fig. S5a). Genes involved in meristem identity such as apple homologues to UNUSUAL FLORAL ORGANS (UFO), CUP-SHAPED COTYLEDON 1 (CUC1), ARABIDOPSIS THALIANA HOMEOBOX GENE 1 (ATH1), APETALA2 (AP2), PROTODERMAL FACTOR 2 (PDF2), and REVOLUTA (REV) were conversely all found to be predominantly up-regulated in RNAi lines (Supplemental Fig. S5b). Interestingly this trend is contrary to that observed in Arabidopsis ag mutants at similar late floral stages[10].
Hormone-induced fruit set of ag lines yields coreless apples
-
To test whether fruit development could be triggered in the absence of carpel tissues, hormone-induced fruit initiation was used in place of pollination which had failed to produce fruit in the extreme RNAi lines. In apple, hormone treatments to induce fruit set vary widely between cultivars in terms of timing and number and type of treatments[20, 21]. In this study, different regimes were trialled over three years (Table 1). In the first year, auxin and gibberellin applied separately did not yield fruit. In the second year, treatment with gibberellin A4 (GA4) and indole-3-acetic acid (IAA) was sufficient to induce fruit set in 4.5% of cases in control lines and yielded a single fruit in line ag-05. Treatment with GA4, IAA and 6-benzylaminopurine (6-BAP) induced fruit set in 6.1% of cases in control lines, and produced one fruit in the extreme line ag-21. In year three, to increase incidence, all flowers were treated with the same treatment of GA4, IAA, and 6-BAP with more regularity and for a longer duration, which yielded 63 fruit in partial line ag-05 and 5 fruit in extreme line ag-21.
Table 1. Hormone treatments to induce pathenocarpy.
Year Conditions of hormone treatment Line No. of flowers treated No. of mature fruit 1 Single 1.5 mM GA4 from -7 to -4 DAFB (no auxin treatment) ag-05 65 0 Single 1.5 mM GA4 from -7 to -4 DAFB with daily 5.7 µM IAA from 0 to 1 DAFB, until appearance of yellowing ag-05 55 0 Daily 5.7 µM IAA from 0 to 1 DAFB, until appearance of yellowing ag-05 50 0 2 602 µM GA4, 5.7 µM IAA, beginning -7 DAFB, repeated every 3−4 days for 2 weeks WT 110 5 ag-05 110 1 ag-21 40 0 602 µM GA4, 5.7 µM IAA, 444 µM 6-BAP, beginning -7 DAFB, repeated every 3−4 days for 2 weeks WT 115 7 ag-05 115 0 ag-21 45 1 No Hormone control, beginning -7 DAFB, repeated every 3−4 days for 2 weeks WT 55 0 ag-05 65 0 ag-21 40 0 3 602 µM GA4, 5.7 µM IAA, 444 µM 6-BAP, beginning -7 DAFB, repeated every 2 days for 3 weeks, then every 3 days for 1 week. ag-05 193 63 ag-21 123 5 Fruit by hormone induction in control lines retained locular cavities with remnant integument-derived seed coats present in seed locules (Fig. 4a). In year two, fruit from partial line ag-05 had a reduced core located more distally with double the normal locule number. Fruit from extreme line ag-21 had no core tissues (Fig. 4a). Vascular traces in the RNAi lines were similar to controls despite reduction or removal of locular cavities (Fig. 4a). In year three, in order to increase the incidence of fruit initiation, hormones were applied more regularly and over a longer period. In this year, fruit were more irregularly-shaped owing in part to development of fruit tissues downwards along the pedicel beyond the hypanthium. In longitudinally sectioned fruit, more fibrous pedicel-like tissues were visible connecting with central vascular bundles (Fig. 4b). Occasionally, a hole formed in mature fruit in the region where pedicel tissues separated from fruit vascular tissues (Fig. 4b, arrow). Calculation of flesh area in ag lines was in addition found to be not significantly different from that of pollinated wild-type controls (Student’s t-test, p ≤ 0.05; Fig. 4c).
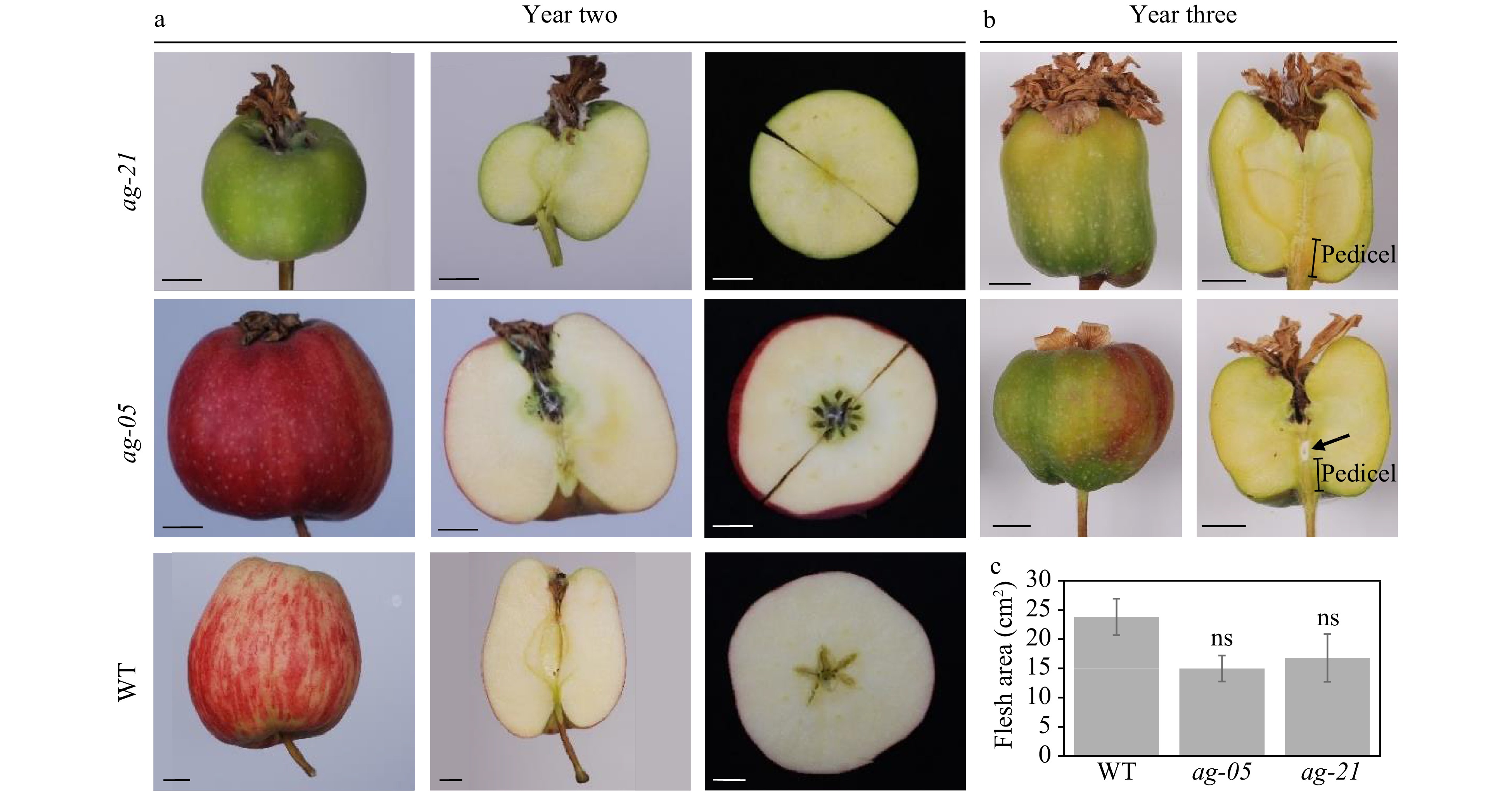
Figure 4. Mature apples (approx. 120 DAFB) produced by hormone-induction in wild-type and RNAi lines in (a) year two and (b) year three. Classic parthenocarpic fruit phenotype in wild-type, compared with coreless phenotype in ag-21. Scale bar, 1 cm. (c) Comparison of flesh area in wild-type and RNAi lines, n = 5 per genotype; Student’s t-test, p ≤ 0.05.
Coreless apples undergo fruit ripening
-
For coreless apples to be acceptable to the consumer it is essential that they undergo fruit ripening, therefore coreless apples were assessed for both competence and degree of ripening. Apple is a climacteric fruit, and so many ripening events respond at varying sensitivities and dependencies to endogenous auto-catalytic ethylene[35]. Compared with pollinated controls, coreless apples produced a similar amount of ethylene at harvest and during storage as the fruit matured (Fig. 5). The early ripening events such as starch clearance were somewhat more advanced in the RNAi lines than in the wild-type, with near-complete clearance at-harvest. The background skin colour changed from green to yellow but was slightly delayed compared with that in the wild type, and finally both RNAi lines underwent softening (Fig. 5). While these results show that coreless apples were able to undergo a ripening processes a more detailed analysis with larger numbers of fruit are needed to verify the earlier ripening phenotype.
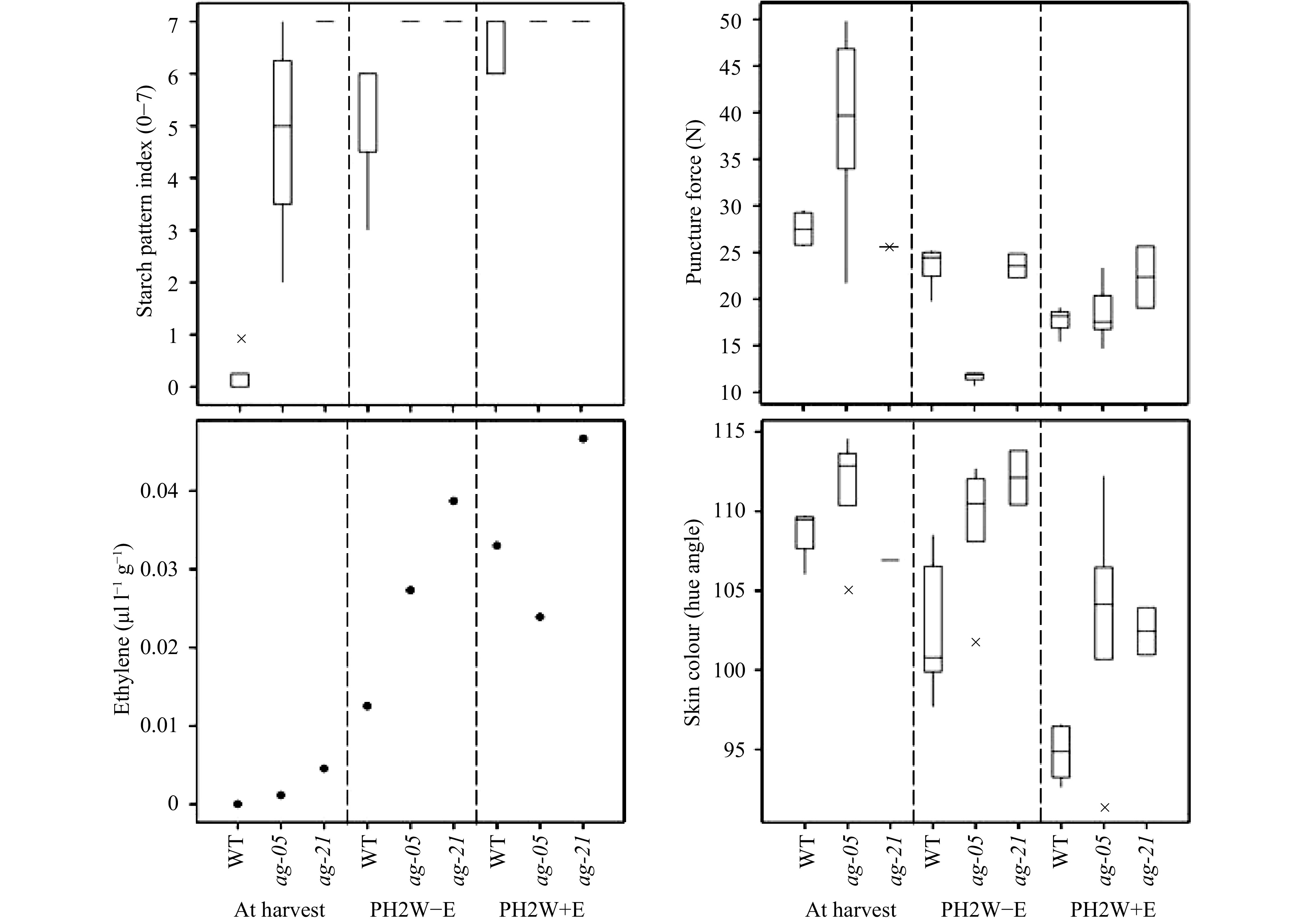
Figure 5. Fruit ripening in mature pollinated ‘Royal Gala’ (wild-type; WT) (n = 5 fruit), and RNAi lines ag-05 (n = 5 fruit) and ag-21 (n = 1−2 fruit) apples. Starch content on a scale of 1 (high starch) to 8 (no starch), flesh firmness measured by puncture force, emission of ethylene using head space analysis, background skin colour measured by hue angle from green (115) to yellow (95) as observed during ripening. PH2W−E are postharvest fruit ripened for 2 weeks at room temperature in the absence of ethylene, PH2W+E are postharvest fruit ripened for 2 weeks in the presence of 100 ppm ethylene.
-
This study demonstrates the feasibility of generating a coreless pipfruit through the suppression of AG-like genes and hormone-induced fruit development. Given that all six genes in the AG clade were suppressed in the RNAi lines (more so in the extreme line), there is the possibility that all genes need to be suppressed for complete removal of carpel tissues. This is consistent with previous work in tomato[8] and apple[9] where there appears to be partial redundancy in function within the AG clade. Additionally, SHP-driven AG-independent carpel-development pathways exist in Arabidopsis and SHP and STK are expressed in carpel and carpel sub-structures[12, 13]. In apple ag lines it was therefore unknown whether suppression of AG clade genes was due to direct RNAi regulation or from absence of carpel tissues; especially given there is a direct regulation of SHP1/2 by AG[10, 13].
The coreless apple phenotype in line ag-21 is consistent with core identity and development being determined by AG-like clade. Previously, it was shown that apple flesh identity, development and ripening are predominantly directed by E-type SEP1/2-like MADS8/9[17]. The coreless phenotype is evidence that C-type AG-like and E-type SEP1/2-like genes act independently in their facilitation of fruit tissue development. In cherry tomato, elevated expression of TOMATO AGAMOUS1 (TAG1) causes homeotically-normal, mature sepals to undergo ripening, especially when cultured at low temperatures[36], suggesting a role for AG in fleshy fruit development in ovary-derived fruit. In apples, suppression of AG did not remove fleshy fruit characters, which shows AG is not required for flesh identity in apple. Therefore, in tomato, ectopic expression which resulted in fleshy sepals was probably due to ectopic carpel identity later defined by other genes to be flesh competent.
Decoupling of flesh and core development in ag lines demonstrates inherent flesh-competency of the hypanthium. This observation aligns with the flesh competency model proposed by Liu, Ma[37]. The apple hypanthium has all three properties the authors propose are needed for flesh competency: one, co-existence with photosynthetic capability; two, ability to promote expression of fruit-related MADS-box genes i.e. MdMADS8/9[17]; and three, potential to inactivate auxin/GA signalling repressors, demonstrated here by fruit set with hormone treatment and also suggested by non-elimination of auxin/GA signalling repressors with carpel removal. Typically the sepal is exemplified as a non-competent organ, and strong support exists for the hypanthium being predominantly sepal-derived[38]. While AG-like MADS15 is not needed for flesh identity, it is expressed in the hypanthium[17, 24]. Its role here may be to promote auxin/GA signalling, a role circumvented by exogenous hormones. In accordance with this, direct targets of AG include auxin, GA and cytokinin signalling pathway components[10].
Different hormone treatment regimes were trialled over the course of this study, but full development of coreless fruit appeared to require all three fruit-set hormones. Treatments of single hormones in the first year failed, including a staged approach of GA then IAA. Comparison of the need for cytokinin in year two produced inconclusive results in ag lines; however, controls had a small increase in yield when cytokinin was included. Treatments with single hormones have had success in other species, such as auxin in citrus and strawberry, gibberellin in grapes, rose and apples, and cytokinin in watermelon, pear, and kiwifruit[19, 22]. Apple cultivars have varying requirements[20, 21] but usually see improved yields with a combination of all three hormones, and indeed the underdevelopment of ovary tissues in gibberellin-induced ‘Honeycrisp’ apples was putatively attributed to a lack of cytokinins[22]. In this study, all three hormones together produced coreless fruit. Of note was the need for repeated treatments, when in practice, in the field, fruit set is usually triggered after a single spray[20, 22]. The extended treatment regime in year three also altered fruit shape compared with that in year two, highlighting the need to find a minimal optimal dose.
It is hypothesised that seed auxin concentrations regulate fruit development, and the onset of seed dormancy depletes fruit auxins, thereby enabling ripening progression[31, 39]. Seedless fruits would therefore be expected to undergo an altered ripening progression compared with seeded fruit. This is indeed the case in avocado[40] and grape[41] which both observe early ripening in seedless or low-seeded varieties. The same appears to occur with coreless apples, providing an opportunity for earlier harvests.
-
Fleshy fruit formation and ripening is therefore inherent to the apple hypanthium and is possible in the absence of carpel tissues. The lack of integration of fruit initiation signals normally orchestrated by the carpel can be overcome with exogenous treatment of fruit initiation phytohormones, but ripening signals are able to proceed in the absence of carpel tissue and without exogenous treatments of any kind. This study provides a proof of concept for fruit formation in the absence of carpels and sexual reproduction, thus bypassing the need for pollination and fertilisation altogether.
-
Apple genes homologous to Arabidopsis AGAMOUS (AG) were identified using BLASTp search against an apple EST collection[42]. The apple MADS-box gene MADS15[24] was found to be most similar to Arabidopsis AG and was later verified using whole genome data[23]. An RNAi hairpin construct was created in the binary vector pSAK277[43] with the 35S promoter driving an inverted repeat of the first 430 bp of MADS15. Twelve independent lines of transgenic Malus × domestica ‘Royal Gala’ were generated using Agrobacterium-mediated transformation[44]. Plants were grown in contained glasshouse conditions. Flowers were scored for homeotic changes and harvested for microscopy.
Microscopy
-
Plant tissue samples were fixed in FAA (4% formalin/50% alcohol/5% acetic acid in water to 100%). Tissues were dehydrated with an ethanol series (50%, 70%, 90%, and x3 100%), cleared with xylene/ethanol (1:1) followed by two changes of xylene (all steps 3 h) followed by three changes of molten wax held at 58−60 °C 12 h per step. All steps were carried out using a Leica TP1020 tissue processor (Leica Microscopy Systems Ltd, Heerbrugg, Switzerland) using both agitation and vacuum cycling. The tissue was embedded in wax blocks using a Leica EG1160 wax embedding station (Leica Microscopy Systems Ltd, Heerbrugg, Switzerland) using Paraplast wax (Oxford Labware, St Louis, MO). Material was sectioned at 15 µm using a Leitz 1512 rotary microtome (Ernst Leitz, Weitzer, Germany) mounted on glass slides and allowed to dry at 42 °C. Sections were dewaxed with two changes of xylene followed by two changes of absolute ethanol (each 5 min) and then stained using the safranin fast green staining method[45]. Sections were observed and imaged using either an Olympus Vanox AHBT3 (Olympus Optical Co. Ltd, Tokyo, Japan) equipped with a CoolSnap digital camera (Roper Scientific Ltd, Tucson, Arizona) or a Leica FLZIII (Leica Microscopy Systems Ltd, Heerbrugg, Switzerland) fitted with an Infinity2-3C digital camera (Lumenera Corporation, Ottawa, ON, Canada).
Hormone-induced fruit set and measurement of ripening
-
In year one, between -7 and -4 days after full bloom (DAFB) flowers of a single line (ag-05) received a single treatment of 1.5 mM gibberellin A4 (GA4) by dipping, followed by no or multiple applications of 5.7 µM indole-3-acetic acid (IAA) by painting onto the hypanthium. In year two, starting at -7 DAFB, hypanthia of whole flowers of three extreme (ag-05, ag-06, ag-21), one intermediate (ag-11), and two control lines were painted with either 602 µM GA4 and 5.7 µM IAA, 602 µM GA4, 5.7 µM IAA, and 444 µM 6-benzylaminopurine (6-BAP) or blank solution every 3–4 days for two weeks. In year three, flowers of lines ag-05 and ag-21 were treated with 602 µM GA4, 5.7 µM IAA, and 444 µM 6-BAP every 2 days for 3 weeks then 3 days for 1 week over the period from -7 to 21 DAFB (Table 1). Control lines in year two were ag lines with wild-type leaf, floral and fruit phenotype and were bagged during hormone treatments. Mature apples were harvested and ripened with 100 μl L–1 ethylene[17]. Fruit were assessed for weight, background skin colour, firmness, and starch clearance as per Johnston et al[35]. For headspace ethylene, fruit were held for 1 h in 2 L headspace tubs, fruit treated with ethylene were aerated for 1 h before being placed in the tubs. A 1 ml headspace sample was extracted and analysed for ethylene using gas chromatography as per Johnston et al.[35]. Flesh area was calculated in ImageJ using five fruit per genotype.
Phylogenetic analyses
-
Analysis was performed using Geneious 10.0.3 (Biomatters, www.geneious.com). Multiple sequence Geneious alignment using Blosum62 cost matrix was used to align apple protein models based on Malus × domestica ‘HFTH1’ genome[46] (Genome Database for Rosaceae, www.rosaceae.org), the putative hybrid sequence of apple MxMADS15 (Supplemental Table S1) and Arabidopsis protein sequences (TAIR, www.arabidopsis.org). A conserved 240-residue region was extracted from the alignment and used to construct a PHYML phylogenetic tree[47] and bootstrap analysis of 100 datasets.
mRNA sequencing and analysis
-
Three biological replicates from whole flowers at two stages of flower development (unopened balloon and open flower) from untransformed Malus × domestica ‘Royal Gala’ and two RNAi lines showing partial and extreme ag-like phenotypes (ag-05 and ag-21) were harvested and preserved in liquid N2. RNA was isolated by rapid CTAB extraction[48]. Five µg total RNA was used to construct strand-specific poly-A RNA libraries following Zhong, Joung[49]. Libraries were sequenced by Macrogen using HiSeq2000 sequencing system (Illumina) to produce reads per library ranging from 20–27 million. Reads were processed using FastQC and Trimmomatic, aligned independently with STAR[50] to Malus × domestica ‘Hanfu’ Triple Haploid (HFTH1)[46] and Malus × domestica ‘Golden Delicious’ Double Haploid (GDDH13)[51] genomes and counted with HTSeq. Two apple genomes were used owing to differences in quality of gene model predictions. The statistical software R[52] was used to calculate transcripts per million (TPM) and perform statistical analyses.
Homology analysis was performed with a publicly available BLASTp analysis of HFTH1 to TAIR peptides (Genome Database for Rosaceae, www.rosaceae.org). Keyword analysis was performed using combined lists of TAIR loci with flowering-related GO Biological Processes, GO molecular function, Growth & Development Stages, and Plant Anatomical Entity (Plant Structure) in conjunction with the HFTH1-TAIR BLASTp results.
-
Malus AG clade accession numbers and gene model numbers are provided in Supplemental Table S1.
The RNA-seq data can be found in: NCBI project PRJNA357936
- The authors would like to thank the glasshouse staff, Monica Dragulescu and Gnanaseela Wadasinghe, for maintaining the apple trees. Prof. Richard Newcomb and Dr Zac Hanley for comments on the manuscript. The New Zealand Ministry for Business Innovation and Employment programme ‘Pipfruit: a juicy future’ (C06X0705) and the New Zealand Ministry for Primary Industries CORE funding supported this work.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Deconvolution of apple AGAMOUS clade accession numbers. Arabidopsis thaliana genes AGAMOUS (AG), SHATTERPROOF1/2 ,(SHP) and SEEDSTICK (STK) have two homologous genes in apple consistent with its ancient whole genome duplication.
- Supplemental Table S2 Summary of phenotypes of RNAi lines. Twelve independent RNAi lines were generated in Malus × domestica ‘Royal Gala’ background; five of these lines bore the multiple petal whorl phenotype characteristic of AGAMOUS loss of function mutants.
- Supplemental Fig. S1 MADS15 RNAi construct details. The insert was amplified with AttR1 and AttR2 adapted primers and inserted into pTKO2 plasmid.
- Supplemental Fig. S2 mRNA-seq analysis of apple SOC1-like gene models in wild-type (WT) and RNAi lines ag-05 and ag-21. Gene expression (transcripts per million, TPM) as found in whole flowers at two development stages (balloon and open flower). Values are mean ± SEM (n = 3). * p ≤ 0.05, −1 ≥ Log2FC ≥ 1 compared with WT stage.
- Supplemental Fig. S3 Comparison of transcriptomes of extreme and partial apple RNAi lines. ( a ) proportion of floral related annotations in four genes sets. ( b ) Proportion of hormone related annotations in four gene sets.
- Supplemental Fig. S4 mRNA-seq analysis of apple gene models to fruit initiation genes in wild-type (WT) and RNAi lines ag-05 and ag-21. (a) Out of 33 auxin-response factors (ARF), the only two significantly down-regulated (p ≤ 0.05, Log2FC ≤ −1) genes in ag-21 open flowers, compared with WT, were two ARFs with expression known to correlate with negative fruit set regulation (Devoghalaere et al. 2012). (b) MdARF105 and MdARF107 are suppressed in carpel-free ag-21 but not totally eliminated. (c) Gibberellin-signalling DELLA transcription factors. Gene expression (transcripts per million, TPM) as found in whole flowers at two development stages (balloon and open flower). Values are mean ± SEM (n = 3). * p ≤ 0.05, −1 ≥ Log2FC ≥ 1 compared with WT stage.
- Supplemental Fig. S5 mRNA-seq analysis of apple homologous gene models to Arabidopsis meristem identity and maintenance genes in wild-type (WT) and RNAi lines ag-05 and ag-21. Genes predominantly down-regulated (a) and up-regulated (b) in RNAi lines. Gene expression (transcripts per million, TPM) as found in whole flowers at two development stages (balloon and open flower). Values are mean ± SEM (n = 3). * p ≤ 0.05, −1 ≥ Log2FC ≥ 1 compared with WT stage.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ireland HS, Tomes S, Hallett IC, Karunairetnam S, David KM, et al. 2021. Coreless apples generated by the suppression of carpel genes and hormone-induced fruit set. Fruit Research 1: 2 doi: 10.48130/FruRes-2021-0002

Coreless apples generated by the suppression of carpel genes and hormone-induced fruit set
- Received: 19 February 2021
- Accepted: 27 April 2021
- Published online: 13 May 2021
Abstract: Seedless fruits have high consumer appeal and have made seeded varieties obsolete in some crops. In seedless apple varieties, core tissues which normally contain the seed can be unpalatable, reducing the seedless appeal. Apples are accessory fruit with edible flesh derived from hypanthial tissue – a floral tube fused to a compound ovary. Here we show that through suppression of AGAMOUS-like carpel identity genes and hormone induced fruit set, it is possible to generate coreless and therefore seedless apples. Suppression of AGAMOUS-like genes increased petal whorls and fully eliminated carpel development. Treatments with a combination of gibberellin, cytokinin and auxin, rather than single treatments, were required for fruit initiation in these lines. Transcriptomic analysis of agamous RNAi lines suggested conservation of AGAMOUS-dependent gene networks between apple and Arabidopsis. In the absence of all sexual tissues, the developing fruit continues to grow and follow a ripening process similar to that of a regular apple. The coreless phenotype offers a new concept for pipfruit consumers improving convenience and reducing food waste.
-
Key words:
- Apple /
- Malus Domestica /
- fruit development /
- Agamous