









-
Rice (Oryza sativa L.
) is a staple food for more than half the world’s population. Its production is important for food security worldwide. However, rice yield is largely limited by disease, such as bacterial blight and fungal blast. These diseases are caused by Xanthomonas oryzae pv. oryzae (Xoo) and Magnaporthe oryzae (M. oryzae), respectively, both of which are currently the leading causes of rice crop loss worldwide[1]. Increasing rice yield is a major challenge for modern agriculture, and maximizing disease-resistance while maintaining high yield remains difficult[2]. Understanding of the molecular mechanisms underlying the infection of the two pathogens will benefit the genetic breeding for disease-resistant and high-yield rice crops. Plants have evolved many receptor-like cytoplasmic kinases (RLCKs) to cope with the constant challenges of biotic and abiotic stresses. The RLCKs contain an intracellular kinase domain but lack extracellular and transmembrane domains[3], however, some RLCKs can anchor to the plasma membrane through N-terminal palmitoylation and/or myristoylation motifs[4,5]. Most RLCKs are active downstream of the receptor-like kinases (RLKs) and receptor-like proteins (RLPs), and play crucial roles in innate immunity and hormone signaling[6−8]. In Arabidopsis, after flagellin is recognized, the RLCK VII subgroup member Botrytis-induced kinase1 (BIK1) is phosphorylated by the FLS2/BAK1 complex and then dissociates from the complex to activate the downstream MAPK cascade response[9,10]. Other RLCKs, such as PBS1 and PBL1 (PBS1-like 1), play redundant roles with BIK1 in pathogen-associated molecular pattern-triggered immunity (PAMP-triggered immunity, PTI)[9,11]. Similarly, in pepper and tomato, the receptor-like cytoplasmic protein kinase1 (CaPIK1) and tomato protein kinase 1b (TPK1b) are also involved in the basal resistance to various pathogens[12,13].
Several RLCKs have also been found to be active in effector-triggered immunity (ETI) response. For example, PBS1 and RPM1-induced protein kinase (RIPK), both subgroup VII RLCKs, function as targets of bacterial type III effectors[14,15]. PBS1 can be cleaved by Pseudomonas syringae effector AvrPphB, and this cleavage is crucial for the activation of RPS5-mediated ETI responses[14,16−18]. RIPK is targeted by at least three effectors, AvrB, AvrRpm1, and XopAC, leading to activation of RPM1-mediated ETI responses[15,19]. Similarly, Pto, a RLCK in tomato, can confer race-specific ETI resistance to P. syringae by interacting with AvrPto or AvrPtoB[20]. In addition, an abundance of evidence suggests that RLCKs play various roles in brassinolide (BR), salicylic acid (SA), jasmonic acid (JA) and ethylene (ETH) mediated signaling, self-incompatibility, and modulating various plant growth and development processes[4−6,21−26].
The monocot rice genome encodes 379 members of RLCKs, and RLCK genes are distributed across 12 chromosomes[21]. A limited number of RLCK genes have been functionally characterized, mainly focusing on subgroup 34. OsRLCK185 serves as a bridge connection between the chitin receptor OsCEBiP and the MAPK cascade after chitin perception. OsRLCK185 interacts with the pattern recognition receptor OsCERK1 and is phosphorylated by the OsCERK1[27]. The phosphorylation-activated OsRLCK185 transmits a signal to several MAPKKKs including OsMAPKKKε, OsMAPKKK11, and OsMAPKKK18 to activate immune signaling[28,29]. In addition, OsRLCK185 interacts with the cyclic nucleotide-gated channel protein OsCNGC9 and then phosphorylates OsCNGC9 to activate the channel activity, leading to calcium influx, accumulation of ROS, and expression of downstream defense genes[30].
Similar to OsRLCK185, OsRLCK176 interacts with OsCERK1 in response to chitin and peptidoglycan[31]. OsRLCK176 acts downstream of the monocot receptor-like kinase SPL11 cell-death suppressor 2 (SDS2) and induces plant immunity by transmitting signals to OsRbohB, subsequently activating ROS production and programmed cell death[32]. OsRLCK57, OsRLCK107, and OsRLCK176, which also belong to subgroup 34, positively regulate immune response by altering the expression of Xa21 but negatively regulate brassinosteroid signaling and influence leaf angle, tillering, and seed set rate[7]. The rice BSR1 (OsRLCK278) also belongs to subgroup 34. OsRLCK278 positively regulates resistance against Xoo and M. oryzae in rice[33,34]. OsRLCK55 and OsRLCK185 function redundantly in the ETI immune response targeted by Xoo effector Xoo1488[27]. Subgroup 34 is the largest subgroup in the rice RLCK family and has around 54 members[21]; however, the function of most RLCKs in this subgroup remains uncharacterized.
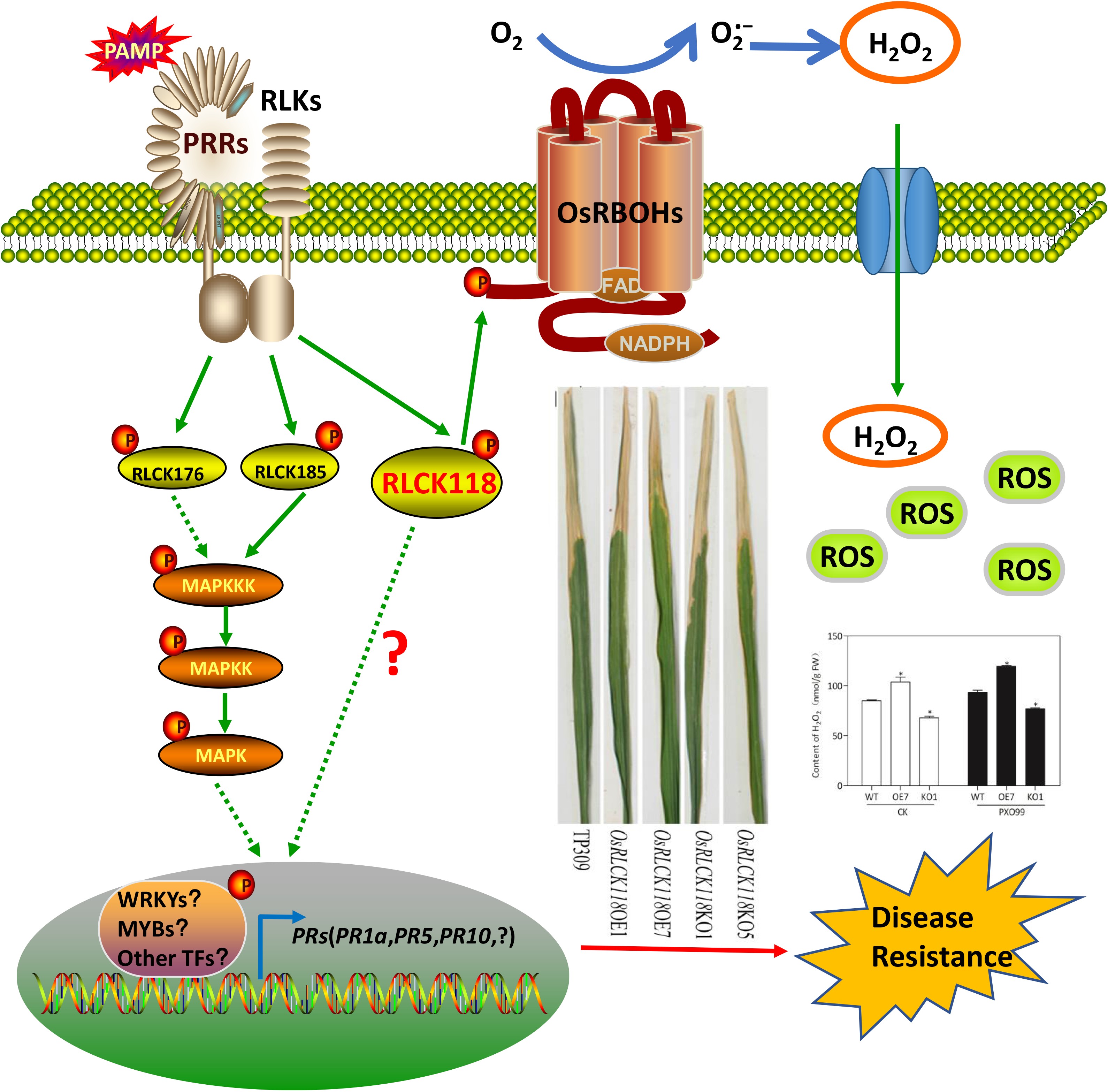
Here, we performed a functional characterization of kinase OsRLCK118, a member of subgroup 34, in rice. We show that OsRLCK118 regulates plant growth and development in terms of shoot length, plant height and leaf angle. In addition, OsRLCK118 is essential for disease resistance to bacterial blight as well as fungal blast. These results provide a new insight into the role of the OsRLCKs in rice development and immunity.
-
After infection with bacterial blight pathogens PXO68 and PXO99, we analyzed the expression patterns of OsRLCK118 in japonica rice variety, Dongjin (DJ), at 0, 12, and 24 h post-inoculation. Transcription levels of OsRLCK118 were remarkably activated at 12 h post-inoculation, and then slightly declined at 24 h (Fig. 1a), suggesting that OsRLCK118 may respond to biotic stress.
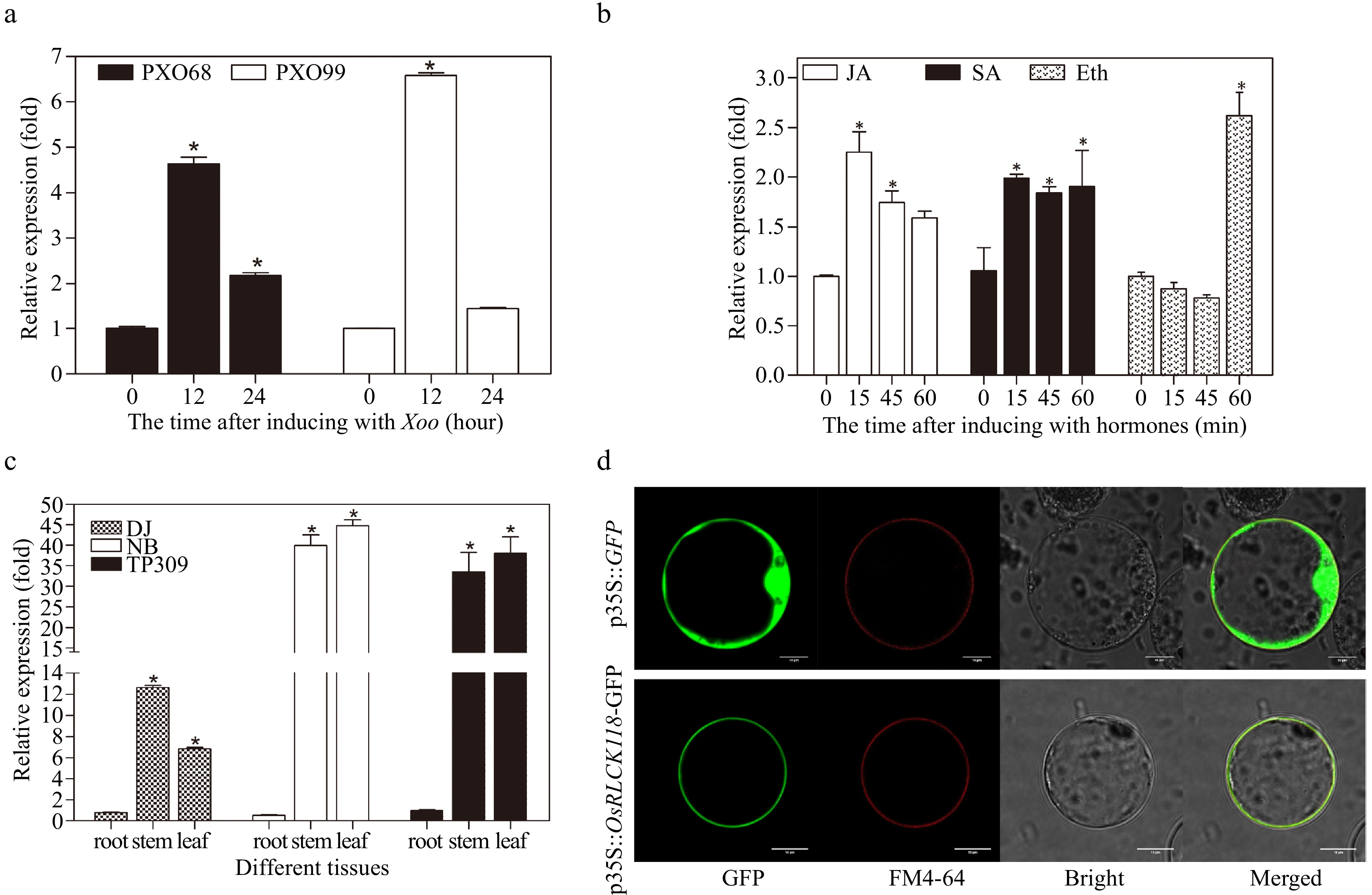
Figure 1.
Expression pattern and subcellular localization of the receptor-like cytoplasmic kinase OsRLCK118. (a) Transcriptional levels of OsRLCK118 in plants inoculated with pathogens Xoo strains PXO68 and PXO99 via qRT-PCR. (b) Expression of OsRLCK118 in plants sprayed with JA, SA and ETH, respectively. (c) Relative expression levels of OsRLCK118 in root, stem and leaf of TP309, DJ and NB rice plants. Relative expression levels of OsRLCK118 in treatments are compared against control plants treated with water. Relative expression levels of OsRLCK118 were characterized by normalization to reference GAPDH gene. Three biological replicates were performed. Error bars represent standard deviation (SD). Asterisks indicate significant differences (P < 0.05) by one-way ANOVA followed by Tukey HSD. (d) Subcellular localization of OsRLCK118 fused with green fluorescent protein (GFP) in rice protoplast. Naked-GFP-expressing construct (35S::GFP) was used as control. The fluorescence signals were detected under a Laser confocal microscopy (Leica Microsystems, Wetzlar, Germany).
We next examined the expression patterns of OsRLCK118 in DJ treated with three plant defense-related hormones at 0, 15, 45, and 60 min after spraying. The transcription levels of OsRLCK118 were all increased when treated with SA, JA and ETH. OsRLCK118 expression in the ETH treatment did not significantly increase before 1 h after spraying. In contrast, peak expression levels of OsRLCK118 occurred earlier in SA and JA treatments (Fig. 1b), suggesting that these plant defense-related hormones can induce expression of OsRLCK118.
OsRLCK118 is ubiquitously expressed and localized in the plasma membrane
-
To investigate the spatiotemporal expression of OsRLCK118, the real-time RT-PCR assays were performed to analyze the expression of OsRLCK118 in different tissues and different rice varieties (Fig. 1c). The result showed that OsRLCK118 was expressed at the higher levels in leaves and stems but at lower levels in roots in DJ. Similar results were also obtained in different rice varieties Nipponbare (NB) and TP309.
To examine the subcellular localization of OsRLCK118, the plasmids 35S::OsRLCK118-GFP and 35S::GFP were transformed into rice protoplasts with incubation of 16 h in the dark at room temperature[35], then the GFP signals were detected with confocal microscopy. OsRLCK118-GFP signals were co-localized with membrane marker FM4-64 to the plasma membrane (PM), whereas control GFP signals were universally distributed across the nucleus, cytoplasm, PM (Fig. 1d). Thus, OsRLCK118 seems to be localized to the PM.
Silencing of OsRLCK118 causes alterations in plant architecture
-
To test whether OsRLCK118 was involved in plant development, we characterized two independent T-DNA insertion mutants, osrlck118-1 and osrlck118-4, in which expression of OsRLCK118 was significantly suppressed (Fig. 2a). Morphological observations showed that the osrlck118 mutant exhibited altered architecture with defects in shoot length, plant height and leaf angle, compared to control plants. The flag leaf angles of the two mutants were 63.5 ± 3.0 and 63.0 ± 2.3 degrees, much wider than the control DJ (51.5 ± 3.4 degrees) (Fig. 2b & c). Osrlck118 mutant plants grew slower than wild-type DJ after white bud spots appeared,resulting in a shorter shoot length in contrast to control wild-type plants. In particular, plant height of the two osrlck118 mutants were 63.7 ± 1.7 and 61.3 ± 2.4 cm, significantly shorter than wild-type at the mature stage (77.5 ± 2.3 cm) (Fig. 2d). Thus, OsRLCK118 drastically affects plant growth patterns in rice.
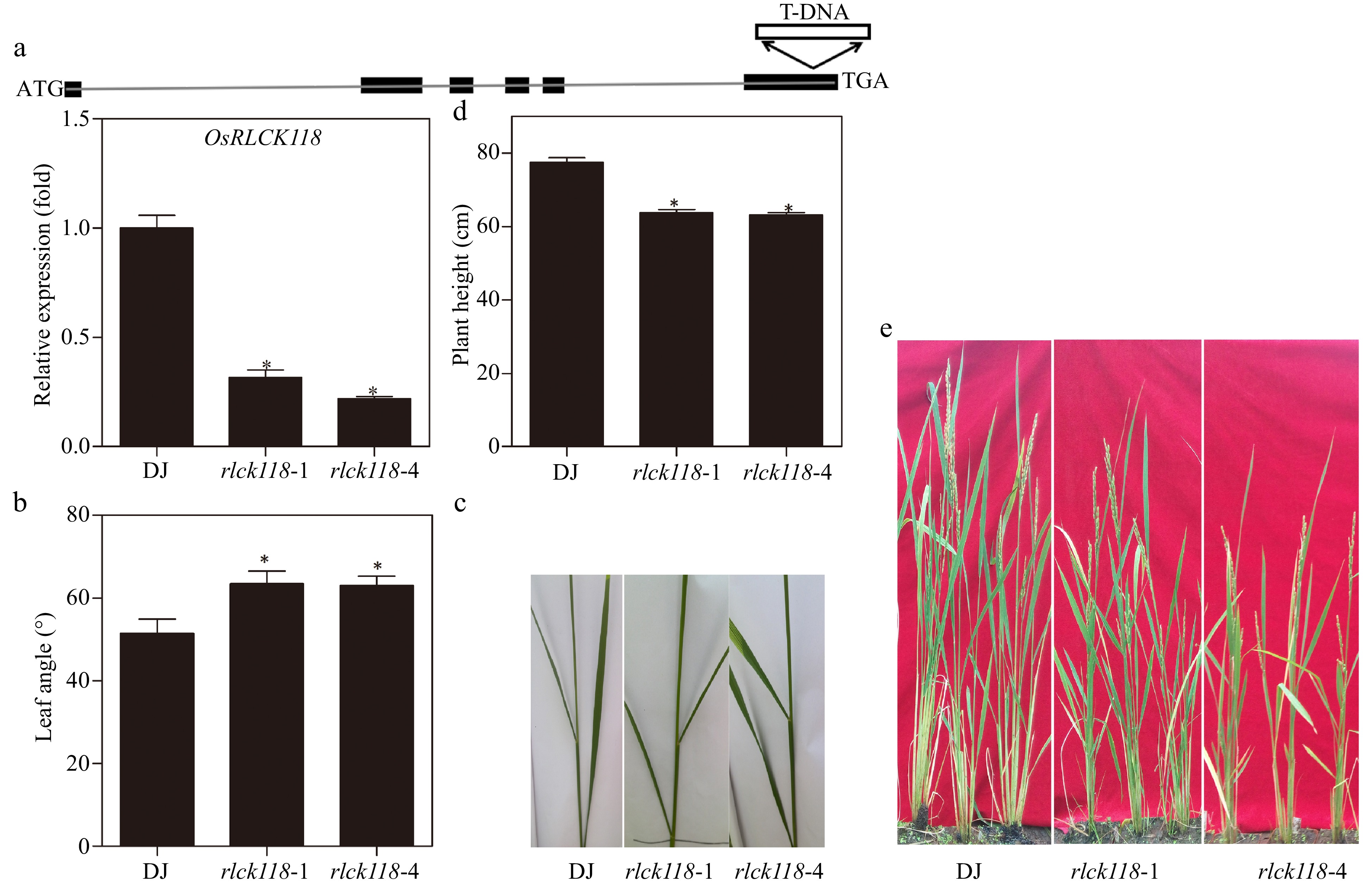
Figure 2.
Heights and leaf angles in OsRLCK118 T-DNA insertion lines. (a) Schematic map of two OsRLCK118 T-DNA insertion plants. Above: The type of T-DNA insertion mutant. Below: Relative transcription levels of OsRLCK118 in wild-type DJ and two independent T-DNA insertion mutant lines (rlck118-1, rlck118-4). (b) Angles of flag leaf and second top leaf of DJ and two mutant (rlck118-1, rlck118-4) plants. (c) Phenotype of angles in DJ and mutant (rlck118-1, rlck118-4) plants. (d) Plant height of different rice lines measured at mature stages. (e) Growth morphology of DJ and mutant (rlck118-1, rlck118-4) plants at maturity stage. Asterisks indicate significant differences (P < 0.05) compared to wild-type DJ by one-way ANOVA followed by Tukey HSD. Averages and SDs were calculated from 20 leaves of representative rice lines as indicated.
OsRLCK118 can partially rescue functional defect of Arabidopsis Atbik1 mutant
-
AtBIK1 was shown to be necessary for flg22 triggered PTI signaling[9]. It is well known that the expression of FRK1 was induced by flg22 and FRK1 was used as a reporter gene in PTI[36]. To investigate whether the OsRLCK118 shares similar function with the Arabidopsis AtBIK1, the 35S::OsRLCK118-Flag construct vector, FRK1::LUC and 35S::RLUC were transiently co-transformed into the leaf protoplasts of Arabidopsis wild type Col-0 and atbik1 mutant for luciferase reporter assay. The LUC activities in Col-0, atbik1, Col-0/OsRLCK118, atbik1/OsRLCK118 were 4.7 ± 0.6, 1.2 ± 0.1, 12.8 ± 0.5 and 3.9 ± 0.1, respectively (Fig. 3a). The results show that OsRLCK118 rescues the functional defect of Arabidopsis atbik1 after treated with flg22 and positively regulates the flg22-triggered immunity, indicating that OsRLCK118 is functionally conserved in plants.

Figure 3.
Basal disease resistance of OsRLCK118 in Arabidopsis and rice. (a) FRK1pro::LUC assay induced by flg22 in Arabidopsis protoplasts. Above: The FRK1::LUC assay using induction of flg22 treatment for 3 h in Arabidopsis protoplasts. Protoplasts prepared from Col-0 and bik1 leaves were co-transfected with/without 35S::OsRLCK118-FLAG together with FRK1::LUC and 35S::RLUC plasmids. Below: Western blot assay, showing the expression level of OsRLCK118 protein fused with Flag-tag Peptide in protoplasts. Anti-Flag, Anti-Flag antibody; CCB, Coomassie blue staining; Col-0, wild-type; bik1, Arabidopsis bik1 mutant; bik1+OsRLCK118, transiently overexpression of OsRLCK118 in the bik1 protoplasts; Col-0+OsRLCK118, transiently overexpression of OsRLCK118 in wild-type Col-0 protoplasts. flg22/mock means that the ratio of LUC activities after treated with flg22 and water. Asterisks indicate significant differences (P < 0.05) compared to wild-type Col-0 by one-way ANOVA followed by Tukey HSD. (b) Lesion lengths and areas of different rice lines after inoculation with M. orzae Y34. Lesion lengths (left Y-axis) and areas (right Y-axis) were measured with Image J software at day 7 post-inoculation with Y34. (c) Lesion phenotype on representative leaves from DJ and mutant (rlck118-1, rlck118-4) at day 7 post-inoculation with Y34. (d) Lesion lengths of different rice lines after inoculation with Xoo strains PXO68 and PXO99. Lesion lengths were measured at day 14 post-inoculation. (e) Lesion lengths on representative leaves from DJ and T-DNA insertion mutant (rlck118-1, rlck118-4) at day 14 post-inoculation with PXO68 and PXO99. Asterisks indicate significant differences (P < 0.05) compared to wild-type DJ by one-way ANOVA followed by Tukey HSD.
Silencing of OsRLCK118 increases susceptibility to bacterial blight and fungal blast in rice
-
To test whether OsRLCK118 participated in rice immunity, we inoculated two T-DNA insertion mutants (osrlck118-1; osrlck118-4) with blast fungal strain Y34. Osrlck118 exhibited increased susceptibility to blast fungus Y34 and showed larger lesions than wild-type DJ (Fig 3b & c). Similar results were obtained when inoculated with Xoo strains PXO99 and PXO68 via leaf-cutting. The lesion length in osrlck118 was ~14 cm, which was longer than that in the control plants DJ at 14 days post-infection (Fig 3d & e). The results showed that OsRLCK118 may positively regulate rice disease resistance.
Overexpression and knockout of OsRLCK118 validates its positive role in resistance to bacterial blight in rice
-
To confirm the function of OsRLCK118 in rice disease resistance, we produced the OsRLCK118 knock-out (OsRLCK118KO) and overexpressing (OsRLCK118OE) plant lines. For OsRLCK118 knockout, we used the CRISPR/Cas9 technology and chose a 20-nt sequence that specifically targeting the first exon of OsRLCK118. We generated multiple transgenic lines and sequenced the target regions after PCR amplification. OsRLCK118KO1 carries a one-base deletion, whereas OsRLCK118KO5 carries a five-base deletion in the target site (Fig. 4a), both truncating the OsRLCK118 open reading frame. Two independent homozygous lines (OE1 and OE7) with higher transcription levels of OsRLCK118 were selected for disease evaluation (Fig. 4b). As expected, the lesions on the leaves of OE plants were significantly smaller than the leaves of wild-type, whereas OsRLCK118KO lines developed larger lesions than wild-type control (Fig. 4c & d).

Figure 4.
Leaf clipping inoculation of OsRLCK118OE and OsRLCK118KO plants in bacterial blight resistance. (a) Information for the OsRLCK118 knockout plants. (b) Relative expression levels of OsRLCK118 in two overexpressor lines (OsRLCK118OE1, OsRLCK118OE7). (c) Lesion lengths were measured at day 14 post-inoculation with PXO99 using the leaf clipping method. (d) Lesion phenotype on representative leaves from wild-type (TP309), OE lines (OsRLCK118OE1, OsRLCK118OE7) and knockout lines (OsRLCK118KO1, OsRLCK118KO5) at day 14 post-inoculation with PXO99. Asterisks indicate significant differences (P < 0.05) compared to wild-type by one-way ANOVA followed by Tukey HSD.
OsRLCK118 affects defense-related gene expression in rice
-
To investigate whether OsRLCK118 regulates the expression of defense-related genes, we measured the expression level of OsNH1, OsPR1a, OsPR10, OsICS1, OsPAL1 and OsRbohE in OsRLCK118-OE7 and osrlck118-KO1 plant lines. The OsICS1 and OsPAL1 genes were reported to encode key enzymes for SA biosynthesis via the isochorismate pathway and the phenylpropanoid pathway[37,38], respectively, however, the transcript level of OsICS1 and OsPAL1 were significantly down-regulated in osrlck118-KO1 line as compared to TP309 (WT) (Fig. 5). OsPR1a, OsRP5 and OsPR10 have been reported to be induced by SA or JA and function in hormone mediated signaling defense response[39−42], our results showed the expression level of OsPR1a, OsRP5 and OsPR10 were significantly lower in osrlck118-KO1 line as well as significantly higher in OsRLCK118-OE7 line compared to TP309 (WT). In addition, the expression level of OsRbohE was also significantly reduced in osrlck118-KO1 line whereas elevated in OsRLCK118-OE7 line, which showed OsRLCK118 might probably alter the production of ROS. Taken together, our results indicate OsRLCK118 is involved in the defense-response via regulating hormone mediated pathogenesis-related (PR) gene expression.

Figure 5.
Relative expression levels of defense-related genes in different rice plants via qRT-PCR. Relative expression levels of NH1, PR5, PR1a, PR10, ICS1, PAL1 and RbohE were measured. WT: wild-type TP309; OsRLCK118OE7: OsRLCK118 overexpressor plant line; OsRLCK118KO1: OsRLCK118 knockout plant line. Asterisks indicate significant differences (P < 0.05) compared to wild-type DJ by one-way ANOVA followed by Tukey HSD. Error bars indicate standard errors for three biological replicates.
OsRLCK118 affected PAMP-triggered ROS burst after bacterial blight treatment
-
To assess the role of OsRLCK118 in the PTI signaling pathway, we characterized PTI-induced ROS responses in OsRLCK118KO1, OsRLCK118OE7, and TP309 (wild-type) plants after inoculation with Xoo. Remarkably, OsRLCK118KO line abolished ROS burst after treatment with Xoo, while the RLCK118OE line increased ROS burst compared with wild-type (Fig. 6).
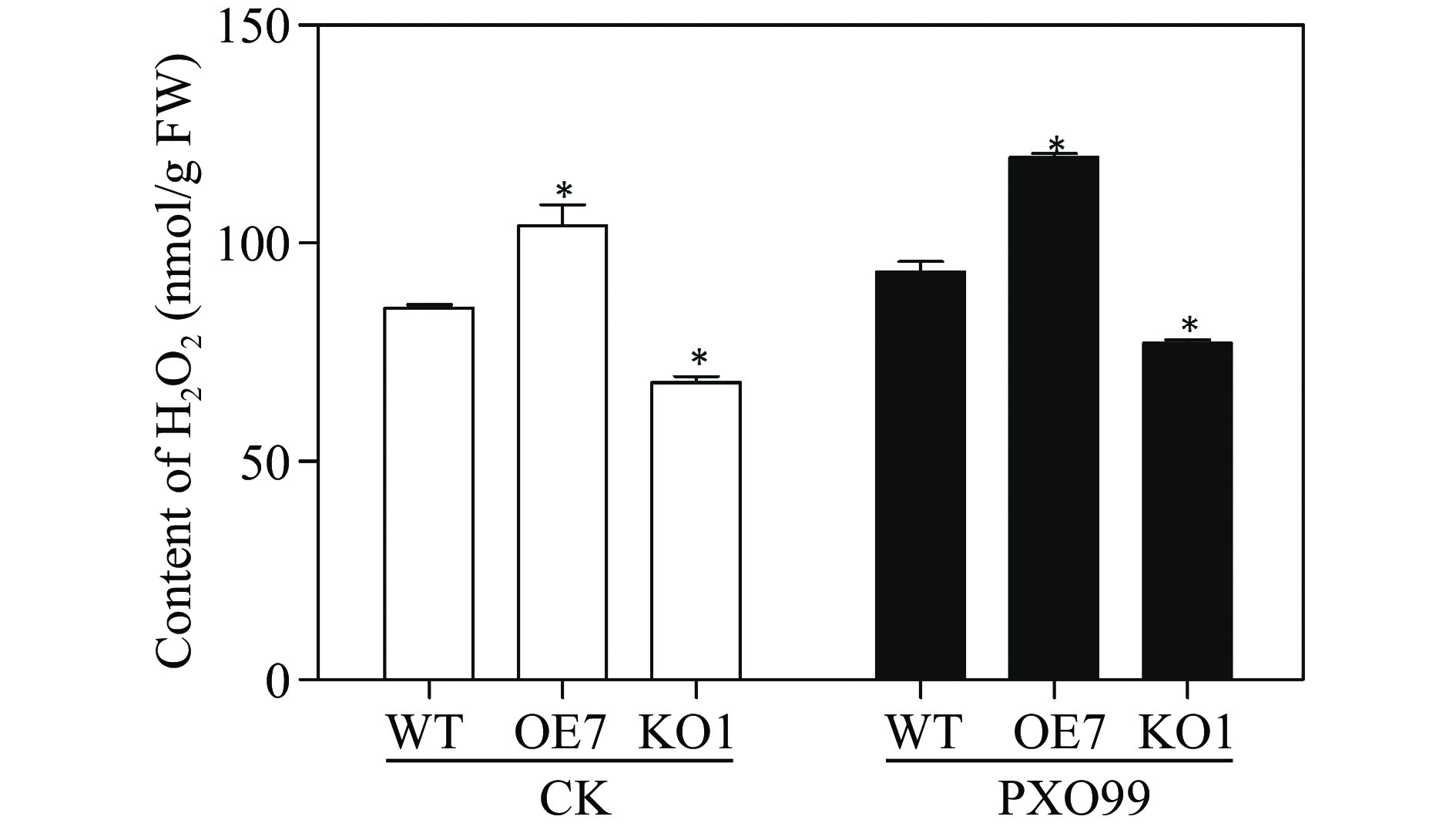
Figure 6.
Measurement of H2O2 in different rice plants inoculated with Xoo. WT: wildtype TP309; OE7: overexpressor plant line OsRLCK118OE7; KO1: knockout plant line OsRLCK118KO1. FW: fresh weight; CK: rice plants inoculated with water; PXO99: rice plants inoculated with Xoo. Asterisks indicate significant differences (P < 0.05) compared to control.
-
The Arabidopsis and rice genomes encode two large families of kinases in plants, which have almost 149 and 379 RLCKs, respectively[3,21]. In rice, the expression levels of 120 RLCKs are significantly changed under pathogen infection. In addition, RNA levels of about 100 OsRLCKs were different across rice growth stages[21]. These results suggest that rice OsRLCKs not only respond to pathogen stimulation but are also involved in many plant developmental processes. However, functions of OsRLCKs in disease resistance and development in rice remain poorly understood.
Transcription levels of four OsRLCK genes (OsRLCK57, OsRLCK107, OsRLCK118, and OsRLCK176) were induced by Xoo in a Xa21-dependent manner, but the transcription levels of these four genes were down-regulated in wild-type Kitaake after treatment with Xoo[7]. In this study, the RNA level of OsRLCK118 significantly increased in wild-type (DJ) after treatment with PXO68 or PXO99, independently (Fig. 1a). In Arabidopsis, BIK1 is required for flg22-mediated immunity in Arabidopsis[11]; furthermore, overexpression of BSR1 can enhance immune response to both Xoo and M. grisea in rice and the response to multiple MAMPs[33,34]. Similarly, in this study, overexpression of OsRLCK118 in Arabidopsis protoplasts could enhance disease resistance to flg22 (Fig. 4a). However, AtBIK1-overexpressed Arabidopsis did not exhibit increases in fungal disease resistance compared to wild-type Col-0 plants[10]. Multi-sequence alignment results showed amino acid differences between OsRLCK118 and AtBIK1 (Supplemental Fig. S1). These results suggest that OsRLCK118 would be functionally different from AtBIK1. Silencing of OsRLCK57, OsRLCK107, OsRLCK102, OsRLCK118, or OsRLCK176 could compromise Xa21-mediated immunity but not the plant basal resistance to Xoo infection[7,43] . However, in this study, we found that silencing OsRLCK118 resulted in more susceptibility to bacterial blight and blast in rice compared to wild-type plants (Fig. 4). Our results imply that OsRLCK118 could modulate the resistance to bacterial and fungal pathogens.
RLCKs also modulate various processes of plant growth and development. In Arabidopsis, knocking-out BIK1 results in serrated leaf margins, wrinkled surfaces, and weakened stem strength, indicating that BIK1 plays an important role in leaf and stem development[10]. In tobacco (Nicotiana tabacum), two RLCK genes (NtPK1 and NtPK2) were involved in pollen germination and pollen tube growth[44]. Other RLCKs, such as BSKs and CDG1 are involved in BR-mediated plant development through interactions with BRI1[25,45,46]. Moreover, reduction in OsRLCK102 expression could alter plant architecture[43]. In this study, mutations in OsRLCK118 caused defects in shoot length, plant height and leaf angle, indicating that OsRLCK118 plays an important role in rice architecture. Our results provide new information for future studies for the regulatory mechanisms of RLCKs that are involved in plant growth and development.
Pathogenesis-related proteins (PR-proteins) function to inhibit pathogen spread and are responsible for immune response in plants. Studies have shown that PR-proteins are related to hormone signaling[47]. For example, phenylalanine ammonia-lyase 1 (PAL1), OsNH1 and OsICS1, which participates in SA synthesis, plays an important role in plant defense[48]. In pepper, CaPAL1 is crucial to plant defense and response to microbial pathogens[49]. In this study, expression of OsPAL1 was significantly decreased in rlck118 mutants, compared to control DJ, suggesting that OsPAL1 may act downstream of RLCK118 affecting its regulation/expression (Fig. 3).
It is well known that, rice PR1a, PR1b, PR5 and PR10/PBZ1 were JA-/ETH-responsive pathogenesis-related (PR) genes[39,50−53] . Expression of defense-related genes, such as PR1 and Ethylene response factor 1 (ERF1), is influenced by BIK1, as demonstrated by their upregulation in the BIK1 mutant[54,55]. In addition, the AtBIK1 not only played positive roles in defense response against fungal and bacterial pathogens but also negatively regulated plant defense against aphids[54]. Meanwhile, PR1 expression positively correlates with resistance to biotrophic pathogens but negatively correlates with resistance to Botrytis in some Arabidopsis mutants[56,57]. Thus, AtBIK1 has a distinct role in plant resistance to different pathogens by affecting the expression of defense-related genes. Constitutive expression of NPR1/NH1 rendered rice plants susceptible to viral infection and hypersensitive to abiotic stresses[58] . The defense strategy of resistance to necrotrophic pathogens is largely distinct from that considered to be effective against biotrophs, which was regulated by SA signaling. While against necrotrophic pathogens, the defense mechanisms in plants is mainly regulated by JA/ETH-dependent signaling routes. Our results showed that RLCK118 mutants were more susceptible to both Xoo and M. oryzae, likely by reducing expression levels of PR1, PR5, PR10, PAL1 and NH1 (NPR1-like gene) (Fig. 5). Interestingly, OsRLCK118 may possess yet unknown complex functions in disease defense and plant development and regulated by hormone-mediated signaling pathway. Nevertheless, more studies are required to further detail the many functions of OsRLCK118.
-
Two Xanthomonas oryzae pv. Oryzae (Xoo) strains, PXO68 and PXO99, were used for bacterial blight inoculation. Magnaporthe oryzae Y34 was used for fungal blast inoculation.
Arabidopsis ecotype Columbia [Col-0, wild-type], atbik1 mutant (Col-0 background), rice cultivar Dongjin (DJ, wild-type), and osrlck118 T-DNA knock-down mutant (DJ background) were purchased from Pohang University of Science and Technology, Korea (
www.postech.ac.kr ).Arabidopsis plants were grown in growth chambers at 22 °C/20 °C, 3000 Lx, 10 h d−1 and 70% room humidity (RH). Rice plants were grown in growth chambers at 28 °C/25 °C, 3000 Lx, 14 h d−1 and 70% relative humidity for hormone treatment and then grown in rice fields for disease resistance assessments.
Hormone treatment
-
The leaves of four-leaf stage seedlings were sprayed with 0.1 mmol/L JA, 1 mmol/L SA, 0.1 mmol/L ABA, and 100 mg/L ETH, respectively. Leaf samples were collected at 0, 15, 30, and 60 min after treatment, frozen in liquid nitrogen, and stored at −80 °C for subsequent analyses. A water treatment was used as a control.
RNA extraction and real time RT-PCR analysis
-
Total RNA was extracted from different frozen rice leaves using Trizol. cDNA synthesis was performed per instructions of the RevertAid First Strand cDNA Synthesis Kit (Thermo Fermentas). Real-time PCR assays were carried out via manufacturer’s instructions for SYBR® Premix Ex TaqTMII (Tli RNaseH Plus) kit (TAKARA, Japan). All primers used in this study are listed in Supplemental Table S1.
Vector construction
-
The first-strand cDNA was diluted ten-fold and then used as a template for the second PCR step. The full length CDS of OsRLCK118 was amplified by PCR using PrimerstarTM DNA polymerase (Takara, Japan). The PCR product was inserted into pCAMBIA35S-4xMyc-MCS-3xFLAG vector to form the OsRLCK118 overexpression construct for rice transformation. For protoplast transformation, the OsRLCK118 was ligated into pUC19-35S-FLAG or pUC19-35S-GFP-RBS, producing 35S::OsRLCK118::FLAG or 35S::OsRLCK118::GFP constructs for protoplast transformation. For FRK::LUC reporter assay, two control vectors FRK::LUC and 35S::RLUC were purchased from Arabidopsis Biological Resource Center (Ohio State University).
CRISPR/Cas9 construction and rice transformation
-
For targeted genome editing of OsRLCK118, the sgRNA (aaggatgggagcccgcaaccggg) in the first exon of the OsRLCK118 gene was used for CRISPR/Cas9 construction[59]. Primers are listed in Supplemental Table S1. Agrobacterium-mediated rice transformation was performed as reported previously[60].
Transient expression in different protoplasts
-
For subcellular localization of OsRLCK118, the rice protoplasts were prepared from cultivated young yellow tissues. Then the resulting construct vector 35S::OsRLCK118::GFP was transferred into rice protoplasts for transiently expression assays using the polyethylene glycol (PEG)-mediated transformation method with incubation of 16 h in the dark, at room temperature[35]. The construct expressing a naked GFP protein was used as a control. FM4-64 plasmid was used as a membrane marker. The GFP fluorescence signals were detected using a Leica Laser confocal microscopy system (Leica Microsystems, Wetzlar, Germany).
For LUC activity analysis, plasmids 35S:: OsRLCK118::Flag, FRK1::LUC, and 35S::RLUC were co-transferred into Arabidopsis wild-type Col-0 and atbik1 mutant protoplasts. Transformed protoplasts were then incubated overnight under light conditions at 22 oC. Protoplasts were treated with either 1 µmol/L flg22 or water (control) for 3 h. LUC activity was determined using the dual-luciferase reporter system per manufacturer’s instructions (Promega, Madison, USA). Bioluminescence was measured by a GLoMax 96 Microplate Luminometer (Promega, Madison, USA).
Pathogen inoculation
-
For bacterial blight inoculation, Xoo strains PXO68 and PXO99 were grown on solid PSA medium [1% (w/v) peptone, 1% (w/v) sucrose, 0.1% (w/v) glutamic acid, 1.5% (w/v) bacto-agar, pH 7.0] for 2 d at 28 °C. Bacteria were collected and suspended in distilled water at OD600 = 0.5−0.6. Fully expanded rice leaves were inoculated via the leaf clipping method[27]. For OsRLCK118 expression assays, leaves were sampled at 0, 12, and 24 h post-inoculation. Samples were immediately frozen in liquid nitrogen and stored at −80 °C. For bacterial blight disease assessment, lesion length was measured two weeks post-inoculation with Xoo. Disease symptoms were photographed.
For blast disease assessment, M. oryzae Y34 was incubated on oatmeal medium [3% (w/v) oat and 1.5% (w/v) Agar] for 5 d at 25 °C. The second top leaves at the four-leaf stage were used for M. oryzae inoculation in vitro using punch inoculation method with slight modification[61]. First, leaves were cut and washed with sterile water. Cuttings were placed face-down on filter paper prewetted with 100 mg/L 6-BA. The ends of the leaf cuttings were fixed with cotton. Then, cuttings were inoculated with fungus colonies of a size that would produce a 0.5 cm diameter perforator. One week post-inoculation, the blast lesion lengths and areas were surveyed using Image J software, and disease symptoms were photographed.
ROS assay
-
The ROS detection method was described previously[62]. Briefly, leaves from 2-month-old plants were inoculated with Xoo by the leaf clipping method[63]. Then 0.1 g samples were extracted with 20 mmol/L phosphate buffer (pH 6.5) after grinded with liquid nitrogen. Using the Amplex Red hydrogen peroxide/peroxidase assay kit (Molecular Probes, USA) to detect the content of hydrogen peroxide. Three replicates were performed for each treatment.
Statistical analysis
-
For each experiment, three biological replicates were performed. Data were presented as means ± standard deviations. All results were subjected to statistical analysis using one-way ANOVA, and significant differences among different lines were identified using T-test (P < 0.05).
This study was supported by the National Natural Science Foundation (31860497) and Natural Science Foundation of Hainan Province (No.2019RC013) and Hainan Provincial Department of Education [Hnjg2019ZD-2]. We also thank Prof. Ye de at China Agricultural University for critical reading of the manuscript, Dr. Chen at South China Agricultural University for useful comments, and Dr. Larry Bowman at Yale University for his assistance with English language and grammatical editing.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (http://www.maxapress.com/article/doi/10.48130/TP-2022-0004)
-
Received 25 May 2022; Accepted 29 June 2022; Published online 25 July 2022
-
OsRLCK118 alters rice architecture
OsRLCK118 positively regulates rice immunity
OsRLCK118 influences the production of reactive oxygen species (ROS)
- supplementary Table S1 Details of primers used in this study.
- Supplemental Fig. S1 Protein sequence visual analysis using TBtools software.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xiao X, Wang R, Guo W, Khaskhali S, Fan R, et al. 2022. The receptor-like cytoplasmic kinase OsRLCK118 regulates plant development and basal immunity in rice (Oryza sativa L.). Tropical Plants 1:4 doi: 10.48130/TP-2022-0004 

The receptor-like cytoplasmic kinase OsRLCK118 regulates plant development and basal immunity in rice (Oryza sativa L.)
- Received: 25 May 2022
- Accepted: 29 June 2022
- Published online: 25 July 2022
Abstract: Receptor-like cytoplasmic kinases (RLCKs), which belong to a large subgroup of receptor-like kinases in plants, play crucial roles in plant development and immunity. However, their functions and regulatory mechanisms in plants remain unclear. Here, we report functional characterization of OsRLCK118 from the OsRLCK34 subgroup in rice (Oryza sativa L.). Expression of OsRLCK118 could be induced by infections with Xanthomonas oryzae pv. oryzae (Xoo) strains PXO68 and PXO99. Silencing of OsRLCK118 altered plant height, flag-leaf angle and second-top-leaf angle. Silencing of OsRLCK118 also resulted in increasing susceptibility to Xoo and Magnaporthe oryzae (M. oryzae) in rice plants. OsRLCK118 knock-out plants were more sensitive to bacterial blight whereas OsRLCK118 overexpressor plants exhibited increased disease resistance. Expression levels of pathogenesis-related genes of OsPAL1, OsNH1, OsICS1, OsPR1a, OsPR5 and OsPR10 were reduced in the rlck118 mutant compared to wild-type rice (Dongjin) and knock-out of OsRLCK118 compromised the production of reactive oxygen species. These results suggest that OsRLCK118 may modulate basal resistance to Xoo and M. oryzae, possibly through regulation of ROS burst and hormone mediated defense signaling pathway.
-
Key words:
- Rice /
- Disease resistance /
- OsRLCK118 /
- Reactive oxygen species burst