








-
With the improved awareness of nutrition and health worldwide, the demand for healthy dietary components has received increasing attention, and the consumption of nutrient supplements has been increasing. Numerous studies have emphasized that fruits confer a protective effect against human degenerative diseases such as diabetes, obesity, cardiovascular disease, and other chronic diseases, due to inherent richness in flavonoid compounds[1,2]. Fruits have been noted to play significant roles in nutrition and human health, especially as sources of vitamins, minerals, dietary fiber, and bioactive compounds[3−5].
Flavonols are by far the most widespread flavonoids, and naturally exist in plant vacuoles in the form of their glycoside derivatives. Currently, at least 15 flavonol aglycones have been identified in fruits, of which quercetin, kaempferol and myricetin are the most common ones. Glycosylation, hydroxylation, methylation and acylation enriches the types of flavonol derivatives present. It has been shown that flavonols play an important role in plant growth and development and resistance to stress, including regulating auxin transport, affecting pollen development, promoting lateral root formation, and responding to, and protecting against, ultraviolet (UV) light[6−10]. As bioactive compounds, flavonols are known to exhibit antioxidative, anti-inflammatory, anticancer and other pharmacological activities, and help prevention of cardiovascular disease and diabetes[11−13]. With the development of new chromatographic techniques and molecular biology methods, more flavonols have been identified in different fruits, and the metabolic mechanisms affecting flavonol accumulation have been analyzed. However, most reviews of flavonols in recent years have focused on health benefits and bioavailability[14,15] and information about metabolic mechanisms that determine the accumulation of specific flavonols in dietary fruits is often overlooked.
Here, we review current knowledge concerning molecular structures, distribution, biosynthetic mechanisms, transcriptional regulation, and metabolic engineering of fruit flavonols. Particular attention is paid to the roles of key structural enzymes, other proteins that add specific chemical modifications that affect structure and properties, and transcription factors important in regulating the biosynthesis pathway. Plant responses to environmental factors that influence accumulation of flavonols are also highlighted. Understanding the knowledge of molecular mechanisms controlling flavonol biosynthesis will facilitate future bioengineering programs to produce desirable levels of targeted bioactivities in our dietary fruits.
-
Flavonols are constructed from 15-carbon skeletons and are composed of two aromatic rings (A and B ring) connected via a three-carbon chain (C ring) to form a basic diphenylpropane backbone (C6-C3-C6) with hydroxyl groups at the carbon 3 position (Fig. 1a). The A ring is normally formed from three malonyl-CoA molecules generated via the acetate pathway and exhibits a characteristic hydroxylation pattern at the carbon 5 and 7 sites. The B ring carbon originates from p-coumaroyl-CoA produced from phenylalanine via the shikimate pathway, and is often hydroxylated at carbon 4’, 3’4’, or 3’4’5’ positions (Table 1). Among the flavonol aglycones identified in fruits, kaempferol is the predominant structure and most other types including quercetin, myricetin, isorhamnetin, morin, laricitrin, gossypetin, kaempferide, natsudaidain, quercetagetin, syringetin, sexangularetin, rhamnetin are considered to be kaempferol derivatives carrying substituted hydroxyl groups or methyl groups at the different positions of the flavonol skeletons (Table 1). For example, quercetin, a 3’-hydroxykaempferol, is widespread in fruits[16]. Morin is hydroxylated at the 2’ carbon of kaempferol and accumulates mainly in mulberry[17]. Kaempferide, a 4’-O-methylkaempferol, occurs in grape (Vitis vinifera)[18]. Galangin and fisetin are not regarded as kaempferol derivatives, however. Galangin has no OH group on the B ring and has been reported in grape[19] and blueberry (Vaccinium L.)[20]. Fisetin is not hydroxylated at the 5-carbon position of the A ring and occurs in mulberry[17]. Hydroxylated flavonol aglycones are highly unstable in vivo, and methylation modifications help to enhance stability. Isorhamnetin, with methylation at the 3’ site, is the most common methylated flavonol aglycone and occurs mainly in pear (Pyrus communis L.)[21] and peach (Prunus persica L.)[22].
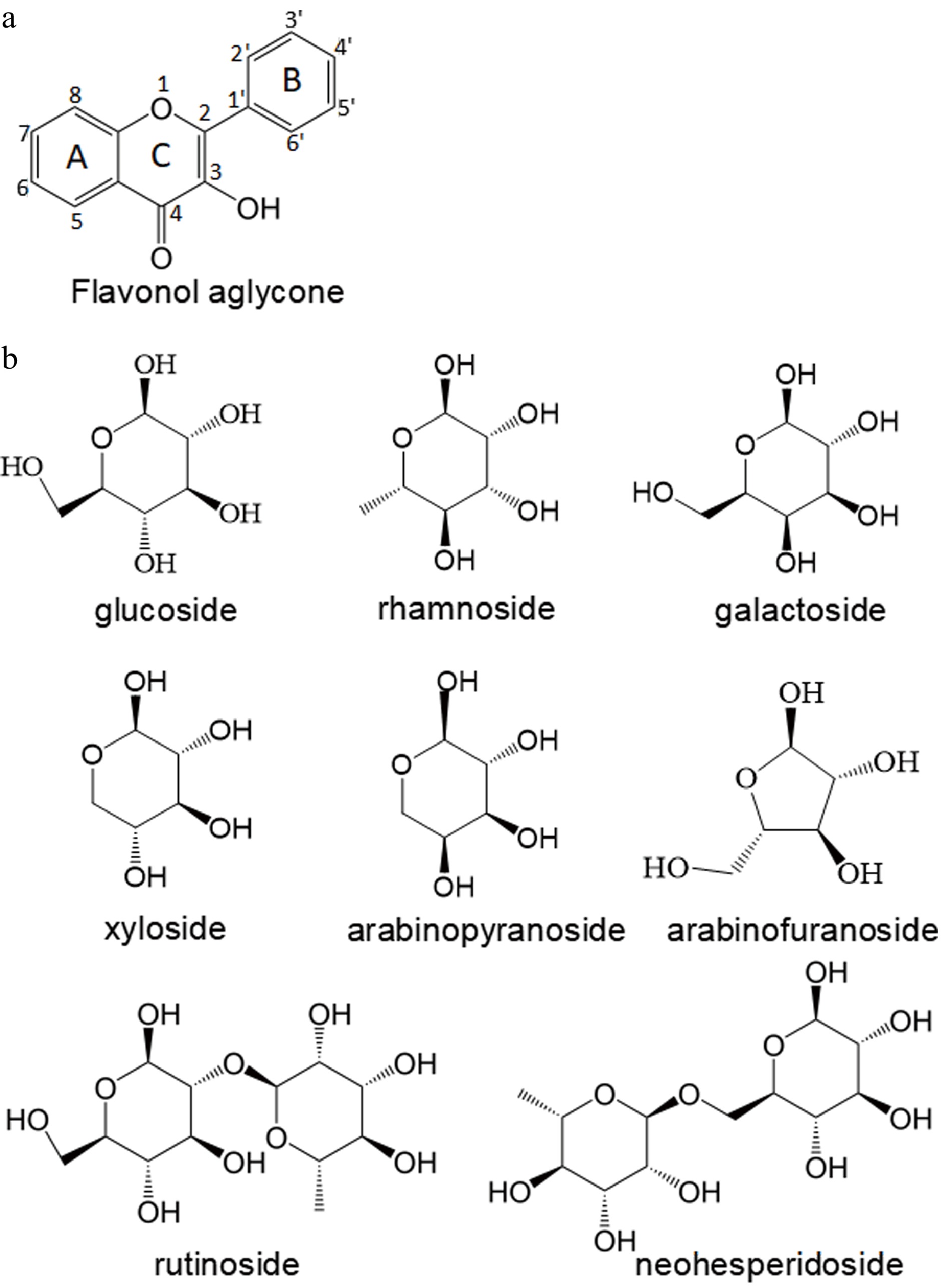
Figure 1. General structure of (a) flavonol aglycones and (b) main glycosides.
Table 1. Summary of flavonol aglycones identified in fruits.
Aglycones 5 6 7 8 2’ 3’ 4’ 5’ 6’ Reference Kaempferol OH H OH H H H OH H H [16] Quercetin OH H OH H H OH OH H H [16] Myricetin OH H OH H H OH OH OH H [16] Galangin OH H OH H H H H H H [19,20] Gossypetin OH H OH OH H OH OH H H [30] Kaempferide OH H OH H H H OCH3 H H [25] quercetagetin OH OH OH H H OH OH H H [31] Laricitrin OH H OH H H OCH3 OH OH H [25] Morin OH H OH H OH H OH H H [17] Isorhamnetin OH H OH H H OCH3 OH H H [16] Natsudaidain OCH3 OCH3 OCH3 OCH3 H H OCH3 OCH3 H [32] Syringetin OH H OH H H OCH3 OH OCH3 H [33] Sexangularetin OH H OH OCH3 H H OH H H [34] Rhamnetin OH H OCH3 H H OH OH H H [17] Fisetin H H OH H H H OH OH H [17] Flavonols are most frequently found in nature in the form of glycosides due to the unstable physicochemical properties of their aglycones. Most of the sugar ligands attached to flavonol aglycones are glucoside, galactoside, rhamnoside, xyloside, and arabinoside, and these sugar moieties usually accumulate in the form of mono-, or diglycosides in fruits (Fig. 1b). The glycosidic linkage can be divided into O-glycosidic bonds and C-glycosidic bonds. Sugar ligands are generally attached to an oxygen atom at carbon 3, 5, 7, 8, 3’, 4’, or 5’ positions to form flavonol O-glycosides, of which 3-oxyglycosides are the most common ones, while flavonol C-glycosides are attached to the carbon atom at position 6 or 8. In addition, hydroxyl or acyl groups can also be attached to the parent ring of flavonols, which contribute to the structural diversity of flavonols and play an important role in their diverse biological functions.
Flavonol compounds in fruits
-
Different types of flavonol metabolites are found in specific fruit, and sugar moieties are most commonly attached to an oxygen atom at carbon 3 (Table 2). With the development and application of high-resolution mass spectrometry, more and more flavonol compounds have been identified, among which kaempferol and quercetin glycosides are the most common dietary flavonols and can be detected in most of fruits, while the distribution of other flavonol glycosides is relatively limited (Table 2). Apple (Malus domestica) is well-known for accumulating quercetin glycosides and estimates obtained by comparing HPLC peak areas with standard curves indicate a content of about 150 mg kg−1 fresh weight (FW) quercetin 3-O-rhamnoside and 100 mg kg−1 FW quercetin 3-O-galactoside respectively[10]. Isorhamnetin and myricetin derivatives are less widespread flavonols compared to kaempferol and quercetin glycosides. It has been demonstrated that isorhamnetin metabolites are the major flavonols in pear with content of isorhamnetin 3-O-galactoside up to 65.15 mg kg−1 FW, quantified by comparing peak area with the standard curves using UPLC[21], and they have also been detected in peach[22], grape[23], strawberry (Fragaria × ananassa)[24] and blueberry[25]. Myricetin compounds are mainly distributed in berry fruits, especially Chinese bayberry (Morella rubra)[11], blueberry[25] and grape[18]. So far, little research has been carried out on less common flavonols in fruits. Rhamnetins have been detected mainly in mango (Mangifera indica L.)[26], and laricitrins and syringetins have been identified mainly in blueberry[25], while sexangularetins has been found only in hawthorn (Crataegus laevigata)[27]. However, the distribution pattern of flavonols in fruits depends on the degree of accessibility to previous illumination due to the fact that their formation is accelerated by light. For example, the content of flavonols is usually higher in the peel of peach and persimmon (Diospyros kaki Thunb.) than in the pulp[28, 29]. Generally, flavonol glycosides are located mainly in the outer parts of fruits such as the peel and they decrease in concentration toward the central core.
Table 2. Distribution of divergent flavonols in fruits. The first three listed compounds are the major flavonols in each fruit.
Fruit species Divergent flavonols Reference Apple Quercetin 3-O-rhamnoside; Quercetin 3-O-galactoside; Quercetin 3-O-arabinoside; Quercetin 3-O-glucoside; Quercetin 3-O-xyloside; Quercetin 3-O-robinobioside; Quercetin 3-O-rutinoside; Quercetin 3-O-neohesperidoside; Kaempferol 3-O-galactoside; Kaempferol 3-O-arabinoside; Isorhamnetin 3-O-galactoside; Isorhamnetin 3-O-glucoside; Rhamnetin 3-O-rutinoside [10,35] Pear
Quercetin 3-O-glucoside; Isorhamnetin 3-O-galactoside; Isorhamnetin 3-O-rutinoside; Isorhamnetin 3-O-malonylglucoside; Isorhamnetin hexoside; Isorhamnetin 3-O-malonylgalactoside; Isorhamnetin 3-O-glucoside; Isorhamnetin; Quercetin 3-O-galactoside; Quercetin 3-O-galactosyl-glucoside; Quercetin 3-O-rutinoside; Quercetin 3-O-arabinoside; Quercetin O-acetylhexoside; Quercetin 5-O-malonylhexosyl-hexoside; Quercetin 7-O-malonylhexosyl-hexoside; Quercetin 4’-O-glucoside; Kaempferol 3-O-galactoside; Kaempferol 3-O-rutinoside; Kaempferol 3-O-acetylglucoside; Kaempferol 3-O-rhamnoside; Rhamnetin hexoside; [21,36,37,38] Peach Quercetin 3-O-glucoside; Isorhamnetin 3-O-rutinoside; Isorhamnetin 3-O-glucoside; Quercetin 3-O-rhamnoside; Quercetin 3-O-galactoside; Quercetin 3-O-rutinoside; Kaempferol 3-O-rutinoside; Kaempferol 3-O-glucoside [22,28] Loquat Quercetin 3-O-glucoside; Quercetin 3-O-galactoside; Kaempferol 3-O-sophoroside; Quercetin 3-O-rhamnoside; Quercetin 3-O-rutinoside; Quercetin 3-O-neohesperidoside; Quercetin 3-O-sophoroside; Quercetin-3-O-galactosyl-glucoside; Quercetin 3-O-sambubioside; Kaempferol 3-O-neohesperidoside; Kaempferol 3-O-sambubioside; Kaempferol 3-O-rhamnoside; Kaempferol 3-O-glucoside; Kaempferol 3-O-rutinoside [39,40] Hawthorn Quercetin 3-O-galactoside; Quercetin 3-O-glucoside; Quercetin 3-O-rutinoside;
Kaempferol 3-O-glucoside; Kaempferol 3-O-neohesperidoside; Sexangularetin; Sexangularetin 3-O-neohesperidoside; Sexangularetin 3-O-glucoside[27,34] Grape Quercetin 3-O-glucoside; Quercetin 3-O-glucuronide; Myricetin 3-O-glucoside; Quercetin 3-O-galactoside; Quercetin 3-O-rutinoside; Quercetin; Kaempferol 3-O-galactoside; Kaempferide coumaroylhexoside; Kaempferol 3-O-glucoside; Myricetin 3-O-glucuronide; Myricetin dihexoside; Myricetin glucoside-glucuronide; Isorhamnetin 3-O-glucoside; Isorhamnetin 3-O-glucoside; Isorhamnetin glucuronide; Isorhamnetin coumaroylglucoside; Isorhamnetin; Laricitrin 3-O-glucoside; Syringetin-dihexoside; Syringetin 3-O-glucoside; Syringetin 3-O-galactoside [18,23,41] Blueberry Quercetin 3-O-galactoside; Quercetin 3-O-rhamnoside; Quercetin 3-O-rutinoside; Quercetin 3-O-glucoside; Quercetin 3-O-pentoside; Quercetin 3-O-glucoside acetate; Quercetin 3-O-arabinoside; Myricetin 3-O-galactoside; Myricetin 3-O-glucoside; Myricetin 3-O-pentoside; Myricetin 3-O-rhamnoside; Kaempferol 3-O-rutinoside; Kaempferol 3-O-glucoside; Laricitrin 3-O-galactoside; Laricitrin 3-O-glucoside; Laricitrin 3-O-rhamnoside; Laricitrin 3-O-pentoside; Isorhamnetin 3-O-galactoside; Isorhamnetin 3-O-rhamnoside; Isorhamnetin 3-O-glucoside; Syringetin 3-O-glucoside; Syringetin 3-O-rhamnoside; Syringetin 3-O-pentoside; Syringetin 3-O-galactoside [20,25,42] Bayberry Myricetin 3-O-rhamnoside; Quercetin 3-O-galactoside; Quercetin 3-O-rhamnoside; Myricetin 3-O-glucoside; Myricetin deoxyhexoside-gallate; Quercetin 3-O-glucuronide; Quercetin 3-O-arabinoside; Kaempferol 3-O-rhamnoside; Kaempferol 3-O-galactoside; Kaempferol 3-O-glucoside; Isorhamnetin 3-O-rhamnoside; Isorhamnetin 3-O-glucoside [11,43] Mulberry Quercetin 3-O-rutinoside; Kaempferol 3-O-glucoside; Quercetin 3-O-glucoside; Quercetin 3-O-rhamnoside; Quercetin 3-O-galactoside; Quercetin 3-O-glucuronide; Quercetin; Myricetin 3-O-rhamnoside; Isorhamnetin 3-O-glucoside; Myricetin; Kaempferol; Isorhamnetin; Fisetin; Morin; Rhamnetin; Galangin; Kaempferide [17] Strawberry Quercetin glucuronide; Quercetin pentoside; Kaempferol coumaroylhexoside; Quercetin 3-O-glucoside; Quercetin 7-O-glucoside; Quercetin 4’-O-glucoside; Kaempferol 3-O-glucoside; Kaempferol 7-O-glucoside; Kaempferol 4’-O-glucoside; Kaempferol glucuronide; Isorhamnetin 3-O-glucoside; Isorhamnetin 7-O-glucoside; Isorhamnetin 4’-O-glucoside; Isorhamnetin glucuronide [24,44] Cherry Quercetin 3-O-rutinoside; Kaempferol 3-O-rutinoside; Quercetin 3-O-glucosil-rutinoside; Quercetin 3-O-rhamnoside; Quercetin 3-O-galactoside; Quercetin 3-O-glucoside; Quercetin 3-O-diglucoside; Kaempferol 3-O-glucoside; Kaempferol 3-O-rhamnoside; Isorhamnetin 3-O-rutinoside [45] Tomato Quercetin 3-O-rutinoside; Kaempferol 3-O-rutinoside; Quercetin glucosyl-glucoside rhamnoside; Quercetin 3-O-rutinoside-7-O-glucoside; Quercetin 3-O-glucoside; Quercetin 3,7-O-glucoside; Kaempferol glucosyl-glucoside rhamnoside; Kaempferol 3-O-glucoside; Kaempferol 3,7-O-glucoside; Kaempferol 3-O-rutinoside-7-O-glucoside [46,47,48] Mango Quercetin 3-O-galactoside; Quercetin 3-O-glucoside; Quercetin 3-O-xyloside; Quercetin diglycoside; Quercetin 3-O-arabinopyranoside; Quercetin 3-O-arabinofuranoside; Quercetin 3-O-rhamnoside; Rhamnetin 3-O-galactoside; Rhamnetin 3-O-glucoside; Rhamnetin 3-O-galactopyranoside; Rhamnetin 3-O-glucopyranoside; Kaempferol 3-O-glucoside; Quercetin; Isorhamnetin 3-O-glucoside [26] Litchi Quercetin rhamnosyl-rutinoside; Quercetin 3-O-rutinoside; Isorhamnetin rhamnosyl-rutinoside; Quercetin rhamnosyl-glucoside; Isomer of Quercetin rhamnosyl-glucoside; Quercetin 3-O-rutinoside-O-rhamnoside; Quercetin glucosyl-rutinoside; Quercetin rhamnosyl-glucosyl-rutinoside; Kaempferol rhamnosyl-rutinoside; Kaempferol 3-O-rutinoside-O-rhamnoside; Kaempferol 3-O-rutinoside; Keampferol rhamnosyl-glucosyl-rutinoside; Isorhamnetin 3-O-rutinoside; Isorhamnetin 3-O-rutinoside-O-rhamnoside; Isorhamnetin glucosyl-rutinoside; Myricetin rutinoside [49,50] Citrus Quercetin 3-O-glucoside; Quercetin 3-O-rutinoside; Quercetin 7-O-glucoside; Quercetin 7-O-rutinoside; Quercetin 3-O-glucofuranoside; Kaempferol 3-O-glucoside; Kaempferol 3-O-rutinoside; Kaempferol 7-O-glucoside; Kaempferol 7-O-rutinoside [51,52] Kiwi fruit Quercetin 3-O-rutinoside; Quercetin 3-O-glucoside; Kaempferol 3-O-rutinoside;
Quercetin 3-O-arabinofuranoside; Quercetin 3-O-rhamnoside; Quercetin 4’-O-glucoside; Kaempferol 3-O-galactoside; Kaempferol 3-O-rhamnoside; Kaempferol 3-O-robinobioside; Kaempferol 3,7-O-diglucoside 8-prenyl derivative; Myricetin 3-O-galactoside; Syringetin[33] -
The mechanisms of flavonol biosynthesis have been widely elucidated and a simplified flavonol metabolic pathway is shown in Fig. 2. Chalcone synthase (CHS) catalyzes the first step in flavonol biosynthesis by converting substrates p-coumaroyl-CoA and malonyl-CoA to product naringenin chalcone[53]. The following second catalytic reaction performed by chalcone isomerase (CHI) and chalcone reductase (CHR) is very important for the corresponding formation of 5,7-oxo and 5-deoxy flavonols. CHI was confirmed to catalyze the stereospecific cyclization of naringenin chalcone to naringenin[54], which is a general precursor for 5,7-oxo flavonols. This step can also proceed spontaneously. CHR, which catalyzes the production of 6’-deoxy chalcone (isoliquiritigenin) through its effects on CHS catalyzed reaction[55], is a key enzyme mediating 5-deoxy flavonol biosynthesis. Flavanone 3-hydroxylase (F3H), flavonoid 3’-hydroxylase (F3’H), flavonoid 3’5’-hydroxylase (F3’5’H), and flavonol synthase (FLS) cover the core metabolic grid of flavonol biosynthesis and the production of different flavonols (Fig. 2). F3H and FLS belong to the 2-oxoglutarate-dependent dioxygenases protein family and catalyze 3-hydroxylation and oxidation of carbon 2 and carbon 3 of flavonols on the C ring. F3’H and F3’5’H are members of the cytochrome P450 protein family and catalyze 3’4’-hydroxylation and 3’4’5’-hydroxylation on the B ring. Thus, the CHI-catalyzed compounds naringenin and liquiritigenin can be converted to the corresponding dihydrokaempferol and garbanzol by F3H[56,57], and then the dihydroflavonols dihydrokaempferol and garbanzol are converted to 5,7-oxo flavonol kaempferol and 5-deoxy flavonol resokaempferol by FLS[57,58]. The 5,7-oxo flavonols quercetin and myricetin are produced directly by FLS consuming the intermediates dihydroquercetin and dihydromyricetin[58,59], which are produced by two hydroxylases: flavonoid 3’-hydroxylase (F3’H) and flavonoid 3’5’-hydroxylase (F3’5’H) respectively[60,61]. Recently, a F3’5’H gene isolated from Chinese bayberry was postulated to be the important factor determining the accumulation of myricetin, because it drives pathway flux towards the trihydroxylated flavonol by hydroxylating kaempferol without the need for a dihydromyricetin specific FLS[62]. Isorhamnetin, a 5,7-oxo methylated flavonol, is produced by the addition of a methyl group to quercetin by O-methyltransferases (OMT)[63]. The 5-deoxy flavonol fisetin is produced by conversion from resokaempferol by F3’H[57].
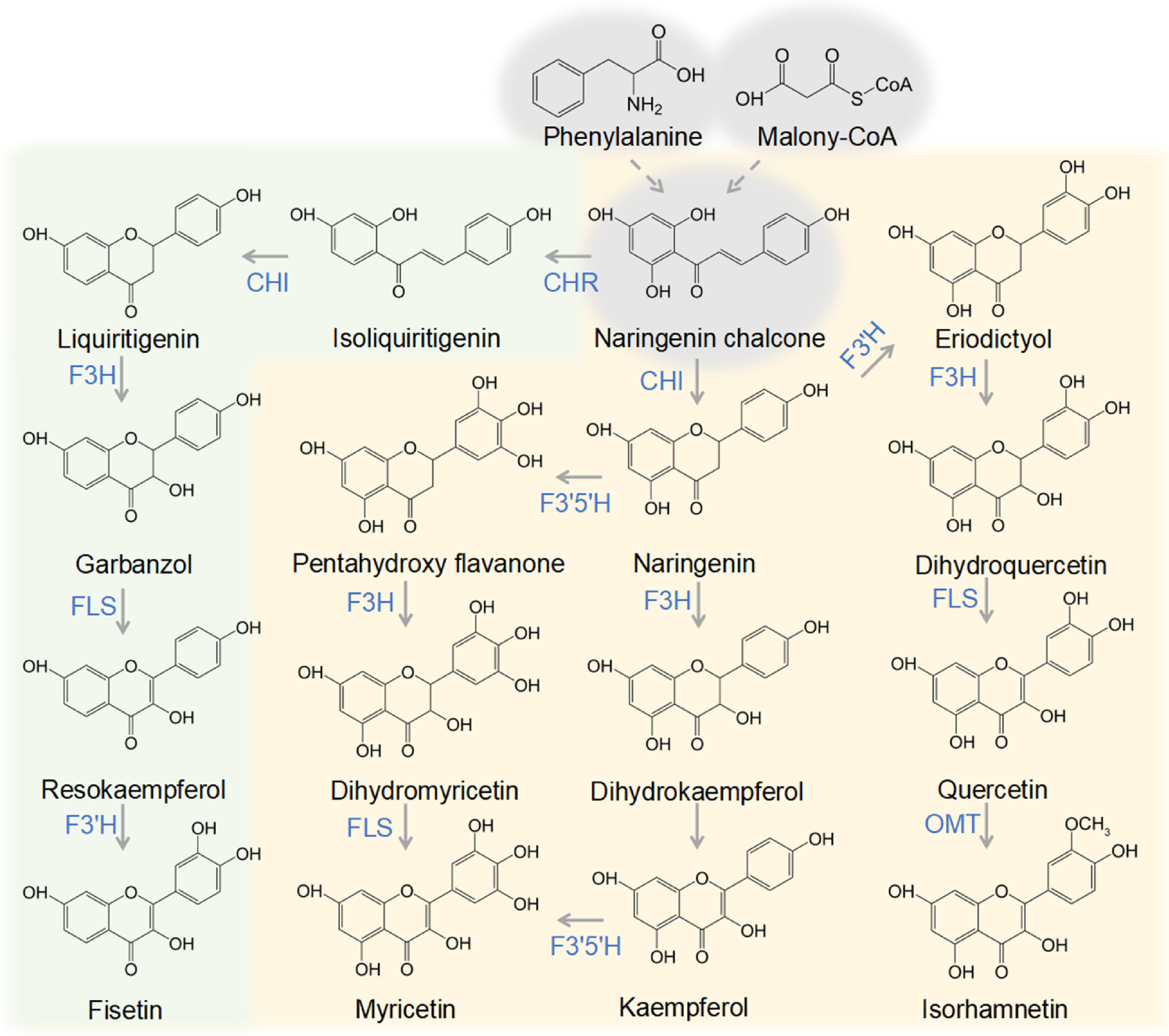
Figure 2. Representative flavonol biosynthetic pathways. The pathways utilize naringenin chalcone, produced from phenylalanine and malonyl-CoA, highlighted in grey. The metabolic pathway of 5,7-oxo flavonols is highlighted in yellow, and biosynthetic pathways for 5-deoxy flavonols are highlighted in green. CHR: chalcone reductase; CHI: chalcone isomerase; F3H: flavanone 3-hydroxylase; F3’H: flavonoid 3’-hydroxylase; F3’5’H: flavonoid 3’5’-hydroxylase; OMT: O-methyltransferases; FLS: flavonol synthase.
Modification of flavonol aglycones
-
Glycosylation, hydroxylation, methylation and acylation are the major modification reactions resulting in the formation of a wide range of flavonol products. These modifications tend to alter the stability, solubility and cellular localization of the corresponding flavonol aglycones. In fruit species, a few genes have now been identified that are involved in catalyzing such decorations of flavonol derivatives.
Flavonols are largely glycosylated by uridine diphosphate glycosyltransferases (UGTs), which use uridine 5-diphosphatesugars (UDP) such as UDP-glucoside, UDP-galactoside, UDP-rhamnoside as the donor molecule. Most fruit UGTs are reported to participate in the generation of 3-oxoglycosylated flavonols. For instance, the enzymes AY519364[64] from citrus (Citrus sinensis), AcF3GT2 from kiwifruit (Actinidia chinensis)[65], and MdUGT71B1 from apple[10] have been confirmed to catalyze the glucosylation of the 3-hydroxyl group of quercetin efficiently, while DkFGT from persimmon[66] and MdUGT75B1 from apple[10] preferentially galactosylated the 3-hydroxyl group of quercetin. In grapevine, VvGT5 was identified as a flavonol-3-O-glucuronosyltransferase that exhibited a strong glucuronosyl transfer activity from UDP-glucuronic acid to kaempferol, quercetin and isorhamnetin. VvGT6 was demonstrated to be a bifunctional glycosyltransferase, which was capable of adding a UDP-glucose or UDP-galactose group to kaempferol, quercetin and isorhamnetin separately[67]. Strawberry UGTs have been reported to be capable of glycosylating at different hydroxyl positions[24]. Using recombinant enzymes, it was shown that both FaGT6 and FaGT7 were able to convert quercetin, kaempferol and isorhamnetin to the corresponding 3-O-glucosides, 7-O-glucosides, and 4’-O-glucosides, respectively. FaGT6 was capable of forming a 3’-O-monoglucoside and one diglucoside with quercetin as a substrate, while FaGT7 only formed 3’-O-monoglycoside but no diglucoside[24]. CsUGT76F1 from sweet orange has been shown to carry out glycosylation at the carbon 3 or 7 position of flavonoids, converting kaempferol and quercetin to the corresponding 3-O-glucosides, 7-O-glucosides, and 7-O-rhamnosides. However, the enzyme CsUGT76F1 was found to be capable of converting kaempferol to its 3,7-O-diglucoside but no quercetin 3,7-O-diglucoside product was formed with quercetin as a substrate[51]. In addition to showing preferences for different glycosylation positions, several fruit UGTs have been found to possess selectivity to receptor flavonol molecules. For example, citrus AY519364 glucosylated only the flavonol aglycones quercetin, kaempferol and myricetin[64], and strawberry UGT75T1 exhibited very strict substrate specificity and glucosylated only the flavonol galangin out of 33 compounds tested[68]. Thus, different fruit UGTs have obvious preferences for different flavonol aglycones and glycosylation sites.
Hydroxylation at carbon 3, 3’ and 3’5’ positions of flavonols is largely catalyzed by F3H, F3’H and F3’5’H discussed above, and hydroxylation at the carbon 6 and 8 positions is generally performed by flavonol 6-hydroxylase[69] and flavonoid 8-hydroxylase[70] separately. Methylation of flavonols is almost exclusively catalyzed by OMTs, and several fruit OMT genes have been identified that methylate flavonols, for example from apple[71], tomato (Solanum lycopersicon)[72] and citrus[63]. However, no genes encoding enzymes functional in acylation have been verified in fruits so far. Future studies could address this issue and may reveal other target flavonol substrates and new decoration enzymes.
-
The transcriptional control of flavonol biosynthesis genes is often regulated by myeloblastosis (MYB) transcription factors and has been extensively studied in fruits.
MYB genes belong to one of the largest transcription factor (TF) families in plants and modulate a number of different biological processes. In Arabidopsis, MYBs are divided into subgroups (SGs), according to sequence similarity and SG7 group members, including MYB12, MYB11 and MYB111, have been confirmed as flavonol-specific factors[73,74]. In fruits, the SG7 MYBs, which have been identified as activators, have been comprehensively researched in grape, apple, pear, peach and other plants (Table 3). Generally, members of this subclade of MYBs, participate in flavonol accumulation by activating expression of structural genes encoding enzymes in the biosynthetic pathway. For example, apple MdMYB22 binds to the promoter of FLS directly to induce flavonol accumulation[75]. Overexpression of peach PpMYB15 or PpMYBF1[22] or Morella MrMYB12[76] significantly induced the accumulation of flavonols in tobacco flowers. MYBs belonging to other subclasses, including SG4 (flavonoid repressors clade), SG5 (proanthocyanidin-related subclade), SG6 (anthocyanidin-related subclade) are also related to flavonol accumulation (Table 3). Different members of the SG4 subclass have been identified as both inhibitors and activators. For instance, strawberry FaMYB1 was identified as an inhibitor and heterologous expression of FaMYB1 in tobacco resulted in a clear reduction in the levels of quercetin glycosides[77], while apple MdMYB3 was identified as an activator and higher levels of kaempferol and quercetin were observed in transgenic tobacco flowers overexpressing this gene than in wild type plants[78]. MYBs belong to the SG5 and SG6 subclasses have been shown to be activators, such as pear PbMYB9 (SG5)[8] and crabapple McMYB10 (SG6)[80].
Table 3. Summary of MYB and bZIP transcription factors characterized in a wide range of fruit species regulating flavonol accumulation.
Species Genes Metabolites Subgroup Reference Fragaria ananasa FaMYB1 Flavonol, Anthocyanin SG4 [76] Vitis vinifera VvMYB5a Flavonol, Anthocyanin SG6 [84,85] VvMYBF1 Flavonol SG7 [86] VvMYB12 Flavonol SG7 [85] VvibZIPC22 Flavonol, Anthocyanin bZIPC [81] VvMYBA2 Flavonol, Anthocyanin SG6 [87] VvMYB114 Flavonol, Anthocyanin Unknown [82] Malus domestica MdMYB3 Flavonol, Anthocyanin SG4 [78] MdMYB22 Flavonol SG7 [75] Malus crabapple McMYB10 Flavonol, Anthocyanin SG6 [80] MdMYB8 Flavonol Unknown [83] Pyrus bretschneideri PbMYB9 Flavonol, Anthocyanin SG5 [79] PbMYB12b Flavonol SG7 [37] PbMYB17 Flavonol SG7 [36] Prunus persica PpMYB15, PpMYBF1 Flavonol SG7 [22] Morella rubra MrMYB12 Flavonol SG7 [77] Solanum lycopersicum SlMYB12 Flavonol SG7 [88] Citrus sinensis CsMYBF1 Flavonol SG7 [89] In addition, several other transcription factors have been reported to be involved in the regulation of flavonol biosynthesis. The basic region/leucine zipper (bZIP) family transcription factors VvibZIPC22 and VvMYB114 from grape were identified as activators and shown to be involved in transcriptional regulation of flavonol metabolic pathway related genes[81,82]. Similarly, MdMYB8 from crabapple was confirmed as an active regulator of flavonol biosynthesis that activates the MdFLS promoter[83]. In apple, the promoter of FLS was activated by ELONGATED HYPOCOTYL 5 (HY5), which is involved in response to light and could be enhanced by the presence of MYB22[9]. Although studies on transcriptional regulation of flavonols have mostly been focused on MYBs, new regulatory mechanisms affecting the flavonol biosynthetic pathways should be given more attention.
Factors affecting the biosynthesis of flavonols
-
The biosynthesis of flavonols is determined by an intricate system of genetically controlled enzymes and influenced by extrinsic factors such as light in fruit species. Most research has shown that formation of flavonols is significantly accelerated by light. In grape, flavonols were shown to be the most drastically reduced flavonoid compounds following shading and leaf removal treatments, and this was related to VvMYB12-mediated reduction in expression of VvFLS. In contrast, exposure to sunlight substantially induced the accumulation of grape flavonols compared to shading[85]. Similarly, the content of flavonols in peels of apple exposed to sunlight were higher than shaded peels[90]. Further, flavonol accumulation in Cabernet Sauvignon grape was dramatically enhanced by increasing sunlight irradiance and exposure time[91]. However, the level of flavonols can be significantly changed in response to different shade treatments. In crabapple, for example, shading decreased the content of flavonols at 15 days after shading while it increased the level of flavonols at 35 and 50 days after shading[92].
Flavonols are considered as effective UV-absorbing compounds, and are generally induced by UV light, particularly damaging UVB radiation. In grape, supplementing UV with white light treatment drastically increased the accumulation of flavonols by inducing the expression of VvCHS2, VvCHS3, VvCHI1, VvF3H2, VvF3’5’H, VvFLS4, VvMYB12, and VvHY5 genes[93,94]. Conversely, the concentration of grape flavonols was greatly reduced in response to exclusion of UVB[95]. Similarly, lower levels of flavonols occurred in UVB-excluded apples compared to solar UVB-exposed fruits[9]. In several berry fruits such as blueberry[96], grape[97]and strawberry[98], it has been reported that UVC treatment significantly enhanced the content of flavonols.
The accumulation of flavonols in fruits is affected by other abiotic factors. Blackberries treated with methyl jasmonate (0.01 and 0.1 mM) had higher quercetin 3-O-glucoside and quercetin 3-O-rhamnoside content[99]. High medium pH values induced the content of flavonols in crabapple leaves, and this was related to up-regulation of McFLS transcript levels[100]. The plant growth regulator 24-epibrassinolide and 5-aminolevulinic acid up-regulated the expression of the structural gene MdFLS, which was decreased by brassinazole[101]. High nitrogen treatment reduced the overall content of total flavonoids in apple by 19.01%, although kaempferol-3-O-arabinoside increased while quercetin and rhamnetin derivatives decreased[35]. Temperature had little effect on the flavonol content of grape berry skins, although lower temperature (15 °C) increased the content with white and supplementary UV light conditions[93]. In apple, however, lower temperatures (10 °C) inhibited the accumulation of quercetin glycosides compared with 20 °C under both UVB and visible light irradiation[102].
-
To our knowledge, dietary flavonols with potent bioactivity and good biosafety are regarded as natural health metabolites and are derived primarily from fruit sources. Engineering of fruits to enrich for desirable flavonols has recently become the focus of scientific attention. The directed manipulation of target gene expression is regarded as a useful tool to induce the accumulation of flavonol constituents especially in model fruit, such as tomatoes, which are consumed in large volumes. Overexpression of petunia CHI in tomato variety FM6203 produced 16.52 mg g−1 dry weight (DW) quercetin and 2.05 mg g−1 DW kaempferol, indicating increases of 66- and 57-fold over control peel extracts, respectively[103]. Subsequently, Luo et al.[46] introduced AtMYB12 into the tomato MicroTom and Money Maker background separately and the contents of flavonols in transgenic fruits were increased to 72 mg g−1 DW and 48 mg g−1 DW on a whole-fruit basis, representing increases of up to 65-fold compared to control fruits. Based on the AtMYB12 mediated genetic background, a crossed phenotype termed Indigo (anthocyanin-enrich Del/Ros1 parent × flavonol-enrich AtMYB12 parent) tomato had even greater content of flavonols in fruits, approximately 3-fold more than parental AtMYB12 tomatoes[104]. In addition, introducing AtMYB11 into tomato resulted in increased flavonol levels in fruit peels but showed a smaller effect on flavonols compared to AtMYB12[48]. With the continuous development and improvement of experimental technology, the prospects of enhancing the accumulation of flavonols in non-model fruits by altering transcript levels of genes related to flavonol metabolic pathway looks promising. Overexpression of either MdMYB22 or MdMYB8 in 'Orin' apple callus significantly promoted flavonol accumulation[75,83]. The concentrations of most flavonol metabolites were up-regulated by overexpressing PbMYB12b in pear fruits, except for quercetin 3-O-arabinoside[37].
Biotechnological production of flavonol compounds using microorganisms could possibly meet the increasing market demand for fruit flavonols. For instance, vectors containing citrus F3H and FLS genes were introduced into E. coli resulting in production of 15.1 mg L−1 kaempferol with tyrosine supplement and 1.1 mg L−1 galangin with phenylalanine supplement[105]. Fisetin has also been produced at a concentration of 0.3 mg L−1 by overexpressing flavonol biosynthesis-related genes in E. coli with 0.5 mM L-tyrosine supplement[57]. In recent years, de novo production of kaempferol, myricetin, quercetin using the actinomycete Streptomyces coelicolor, and fisetin in the host yeast Saccharomyces cerevisiae grown on a cheap carbon source has been described[106].
-
Flavonols with their extensive double bonds and polyphenolic nature are important secondary metabolites and have diverse functions in animals, plants, and microorganisms. In this article, we have attempted to summarize recent advances in the understanding of the structure, distribution, biosynthesis, regulation and metabolic engineering of fruit flavonols. Recent development in metabolomics, particularly the widespread adoption of high-resolution mass spectrometry, have considerably improved detection and identification of flavonol metabolites. The increasing development of functional genomics and transcriptomics and improvement of experimental systems for modifying gene expression have given a significant boost to studies on the biosynthesis, regulatory mechanisms and modification of flavonol content. Whilst most research on regulation of flavonol production to date has focused on MYB transcription factors, there is a need to better understand how environmental and stress responses affect the production of flavonols and identify other participating transcription factors. Furthering our understanding of the factors affecting the structure, accumulation and distribution of fruit flavonols will facilitate production of metabolically engineered plants containing desirable bioactive compounds and promote consumption of healthier fruit.
- This work was supported by the Key Research and Development Program of Zhejiang Province (2021C02001), the Key Project for New Variety Breeding in Agriculture of Zhejiang Province (2021C02066-3), the National Natural Science Foundation of China (31872067), the 111 project (B17039), and the Fundamental Research Funds for the Central Universities.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xing M, Cao Y, Grierson D, Sun C, Li X. 2021. The chemistry, distribution, and metabolic modifications of fruit flavonols. Fruit Research 1: 11 doi: 10.48130/FruRes-2021-0011

The chemistry, distribution, and metabolic modifications of fruit flavonols
- Received: 18 September 2021
- Accepted: 18 October 2021
- Published online: 28 October 2021
Abstract: Fruits are considered as healthy foods because they provide a rich source of vitamins, antioxidants and other nutrients, including a range of essential bioactive flavonoid compounds. Flavonols, with diverse chemical properties and biological activities, are the most ubiquitous flavonoids that occur naturally in fruits and they are nutritionally important to animals and humans. Numerous investigations have emphasized that significant intake of dietary flavonols is associated with lower incidences of degenerative diseases. Here, we review current knowledge concerning the molecular structures, composition and distribution, regulation, and structural modification of fruit flavonols. In addition, we consider biotechnological approaches to enhance the levels of flavonols in plants or microorganism. An understanding of the factors determining production of flavonols in fruit crops will improve breeding programs and facilitate the production of fruits or bio-products with desirable contents of bioactive flavonols of benefit to humans.
-
Key words:
- Fruit /
- Flavonols /
- Chemistry /
- Metabolite modification /
- Biotechnology