










-
Traditional Chinese Medicine (TCM) constitutes an integral component of China's healthcare system and is increasingly recognized worldwide for its therapeutic value. The production and processing of TCM, however, generate approximately 30 million tons of herbal residues annually, of which a significant proportion is disposed of improperly[1]. To address these challenges, various strategies for the comprehensive utilization of TCM herbal residues have been proposed, including the extraction of residual bioactive compounds, the cultivation of edible fungi, the production of animal feed, and agricultural composting[2]. Notably, recent studies have drawn increasing attention to the potential value of herbal residues in improving livestock and poultry production systems. For instance, dietary supplementation with fermented herbal tea residues has demonstrated potential to stabilize physiological functions in goats[3]. Similarly, incorporating herbal residues into the diets of weaned piglets has been associated with improvements in intestinal function[4]. In poultry, supplementing diets with fermented herbal residues has enhanced intestinal barrier functions and modulated inflammatory responses[5]. These findings underscore that herbal residues represent not merely waste, but valuable resources with substantial potential for sustainable agriculture and animal husbandry. Furthermore, their utilization extends the industrial chain of TCM, aligns with circular economy principles, and thereby supports the achievement of sustainable development goals. Consequently, comprehensive further research is imperative to clarify and refine the practical use of herbal residues.
Radix isatidis, commonly known as Banlangen in Chinese, is an annual herbaceous plant in the Cruciferae family[6,7]. Since 1985, the dried roots and leaves of R. Isatidis have been officially listed in the Chinese Pharmacopoeia[6]. More than two hundred constituents have been characterized from this species so far, spanning alkaloids, lignans, organic acids, and various flavonoids[8]. R. Isatidis is known to exert a broad spectrum of biological actions, such as antiviral, antibacterial, anti-inflammatory, antitumor, and immune-regulating effects[6,9]. Analysis revealed that R. Isatidis herbal residue (RIHR) enhances immunity and exerts anti-inflammatory effects in laying hens; these benefits are primarily mediated by key bioactive compounds, including epigoitrin, deoxyvasicinone, 3-indoleacetonitrile, indigo, and indirubin[10]. Notably, epigoitrin has demonstrated efficacy in mitigating inflammation in mice, enhancing immune function, and decreasing susceptibility to the influenza virus[11]. Furthermore, indigo and indirubin, the principal bioactive constituents of indigo, synergistically enhance intestinal barrier integrity and ameliorate ulcerative colitis in mice[12]. The effects of RIHR on broilers, which constitute a major component of global poultry output, have yet to be systematically investigated. Given RIHR's bioactive compounds and their known functions in immunomodulation and inflammation control, its inclusion in broiler diets could enhance productivity and health.
Valorization of herbal residues in animal feed not only enhances waste resource utilization but also advances circular economy principles within the TCM industry. This work examines how RIHR influences growth traits, meat characteristics, and serum indicators in broilers, and further incorporates analyses of the intestinal microbial community, lipid metabolites in the pectoral muscle, and transcriptomic patterns in the ileum to clarify underlying mechanisms. This multi-dimensional approach aims to establish sustainable dietary strategies for enhancing production efficiency and poultry health.
-
The preparation of RIHR and the analysis of its routine nutritional composition, along with the resulting data, were conducted with reference to the methodologies and findings previously established by our research team (Supplementary Tables S1–S3).
Test animals and design
-
In this study, 320 white-feather broiler chickens (Arbor Acres; one-d-old) were randomly allocated to four groups. Each group contained eight replicate pens, with ten chickens in each pen (balanced by sex: five males and five females). The treatments consisted of a control group receiving only the basal diet, and three RIHR-supplemented groups (RIHR-L, RIHR-M, RIHR-H) formulated with 5, 10, and 15 kg of RIHR per ton of feed, respectively. The basal diet fulfilled or surpassed the nutrient recommendations of the NRC (1994), and its ingredients and nutrient profile are summarized (Table 1). The trial lasted for 42 d, during which body weight and feed intake were recorded on days 1, 22, and 43. At the end of the feeding period, blood samples were obtained, and the chickens were slaughtered. Intestinal tissues, chyme, viscera, and muscle samples were then separated and collected for further analysis. The study adhered to the ethical guidelines of 'Replacement, Reduction, and Refinement' to minimize animal use and distress.
Table 1. Ingredients and nutrient levels of the experimental basal diets (on an air-dried basis, %).
Items Diets Phase 1
(d 1−21)Phase 2
(d 22−42)Ingredients Wheat middling and reddog 11.20 10.00 Corn protein powder 4.30 3.00 Soybean oil 2.00 4.95 DL- Met 0.19 0.15 Soybean meal 30.35 27.77 Corn 47.25 49.90 Limestone 1.62 1.49 CaHPO4·2H2O 1.44 1.24 NaCl 0.30 0.30 L- Lys 0.35 0.20 Premix1 1.00 1.00 Total 100.00 100.00 Nutrient levels2 Crude fiber (CF) 3.82 3.60 Met + Cys 0.89 0.79 Calcium (Ca) 1.00 0.90 Crude protion(CP) 21.01 19.01 Metabolizable energy (ME, MJ/kg) 12.14 12.96 Methionine (Met) 0.55 0.47 Lysine (Lys) 1.33 1.13 Total P 0.70 0.65 Available P 0.43 0.38 1 The premix was formulated to supply the complete diet with the following levels of micronutrients (per kg of feed): 12,000 IU of vitamin A, 2,500 IU of vitamin D3, 20 mg of vitamin E, 3 mg of vitamin K3, 3 mg of vitamin B1, 8 mg of vitamin B2, 7 mg of vitamin B6, 0.03 mg of vitamin B12, 20 mg of D-pantothenic acid, 50 mg of nicotinic acid, 0.1 mg of biotin, 1.5 mg of folic acid, 9 mg of copper (from copper sulfate), 110 mg of zinc (from zinc sulfate), 100 mg of iron (from ferrous sulfate), 100 mg of manganese (from manganese sulfate), 0.16 mg of selenium (from sodium selenite), and 0.6 mg of iodine (from potassium iodide). 2 The nutritional level is a calculated value. Growth performance, slaughter metrics, and meat quality assessment
-
Broilers were fasted and repeatedly weighed on days 1 and 43, with cumulative mortality and feed intake documented. Average daily feed intake (ADFI), average daily gain (ADG), and feed conversion ratio (FCR) were derived for the growth phase (days 1-42). The pen (n = 8 per group, 10 birds per pen) was used as the experimental unit. On day 43, four chickens per group with average body weights were selected for slaughter to assess performance indicators (n = 4), including slaughter rate, semi-eviscerated rate, eviscerated rate, and abdominal fat rate, following the Poultry Production Performance Terminology and Measurement Statistics Method (NY/T 823-2004).
Serum biochemical analysis
-
On day 43, after an 8-h fasting period, blood was drawn from the jugular vein in the morning, and isolated serum was used for subsequent biochemical analysis (n = 4). Serum concentrations of glucose (GLU), total protein (TP), low-density lipoprotein cholesterol (LDL-C), triglycerides (TG), total cholesterol (TC), uric acid (UA), high-density lipoprotein cholesterol (HDL-C), albumin (ALB), and the activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were measured using an automatic biochemical analyzer (Excellence 450, Shanghai Kehua). An automatic biochemical analyzer (Excellence 450, Shanghai Kehua) was used to measure the activities of aspartate aminotransferase (AST) and alanine aminotransferase (ALT). Biochemical assay kits were purchased from Kehua Bioengineering Co, Ltd, Shanghai, China.
Enzyme-linked immunosorbent assay (ELISA)
-
Serum concentrations of immunoglobulins IgA, IgG, and IgM were quantified using ELISA kits and processed following the manufacturer's protocol (n = 4). All ELISA kits were purchased from Shanghai Enzyme-linked Biotechnology Co., Ltd, Shanghai, China.
Intestinal microbiota analysis
-
On day 43 of the feeding trial, three healthy broiler chickens, which had body weights near the group average, were chosen from each group for slaughter. Immediately after slaughter, the entire contents of the anterior intestine (duodenum, jejunum) and the distal ileum (the last 5–10 cm segment before the ileocecal junction) and posterior intestine (cecum, colon) were harvested and placed into sterile 10 mL centrifuge tubes. These samples for subsequent analysis were then stored at −80 °C. The microbiota communities in the anterior and posterior intestines were characterized using high-throughput 16S rDNA sequencing (n = 3). Total DNA was obtained directly from the intestinal contents for metagenomic sequencing to provide a detailed profile of the microbial communities. Operational taxonomic units (OTUs) were generated by clustering qualified sequences at a 97% similarity threshold using Uparse algorithm of Vsearch (v2.7.1) software. QIIME (v1.8.0) was employed to generate rarefaction curves and compute diversity indices and richness based on the OTU information, and R (v3.6.0) was used for plotting. PCA was performed in R (v3.6.0) to assess the dissimilarity among samples based on OTU data. The β-Diversity distance matrix between samples was assessed using the Bray-Curtis algorithm. Additionally, the relative abundance of taxa at different levels was determined, and LEfSe analysis was conducted using Python (v2.7).
Transcriptome sequencing of ileum
-
Transcriptome sequencing was performed to examine the effects of RIHR on distal ileum gene expression profiles. Distal ileum samples (the last 5–10 cm segment before the ileocecal junction) were collected from three broiler chickens in both the group showing the optimal effect and the control group, placed in cryovials, and promptly frozen in liquid nitrogen (n = 3). mRNA was purified using oligo(dT)-bound magnetic beads. The isolated mRNA was then fragmented into smaller pieces by incubating with fragment buffer at the recommended temperature. First-strand cDNA was synthesized using random hexamer primers, followed by second-strand synthesis and purification with AMPure XP Beads (Beckman Coulter, USA). Next, A-Tailing Mix and RNA Index Adapters were added to repair the cDNA ends. The resulting cDNA fragments were subsequently amplified via PCR, and the products were purified with AMPure XP Beads (Beckman Coulter, USA) to obtain the final library. Library quality control was performed using an Agilent 2100 Bioanalyzer. Sequencing reads containing adapters, reads with ambiguous bases (N) exceeding 0.5%, and low-quality reads (quality score < Q20) were filtered out to generate clean reads. The resulting clean reads were then aligned to the reference genome using HISAT2 (v2.1.0) while being simultaneously mapped to reference transcript sequences via bowtie2 (v2.3.5). Utilizing RSEM (v1.3.1), read counts mapped to each transcript were quantified based on bowtie2 alignment results, followed by FPKM normalization. DEG analysis was conducted with DESeq2 (v1.22.2) using thresholds of FDR less than 0.05 and absolute |log2FC| greater than 1. For GO enrichment, GOseq (v1.22) was employed with significantly enriched terms defined as q-value less than 0.05. KEGG annotation was performed using KOBAS (v3.0). The pathways (q-value less than 0.05, which had been Benjamini-Hochberg corrected) were identified as significantly enriched through R-based pathway enrichment analysis.
Gene expression level measurement
-
Using the Trizol method, total RNA was extracted from distal ileum tissues (n = 3). Subsequently, cDNA was synthesized using the RT Mix Kit with gDNA Clean for qPCR (Aikerui Biological Company). Detailed information on primers is shown in Table 2. Quantitative PCR was performed using SYBR Green Pro Taq HS qPCR Kit (Aikerui Biological Company, China).
Table 2. Primer sequence information.
Gene name Sequence Accession number β-actin F: ATGATGATATTGCTGCGCTCGT NM_205518.2 R: CCCATACCAACCATCACACCCT FABP1 F: AGAGAGCTTTCCATCATGAGC NM_204192.4 R: TTATAGCAACAGCAAAGAGGTTT ACC F: TCCAGCAGAACCGCATTGACAC NM_205505.2 R: GTATGAGCAGGCAGGACTTGGC SCD1 F: GCCTTCCAGAATGACATCTATG XM_010889078.5 R: AAATCACTCAGGTCCAGCTTC FAS F: AATCTGCCGTCTGGAACTGAATGG XM_032706323. 1 R: CATCCTGTGACTGGTCGTGTTCTC PPARα F: ACCAGCATCCAGTCCTTCATCC XM_063395323.1 R: TTCACAAGCATGTACTCCGTAATGG Lipid metabolomics of breast muscle
-
Breast muscle samples were collected from the same four chickens per group (n = 4) for lipid metabolomic profiling. This study used high-performance liquid chromatography (HPLC) and mass spectrometry (MS) to investigate lipid metabolites of breast muscle. HPLC-grade solvents, including acetonitrile, methanol, isopropanol, and methyl tert-butyl ether, were used for sample preparation, with lipid standards obtained from Sigma-Aldrich (USA) and Avanti Polar Lipids (USA). Tissue samples stored at −80 °C were homogenized and extracted with a methanol-methyl tert-butyl ether solution containing internal standards, followed by reconstitution for LC-MS/MS analysis. The lipid extracts were examined using an LC-ESI-MS/MS system equipped with a Thermo Accucore™ C30 column, using a gradient of acetonitrile, water, and isopropanol as the solvent system. The triple quadrupole-linear ion trap mass spectrometer (QTRAP) was operated in negative and positive ionization modes, with parameters optimized for precise metabolite detection. For the subsequent data processing, principal component analysis (PCA) together with hierarchical cluster analysis (HCA) were conducted, and the analytical outcomes were presented in the form of heatmaps. To determine the differential metabolites, variables with VIP values greater than 1 and p-values below 0.05 were selected based on the OPLS-DA model, with permutation testing to prevent overfitting. The detected metabolites were matched to entries in the KEGG database for functional annotation, and pathway enrichment analysis was performed to determine significant biological pathways through metabolite set enrichment analysis (MSEA).
Data processing
-
Experimental data were organized using Excel 2019. Growth performance, slaughter performance, organ indices, meat quality and serum biochemical data were treated using SPSS 23.0 statistical software. Group-level variation was assessed through the Tukey–Kramer procedure, while the slice-related measurements were examined by applying a one-way ANOVA framework. All results are expressed as 'mean ± the standard error'. Differences were deemed statistically significant at p < 0.05 and highly significant at p < 0.01. Graphs were generated using GraphPad Prism 9.0 software (* indicating p < 0.05 and ** indicating p < 0.01).
-
In Table 3, compared with control group, all dietary RIHR supplementation groups exhibited increased FCR, ADG, and ADFI in broilers, though no significant alterations were observed. These findings suggest that RIHR may enhance growth performance in broilers, while confirming that RIHR supplementation imposes no adverse effects on FCR in 42-d-old broilers under the experimental conditions.
Table 3. Effects of RIHR on the growth performance.
Items Group p-value Control RIHR-L RIHR-M RIHR-H ADG (g/d) 43.38 ± 3.15 45.71 ± 0.83 44.42 ± 2.58 47.93 ± 2.47 0.596 ADFI (g/d) 62.12 ± 3.4 68.34 ± 1.26 72.60 ± 3.71 69.29 ± 1.64 0.126 FCR 1.44 ± 0.04 1.50 ± 0.05 1.64 ± 0.09 1.45 ± 0.04 0.149 Values marked with different lowercase letters within a row differ significantly (p < 0.05), while sharing a letter or having no mark denotes a lack of significant difference (p > 0.05). Effect of RIHR on slaughter performance
-
Table 4 data indicate that, against the control group, all RIHR-supplemented dietary groups showed numerical increases in semi-eviscerated rate, slaughter rate, and eviscerated rate, though these differences lacked statistical significance (p > 0.05). This demonstrates that under the present experimental conditions, dietary RIHR supplementation did not elicit significant improvements in broiler slaughter performance, while confirming the absence of detrimental effects on these parameters.
Table 4. Effect of RIHR on the slaughter performance.
Items Group p-value Control RIHR-L RIHR-M RIHR-H Slaughter rate (%) 92.35 ± 2.13 94.33 ± 0.40 94.51 ± 0.84 93.92 ± 0.68 0.594 Eviscerated rate (%) 74.86 ± 2.48 76.62 ± 0.99 75.13 ± 0.31 75.05 ± 0.90 0.805 Semi-eviscerated rate (%) 86.40 ± 2.18 88.46 ± 0.78 88.20 ± 0.61 87.58 ± 0.34 0.639 Abdominal fat rate (%) 1.22 ± 0.47 1.33 ± 0.32 0.79 ± 0.25 1.37 ± 0.10 0.185 Effect of RIHR on meat quality
-
As presented in Table 5, the dietary supplementation of RIHR elicited no statistically significant differences in most meat quality parameters compared to the control group. However, non-significant trends were observed: compared to the control group, both RIHR-M and RIHR-H groups exhibited reduced cooking loss rates alongside elevated pH24h, a*, b*, and springiness. Although directional patterns in RIHR-mediated modulations of meat textural properties, visual characteristics, and hydration retention were observed, these parameter shifts remained statistically non-significant within the current experimental design.
Table 5. Effect of RIHR on the meat quality.
Items Group p-value Control RIHR-L RIHR-M RIHR-H Shear force (N) 21.47 ± 1.46 26.24 ± 2.47 22.07 ± 5.38 20.64 ± 0.86 0.602 Cooking loss rate (%) 12.48 ± 1.76 12.55 ± 1.64 11.64 ± 1.56 10.26 ± 1.29 0.718 Drip loss rate (%) 0.13 ± 0.10 0.12 ± 0.03 0.12 ± 0.21 0.76 ± 0.19 0.316 pH45min 6.53 ± 0.21 6.21 ± 0.08 6.19 ± 0.11 6.67 ± 0.11 0.127 pH24h 5.71 ± 0.13 5.77 ± 0.99 5.83 ± 0.12 5.80 ± 0.04 0.815 Lightness (L*) 41.13 ± 2.47 41.59 ± 2.79 42.06 ± 0.52 39.85 ± 3.47 0.935 Redness (a*) 2.40 ± 0.36 2.22 ± 1.14 3.38 ± 1.71 3.89 ± 1.71 0.815 Yellowness (b*) 2.83 ± 0.28 2.81 ± 0.38 3.47 ± 0.25 4.00 ± 0.99 0.423 Springiness (mm) 1.89 ± 0.09 1.72 ± 0.11 2.33 ± 0.43 2.19 ± 0.18 0.310 Gumminess (N) 6.27 ± 1.87 6.77 ± 0.98 8.06 ± 1.05 17.01 ± 6.59 0.181 Chewiness (mj) 12.17 ± 3.93 11.40 ± 0.93 18.10 ± 2.40 36.87 ± 14.81 0.150 Effect of RIHR on serum biochemical indicators
-
As presented in Table 6, the RIHR-M and RIHR-H groups significantly or extremely significantly up-regulated serum TP, ALB, A/G, and GLU concentrations, concurrently with reduced LDL-C. These findings indicate that RIHR supplementation may enhance protein anabolism and glucose utilization while improving lipid metabolic health.
Table 6. Effect of RIHR on the serum biochemistry.
Items Group p-value Control RIHR-L RIHR-M RIHR-H IP (mmol/L) 3.15 ± 0.19 3.15 ± 0.98 3.06 ± 0.14 2.91 ± 0.15 0.626 Ca (mmol/L) 2.75 ± 0.10 2.49 ± 0.32 2.64 ± 0.11 2.55 ± 0.99 0.230 ALT (U/L) 4.84 ± 0.97 5.79 ± 1.92 3.35 ± 0.44 3.33 ± 0.56 0.226 AST (U/L) 511.98 ± 44.30 480.54 ± 36.14 510.48 ± 51.33 463.13 ± 36.15 0.814 TP (g/L) 34.65 ±
2.89b43.43 ±
1.08a45.07 ±
3.23a37.87 ±
1.84ab0.019 ALB (g/L) 9.05 ± 2.09b 18.02 ± 0.47a 19.15 ± 0.74a 17.41 ± 0.85a < 0.001 GLB (g/L) 25.43 ± 3.44 25.14 ± 1.06 25.86 ± 2.46 20.57 ± 1.15 0.331 A/G 0.42 ± 0.11b 0.72 ± 0.39a 0.76 ± 0.43a 0.86 ± 0.11a 0.001 GLU (mmol/L) 9.90 ± 0.41b 10.61 ± 0.91b 13.96 ± 0.77a 14.94 ± 0.94a < 0.001 UA(μmol/L) 318.00 ± 24.13 333.71 ± 28.85 422.43 ± 56.35 376.14 ± 61.89 0.392 TC (mmol/L) 3.56 ± 0.13 3.71 ± 0.32 4.43 ± 0.22 3.99 ± 0.23 0.071 TG (mmol/L) 0.81 ± 0.10 0.49 ± 0.05 0.52 ± 0.26 1.45 ± 0.98 0.489 HDL-C (mmol/L) 2.00 ± 0.16 2.40 ± 0.26 2.74 ± 0.22 2.28 ± 0.22 0.154 LDL-C (mmol/L) 2.43 ± 0.32a 0.50 ± 0.67b 0.83 ± 0.52b 0.71 ± 0.73b < 0.001 Note: Different lowercase letters in the same row indicate significant differences (p < 0.05), while the same letters or no letters indicate no significant differences (p > 0.05), the same below. Promotion of immunity by RIHR
-
Figure 1 illustrates the impact of RIHR supplementation on serum immunoglobulin levels in broilers. Compared with the control group, all RIHR-treated groups exhibited extremely significant up-regulations in serum IgA levels (Fig. 1a), while no significant alterations were found in IgM or IgG concentrations.
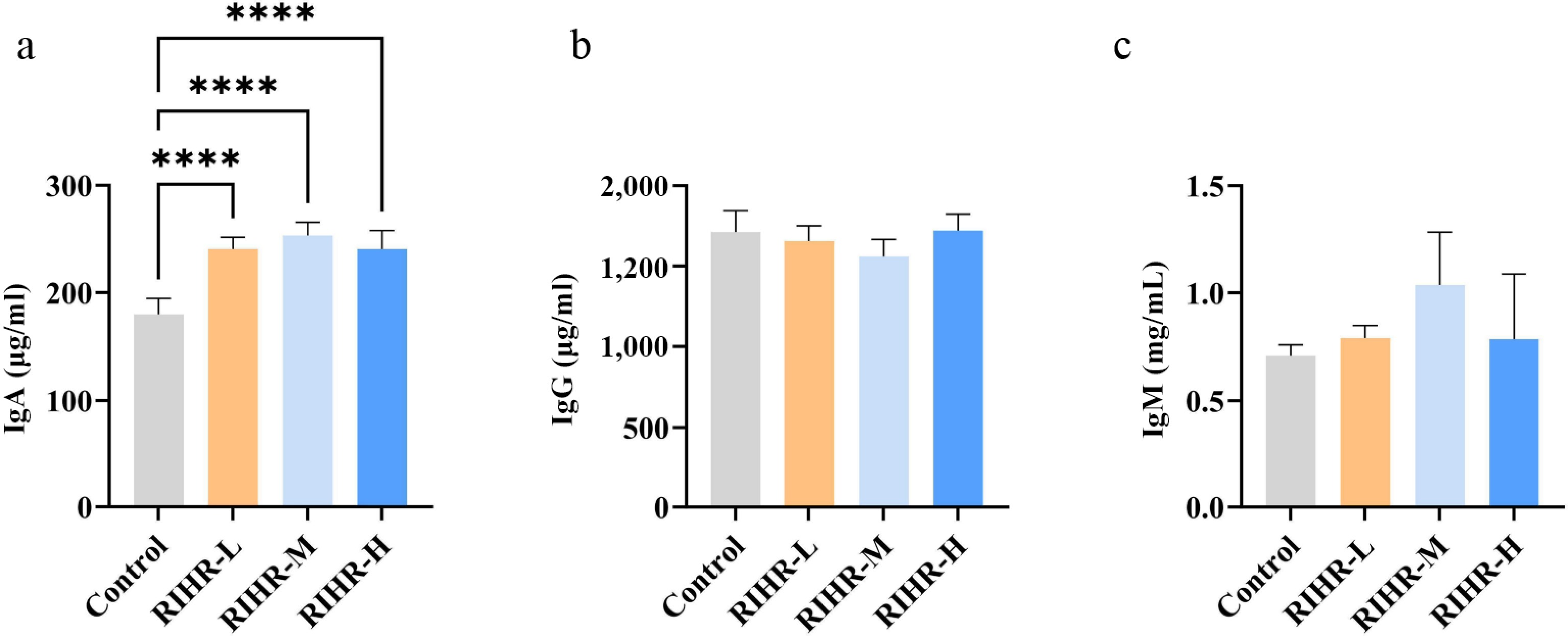
Figure 1.
Effects of RIHR on Immunity. (a)–(c) Measurement of IgA, IgM, and IgG in serum using ELISA. * Indicates significant difference (p < 0.05), **, *** and **** indicate extremely significant differences (p < 0.01), and no * indicates no significant difference (p > 0.05).
Effects of RIHR on the foregut microbiota
-
Our earlier work has demonstrated that Radix isatidis polysaccharides influence physiological outcomes mainly by affecting intestinal microorganisms[10]. Similarly, we examined the effect of RIHR on gut microbiota in broilers. Analysis using Venn diagrams showed that the supplementation of RIHR in the diet resulted in changes to the composition of the foregut microbiota (Fig. 2a). Species richness in all RIHR-treated groups remained comparable to that of the control, with no statistically significant differences detected. (Fig. 2b). The community was primarily composed of Firmicutes, Proteobacteria, Bacteroidota, and Chloroflexi at the phylum level. (Fig. 2c). The abundance of Lachnospiraceae rose across all RIHR groups at the genus level. Notably, the RIHR-H group showed a significant rise in Alistipes compared to the control group (Fig. 2d). The LDA score > 3 as the screening criterion, LEfSe analysis indicated that the control group was characterized by an enrichment of f_Anaerovoracaceae, the RIHR-M group of g_Rothia; and the RIHR-H group of g_norank_f_UCG-010, c_Coriobacteriia, f_UCG-010 (Fig. 2e). Furthermore, significant differences were observed between RIHR and control groups in Lachnospiraceae_NK4A136_group, norank_f_Rikenellaceae, Helicobacter, NK4A214_group, MND1, Hyarogenophaga (Fig. 2f).
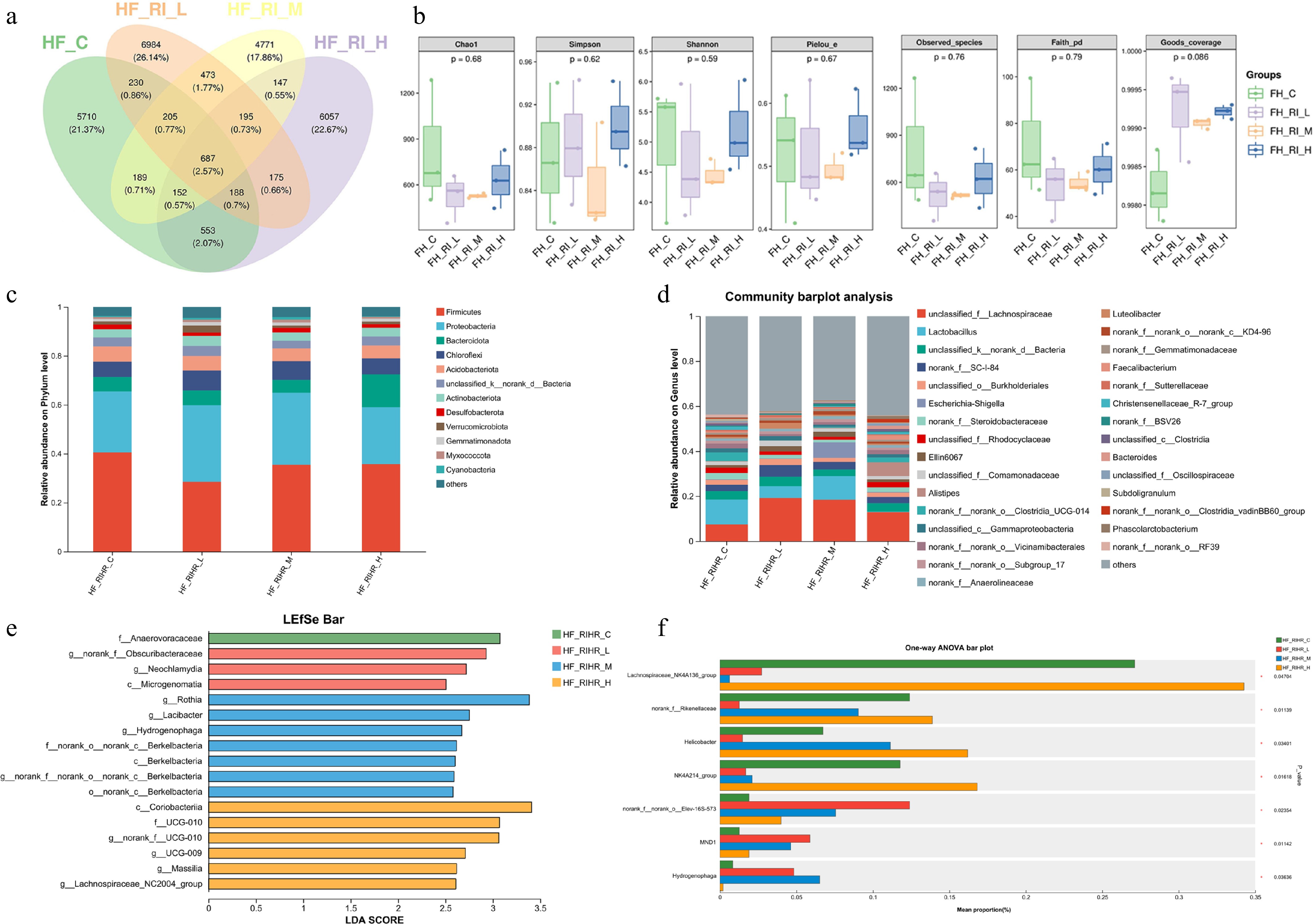
Figure 2.
Effect of RIHR on foregut microflora of 42-day-old broilers. (a) Venn diagram community analysis of foregut microbiota. (b) Analysis of differences in alpha diversity indices among foregut microbiota. (c) Bar chart showing species composition at the phylum level for each group. (d) Bar chart showing species composition at the genus level for each group. (e) LEfSe analysis for multilayer species differentiation. (f) Inter-group species difference analysis. * In the figure indicates a significant difference (p < 0.05), while no * indicates no significant difference (p > 0.05).
Effects of RIHR on the hindgut microbiota
-
The constitution of microbiota in hindgut was changed due to RIHR supplementation, too (Fig. 3a). No significant differences in species richness were detected among the RIHR groups when compared with the control. (Fig. 3b). Although no statistically meaningful differences were observed at the phylum level between the RIHR groups and the control (with Firmicutes and Bacteroidota being dominant), the Firmicutes/Bacteroidota ratio specifically decreased in the RIHR treatment group (Fig. 3c). Genus-level analysis indicated that the RIHR-H group was associated with elevated abundances of Alistipes and Bacteroides (Fig. 3d). Relative to the control group, the RIHR-H group markedly enriched f_Bacteroidaceae, g_Bacteroides, c_Actinobacteria (Fig. 3e). Significant differences were also observed between the RIHR and control groups in norank_f_norank_o_Clostridia_UCG-014 and CHKCI001 (Fig. 3f).
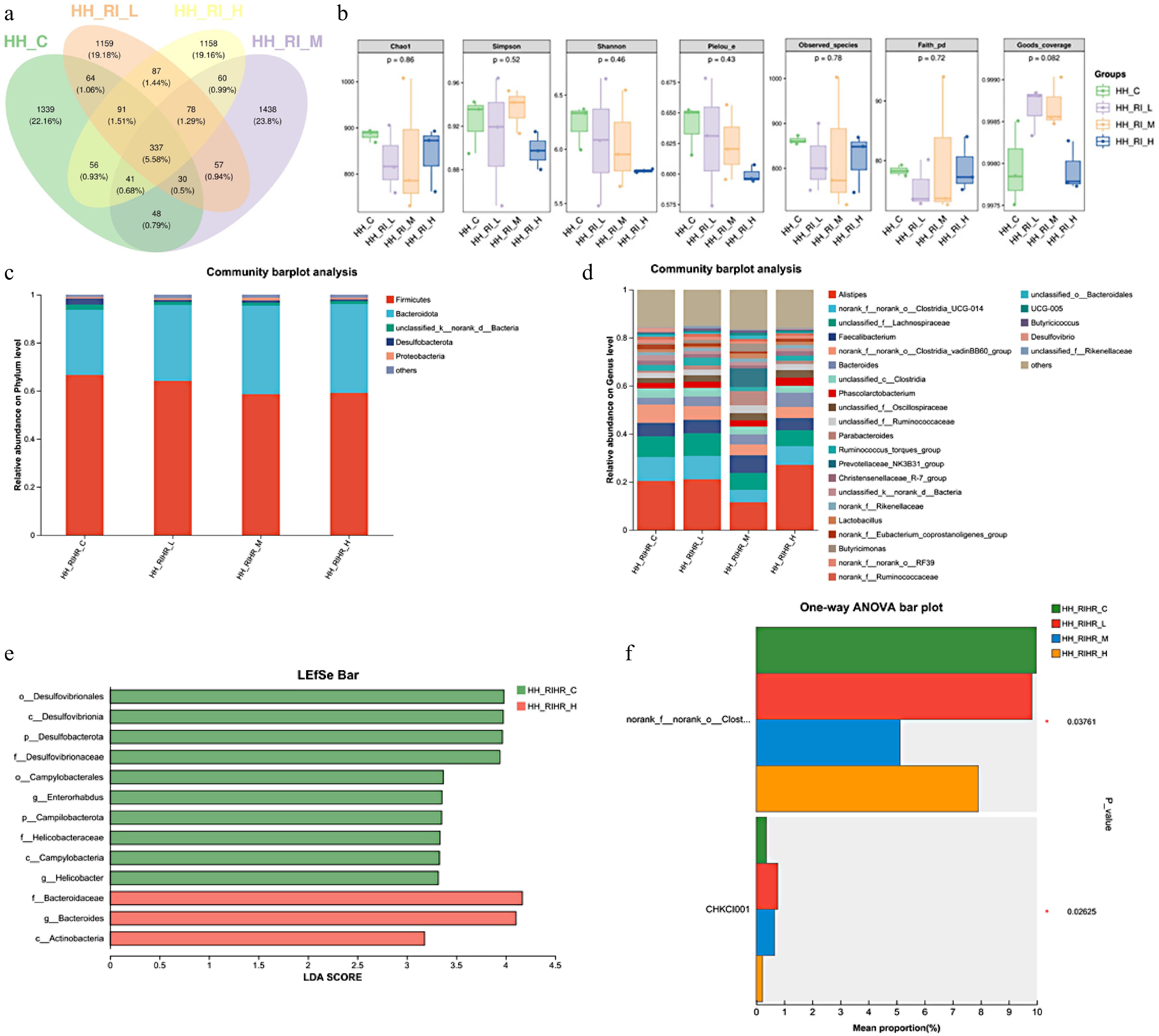
Figure 3.
Effect of RIHR on hindgut microflora of 42-day-old broilers. (a) Venn diagram community analysis of hindgut microbiota. (b) Alpha diversity index analysis of hindgut microbiota among groups. (c) Bar chart of species composition at the phylum level for each group. (d) Bar chart of species composition at the genus level for each group. (e) LEfSe multi-level species differential discrimination analysis. (f) Inter-group species differential analysis. * In the figure indicates a significant difference (p < 0.05), while no * indicates no significant difference (p > 0.05).
Correlation analysis between metabolome and gut microbiota
-
Based on serum biochemical results, correlation analysis was conducted with the microbiota. A significant inverse relationship with TP was detected for Dependentiae at the phylum level (Fig. 4a). At the genus level, Lactobacillus exhibited a significant negative association with HDL-C and ALB, and a significant positive association with ALT (Fig. 4b). At the hindgut phylum level, Desulfobacterota was strongly and positively related to LDL-C and ALT, whereas its relationships with TP, ALB, and A/G were significantly negative. Campilobacterota also demonstrated a notable positive correlation with LDL-C (Fig. 4c). At the genus level in the hindgut, serum HDL-C was significantly positively correlated with Bacteroides and significantly negatively associated with norank_f__Eubacterium_coprostanoligenes_group and Subdoligranulum. Furthermore, analysis revealed that LDL-C was positively linked to Desulfovibrio and Helicobacter, while TP showed significant inverse relationships with Helicobacter and norank_f__norank_o__Clostridia_UCG-014. In addition, ALB and A/G were positively associated with unclassified_f__Ruminococcaceae and negatively associated with Desulfovibrio (Fig. 4d).
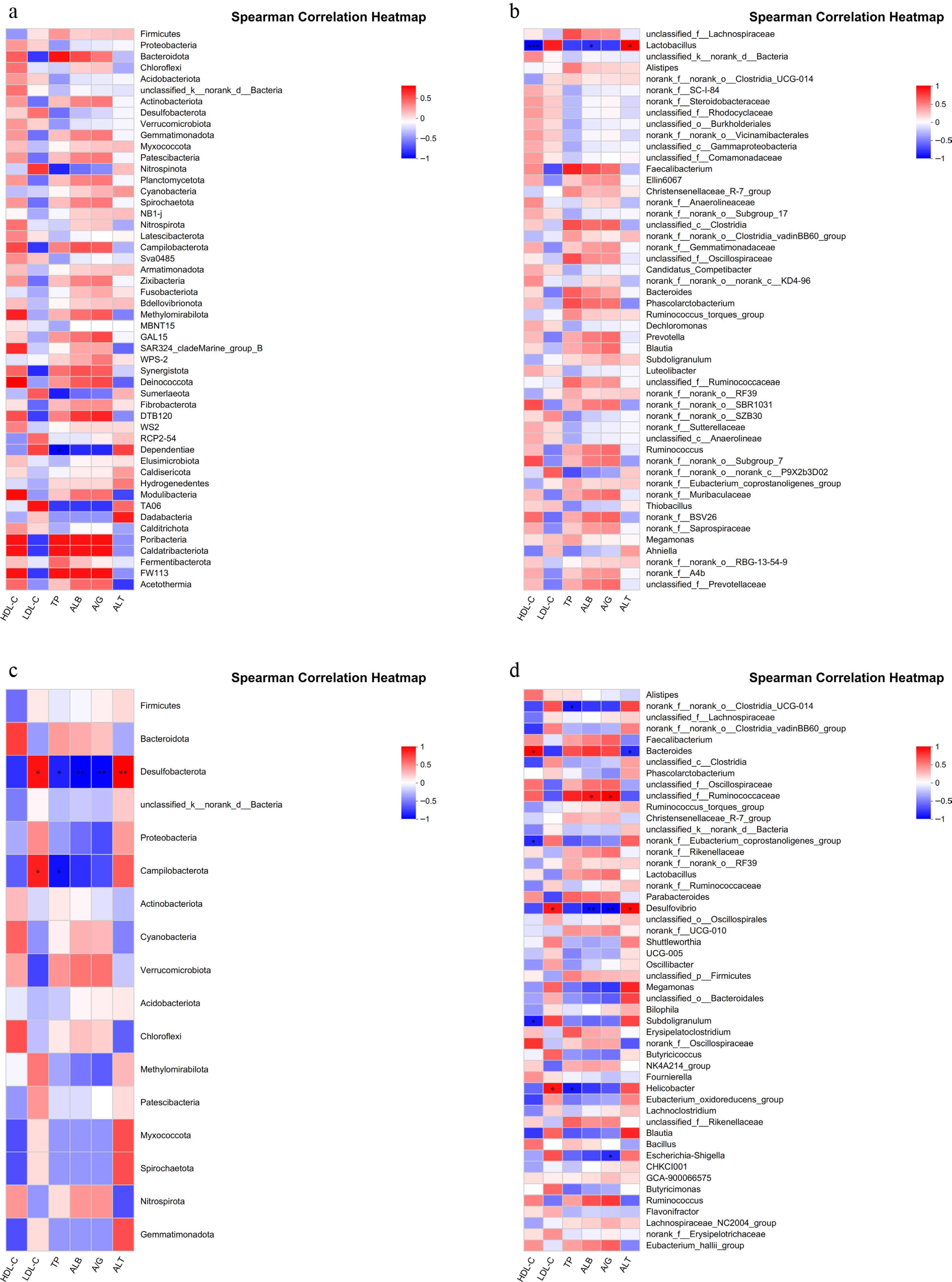
Figure 4.
Correlation analysis of gut microbes with metabolome. (a) Correlation analysis between foregut microbiota at the phylum level and serum biochemistry. (b) Correlation analysis between foregut microbiota at the genus level and serum biochemistry. (c) Correlation analysis between hindgut microbiota at the phylum level and serum biochemistry. (d) Correlation analysis between hindgut microbiota at the genus level and serum biochemistry. * Indicates significant difference (p < 0.05), ** and *** indicate extremely significant differences (p < 0.01).
Transcriptomics of ileum
-
To investigate the regulatory impact of RIHR on gene expression in the ileum of broilers, we performed transcriptomic analysis of ileal tissues. The 3D representation of principal component analysis (PCA) demonstrated that RIHR significantly altered the gene transcript profile in the broiler ileum (Fig. 5a). Furthermore, 673 up-regulated and 691 down-regulated differentially expressed genes (DEGs) were screened (Fig. 5b). Gene Ontology (GO) analysis indicated that the DEGs were significantly enriched in terms related to transmembrane receptor protein tyrosine kinase activity, long-chain fatty acid metabolic process, high-density lipoprotein particle binding (Fig. 5c). Notably, several pathways associated with lipid metabolism, such as the PPAR signaling pathway, fat digestion and absorption, cholesterol metabolism, pyruvate metabolism, glucagon signaling pathway, adipocytokine signaling pathway, fatty acid degradation, and AMPK signaling pathway, were identified. These findings demonstrate that RIHR intervention influences lipid metabolism in broilers (Fig. 5d).
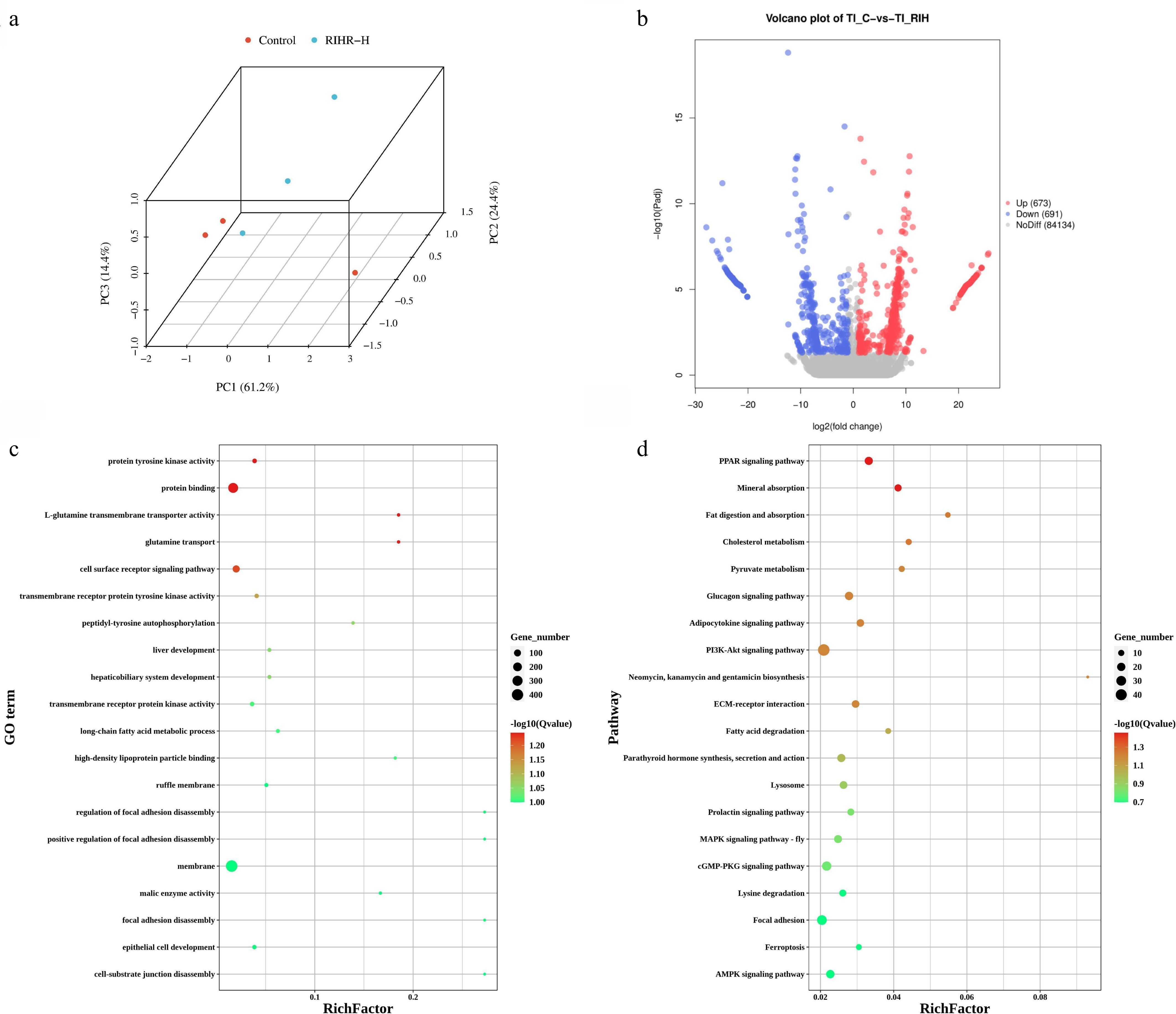
Figure 5.
Transcriptomic analysis of the intestine from 42-day-old broiler chickens. (a) 3D PCA of the ileum from the control (TI-C) and RIHR-H (TI-RIHR) groups. (b) Volcano plot analysis of DEGs isoforms in the ileum from the control (TI-C) and RIHR-H (TI-RIHR) groups. Each point in the volcano plot represents a gene isoform, with blue points indicating downregulated genes, red points indicating upregulated genes, and gray points representing genes with no significant differences. (c) GO enrichment analysis of DEGs in the ileum from the control (TI-C) and RIHR-H (TI-RIHR) groups. (d) KEGG pathway enrichment analysis of DEGs in the ileum from the control (TI-C) and RIHR-H (TI-RIHR) groups.
Improvement of lipid metabolism by RIHR via the PPAR pathway
-
Based on the KEGG enrichment results, several key genes associated with the PPAR signaling pathway in the ileum were identified, including PPARα, FABP1, FAS, ACC, and SCD1. RT-qPCR analysis showed that, relative to the control group, RIHR supplementation reduced the expression of ACC, FAS, and SCD1, and markedly lessened the expression of FABP1. Additionally, PPARα expression was found to increase (Fig. 6a–e).
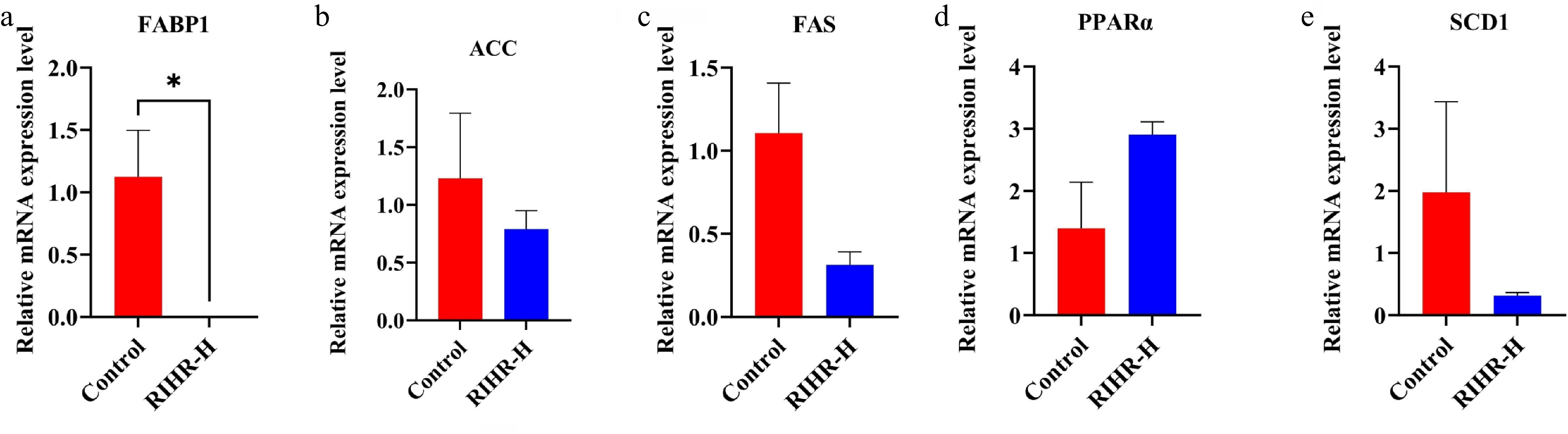
Figure 6.
RT-PCR validation of RIHR on PPAR expression in ileum. (a) FABP1, (b) ACC, (c) FAS, (d) PPARα, and (e) SCD1. * In the figure indicates a significant difference (p < 0.05), while no * indicates no significant difference (p > 0.05).
Lipid metabolomics of breast muscle
-
Transcriptomic analysis of the ileum revealed that RIHR significantly altered the ileal gene expression profile, with DEGs predominantly enriched in lipid metabolism-related pathways. To further elucidate the role of RIHR, we conducted lipid metabolomic profiling of broiler breast muscle. The volcano plot found that compared to the control group, the RIHR group revealed an upregulation of 46 different metabolites (DMs) and a downregulation of 14 DMs in breast muscle (Fig. 7a). The visualization of the DMs (Fig. 7b) revealed that RIHR significantly increased the levels of unsaturated fatty acids, including FFA(22:5), PC(14:0_22:6), PC(14:0_18:3), LNAPE(20:4/N-18:2), PE(20:3_20:4), PE(18:2_22:5). Cluster analysis indicated that RIHR significantly changed the lipid metabolism profile of broiler breast muscle, with obvious intra-group clustering and inter-group differences (Fig. 7c). Analysis of KEGG showed that DMs were mainly enriched in glycerophospholipid metabolism, alpha-linolenic acid metabolism, phosphatidylinositol signaling system, arachidonic acid metabolism, linoleic acid metabolism, glycerlipid metabolism (Fig. 7d).
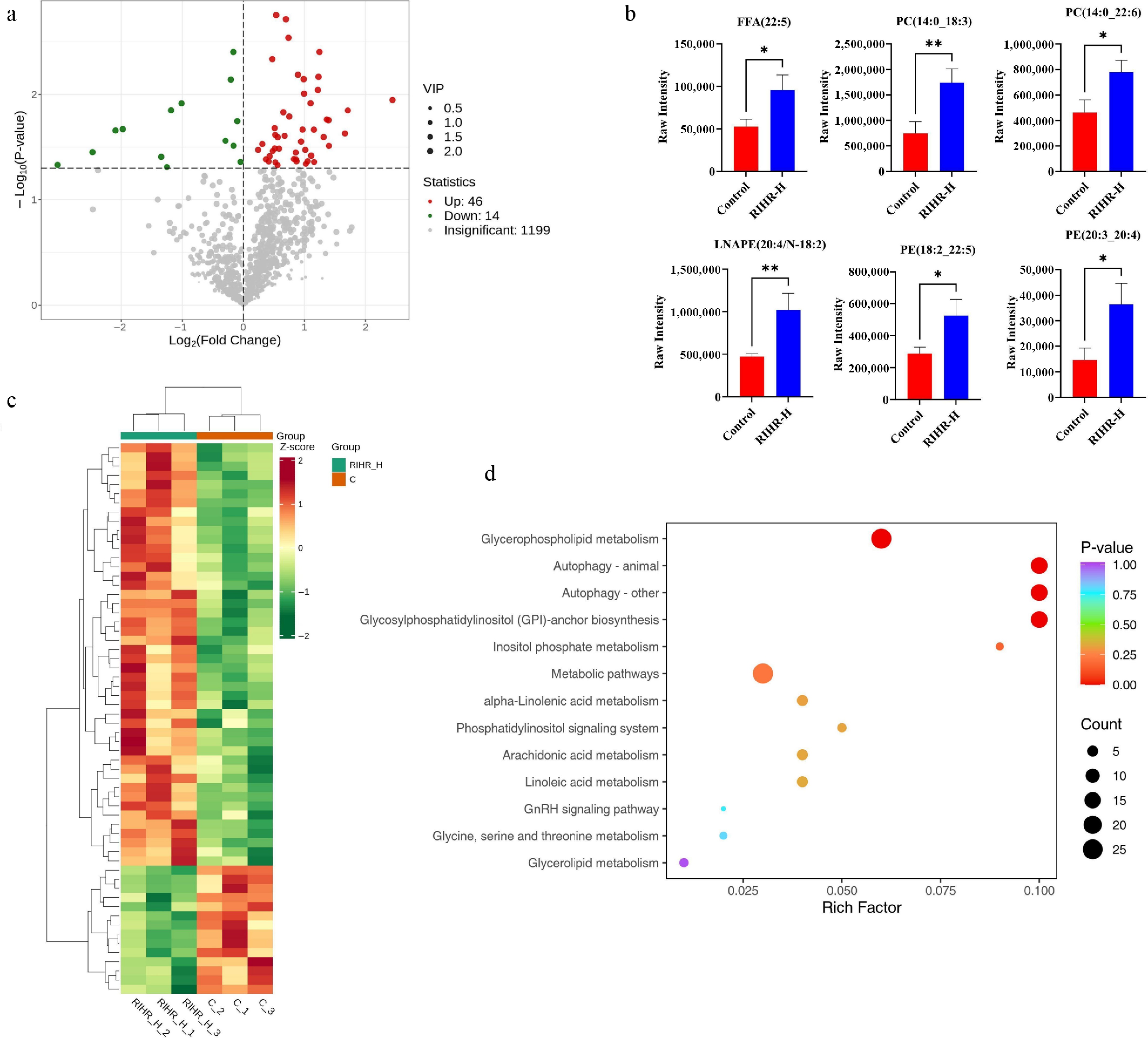
Figure 7.
Metabolomic analysis of breast muscle of broiler chickens at 42 d of age. (a) Volcano plot of differential lipids in breast muscle between the control and RIHR-H groups; green points represent downregulated differential lipids, red points represent upregulated differential lipids, and gray points indicate lipids that were detected but not significantly different. (b) Bar chart of DMs in breast muscle. (c) Cluster heatmap of DMs in breast muscle.(d) KEGG enrichment analysis of DMs in breast muscle.
-
The burgeoning industrialization of TCM, while instrumental in safeguarding human and animal health, generates substantial quantities of herbal residues[13]. The sustainable valorization of these residues is of paramount importance for extending the TCM industrial chain and promoting circular economy principles[14]. Consequently, exploring high-value, resource-efficient utilization pathways for TCM residues, thereby achieving 'waste-to-resource' conversion, constitutes a critical research focus within TCM resource recycling[15]. Furthermore, the comprehensive prohibition of growth-promoting antibiotics in animal feed within China necessitates the urgent identification of suitable antibiotic alternatives for the livestock industry. Compared to raw medicinal herbs, herbal residues, as post-extraction byproducts, offer distinct advantages: their cost is substantially lower, they retain a significant portion of bioactive constituents, and they possess a disrupted lignocellulosic matrix that facilitates the release of active compounds and enhances digestibility and absorption in animals[16,17].
It has been reported that adding Chinese herbal residues to the diet can significantly improve the growth and slaughter performance, meat quality, and immunity of broiler chickens, which is worth further study[5,18,19]. As a byproduct of pharmaceutical extraction, RIHR retains residual pharmaceutical activity. Its dietary supplementation demonstrates dual industry advantages: reducing production costs for TCM manufacturers while enhancing production efficiency in livestock operations[1,20]. The results showed that dietary supplementation with RIHR resulted in numerical increases in ADFI, ADG, and improvements in FCR in broilers aged 1–42 d, although these differences did not reach statistical significance (p > 0.05). These observations are consistent with previous reports suggesting that plant-derived residues can improve nutrient utilization and growth trends in poultry by modulating intestinal digestion and absorption processes[21]. However, further studies with larger sample sizes may be required to confirm these performance benefits. To assess the effects of dietary RIHR supplementation on meat production capacity and quality, we measured key parameters of slaughter performance and meat quality. The data indicated a tendency for dietary RIHR supplementation to improve broiler slaughter rate, semi-eviscerated rate, eviscerated rate. Similarly, regarding meat quality, RIHR reduced cooking loss rates while elevating pH24h, a*, b*, and springiness. These findings demonstrate RIHR's potential to enhance both meat quantity and quality, although statistical significance was not achieved in the current result. In addition, we also measured serum biochemical and immunoglobulin levels, and the results showed that serum TP, ALB, A/G, and GLU concentrations were significantly up-regulated, concurrently with reduced LDL-C. Regarding immune function, serum IgA concentrations were significantly elevated across all RIHR supplementation groups in broilers. The observed alterations in these serum parameters suggest that the effects of RIHR may be achieved through modulation of these sensitive indicators, which are functionally linked to protein and lipid metabolism pathways.
Chinese herbal medicine exerts beneficial effects on the host by modulating gut microbiota composition. This study further investigated the impact of RIHR on intestinal microbiota in broilers. The results demonstrated that RIHR modulated the composition of gut microbiota throughout the intestinal tract of broilers. However, the dominant phyla in the foregut (Firmicutes and Proteobacteria) and hindgut (Firmicutes and Bacteroidota) remained unaltered. Consequently, both digestive/absorptive capacities and fermentation-dependent immune functions were preserved in broiler intestines. It is noteworthy that the decreased Firmicutes-to-Bacteroidetes ratio in the hindgut of broilers from the RIHR-treated group primarily suggests that the intestinal degradation of complex dietary fiber and plant polysaccharides is enhanced, enhancing the capacity to produce short-chain fatty acids[22]. Genus-level analysis revealed that the foregut of the RIHR-H group exhibited significantly increased abundances of Lachnospiraceae and Alistipes. Furthermore, LEfSe analysis revealed significant enrichment of c_Coriobacteriia and f_UCG-010 in this group. Concomitantly, the hindgut demonstrated elevated abundances of Alistipes and Bacteroides, with multiple taxa within phylum Bacteroidota showing significant enrichment in LEfSe analysis. Previous research has established that Lachnospiraceae represent significant producers of SCFAs, particularly acetate and butyrate, which may modulate ileal lipid metabolism through GPR41/43 receptor signaling pathways[23]. Concurrently, Bacteroides demonstrate high propionate and secondary bile acid production, both serving as endogenous agonists of the PPAR signaling. Similarly, Alistipes contributes to propionate biosynthesis[24]. Therefore, enrichment of these taxa suggests a potential increase in microbial metabolites capable of interacting with host lipid metabolic pathways[25]. According to published KEGG functional annotations and previous reports, members of the genera Alistipes and Bacteroides harbor genes involved in fatty acid metabolism, bile acid transformation, and PPAR signaling-related pathways, suggesting their potential roles in modulating host lipid metabolism[26,27]. In addition, these genera have been reported to participate in intestinal immune regulation, including IgA-associated mucosal responses and NOD-like receptor signaling, indicating a possible contribution to immune homeostasis[26]. Correlation analyses further substantiated significant associations between intestinal microbiota and serum lipid metabolites. The ileum constitutes the principal site for bile acid recirculation and active lipid transport in broilers. Consequently, lipid-metabolism-associated pathways in the ileum exhibit heightened susceptibility to modulation by gut microorganisms and their metabolic derivatives[28]. However, SCFAs and bile acid concentrations in intestinal contents were not directly quantified in the present study. Therefore, the causal relationship between microbiota-derived metabolites and PPAR pathway activation remains to be further verified. Future studies incorporating targeted SCFA and bile acid measurements will be required to establish a direct mechanistic connection.
Among the three supplementation levels, the RIHR-H group showed the most pronounced modulation of serum biochemical indices, immunoglobulin levels, and gut microbiota composition. Although growth performance parameters were not statistically significant, a dose-dependent trend was noted across RIHR-L, RIHR-M, and RIHR-H groups. Based on these trends and microbiota analyses, the RIHR-H group was chosen for subsequent transcriptomic and metabolomic profiling to explore potential molecular mechanisms, while acknowledging that further validation is required to confirm causality. Based on the phenotypic parameters and gut microbiota analysis, we conducted transcriptomic profiling of the ileum in broilers, which revealed significant alterations in gene expression relative to the control group, with 673 significantly upregulated and 691 downregulated DEGs identified. These DEGs demonstrated enrichment in GO terms, including transmembrane receptor protein tyrosine kinase activity, long-chain fatty acid metabolic process, and high-density lipoprotein particle binding, indicating enhanced microbiota metabolite-host receptor interactions and intensified lipid catabolic processes[29,30]. KEGG pathway analysis suggested enrichment in the PPAR signaling pathway and other lipid-metabolism-related pathways. Metabolites within these pathways may modulate PPAR signaling through substrate activation or ligand provision[31]. Moreover, we quantified the expression of key genes involved in the PPAR signaling pathway in the ileum. The five genes selected for qPCR validation (FABP1, ACC, FAS, PPARα, and SCD1) represent key regulatory nodes of intestinal lipid absorption, fatty acid synthesis, and PPARα-mediated transcriptional control, and exhibited strong differential expression in transcriptomic analysis. Nevertheless, we acknowledge that validation of a broader panel of lipid metabolism-related DEGs would further strengthen the evidence for PPAR pathway enrichment. It should be noted that for serum biochemical, immunoglobulin, intestinal microbiota, transcriptomic, qPCR, and muscle metabolomic analyses, only four chickens per group were sampled (n = 4). This relatively small sample size may limit the statistical power to detect subtle or moderate effects. Consequently, some observed trends, such as changes in growth performance, slaughter performance, meat quality, and microbiota composition, did not reach statistical significance despite showing consistent directional effects. Due to limited remaining biological samples, additional qPCR validation could not be performed in the present study. Future studies will include expanded gene validation to further confirm the transcriptomic findings. The results of the exhibition showed that broilers in the RIHR-H group exhibited upregulated PPARα expression, a significant downregulation of FABP1, and downregulation trends in the mRNA levels of lipogenic genes ACC, FAS, and SCD1. Activation of PPARα has been demonstrated to upregulate the expression of ApoA-I and ApoA-II, while enhancing ABCA1-mediated cholesterol efflux, thereby promoting HDL-C biogenesis and reverse cholesterol transport. Previous studies have provided a reference framework for this mechanism, which is typically associated with elevated serum HDL-C levels in clinical settings[32]. Significant downregulation of FABP1 in the ileum implies a reduction in the intracellular transport capacity of enterocytes for fatty acids and cholesterol. This attenuation consequently leads to diminished enterocytic lipid efflux and cholesterol absorption, ultimately altering the peripheral lipoprotein profile by reducing the secretion of VLDL and LDL-C[33]. As a negative acute-phase protein, serum albumin levels increase in response to reduced inflammation or improved nutritional status. This increase may result from the activation of the PPARα signaling pathway[34]. Although extracted, RIHR retains multiple bioactive components. Our previous study confirmed that dietary supplementation with RIHR enhances serum IgA levels in laying hens[10]. Furthermore, increased abundance of SCFA-producing bacteria has been shown to promote IgA class switching in B cells[35]. Collectively, these findings suggest that RIHR may ameliorate lipid metabolism and enhance mucosal immune function in broilers by modulating the abundance of intestinal SCFA-producing bacteria and regulating lipid metabolic pathways in the ileum. To investigate if RIHR's influence on lipid metabolism extends to muscle tissue, we performed lipid metabolomic analysis of broiler breast muscle. The results showed that RIHR significantly altered lipid metabolism in breast muscle, as evidenced by the upregulation of 46 DMs and the downregulation of 14 DMs in the RIHR-H group, accompanied by distinct changes in metabolite clustering. Furthermore, RIHR feeding markedly increased the levels of several unsaturated fatty acid metabolites in breast muscle, which are of potential nutritional and health significance and considered beneficial to consumers. The literature suggests that plant-derived compounds may influence poultry lipid metabolism by enhancing antioxidant capacity and modulating gene expression[32], providing a conceptual basis to explore potential effects of RIHR on muscle lipid composition. We therefore speculate that multiple bioactive components in RIHR may influence breast muscle lipid metabolism through similar mechanisms. In support of this, an integrated metabolomic and transcriptomic study on chicken meat quality and flavor revealed that muscle differences between chicken breeds were significantly enriched in glycerophospholipid metabolism and α-linolenic acid metabolism; notably, these two pathways were also significantly enriched in the present study[36]. Since such pathways are commonly associated with phospholipid composition of cell membranes, energy metabolism, and the generation of flavor precursors, our findings collectively indicate that RIHR may indirectly affect meat quality traits by modulating muscle lipid composition.
-
The present study indicates that dietary supplementation of RIHR may improve growth trends in broilers, modulate mucosal immune function, and influence lipid metabolism. Specifically, RIHR supplementation was associated with changes in gut microbiota composition, including increased abundance of c_Coriobacteriia and f_UCG-010 in the foregut and altered Firmicutes-to-Bacteroidetes ratio in the hindgut, which could enhance intestinal fiber degradation and SCFA production. Although transcriptomic and metabolomic analyses suggest potential activation of the PPAR signaling pathway in the ileum and altered lipid metabolic activity, these findings require further experimental validation before mechanistic conclusions can be confirmed. In addition, changes in breast muscle lipid composition may contribute to meat quality traits, though the direct mechanisms remain to be elucidated.
-
All procedures were reviewed and preapproved by the Animal Ethics Committee of Hunan Agricultural University (Approval No. 2023159) (January 1, 2023). The study adhered to the ethical guidelines of 'Replacement, Reduction, and Refinement' to minimize animal use and distress. Comprehensive information on animal housing, husbandry, and analgesia is included in this article to ensure that any potential discomfort to the animals during the experimental procedures is kept to a minimum.
-
The authors confirm contribution to the paper as follows: conceptualization: Huang P; methodology: Xiao Y, Xiao H, Zeng C, Ouyang W, Wang W, Huang P; software: Xiao Y, Xiao H, Zeng C; validation: Xiao Y, Xiao H, Zeng C, Ouyang W; formal analysis: Ouyang W; in-vestigation: Xiao Y, Xiao H, Zeng C, Ouyang W; resources: Xiao Y, Xiao H, Zeng C, Ouyang W, Wang W; data curation: Xiao Y, Xiao H, Zeng C, Ouyang W; writing − original draft preparation: Xiao Y, Xiao H, Zeng C; writing − review and editing: Huang P, Song M, Javed S; visualization: Xiao Y, Xiao H, Zeng C, Ouyang W; supervision: Huang P, Song M, Javed S; project administration, funding acquisition: Huang P. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets were deposited in a repository and are available at https://dataview.ncbi.nlm.nih.gov/object/PRJNA1101730 Information can be made available from the authors upon request.
-
This research was supported by the National Key R&D Program of China (Grant No. 2023YFD1301200). The authors thank all the participants and researchers who contributed and collected data.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/animadv-0026-0003.
-
# Authors contributed equally: Yu Xiao, Haibo Xiao, Cheng Zeng
- Supplementary Table S1 The determination results of amino acids in RIHR.
- Supplementary Table S2 Compound names and average content in RIHR (μg/g).
- Supplementary Table S3 The analysis results of the conventional nutritional components of RIHR.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xiao Y, Xiao H, Zeng C, Ouyang W, Wang W, et al. 2026. Radix isatidis herbal residue promotes growth performance, mucosal immunity, and muscle lipid metabolism in broilers via the gut microbiota and ileal PPAR signaling pathway. Animal Advances 3: e016 doi: 10.48130/animadv-0026-0003 

Radix isatidis herbal residue promotes growth performance, mucosal immunity, and muscle lipid metabolism in broilers via the gut microbiota and ileal PPAR signaling pathway
- Received: 13 December 2025
- Revised: 24 January 2026
- Accepted: 10 February 2026
- Published online: 28 May 2026
Abstract: This study examined the effects of dietary Radix isatidis Herbal Residue (RIHR) on growth performance, mucosal immunity, breast muscle lipid metabolism, and its underlying mechanisms in white-feathered broilers. A total of 320 one-d-old broilers were randomly assigned to four groups by random allocation: a control group received the basal diet, whereas the remaining three groups were offered the same diet fortified with 5, 10, or 15 kg/t of RIHR for the entire 42-d-period. From the collected observations, RIHR supplementation tended to improve ADFI, as reflected by a numerical increase, without negatively affecting FCR. Moreover, the RIHR-H group exhibited significantly elevated serum IgA and HDL-C levels, along with a significant decrease in LDL-C. In the foregut, the abundance of Lachnospiraceae and Alistipes appeared at higher levels, accompanied by a marked rise in c_Coriobacteriia and f_UCG-010. Changes were also evident in the hindgut, where both Alistipes and Bacteroides showed greater representation, along with an overall increase in the Firmicutes-to-Bacteroidetes ratio. Ileal transcriptomic analysis revealed activation of the PPAR signaling pathway and improved lipid metabolism in the ileum. Notably, breast muscle lipid metabolomics indicated a significant increase in the content of multiple unsaturated fatty acids in the RIHR-H group. In conclusion, dietary RIHR promotes growth, enhances mucosal immune function, and modulates lipid metabolism in both the intestine and breast muscle by regulating the gut microbiota.