








-
Mango (Mangifera indica L.) is a major tropical fruit crop belonging to the family Anacardiaceae[1−3]. It ranks as the third most economically important tropical fruit globally, and China is currently the world’s second-largest producer[4,5]. Production is concentrated in southern China, particularly in Yunnan Province, one of the country’s primary mango-growing regions[4−6]. Commercial orchards rely on grafted systems using locally adapted rootstocks and elite scions, with Tainong No.1, Guifei, and Jinhuang among the most widely cultivated cultivars[4,5].
Optimal mango production is typically achieved 4–5 years after establishment and requires substantial investment in irrigation, fertilization, and orchard management[4,5,7]. However, climate change poses increasing risks to mango cultivation worldwide. Approximately 26% of currently suitable mango-growing areas are projected to experience negative impacts due to warming and altered climatic regimes[8−11]. Warming trends are particularly pronounced at lower elevations[12−14], threatening established orchards in valley and lowland systems.
Within the framework of circular agriculture systems, long-term sustainability depends on enhancing resource-use efficiency, minimizing external inputs, and increasing resilience to environmental variability. Instead of increasing fertilizer or irrigation to offset climatic stress, spatial adaptation strategies such as shifting cultivation along natural altitudinal gradients—offer a viable alternative. Relocation to higher altitudes has been proposed as a promising climate adaptation strategy for mango[8,10,13,14]. However, implementation requires robust field-based evidence of physiological responses across elevation gradients[8,15].
Mango has been reported to grow across altitudes ranging from 450 to 1,200 m above sea level in tropical regions[16−18]. Yet, growth limitations and increased seedling mortality have been observed above 1,000 m due to low-temperature stress[8,11,19]. Most previous studies are observational or conducted under controlled or homogeneous orchard conditions[9,10], limiting their applicability to heterogeneous production landscapes.
Honghe County in Yunnan Province provides a natural experimental gradient, where orchards are distributed across varying altitudes and exposed to distinct microclimatic conditions[4−6,18]. Climate projections indicate continued temperature increases in this region, with warming patterns closely tied to seasonal dynamics[12,20,21]. Elevated temperatures may reduce seedling survival, prolong vegetative phases, delay flowering, and ultimately compromise productivity[9,11,22]. These effects are expected to intensify during the hot-dry season[9−11,19,23,24].
Tracking early growth patterns across different altitudes and seasons is key to creating sustainable orchards. This approach aligns with circular agriculture by optimizing where we plant, making nutrient use more efficient, and lowering the demand for external inputs.
-
The study was conducted in Honghe County, Yunnan Province, China (Fig. 1a, b), from August 2018 to September 2019. Seasons were classified based on the regional climatic regime as cold–dry (November–February), hot–dry (March–May), and wet–warm (June–October). Experimental mango plantations were established at two elevations representing contrasting climatic conditions: 450 (23°25'34" N, 102°15'50" E), and 1,050 m (23°24'30" N, 102°14'47" E) above sea level (a.s.l.) (Fig. 1c).
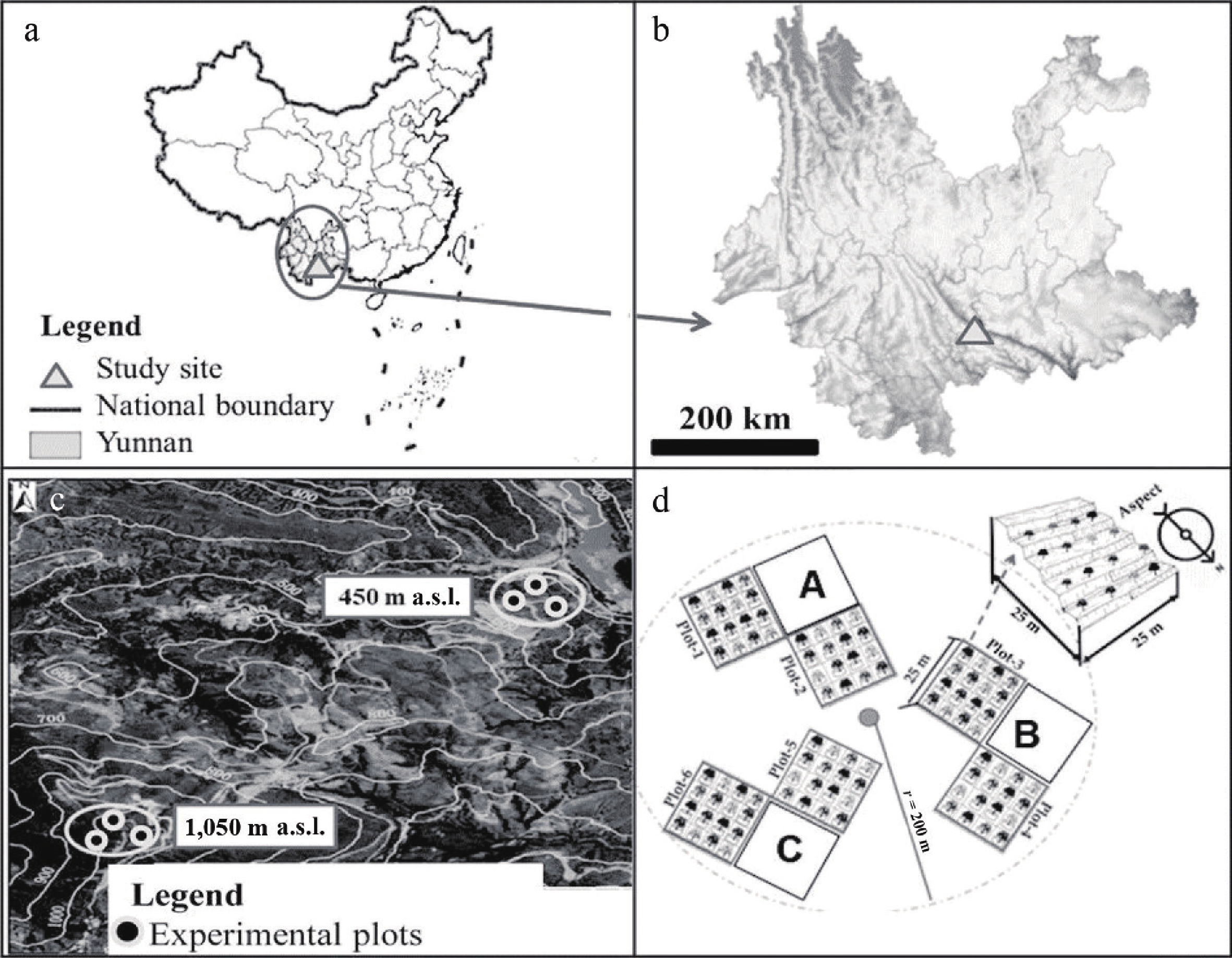
Figure 1.
(a) Experimental plots located at altitude 450 and 1,050 m a.s.l. (a), (b) Map of the study location at Honghe County, Yunnan Province, China (c) Experimental plots at 450 or 1,050 m a.s.l. (d) Illustration of experimental plots layout located within a radius of (ca. 100 m), and facing north. (GIS clip from global map).
At each altitude, six north-facing, 25 m × 25 m plots were established (designated as Plots A–C; Fig. 1d). Local climate data were retrieved from automated weather stations (HOBO-Onset U30, Onset Computer, Bourne, Massachusetts, USA) installed approximately 100 m from the study plots at both elevations (Fig. 2a–c). To assess the initial soil chemical composition, 60 composite samples were collected—30 per altitude and five per plot. These were randomly sampled from a depth of 0–10 cm using a 38 mm diameter auger. The samples were then analyzed via conventional chemical methods, with the findings summarized in Table 1.
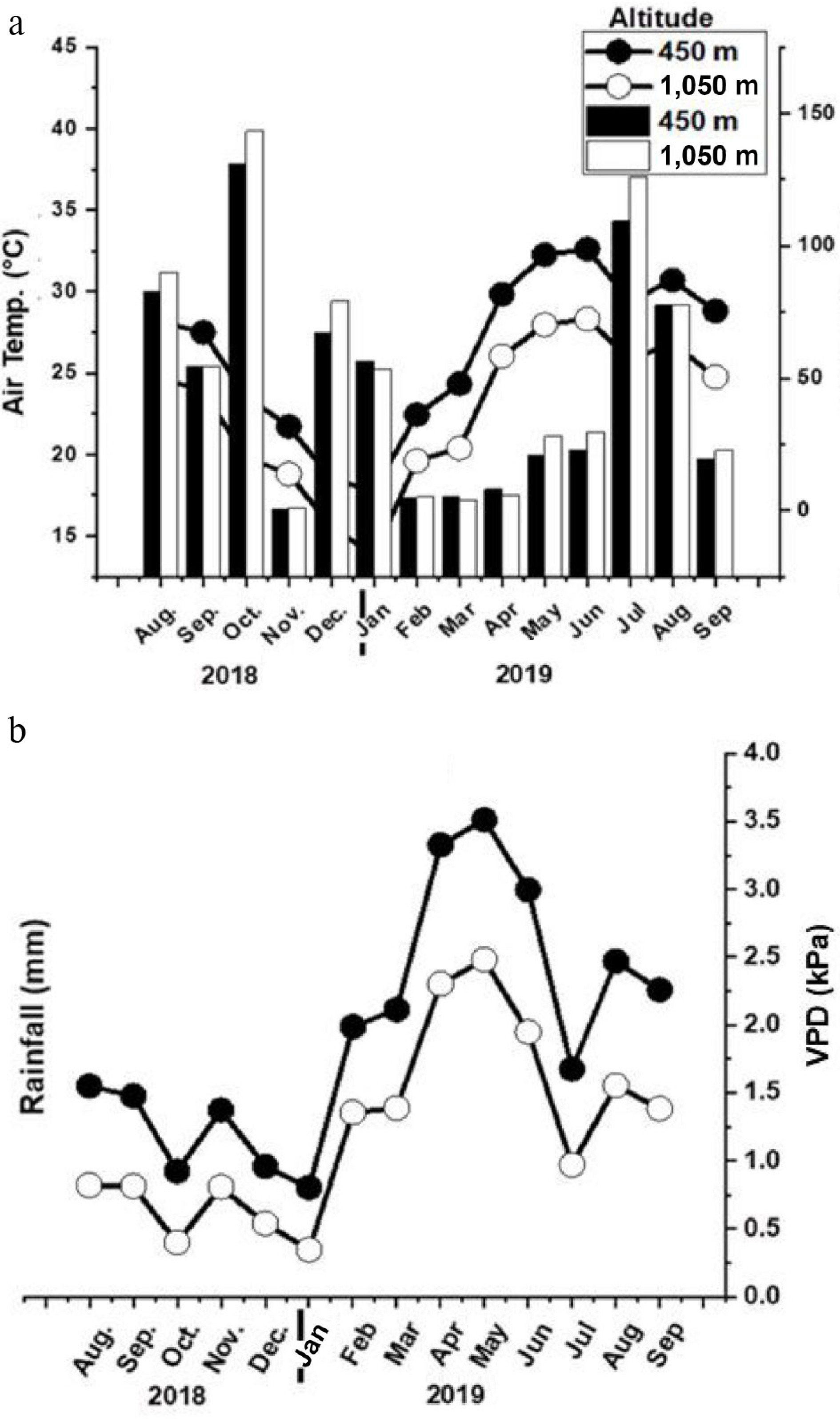
Figure 2.
(a) Monthly mean air temperature (°C), and rainfall (mm). (b) Vapor pressure deficit (VPD kPa) of the study site at 450 and 1,050 m a.s.l.
Table 1. Soil chemical composition of the study plots located at 450 and 1,050 m altitudes before any treatment is applied. Different letters indicate the difference between means is statistically significant, whereas similar letters indicate the absence of a statistically significant difference between means, p ≤ 0.05.
Composition Unit Altitude (m) 450 1,050 pH − 6.72 ± 0.05a 6.67 ± 0.07a Soil organic carbon g/kg 8.89 ± 0.8a 7.61 ± 0.4a Total nitrogen g/kg 1.21 ± 0.7a 1.016 ± 0.03a Total phosphorous g/kg 0.5 ± 0.01a 0.54 ± 0.02a Available phosphorous mg/kg 9.49 ± 0.34a 8.93 ± 0.68a Total potassium g/kg 26.12 ± 0.7a 27.3 ± 1.0a Available potassium g/kg 0.224 ± 0.02a 0.22 ± 0.01a Two-year-old grafted mango tree seedlings were purchased from a local nursery. The graft union comprised four scions (a detached shoot grafted to rootstock) cultivars, namely 'Tainong No. 1', 'Guifei', 'Jingdong', and 'Jinhuang', grafted to the local rootstock/trunk (the portion of the stem to which a scion is grafted) cultivar 'Daheman'. Initial heights and diameters of the scions and the trunk were equivalent, and they were planted randomly at a similar depth, with planting density of 5 m × 3 m, at 450 and 1,050 m a.s.l. During seedling transplantation, 2 kg of compost was mixed into the soil. Next, 0.15 kg of urea per tree was applied in April, and 0.8 kg of compound fertilizer N, P, K (ratio = 16:16:16) per tree was applied in June and August, 2019. Initial soil chemical properties did not differ significantly between altitudes (Table 1), and mango trees at both sites were managed under identical fertilization and irrigation regimes following local commercial practices. However, soil nutrients were not monitored continuously during the experimental period.
Growth measurements were obtained from 96 trees at each altitude, comprising 24 trees per cultivar per site. Replication was therefore based on multiple trees per cultivar at each altitude. Plant height, measured from ground level to the tip of the apical shoot, and trunk height, measured from ground level to the graft union (representing rootstock height), were determined using a measuring tape. Trunk (rootstock) and scion diameters were measured at 3 cm above ground level and 3 cm above the graft union, respectively, using a digital caliper. Height and diameter increments were recorded at 30-d intervals and subsequently used for statistical analyses.
Leaf samples were collected from 72 trees at each altitude, comprising 18 trees per cultivar per site. From each sampled tree, three fully expanded mature leaves were collected from a mid-canopy position to ensure uniform sampling. Leaf area was measured using a leaf area meter (Li-Cor 3001A, USA).
To minimize potential residual nursery effects on nutrient status, leaf sampling was conducted one year after field establishment (August 2019). To ensure a representative nutrient profile, three fully expanded leaves were collected from the mid-canopy of each mango tree. To minimize intra-tree variability, these leaves were pooled into a single composite sample per tree prior to processing. Samples were oven-dried at 70 °C for 48 h to determine dry weight, then ground using a mortar and pestle and passed through a No. 100 mesh sieve to obtain a homogeneous powder for nutrient analysis.
Leaf nutrient concentrations were determined using mid-infrared diffuse reflectance spectroscopy (MIR), with an ALPHA II FT-IR Base Spectrometer (Bruker, Germany) based on a calibrated prediction model. To develop the spectral calibration, a representative subset (n = 48) was randomly selected for primary chemical analysis. Carbon (C) and nitrogen (N) concentrations were determined via dry combustion using an elemental analyzer (vario MICRO Cube, Elementar, Germany) on 10 mg subsamples. Phosphorus (P), potassium (K), magnesium (Mg), and calcium (Ca) were quantified using atomic absorption spectrometry (PinAAcle 900T, PerkinElmer, USA). These primary measurements served as the reference data to calibrate and validate the Mid-Infrared (MIR) spectral prediction models; performance statistics for these models are summarized in Table 2. For the broader MIR analysis, 100 mg of finely ground leaf tissue per sample was scanned. The resulting spectral data were exported and processed in R to predict nutrient concentrations across the remaining sample set.
Table 2. Calibration and validation statistics for the mid-infrared (MIR) spectroscopy models of mango leaf nutrients (n = 48).
Nutrient Unit R2cal R2Val RMSE RPD Carbon (C) g/kg 0.88 0.84 20.53 2.4 Nitrogen (N) g/kg 0.91 0.87 2.96 2.8 Potassium (K) g/kg 0.89 0.85 1.15 2.5 Calcium (Ca) g/kg 0.92 0.88 0.55 2.9 Phosphorus (P) g/kg 0.85 0.81 0.27 2.1 Magnesium (Mg) g/kg 0.86 0.82 0.06 2.2 Individual trees were considered the experimental units for statistical analyses, while plots served as spatial blocks within each altitude to maintain uniform site conditions. Accordingly, growth and nutrient measurements were analyzed at the tree level. The experiment followed a completely randomized design to evaluate the effects of altitude on vegetative growth and leaf nutrient composition of four mango cultivars. To compare vegetative growth and nutrient concentrations between the two altitudes (450 and 1,050 m), independent-sample t-tests were performed for each cultivar. Statistical significance was determined at (α = 0.05). Differences were considered significant at (p ≤ 0.05), and highly significant at (p < 0.01). Analyses were conducted using Statistix (Version 8.1), and results are presented as means ± standard error (± SE). Multivariate regression analysis was performed in R (Version 4.2.0) to quantify the influence of climatic variables on growth. The relative contribution of each factor was determined using standardized regression coefficients (β), normalized to 100% to represent the proportional influence of temperature, vapor pressure deficit (VPD), and rainfall. Graphical representations were generated using OriginPro 2021 (Version 9.65).
-
Altitudinal relocation significantly influenced vegetative growth dynamics of mango cultivars, demonstrating strong climatic regulation of biomass accumulation and structural development.
Across cultivars, mean annual growth in plant and scion heights increment was significantly greater at 450 m compared to 1,050 m (p < 0.01; Fig. 3a, b), whereas trunk height increment was significantly lower at 450 m (Fig. 3c). This contrasting response suggests a shift in vertical growth allocation along the altitudinal gradient, with lower elevation promoting rapid shoot elongation and higher elevation relatively favoring trunk extension under cooler conditions.
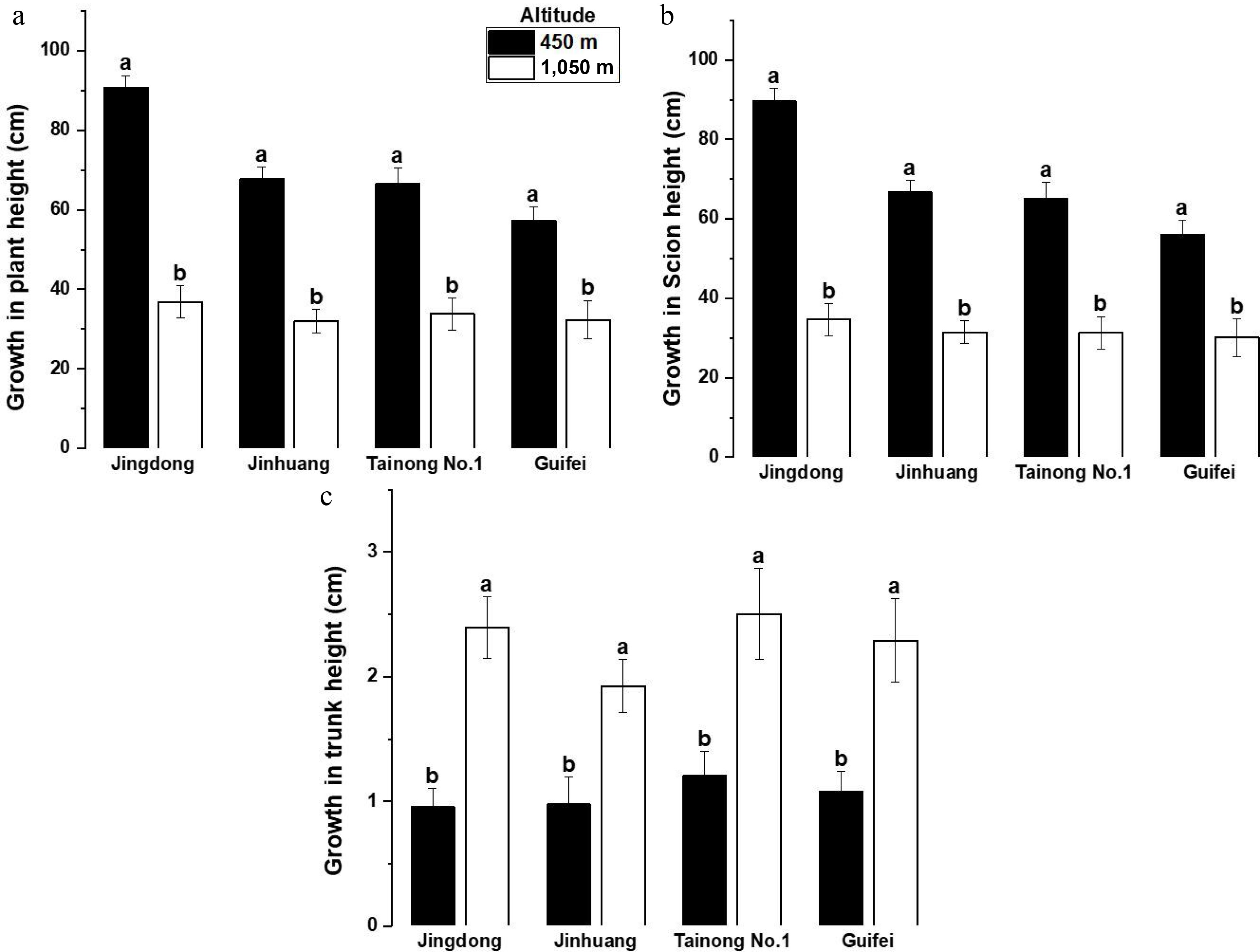
Figure 3.
Mean ± SE annual growth in (cm). (a) Plant height, (b) scion heights, (c) trunk height of four mango cultivars grown at altitudes of 450 and 1,050 m a.s.l. Different letters indicate differences between means are statistically significant p < 0.01.
Mean annual scion and trunk diameter increments also differed significantly between altitudes (p < 0.01), with consistently greater radial growth observed at 450 m (Fig. 4a, b). The combined increase in height and diameter growth at lower altitude indicates enhanced early biomass accumulation and underscores altitude as a critical environmental determinant of vegetative performance.
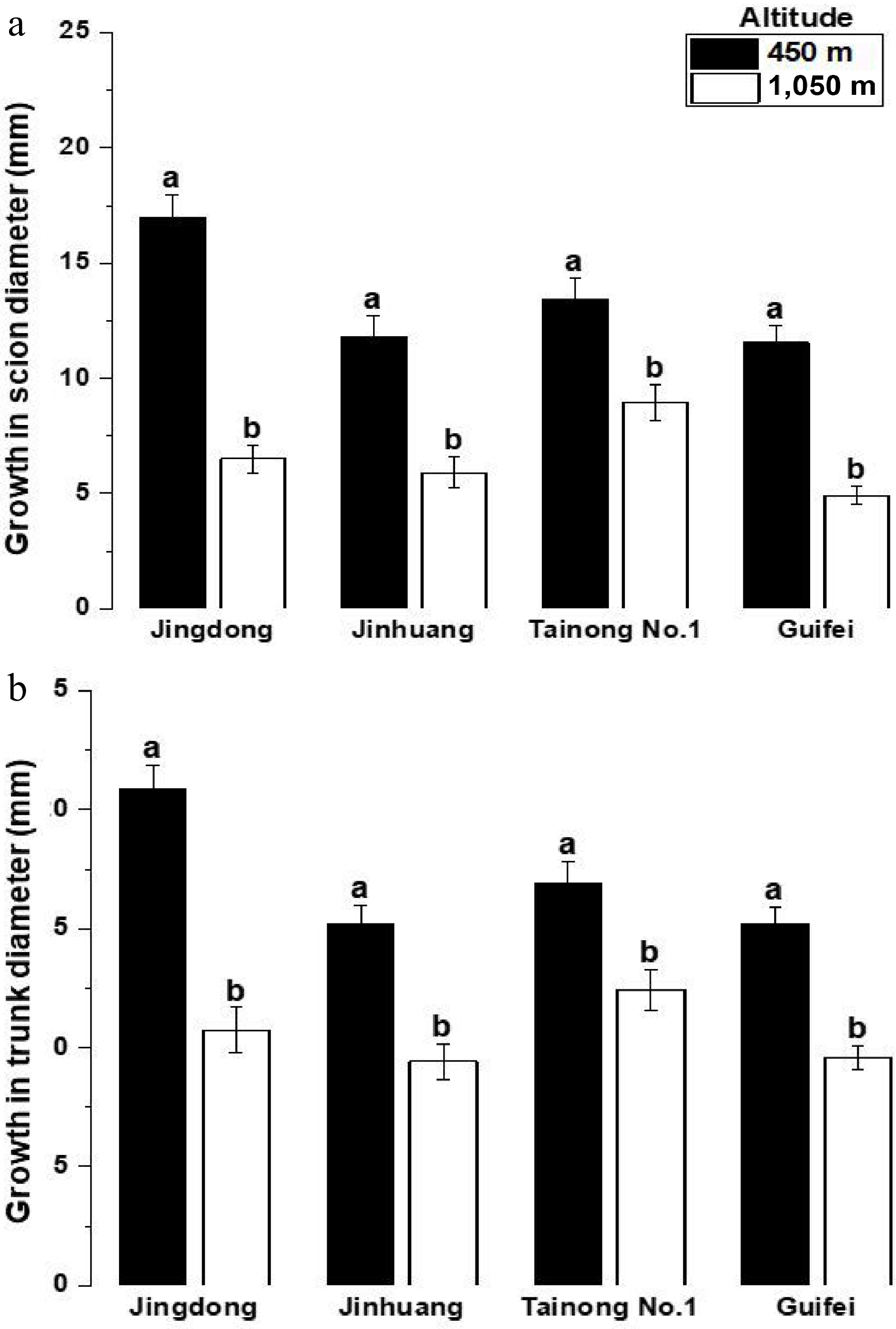
Figure 4.
Mean ± SE annual growth in (mm). (a) Scion, (b) trunk diameters of mango cultivars grown at altitudes of 450 and 1,050 m a.s.l. Different letters indicate the difference between means is statistically significant, p < 0.01.
Vegetative growth dynamics were strongly governed by environmental variables (R2 = 0.87; Fig. 5), with temperature and vapor pressure deficit (VPD) emerging as the predominant drivers of mango canopy development.
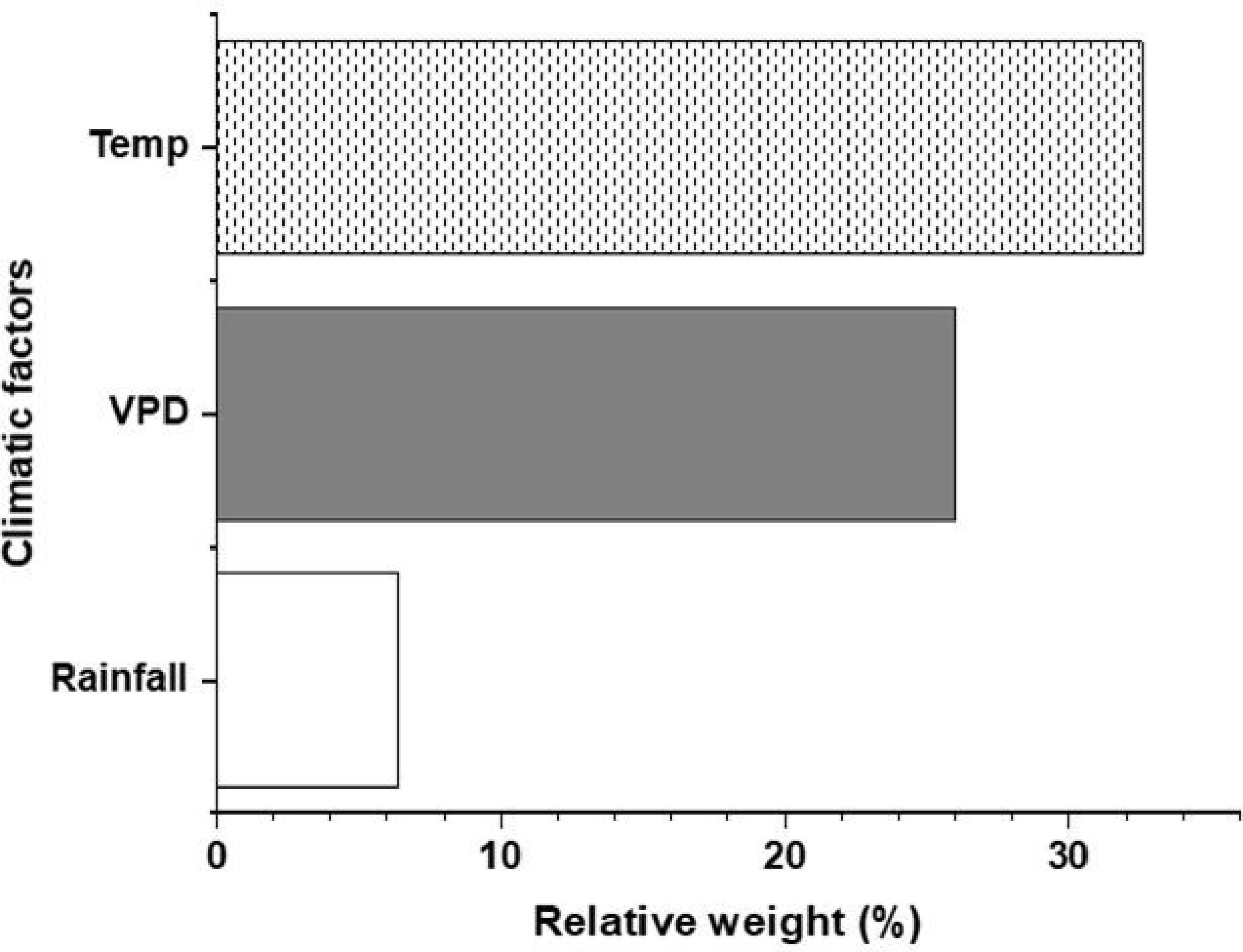
Figure 5.
Relative weight (%) of climatic factors to vegetative growth variation (R2 = 0.87).
These growth responses closely mirrored the climatic differences observed between sites. Temperature and vapor pressure deficit (VPD) were consistently higher at 450 m (Fig. 2a, b), and multivariate analysis confirmed that temperature and VPD were the dominant climatic drivers of vegetative growth (Fig. 5). Such conditions are widely recognized to stimulate mango vegetative development[8,10,11]. Optimal vegetative performance in mango has been reported under day/night temperature regimes between 20/25 °C and 30/25 °C[15], which were more closely approximated at the lower elevation site. Elevated VPD at 450 m may have further enhanced transpiration-driven nutrient transport and metabolic activity[8,9,11,25], supporting increased shoot elongation and radial expansion. Comparable responses have been documented in silver birch, where VPD altered foliar metabolites and growth dynamics[26], reinforcing the role of atmospheric demand in regulating vegetative performance.
In contrast, vegetative growth was uniformly restricted at 1,050 m (Fig. 3a, b; Fig. 4a, b), most likely due to prolonged exposure to lower temperatures that constrained photosynthesis and cambial activity, as demonstrated in both field and controlled studies[10,15,27,28]. These findings collectively confirm that mango vegetative growth is strongly temperature-limited. Nevertheless, cooler conditions have also been associated with enhanced floral induction[9,11,20,24,28], suggesting a vegetative–reproductive trade-off along the altitudinal gradient. This trade-off implies that higher elevations may become increasingly favorable for sustaining reproductive stability under projected warming scenarios[13], even if vegetative growth remains comparatively moderate.
Seasonal variation further confirmed the dominant influence of temperature and atmospheric demand on growth dynamics. Height increments (Fig. 6a, b) and scion and trunk diameter increments (Fig. 7a, b, d) were significantly greater during the hot-dry and wet-warm seasons than during the cold-dry season at both altitudes, except for scion diameter during the hot-dry season at 1,050 m (Fig. 7c). This seasonal pattern confirms that mango vegetative flushing is closely synchronized with warmer periods[8,11]. Growth suppression during the cold-dry season highlights the sensitivity of vegetative development to cooler temperatures, particularly at higher elevations.
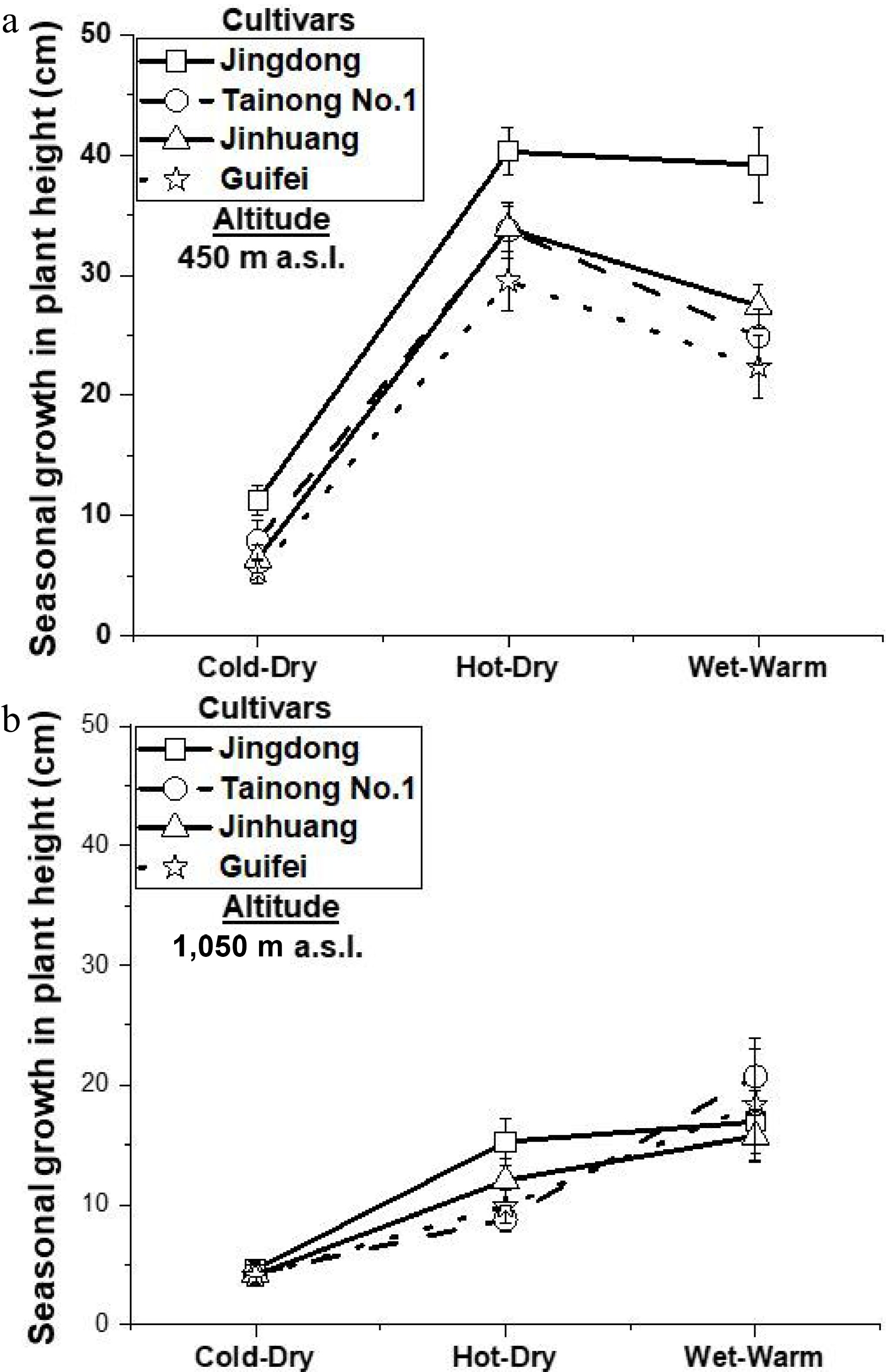
Figure 6.
Mean ± SE seasonal height growth (cm) during cold-dry, hot-dry, and wet-warm of four mango cultivars grown at 450 and 1,050 m, p < 0.01.
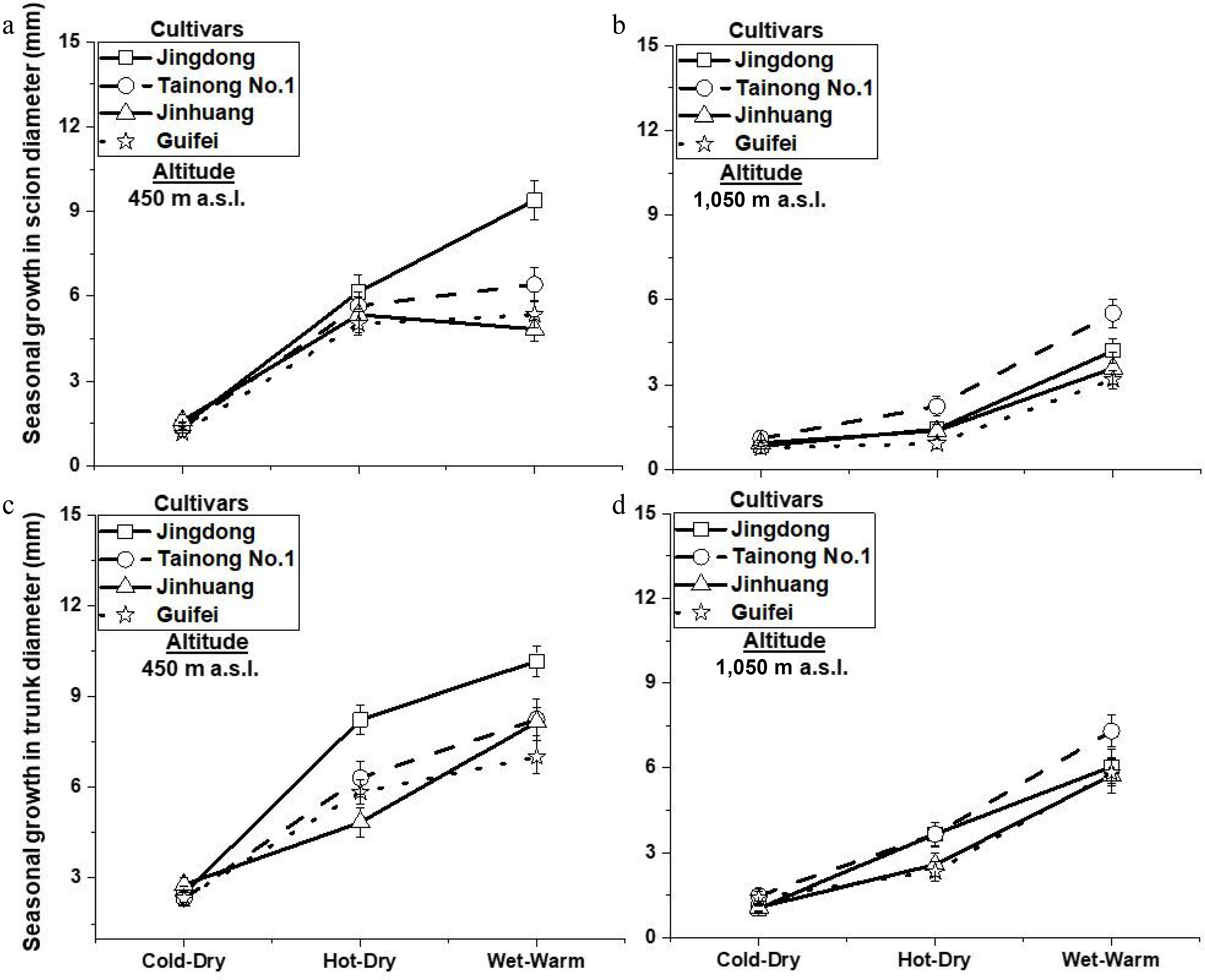
Figure 7.
Mean ± SE seasonal growth in scion and trunk diameter (mm) during cold-dry, hot-dry, and wet-warm of four mango cultivars grown at 450 and 1,050 m, p < 0.01.
Leaf morphological responses were cultivar-dependent. Leaf fresh weight, dry weight, and leaf area were significantly greater at 450 m for Jingdong and Tainong No. 1 (p < 0.05; Table 3), whereas Jinhuang and Guifei exhibited no significant altitudinal differences. In contrast, specific leaf area (SLA) remained stable across altitudes for all cultivars (Table 3). The conservation of SLA suggests that structural investment in leaf construction is largely genetically regulated or linked to cultivar origin, consistent with findings in mango and other woody species[25,29−31]. Thus, altitude primarily influenced total leaf biomass accumulation rather than intrinsic structural density or thickness.
Table 3. Mean ± SE of leaf fresh weight, dry weight, area, specific leaf area of mango cultivars at altitudes of 450 and 1,050 m a.s.l. Different letters indicate that the difference between means is statistically significant, whereas similar letters indicate the absence of statistically significant differences between means (p < 0.05).
Altitude (m) Cultivars Fresh weight (g) Dry weight (g) Leaf area (cm2) Specific leaf area (cm2 g−1) 450 Jingdong 1.72 ± 0.07a 0.75 ± 0.03a 59.8 ± 1.9a 86.52 ± 1.6a Jinhuang 2.30 ± 0.2a 0.97 ± 0.04a 67.2 ± 2.2a 72.28 ± 1.1a Tainong No.1 1.77 ± 0.1a 0.77 ± 0.03a 58.0 ± 1.8a 77.98 ± 1.5a Guifei 2.35 ± 0.01a 1.07 ± 0.04a 72.8 ± 2.4a 72.43 ± 1.1a 1,050 Jingdong 1.35 ± 0.06b 0.60 ± 0.02b 48.3 ± 1.5b 87.22 ± 1.6a Jinhuang 2.00 ± 0.06a 0.85 ± 0.03a 59.8 ± 1.7a 75.10 ± 3.6a Tainong No.1 1.52 ± 0.05b 0.68 ± 0.02b 49.3 ± 1.4b 74.77 ± 1.2a Guifei 2.03 ± 0.08a 0.87 ± 0.04a 64.3 ± 2.2a 77.33 ± 1.6a Leaf nutrient composition exhibited a distinct altitudinal pattern (Table 4). N and K concentrations were significantly higher at 1,050 m across cultivars, whereas calcium (Ca) was generally higher at 450 m except in Tainong No. 1. The C:N ratio was significantly higher at 450 m, while C:P remained largely stable except for Tainong No.1. The elevated N and K concentrations at higher altitude likely reflect reduced biomass dilution under slower vegetative growth rather than enhanced nutrient uptake by itself. Conversely, higher Ca concentrations at 450 m are consistent with greater transpiration flux under elevated VPD, facilitating Ca transport through the xylem[25]. These nutrient shifts illustrate how climatic gradients influence internal nutrient allocation and stoichiometric balance.
Table 4. Mean ± SE of leaf nutrient content of four mango cultivars of mango cultivars at altitudes of 450 and 1,050 m a.s.l. Different letters indicates the difference between means is statistically significant.
Altitude (m) Cultivar N (%) p < 0.01 K (mg g−1) p < 0.05 Ca (mg g−1) p < 0.05 C : P ratio p < 0.05 C : N ratio p < 0.05 450 Jingdong 2.2 ± 0.029b 6.4 ± 0.12b 25.08 ± 0.7a 312.2 ± 5.3a 18.7 ± 0.17a Jinhuang 2.2 ± 0.03b 6.3 ± 0.1b 25.7 ± 0.7a 321.8 ± 5.1a 19.135 ± 0.28a Tainong No.1 2.2 ± 0.028b 6.7 ± 0.12b 24.5 ± 0.8a 305.4 ± 7.8b 18.6 ± 0.13a Guifei 2.2 ± 0.037b 6.1 ± 0.21b 26.7 ± 0.9a 305.4 ± 7.8a 18.7 ± 0.2a 1,050 Jingdong 2.4 ± 0.02a 7.01 ± 0.14a 22.7 ± 0.6b 324 ± 5.5a 17.6 ± 0.17b Jinhuang 2.37 ± 0.027a 7.02 ± 0.12a 23.2 ± 0.4a 322.6 ± 8.1a 17.8 ± 0.18b Tainong No.1 2.4 ± 0.02a 7.01 ± 0.12a 22.8 ± 0.6b 326.45 ± 5a 17.8 ± 0.12b Guifei 2.39 ± 0.03a 7.02 ± 0.1a 23.4 ± 0.5b 318.7 ± 6.4a 17.7 ± 0.26b Scion–rootstock interactions may also have contributed to the observed growth differences. Enhanced scion vigor can stimulate rootstock development, improving hydraulic conductivity, nutrient uptake efficiency, and mechanical stability, as reported in apple and grape systems[32−34]. Such interactions become increasingly important under climatic gradients where efficient resource allocation determines overall system performance.
Honghe County remains one of the largest mango-producing regions in mainland China[4,5], yet projected increases in temperature and VPD across Yunnan Province[22,35,36] may intensify climatic stress, particularly at lower elevations. Temperature increases of up to 2.7°C under RCP8.5 scenarios[22,36] could prolong vegetative phases and disrupt flowering synchrony[24,27,28], potentially reducing yield stability. The 2019 growing season, during which this experiment was conducted, was warmer and drier than previous years, reflecting emerging climate trends. Although warmer conditions enhanced vegetative growth in the present study, consistent with previous findings[9,27,28], continued warming may negatively affect reproductive development[8,10,14,37,38].
Within the framework of circular agriculture systems, these findings have important implications for sustainable mango production under climate variability. Spatial redistribution of orchards along altitudinal gradients offers a strategy to align crop physiological requirements with prevailing climatic conditions rather than relying solely on compensatory external inputs[4,5,22,35,36]. In the present study, higher leaf N and K concentrations observed at 1,050 m under identical fertilization regimes suggest that trees at higher elevations may utilize nutrients more efficiently, offering potential to optimize fertilizer application while maintaining adequate plant nutritional status[9,27,28]. Such altitude-mediated nutrient patterns can enhance internal nutrient cycling within orchard systems, as leaf litter, pruned material, and other organic residues decompose and return essential nutrients to the soil[39]. Stable specific leaf area (SLA) across altitudes further indicates that biomass quality and litter decomposition dynamics are relatively predictable, supporting efficient nutrient retention and turnover within the system[39,40].
By aligning orchard placement with specific climatic and physiological requirements, production systems can become more self-sustaining. Combining spatial optimization with residue recycling and integrated nutrient management allows mango production to reduce dependence on synthetic inputs, maintain soil fertility, and improve long-term sustainability[11,13].
Furthermore, altitudinal relocation buffers production against climatic stress by distributing temperature and vapor pressure deficits across landscapes with varying elevations, effectively diversifying risk and enhancing resilience[39,40]. The predictable nutrient dynamics and biomass quality observed at higher altitudes also provide opportunities to integrate agroecological practices such as cover cropping, mulching, and compost application, which further close nutrient loops and improve water and soil conservation[39].
Adopting these circular principles strengthens the adaptive capacity of orchards by improving resource-use efficiency, reducing the environmental footprint of fertilization, and maintaining productivity under projected warming scenarios[39]. Although this study focused on vegetative growth, these findings indicate that aligning orchard location with climatic requirements creates a more coordinated system where nutrient inputs, plant growth, and biomass turnover are tightly linked. However, future research evaluating flowering phenology, fruit set, yield stability, and long-term soil nutrient dynamics will be essential to fully validate the viability of altitudinal relocation as a climate-resilient, circular mango production strategy[8,10,14,38].
-
Our results indicate that altitudinal relocation significantly modifies vegetative architecture and nutrient partitioning in young mango trees. While low-altitude conditions favor biomass accumulation via shoot and radial elongation, higher elevations promote greater foliar N and K concentrations, suggesting enhanced nutrient retention over rapid growth. Because growth intensity is primarily governed by temperature and VPD, lowland orchards face the highest risk from climate warming. Consequently, moderate upslope expansion serves as a viable adaptive strategy, optimizing the trade-off between vegetative vigor and reproductive resilience while reducing the need for intensive external inputs.
-
The authors confirm their contributions to the paper as follows: conceptualization, methodology, investigation, data curation, and draft preparation: Wubshet TT; supervision, review and editing: Lu P; technical support: Xie X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated and analyzed during the current study are available from the corresponding author upon reasonable request.
-
This work was generously supported by the Ministry of Sciences and Technology of China (2017YFC0505100), Yunnan Provincial Science and Technology Department (202003AD150004) and Chinese Academy of Sciences, President’s International Fellowship Initiative (2026PVC0121). We would like to express our gratitude to ICRAF, Honghe County Kubo Agricultural Development Co. Ltd for providing plots of land for this experiment. We extend our gratitude to colleagues at Kunming Institute of Botany, Chinese Academy of Sciences, and the Centre for Mountain Futures, Honghe in facilitating the necessary logistics during field study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wubshet TT, Lu P, Xie X. 2026. Altitudinal relocation alters growth and nutrient allocation of young mango cultivars: implications for climate-resilient orchard systems. Circular Agricultural Systems 6: e012 doi: 10.48130/cas-0026-0014 

Altitudinal relocation alters growth and nutrient allocation of young mango cultivars: implications for climate-resilient orchard systems
- Received: 20 February 2026
- Revised: 21 March 2026
- Accepted: 23 March 2026
- Published online: 20 May 2026
An Author Correction to this article was published on 30 June 2026, http://doi.org/10.48130/cas-0026-0020
Abstract: Shifting fruit production to higher elevations is a key climate-adaptation strategy to mitigate the impacts of rising temperatures in tropical regions. However, empirical field evidence regarding early tree physiological responses to such shifts remains scarce. This study evaluated the vegetative growth dynamics and leaf nutrient allocation of four commercial mango (Mangifera indica L.) cultivars (Jingdong, Guifei, Jinhuang, and Tainong No. 1) grown at two contrasting altitudes (450 and 1,050 m a.s.l.) in Honghe County, China. Standardized fertilization and irrigation regimes were applied at both sites. Leaf nutrient concentrations were determined using Mid-Infrared (MIR) spectroscopy calibrated against primary chemical analysis (n = 48). At 450 m, scion height and stem diameter increments were significantly greater, driven by warmer temperatures and higher vapor pressure deficit (VPD) (Multiple Linear Regression, R2 = 0.87). Conversely, leaf nitrogen (N) and potassium (K) concentrations were significantly higher at 1,050 m across all cultivars. These findings suggest that higher elevations promote enhanced nutrient retention and utilization efficiency rather than rapid biomass dilution. Within a circular agricultural framework, moderate upslope relocation acts as a potent spatial adaptation strategy. By aligning crop physiological requirements with natural altitudinal gradients, producers can reduce the linear flow of synthetic inputs, optimize internal nutrient cycling, and develop more resource-efficient, climate-resilient orchard systems.
-
Key words:
- Altitude /
- Climate adaptation /
- Vegetative growth /
- Nutrient allocation /
- Mango