









-
Plant cells are wrapped within the cell wall, which can be altered to allow the cells to expand during growth, once the cells reach their final size and shape, many specialized cells produce thicker secondary cell walls (SCWs). Secondary walls are usually composed of cellulose (40−80%), hemicellulose (10−40%), lignin (5−25%), and cell wall protein[1]. Cellulose, as the most typical substance in plant cell walls, consists of a collection of glucan chains interacting with each other via hydrogen bonds. Lignin is the second largest biopolymer in vascular plants and is constructed primarily from three kinds of 4-hydroxyphenyl-propanoids known as monolignols. Both cellulose and lignin provide mechanical protection for cells and plant stress resistance. However, previous studies have not explored the molecular regulatory network involved in the accumulation of cellulose and lignin in apple.
Cellulose is produced by the cellulose synthase complex on the plasma membrane, in contrast, lignin is synthesized by the common phenylpropane pathway and the specific pathway from feruloyl-CoA to monolignin units[2], which can be produced by a three-level regulatory network[3−4]. In Arabidopsis, it is shown that the coordinated expression of secondary wall formation related genes can be organized by a group of NAC (NAM, No apical meristem; ATAF, Arabidopsis transcriptional activation factor and CUC, Cup-shaped cotyledon) master switches, which involve SND1 (secondary wall-related NAC domain protein 1), NST1/2 (NAC secondary wall thickening promoting factor 1/2) and VND6/7 (vascular-related NAC-DOMAIN6/7)[5−9]. There are a high number of MYB gene family members which have various effects on SCW biosynthesis[10−11]. AtMYB46, the master switch of SCW biosynthesis in Arabidopsis thaliana, transcriptionally activates downstream target transcription factors by directly binding to cis-elements in their promoters, of which AtC3H14, MYB52 and MYB63 can continue to make downstream SCW formation related genes active[12]. Being functionally redundant with MYB46, MYB transcription factor, MYB83 is also a target gene for SND1[13]. Other MYB regulatory factors also have the function of transcriptional activation in SCW formation, with the exception of MYB46 and MYB83. OsMYB58 and OsMYB63, the Arabidopsis homologs in rice, directly activate the cellulose synthase gene OsCesA[14]. AtMYB103, the direct target of NST1/2, SND1 and MYB46, is a regulatory factor that is fundamental for S-lignin biosynthesis in the inflorescence stem of A. thaliana, and can also directly bind to the promoter of the cellulose synthase gene and regulate the transcription of CesA8[15−16]. In apple, MdMYB88, MdMYB124, MdMYB46 and SND1 were recently identified and shown to arrange the formation of SCW[17−19]. The regulatory network for SCW formation in fruit trees still needs to be further explored in comparison to model plants.
MYB5, an R2R3 MYB transcription factor, collaborates with the WD40 repeat protein TTG1 (TRANSPARENT TESTA GLABRA1) in seed coat development[20]. Overexpression of AtMYB5 will bring about the formation of more small trichomes in Arabidopsis[20], along with a reduction in the number of branching of plants and leaf trichomes. Meanwhile, MtMYB5 also functions in organizing the seed coat compound biosynthesis mediation in Medicago truncatula[21]. Constitutive expression of MtMYB5 induces more proanthocyanidin stored in M. truncatula hairy roots, in contrast, inhibiting its expression causes the color of the seed coat to become darker and mucilage biosynthesis to be partially blocked. Overexpression of VvMYB5a/b from grape (Vitis vinifera) in tobacco (Nicotiana tabacum) induced proanthocyanidins (PA) accumulation in flowers[22−23], and it was recently discovered that the MYB5-driven MYB-bHLH-WD (MBW) transcription factor complex could recruit VvWRKY26 to improve the expression of target genes involved in vacuolar acidification, flavonoid compound synthesis and transport[24].
In apples, we previously explored the transcriptional regulation network of SCW formation involving MdMYB46 and MdSND1[18−19], and found that the expression level of MdMYB5 showed a strong correlation with the expression levels of MdSND1 and MdMYB46 (Supplemental Fig. 1), which implies that the process of regulating SCW biosynthesis in apples was possibly influenced by MdMYB5. Therefore, we cloned MdMYB5 in apple and studied its function. We observed that MdMYB5, directly activated by MdSND1 and MdMYB46, could initiate SCW formation by promoting the biosynthesis of lignin and cellulose in apple.
-
The MYB transcription factor gene MdMYB5 (numbered MD11G1006500 in the Apple Genome (GDDH13 V1.1) database or LOC103429782 in National Center for Biotechnology Information) was cloned from the GL-3 apple. The result of phylogenetic analysis and multiple sequence alignments suggested that MdMYB5 was an R2R3-type transcription factor and similar to RcMYB5 and VvMYB5b (Fig. 1a, b).
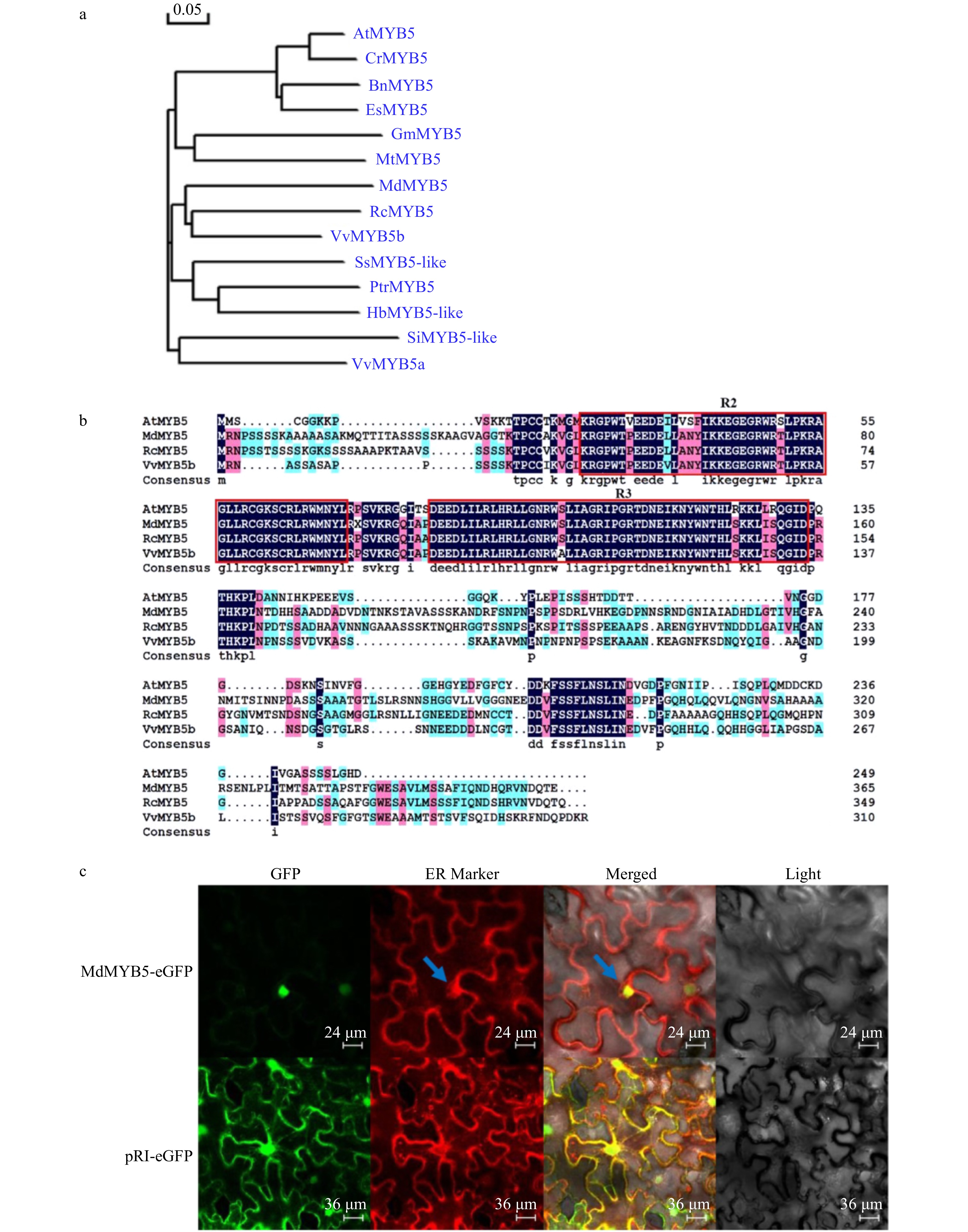
Figure 1. Isolation and characterization of MdMYB5 from apple. (a) Phylogenetic analysis of MdMYB5 compared with AtMYB5 orthologs. Si (Sesamum indicum); Vv (Vitis vinifera); Es (Eutrema salsugineum); Bn (Brassica napus); Cr (Capsella rubella); Rc (Rosa chinensis); Md (Malus domestica); Hb (Hevea brasiliensis); Ptr (Populus trichocarpa); Ss (Syzygium samarangense); Mt (Medicago truncatula); Gm (Glycine max); At (Arabidopsis thaliana). (b) The conserved R2R3 domain in MdMYB5, AtMYB5, RcMYB5 and VvMYB5b. (c) Subcellular localization of MdMYB5-eGFP in the epidermal cells of tobacco (the blue arrow indicates the location of the endoplasmic reticulum).
The endoplasmic reticulum (ER) is an interstitial laminar or small tubular system composed of plastid membranes, connected to mitochondria, cell membranes and nuclear membranes. Given that ER is usually located around the nucleus in the cytoplasm, we used ER markers to indirectly identify the position of the nucleus. It can be seen that the red fluorescence located in ER is wrapped around the green fluorescence of MdMYB5-eGFP (shown by the arrow), and the green fluorescence with high intensity is spherical (Fig. 1c). This revealed that MdMYB5 was functioning in the nucleus.
MdMYB5 regulates the formation of secondary cell wall and may correlate with the resistance to salt and osmotic stress in apple.
-
The studies performed showed that the expression level of MdMYB5 was significantly up-regulated in MdSND1 and MdMYB46 overexpressing apple tissue cultures while substantially down-regulated in the MdSND1-RNAi and MdMYB46-RNAi plants (Supplemental Fig. 1). Due to the vital role of MdSND1 and MdMYB46 in the formation of SCW, MdMYB5 may have effects on forming SCW, implied by the positive correlation of the expression level.
We constructed the MdMYB5-RNAi vector (Fig. 2a), transformed 'GL-3' by Agrobacterium-mediated method, and screened 3 RNAi transgenic apple lines, MdMYB5-RNAi-1, MdMYB5-RNAi-2 and MdMYB5-RNAi-3. Quantitative analysis revealed that the expression levels of MdMYB5 in MdMYB5-RNAi lines were significantly down-regulated (Fig. 2b). Additionally, lignin and cellulose content were significantly down-regulated in the MdMYB5-RNAi lines (Fig. 2c, d). Cross-section observation of the stems showed that cells with thickened SCW in the red frame of GL-3 sections were more densely distributed, in contrast, cells with thickened secondary walls were less and more scattered in MdMYB5-RNAi sections suggesting that MdMYB5 transcription level influenced the formation of SCW (Fig. 2e, f). To further demonstrate the role of MdMYB5, we also examined the transcription levels of lignin biosynthesis related genes (Md4CL and MdCOMT) and cellulose related genes (MdCESA4 and MdCESA8) in MdMYB5-RNAi lines, and found that these genes were also significantly down-regulated in MdMYB5-RNAi lines (Fig. 3a−d), implying that the expression of these genes was also limited in their interference with the transcription of MdMYB5. It is noticeable that the expression levels of genes encoding the key transcription factors MdSND1 and MdMYB46 taking part in the regulatory network of SCW formation did not change significantly in MdMYB5-RNAi lines (Fig. 3e, f), suggesting that MdMYB5 possibly plays a part downstream of MdSND1 and MdMYB46. Meanwhile, the leaves of these MdMYB5-RNAi lines appeared noticeably dark brown in comparison with the wild-type plants under salt and osmotic stress, implying that the RNAi plants were likely more affected by the stress conditions (Fig. 4).
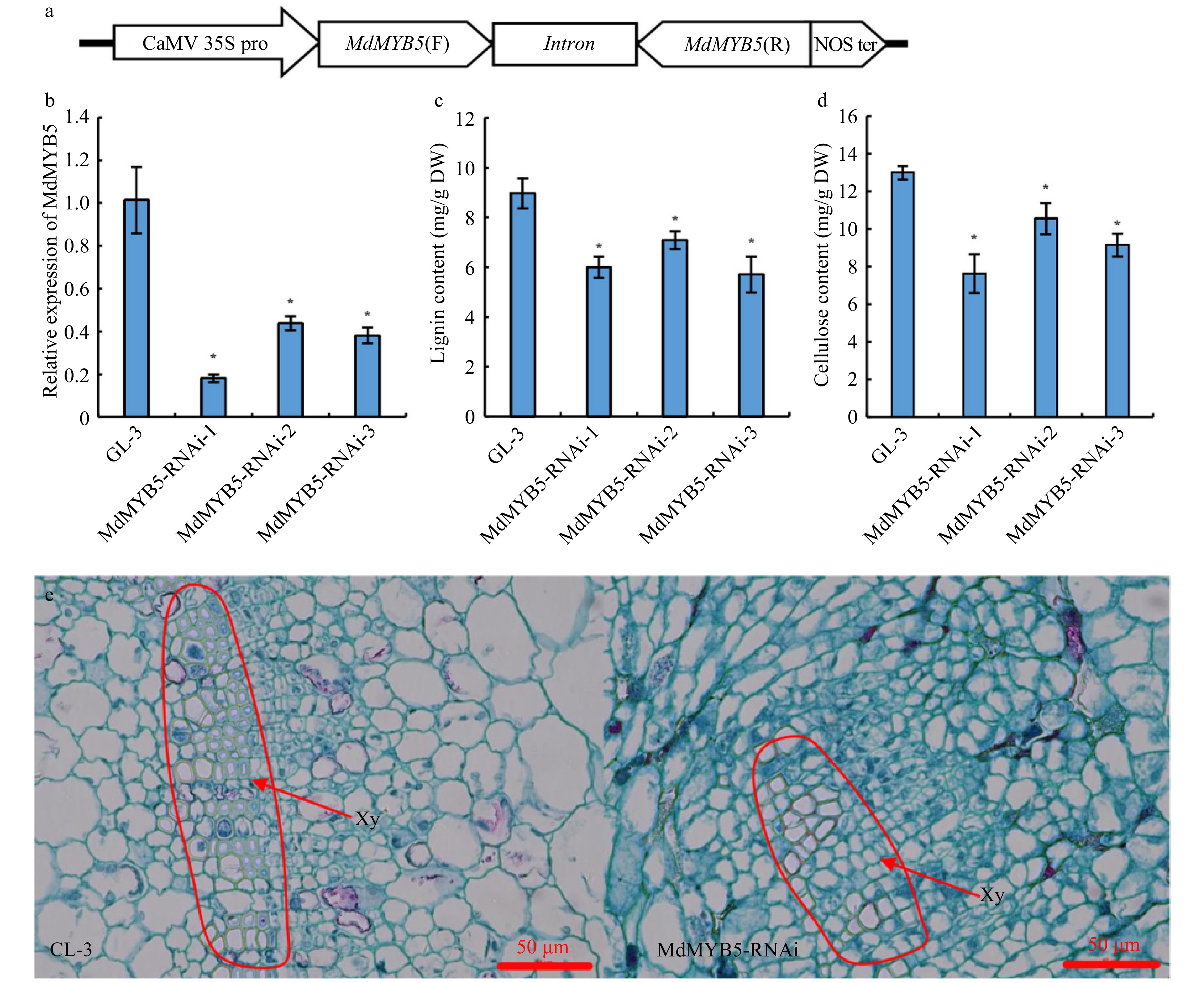
Figure 2. The formation of secondary cell wall was 'blocked' in MdMYB5-RNAi apple tissue cultures. (a) Schematic showing the RNAi vector for MdMYB5 gene. (b) Transcriptional levels of MdMYB5 in RNAi apple tissue cultures. The error bars indicate the standard deviation (SD, n = 3). * Indicates significant differences between the MdMYB5-RNAi apple tissue cultures and GL-3 (p < 0.05, based on t-test). (c) Lignin content in MdMYB5-RNAi apple tissue cultures. (d) Cellulose content in MdMYB5-RNAi apple tissue cultures. (e) The observation of transverse sections by microscopy, which were taken from the basal stems of GL-3 and MdMYB5-RNAi apple tissue cultures. The red arrow indicates the thickened secondary cell wall. Xy = xylem.
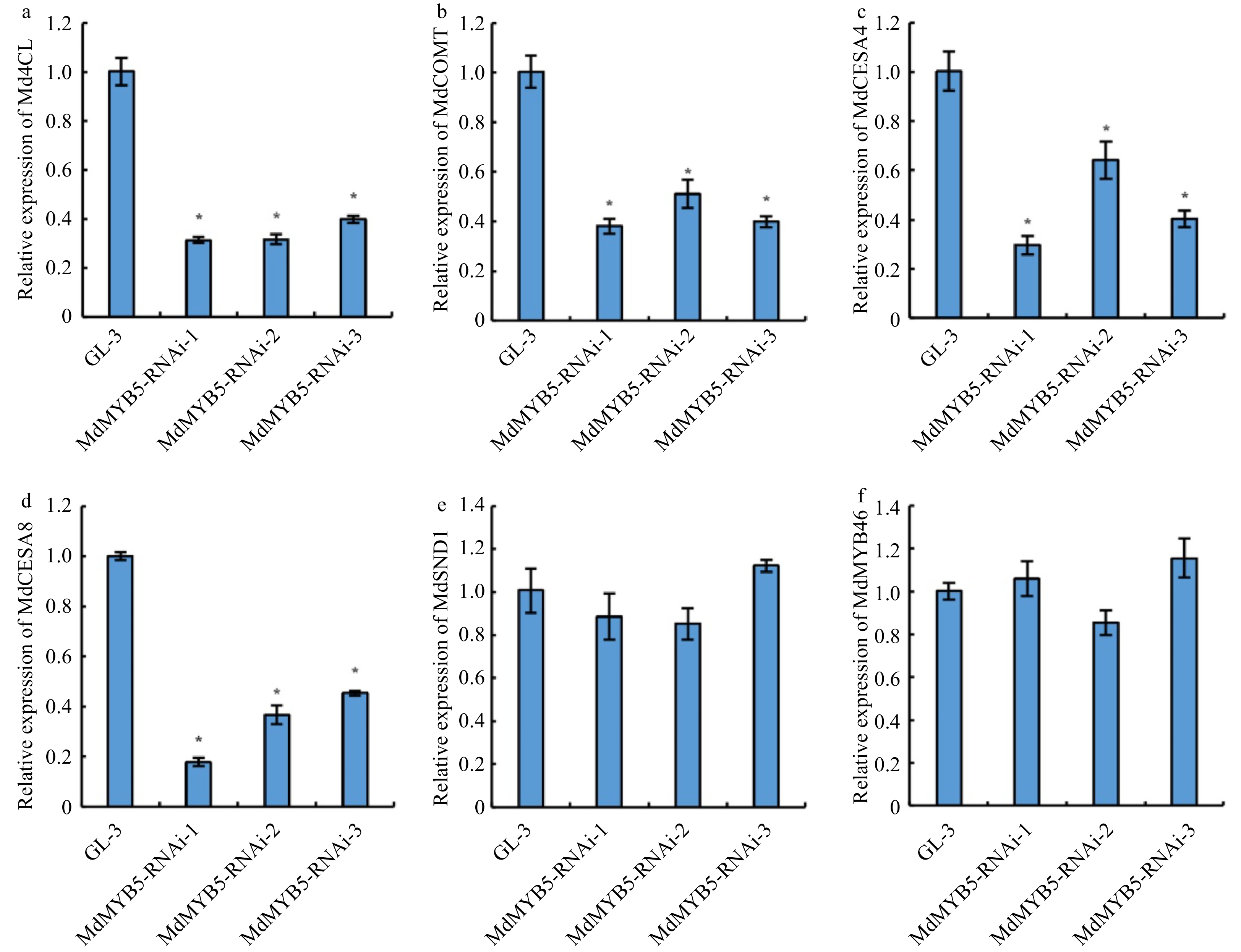
Figure 3. Transcriptional levels of lignin and cellulose biosynthesis related genes in MdMYB5-RNAi apple tissue cultures. (a−f) The error bars indicate the standard deviation (SD, n = 3). * Indicates significant differences between GL-3 and the MdMYB5-RNAi apple lines (p < 0.05, based on t-test).

Figure 4. The phenotype of MdMYB5-RNAi apple tissue cultures under salt and osmotic stress.
Additionally, we constructed the overexpression vector of MdMYB5 (Fig. 5a) and transferred it into Arabidopsis plants to ensure the accuracy of the experimental results. It was found that transgenic Arabidopsis seedlings ectopically expressing MdMYB5 (Fig. 5c) were significantly shorter than wild-type (Fig. 5b). The lignin and cellulose content were also higher than those in the wild-type (Fig. 5d, e). The above results confirmed that MdMYB5 does have a positive regulatory effect on the formation of secondary walls.
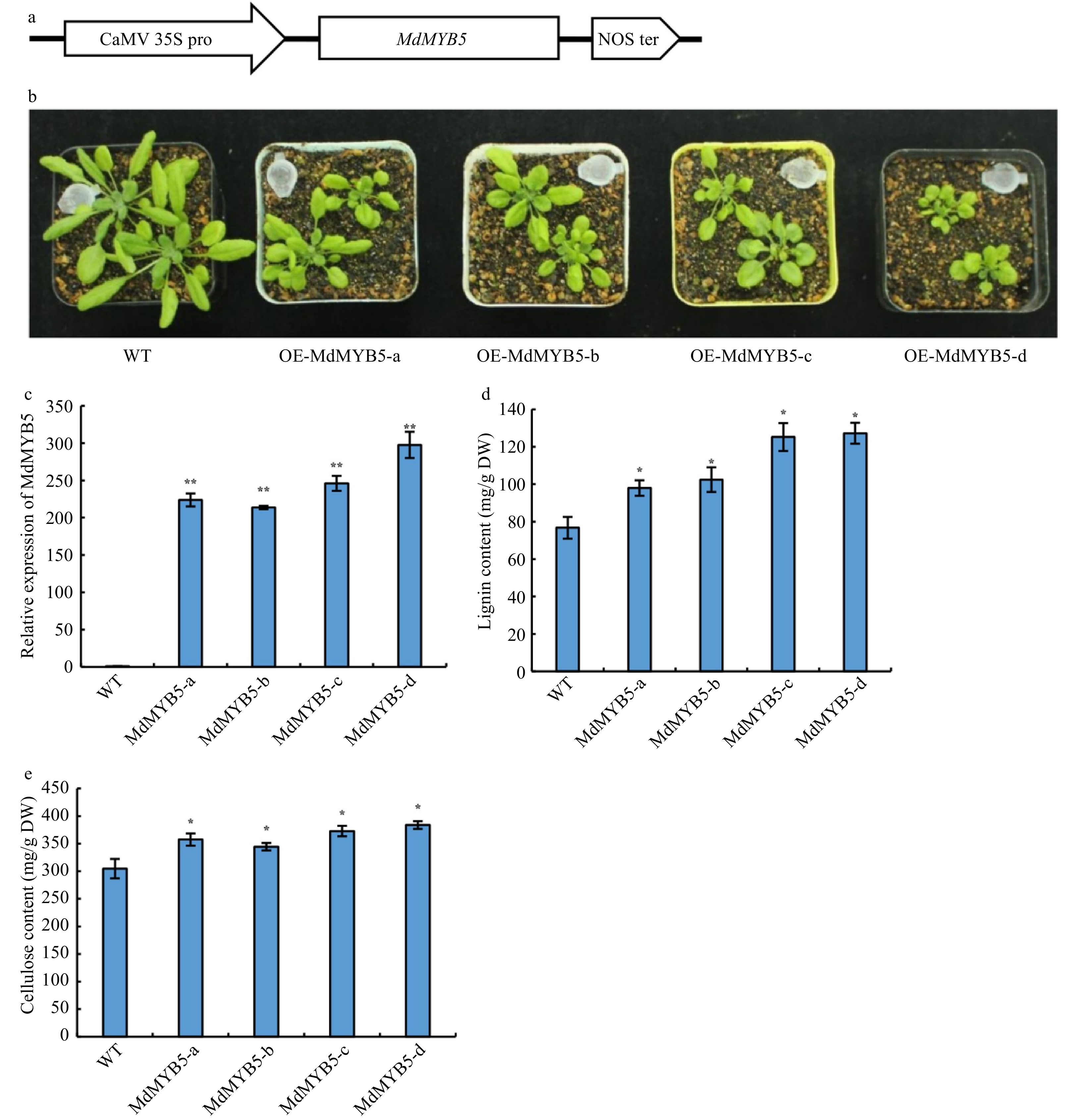
Figure 5. MdMYB5 promotes the formation of secondary cell wall in Arabidopsis. (a) Schematic showing the overexpressing vector for MdMYB5. (b) The phenotype of Arabidopsis ectopically expressing MdMYB5. (c) Transcriptional levels of MdMYB5 in transgenic Arabidopsis lines. (d) Lignin content in stems of the MdMYB5 overexpressing plants and wild-type. (e) Cellulose content in stems of the MdMYB5 overexpressing plants and wild-type. The error bars reveal the standard deviation (SD, n = 3). * shows notable differences between WT and the OE-MdMYB5 Arabidopsis lines (p < 0.05, based on t-test), ** = p < 0.01.
MdSND1 and MdMYB46 regulate the transcription of MdMYB5 by directly binding to its promoter
-
To reveal the direct or indirect regulating relationship between MdMYB5 and MdMYB46, MdSND1, we cloned the 1,947 bp MdMYB5 promoter sequence ( Supplemental Fig. 2) and analyzed the cis-elements. Two specific binding sites of MdSND1 (SNBE1 and SNBE2) and two MdMYB46 specific binding sites (M46RE1 and M46RE2) (Fig. 6a) were found in MdMYB5 promoter. EMSA experiments demonstrated that MdSND1 and MdMYB46 can bind to the SNBE and M46RE elements, respectively (Fig. 6b, c). In order to further verify the binding of MdSND1 and MdMYB46 to the MdMYB5 promoter, the MdMYB5 promoter were divided into two segments (one segment containing two SNBE sites, named proMdMYB5-2; the other containing two M46RE sites, named proMdMYB5-1) and inserted them into the vector containing the luciferase reporter gene, then co-injected the tobacco leaves with 35S:MdSND1 and 35S:MdMYB46, respectively (Fig. 6d, e). Luciferase fluorescence signal and luciferase activity assay showed that MdSND1 and MdMYB46 can bind to MdMYB5 promoter and promote its transcription (Fig. 6f). In vivo ChIP-qPCR experiments also demonstrated that MdSND1 and MdMYB46 can bind to the promoter of MdMYB5 (Fig. 6g, h). The above results show that MdMYB5 could be directly activated by MdSND1 and MdMYB46.
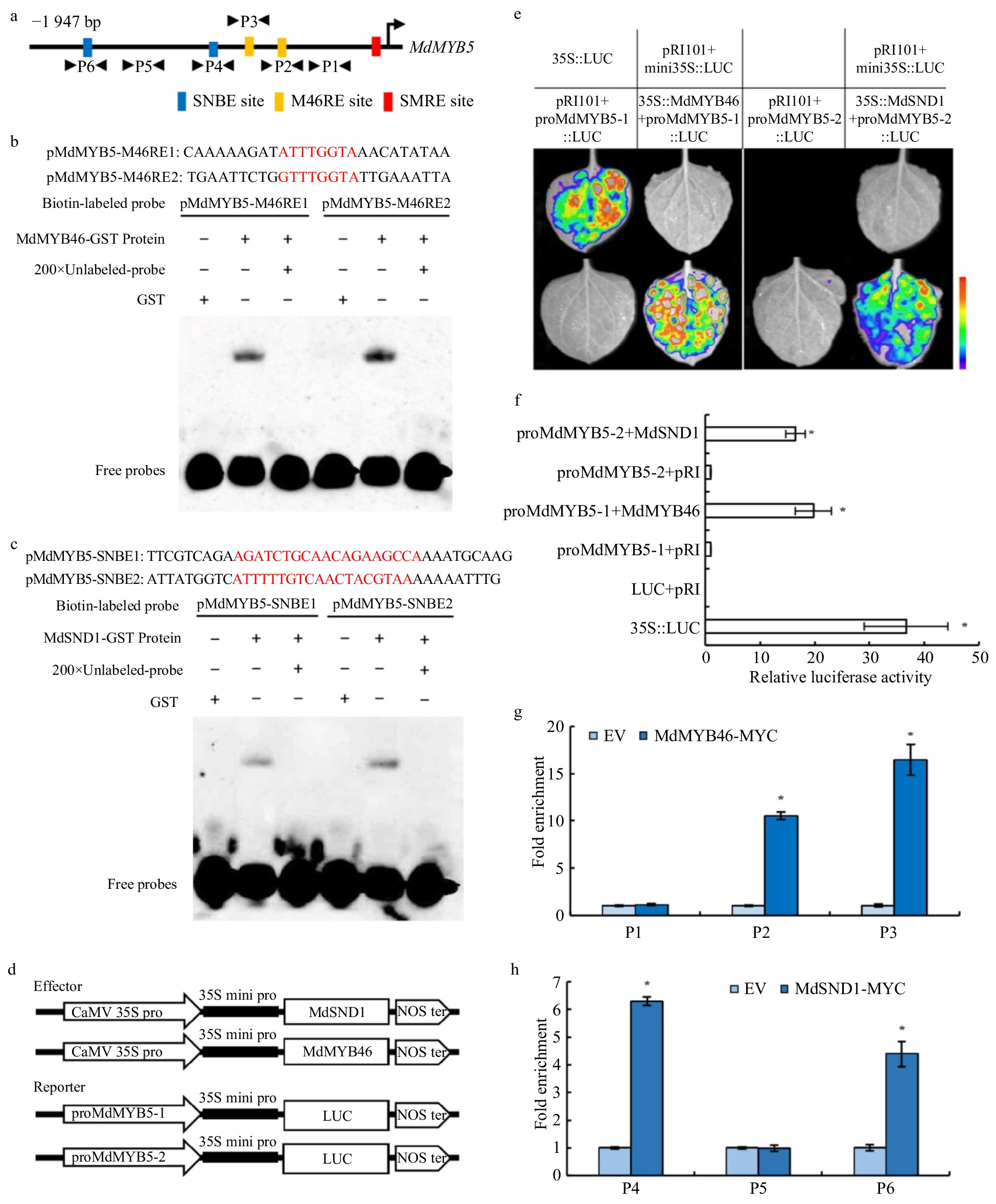
Figure 6. Identification of SNBE, SMRE/M46RE motifs in the promoter of MdMYB5 and the activation effect of MdMYB46 and MdSND1 on their transcription. (a) Schematic representation of MdMYB5 promoter showing the position of MdSND1 and MdMYB46 binding sites. (b) EMSA showing the binding characteristics of the MdMYB46 to M46RE fragments with unlabeled DNA probe (200-fold). Red represents the binding site sequence. (c) EMSA showing the binding characteristics of the MdSND1 to SNBE fragments with unlabeled DNA probe (200-fold). (d) Schematic diagrams of the effector and reporter plasmid structure in the tobacco transient expression assay. (e) Split-luciferase reporter signal showing the interaction between MdSND1, MdMYB46 and MdMYB5 promoter fragments in Nicotiana benthamiana leaves. (f) The comparison of luciferase activity between MdSND1, MdMYB46 and MdMYB5 promoter fragments in Nicotiana benthamiana leaves. (g) and (h) ChIP-qPCR assay of MdSND1 and MdMYB46 binding to the promoter of MdMYB5, respectively. P1 to P6 indicate the fragment of MdMYB5 promoter shown in (a). * Indicates significant difference (p < 0.05, based on t-test). The error bars indicate the standard deviation (SD, n = 3), EV = empty vector.
-
The NAC transcription factor family can regulate multiple downstream target genes by recognizing numerous DNA-binding sequences, such as NAC binding sites with GCTT as consensus binding motif[25], the secondary wall NAC binding elements (SNBE) with (T/A)NN(C/T)(T/C/G)TNNNNN NNA(A/C)GN(A/C/T)(A/T) as core sequence[26] and the specific recognition sequence CA(A/C)G(T/C) (T/C/A)(T/C/A)[27]. SND1 is specifically expressed in interfibers and lignocellulosic fibers in plant stems and can activate the expression of downstream genes by binding SNBE sites[28]. In regulating the network of secondary wall formation in Arabidopsis, MYB46 is a key player at the transcriptional level[29] and activates cellulose and lignin biosynthesis related genes mainly by binding elements such as AC-box, MYBCORE and SMRE in the promoter region[30−31]. In this study, there were SNBE and M46RE/SMRE sites in the promoter of MdMYB5 and the results of EMSA, luciferase reporter assays and ChIP further confirmed that the transcription level of the MdMYB5 gene could be regulated by MdMYB46 and MdSND1, which demonstrated that MdMYB5 had a close relationship with the biosynthesis of cellulose and lignin.
In Arabidophsis, the MYB5 promoter was active in trichomes, stipules, leaf margins, and developing seeds[20, 32]. AtMYB5 collaborated with TTG1 in seed coat development[32]. Additionally, only about 30% and 50% of the PA content were detected in transgenic plants when inhibiting the transcription of MYB5 and MYB14, respectively, and the PA level reduced further in myb5/myb14 double mutants. The function of MYB5 is likely different in different plants. AtMYB5 had only a minor effect on PA biosynthesis in Arabidopsis[20], but ectopic expression of grape VvMYB5a/b in tobacco induced PA accumulation in flowers[22−23]. In Arabidopsis, more small trichomes will be formed with overexpression of AtMYB5[17], as well as a reduction in the number of leaf trichomes and the branching of plants. However, no small trichomes and ectopic trichomes exist in the MdMYB5 ectopically expressing Arabidopsis, demonstrating that there are also functional differences between AtMYB5 and MdMYB5.
Similar to the phenotype of Arabidopsis ectopically expressing MdMYB46[18], the plant became shorter after ectopically expressing MdMYB5, accompanied by the accumulation of lignin and cellulose and the thickening of SCW, suggesting that MdMYB5 can regulate the formation of secondary walls by activating the expression of SCW formation related genes.
Plant cells usually lose water or even die as a result of salt and osmotic stress, nevertheless, lignin and cellulose can help to maintain membrane integrity and reduce plant cell water penetration and transpiration[33]. Many MYB and NAC regulatory genes play a key role in lignin and cellulose biosynthesis and SCW formation in land plants[31]. In apple, the deposition of cellulose and lignin in root cell walls and root xylem vessel formation could be regulated by MdMYB88 and MdMYB124, this leads to a greater hydraulic conductivity and thus increases drought tolerance[17]. In this study, the stress tolerance of MdMYB5-RNAi apple tissue cultures declined, which is possibly caused by the decrease of lignin and cellulose content in the SCW and its capacity in maintaining cell water. MYB transcription factors have a highly conserved DNA binding domain at their N terminus. Previous studies have shown that several MYB proteins, like AtMYBS3, AtMYB2, AtMYB14, and AtMYB15 could affect the resistance to abiotic stress of plants by regulating the inner stress-responsive signal transduction pathways[34−37]. In Arabidopsis, AtMYB2 proteins activated the expression of the ABA-inducible gene under drought stress. As the downstream target of AtMYB46 during secondary wall formation, plant drought resistance was improved by AtMYB52 in regulating the transcription of AtRD29B and AtNCED3[12]. AtMYBS3, AtMYB14 and AtMYB15 could repress the expression of CBF genes and affect the freezing tolerance in Arabidopsis[35−37]. In apple, we previously found that MdSND1 and MdMYB46 could improve the resistance to salt and osmotic stress of apple by directly activating SCW biosynthesis pathways and stress response signals[18−19]. In this study, manipulating the transcription of MdMYB5 significantly reduced the content of lignin and cellulose in apple stems and inhibited the formation of SCWs. Meanwhile, stress treatment experiments also showed that the stress response of apples involves MdMYB5. MdMYB5 likely regulates stress tolerance in apple partially by affecting the activation of stress signals, however the detailed mechanism requires further investigation.
-
Genetic transformation and stress treatment experiments were performed with the apple plant GL-3 (Malus × domestica Borkh.) previously screened in our laboratory, with high efficiency of transformation[38]. Apple tissue cultures were cultured according to the condition described by Chen et al.[19]. The callus of 'Ourin' apple was cultured in the medium (MS medium supplemented with 0.4 mg/L 6-BA, 1.5 mg/L 2,4-D) at 25 °C in the dark. Nicotiana benthamiana was used for subcellular localization and transient expression of genes, and Arabidopsis thaliana for transformation was managed under long-day photoperiod (16 h : 8 h, light : dark) at 23 °C. T test was used to indicate the difference between the wild-type and transgenic plants.
Vector and transformation
-
The MdMYB5 gene sequence was from the new apple genome initiative under locus identifier MD11G1006500. The forward and reverse sequences of MdMYB5 gene were amplified with MdMYB5-RNAi ZF/ZR and FF/FR primer pairs (Supplemental Table 1), respectively. To construct the MdMYB5-RNAi vector, the cloned sequence was inserted into the pRNAi-E vector[39]. The RNAi vectors of MdMYB5 were then transferred into the Agrobacterium tumefaciens EHA105. The apple plant GL-3 was transformed with leaves as explants, according to the method of Dai et al.[38]. Three RNAi transgenic apple lines, namely MdMYB5-RNAi-1, MdMYB5-RNAi-2 and MdMYB5-RNAi-3, were finally screened out and each line was used as a biological replicate.
Meanwhile, the overexpressing A. thaliana lines were also used for the functional analysis of MdMYB5. The MdMYB5 full-length CDS region was inserted into the PRI101-AN vector to construct the overexpression vector of pRI-MdMYB5. Floral dip method was used for A. thaliana transformation via Agrobacterium tumefaciens strain GV3101. The transformed seeds were then screened in 1/2 MS medium supplemented with 30 mg/L Kan and transgenic seedlings were identified by PCR. Four screened overexpressing transgenic Arabidopsis seedlings, namely MdMYB5-a, MdMYB5-b, MdMYB5-c, and MdMYB5-d, were used as biological replicates, respectively.
Microscopic analysis
-
When the transgenic apple plants were 1 month old, the transverse sections of the basal stems were cut and prepared for paraffin sections. The sections were then stained with Safranin and Fast Green FCF, observed and photographed using a microscope. The red part represented the structure of lignification.
ChIP-qPCR
-
The chromatin immunoprecipitation was carried out in accordance with the methods of Chen et al.[18]. The positive calli transformed with pRI-MYC-MdMYB46 and pRI-MYC- MdSND1 of 'Ourin' were screened. The main steps of the chromatin immunoprecipitation experiment were based on the instructions of EPIGENTEC's Plant ChIP kit (Farmingdale, USA). MdACTIN was used as the internal reference gene and the 2−ΔΔCt method was used to calculate the fold enrichment of each promoter fragment in ABI 7500 software v2.0.6. Duncan’s multiple range test was used to perform substantial difference analysis.
Stress treatment
-
The 20-day-old wild-type GL-3 and RNAi plants were used for stress treatment. The young shoot tip of these plants were cut and cultured in a subculture medium containing 200 mM NaCl or 300 mM mannitol for 10 days. The phenotype of these plants were photographed after treatment, and their leaves were collected for further analysis.
Analysis of transcriptional activity
-
The constructed pRI-MdMYB46 and pRI-MdSND1 vector were used as effectors. The clipped region (about 200−500 bp fragment containing the corresponding binding site) of MdMYB5 was inserted into the vector of pRI-mini35S-LUC and used as a report. The conserved bases of the binding sites in the MdMYB5 promoter fragments were mutated by PCR and ligated into the vector of pRI-mini35S-LUC as a control.
The above constructed vector was transferred into the leaves of 1-month-old N. benthamiana for transient expression. The infected tobacco was cultured for 24 h in the dark and then transferred to a light incubator (25 °C, 16 h : 8 h, light : dark). After 48 h, the tobacco was photographed by a living fluorescence imager (Lb985, Berthold, Germany). The luciferase activity of these leaves was measured using a Transgen Biotech firefly luciferase assay kit (Beijing, China).
EMSA
-
The full-length CDS region of MdSND1 and MdMYB46 were inserted into the prokaryotic expression vector pGEX-5X-1 and labeled as pGEX-MdSND1 and pGEX-MdMYB46, respectively. The constructed vector was then separately introduced into the competent cells of Escherichia coli BL21 to induce protein production. The GST protein was purified according to the instructions of the Thermo GST Fusion Protein Purification Kit (Massachusetts, USA). The EMSA probe was prepared according to the Biyuntian Biotin Probe Labeling Kit (Shanghai, China) and the fusion of labeled-probe made with reference to the EMSA kit.
Subcellular localization, qRT-PCR and content of lignin and cellulose
-
The subcellular localization, qRT-PCR and measurement of lignin and cellulose content were carried out in accordance to the methods of Chen et al.[18]. Sequences of primers used in qRT-PCR are shown in Supplemental Table 1.
Gene sequence
-
Sequences of genes used in this study were from the new apple genome initiative under locus identifiers MdMYB5 (MD11G1006500), MdMYB46 (MD03G1176000), MdSND1 (MD06G1121400), MdMYB83A (MD07G1010500), MdMYB83B (MD02G1312700), MdCOMT (MD05G1083900), Md4CL (MD13G1257800), MdCESA4 (MD15G1150300), MdCESA8 (MD10G1276500).
- This work was supported by grants from the National Natural Science Foundation of China (31972380; 31170635).
- The authors declare that they have no conflict of interest.
- Supplemental Fig. 1 MdMYB5 expression levels in MdSND1 and MdMYB46 transgenic and non-transformed apple plants. A. MdSND1 transgenic apple plants; B. MdMYB46 transgenic apple plants. The error bars indicate the standard deviation (SD, n = 3). Asterisk indicates significant differences between the transgenic lines and GL-3 (p < 0.05, based on t-test).
- Supplemental Fig. 2 The sequence of MdMYB5 promoter.
- Supplemental Table 1 Primers used in this study.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen K, Tang X, Song M, Guo Y, Liu L, et al. 2021. Functional identification of MdMYB5 involved in secondary cell wall formation in apple. Fruit Research 1: 6 doi: 10.48130/FruRes-2021-0006

Functional identification of MdMYB5 involved in secondary cell wall formation in apple
- Received: 20 April 2021
- Accepted: 19 July 2021
- Published online: 27 July 2021
Abstract: Being the principal elements of secondary cell wall, cellulose and lignin both play a strengthening role in plant structures and stress resistance. However, little research has been done regarding the molecular mechanisms involved in the formation of cellulose and lignin in apple. In this study, in order to better understand the regulatory network in the formation of secondary cell wall, an R2R3 MYB transcriptional factor MdMYB5 was identified and explored. The subcellular localization experiments showed that MdMYB5 could function in the nucleus. Even though lignin and cellulose content, and the expression of their biosynthesis related genes decreased in the MdMYB5-RNAi apple, the ectopic overexpression of MdMYB5 promotes lignin and cellulose content in Arabidopsis, which contributes to the dwarf phenotype. At the same time, salt and osmotic stress affect MdMYB5-RNAi apple tissue cultures. Further transcriptional activation assays carried out demonstrated that MdMYB5 could be activated by MdMYB46 and MdSND1. In conclusion, MdMYB5 was required for the normal formation of secondary cell wall in apple and could be activated by the key regulatory factors MdMYB46 and MdSND1.
-
Key words:
- Apple /
- MdMYB5 /
- Cellulose /
- Lignin /
- Resistance to salt and osmotic stress