










-
Unlike most animals, vascular plants exhibit continuous postembryonic development, primarily driven by meristematic activity[1]. The shoot apical meristem produces aboveground organs, the root apical meristem generates root tissues, and both meristems contribute to the postembryonic development of vascular plants[2]. The shoot apical meristem, which gives rise to the aboveground parts, can be divided into three functional domains[3]. Located in the center and summit of the shoot meristem, the central zone (CZ) harbors a mass of pluripotent stem cells. During postembryonic development, shoot stem cells continuously divide to generate daughter cells and simultaneously maintain themselves. The daughter cells are pushed laterally into the peripheral zone (PZ) to generate lateral organs, or downwardly to form the stem. A group of specialized cells beneath the CZ comprises the organizing center (OC), which is required for maintaining the stem cells[4].
The elaborate activity of the shoot meristem is controlled by a number of regulatory genes, among which the WUSCHEL-CLAVATA (WUS-CLV) module plays a central role[5]. WUS is specifically expressed in the OC and encodes a homeodomain transcription factor[6]. After translation, WUS migrates upward into the CZ through the plasmodesmata and activates the transcription of CLV3, the stem cell marker gene[7,8]. CLV3 encodes a secreted peptide, which is perceived by multiple sets of receptor complexes[9]. CLV3 signaling restricts WUS expression in the OC through ligand-receptor interactions involving CLV1, CLV2, and CLV3 itself, leading to the inhibition of WUS transcription[10,11]. Defects in WUS result in stem cell consumption, while mutations of CLV3 lead to over-proliferation of stem cells and enlargement of the meristem[6,10]. Thus, WUS and CLV3 form a regulatory feedback loop that controls shoot meristem homeostasis, where WUS activates CLV3 expression and CLV3-signaling restricts WUS expression.
Recent studies have revealed regulatory roles of the L1-miR171-HAM signaling cascade in maintaining the WUS-CLV module. HAIRY MERISTEMs (HAMs) are GRAS family transcription factors that form heterodimers with WUS and repress its ability to activate CLV3 expression in the OC[12−14]. Meanwhile, ARABIDOPSIS THALIANA MERISTEM LAYER1 and PROTODERMAL FACTOR2 specifically activate the transcription of miR171 in the epidermal cell layer[15]. Mature miR171 diffuses downward and then targets and cleaves HAMs transcripts in the CZ, thereby allowing CLV3 expression therein[16]. WUS acts in parallel with the KNOX1 family homeodomain transcription factor SHOOTMERISTEMLESS (STM)[17]. STM is expressed throughout the shoot meristem except for incipient primordia. STM stimulates cytokinin biosynthesis, suppresses differentiation, and maintains the proliferation of meristematic cells[18,19]. A recent study showed that STM interacts with WUS and enhances the binding of WUS to the CLV3 promoter, demonstrating the coordination between STM and the WUS-CLV module[20].
The phytohormones cytokinin and auxin play important roles in regulating the homeostasis of the shoot meristem. The cytokinin biosynthetic gene LONELY GUY4 is expressed in the epidermis of the shoot meristem and probably establishes a cytokinin gradient that extends into the OC, where the expression of AHK4 (encoding the cytokinin receptor ARABIDOPSIS HISTIDINE KINASES 4) overlaps with that of WUS[21,22]. Type-B ARABIDOPSIS RESPONSE REGULATORs (ARRs), the key regulators of cytokinin response genes, directly activate WUS transcription and sustain its expression level via suppressing auxin biosynthesis in the OC[22−26]. Cytokinin signaling is required for stabilizing the WUS protein[27]. WUS in turn represses the expression of type-A ARRs, which encode cytokinin signaling repressors[28]. Therefore, cytokinin signaling and WUS expression form a positive feedback loop to confine the appropriate organization of the shoot meristem. Auxin signaling is functionally connected to stem cell behavior[29]. In the CZ, WUS rheostatically controls auxin signaling via regulating histone acetylation at target loci, including genes involved in the auxin pathway. This permits low levels of auxin signaling, which enables stem cells to be resistant to auxin-induced differentiation, and is required for their maintenance[30]. Another study has shown that AUXIN RESPONSE FACTOR 5 mediates auxin signaling in the CZ by directly repressing the transcription of DORNROSCHEN, which encodes a positive regulator of CLV3 expression[31]. The findings of those studies revealed the molecular mechanism of auxin-controlled stem cell homeostasis.
Under proper in vitro culture conditions, the shoot meristem can be regenerated from somatic tissues[32]. When detached explants are cultured on auxin-rich callus induction medium, they produce callus, a mass of proliferating cells with transcriptional similarities to lateral root primordia but lacking a fully defined meristem identity[33]. After transfer onto cytokinin-rich shoot induction medium, type-B ARRs initiate WUS expression in clusters of callus cells and subsequently reestablish shoot meristems[23]. This process enables regeneration of the entire plant body and provides the essential foundation for in vitro propagation, genetic transformation, and the generation of virus-free plants[34,35]. Several lines of evidence indicate that WUS is essential for the regeneration of the shoot meristem[23,25,36].
Despite their similar structural features, shoot meristems of annual and perennial species exhibit distinct activities[37]. Annual plants preserve shoot meristem activity throughout the growth season and complete their life cycle before winter. However, for perennial plants such as trees in temperate and boreal regions, the shoot meristem perceives photoperiod and/or temperature signals and ceases activity in autumn[38]. Subsequently, bud scales form to protect the meristem from freezing injury[39]. As winter progresses, shoot meristems are gradually released from dormancy, resume activity, and give rise to bud flush[40]. In poplar, the induction of dormancy is controlled by the integration of photoperiod perception and the FLOWERING LOCUS T (FT) module[41]. Light signals are detected by phytochrome receptors and circadian clock components[40]. The circadian components LATE ELONGATED HYPOCOTYL 1 (LHY1), LHY2, GIGANTEA, and CONSTANS regulate the expression of FT2 in response to day length[42−45]. Downregulation of FT2 induces dormancy, while its overexpression prevents growth cessation and bud set[41].
Perennial woody plants account for 42.7% of angiosperms and represent a large proportion of global biodiversity[46,47]. Seasonal dormancy and secondary growth are typical characteristics of woody species. Signal transduction that induces meristem cessation has been studied in detail[48]. However, there is no link between cessation signaling and shoot meristem activity. Whether the maintenance of the shoot meristem affects secondary growth remains unknown. In this study, we investigated the regulatory roles of the poplar genes PagWUS and PagCLV3, orthologs of Arabidopsis WUS and CLV3, respectively. The results reveal their conserved as well as distinct functions compared with those of their orthologs in herbaceous species, consistent with the developmental characteristics of perennial trees.
-
Poplar (Populus alba × Populus glandulosa var. glandulosa) clone 84 K was used as the wild-type background. The leaves of sterile saplings cultured on rooting medium for 4−6 weeks were used as explants for genetic transformation. Phenotypic and histological analyses were performed using sterile saplings cultured on rooting medium for 2 months. Saplings grown on rooting medium for 2 months and transferred to soil for 1 month were used for meristem cessation analysis. Poplar saplings were grown at 25 °C, and Arabidopsis (Arabidopsis thaliana) seedlings at 22 °C. The tobacco (Nicotiana benthamiana) plants used for subcellular localization analysis were grown at 25 °C. All three types of plant materials were grown under a 16-h light/8-h dark photoperiod. The Arabidopsis wus-1 and clv3-2 mutants were prepared as described previously[34,49].
In situ hybridization
-
Shoot tips, internodes, and leaf tips derived from sterile saplings cultured on rooting medium for 2 months were fixed in FAA (alcohol : formaldehyde : acetic acid : water, 10:2:1:7) at 4 °C overnight. The material was then dehydrated and embedded in Paraplast (Sigma-Aldrich). The full-length coding region containing a digaoxin label was synthesized in vitro, and used as a hybridization probe as previously described[23]. The sections with hybridization signals were viewed and imaged under an Olympus BX-51 microscope (Olympus, Tokyo, Japan).
Comparison of the stem cell niche area
-
The height, width, and area of CLV3/WUS expression domains, as well as the number of cells within these domains, were measured from the in situ hybridization images using ImageJ software as previously described[50,51]. The shoot meristem width was measured at the maximum width between leaf primordia.
Plasmid construction and genetic transformation
-
To construct overexpression vectors, the full-length coding sequences of PagCLV3-1, PagCLV3-2, PagWUS1, and PagWUS2 were amplified by PCR and cloned into the pROKII-GFP vector to obtain the 35S::PagCLV3-1-GFP, 35S::PagCLV3-2-GFP, 35S::PagWUS1-GFP, and 35S::PagWUS2-GFP vectors, respectively. CRISPR target sites were designed using the CRISPRdirect online tool (
http://crispr.dbcls.jp ), selecting sites on the basis of specificity, PAM sequence constraints, and minimal predicted off-target effects. The target sequence and the upstream and downstream 10-bp sequence were used as Blast queries against the poplar genome to ensure specificity. One target was designed in the upstream sequence of the open reading frame and one in the functional domain to improve the mutation efficiency. The target sequence with A or G at the 20th bp upstream of NGG was preferentially selected (Supplementary Fig. S1). The target sequence was amplified to construct the gsRNA expression cassette, which was then assembled into the vector pYLCRISPR/Cas9-DN (kindly provided by Yao-Guang Liu, South China Agricultural University)[52]. Sequence changes in transgenic plants were identified by PCR amplification and DNA sequencing (Supplementary Fig. S1). The 4,430-bp sequence immediately upstream of the ATG start codon of AtWUS was used as the promoter, and was amplified and cloned into the pROKII-GFP vector. The PagWUS1 coding sequence was cloned and inserted downstream of the AtWUS promoter to construct the pAtWUS::PagWUS1-GFP vector. The coding region of PagCLV3-2 was amplified and cloned into the PFK-321 vector (kindly provided by Zhong Zhao, University of Science and Technology of China) by the LR reaction to construct the pAtCLV3::PagCLV3-2 vector[31].To analyze the expression patterns of PagWUS2 and PagCLV3-1 at the transcriptional level, the 2,408-bp sequence immediately upstream of the ATG start codon of PagWUS2 was amplified by PCR and cloned into the PZP211-GFP3 vector to generate the pPagWUS2::GFP3 vector (where GFP3 represents the concatemer of three GFP proteins). The 6,186-bp promoter sequence was used in the pPagWUS2::GFP3-L vector. The 4,707-bp sequence immediately upstream of the ATG start codon of PagCLV3-1 was amplified by PCR and cloned into the PBI121-GUS vector to generate the pPagCLV3-1::GUS vector. To construct the pPagWUS2::GFP3 vector, the 4,707-bp promoter sequence of PagCLV3-1 was cloned upstream of GFP3 in the PZP211-GFP3 vector, and the 1,791-bp sequence downstream of the translational stop codon was added downstream of GFP3. The sequences of all primers are listed in Supplementary Table S1.
The vectors were transformed into poplar via Agrobacterium-mediated transformation, using leaves as explants. The floral dip method was used for Arabidopsis transformation.
ChIP-qPCR assay
-
Regenerated shoots of pPagRR13::PagRR13-GFP lines were used for ChIP-qPCR analyses. After harvest, 1 g tissue was cross-linked with 1% (v/v) formaldehyde under vacuum for 10 min. The cross-linking reaction was quenched with 0.125 M glycine for 5 min. The tissue was thoroughly ground in liquid nitrogen, and chromatin was extracted before sonication. Chromatin was sheared by sonication (15 min at 40% power for 10 s on/off) to generate DNA fragments between 200 bp and 1 kb. Of the DNA fragment mixture, 5% of the volume was saved as the input control. The anti-GFP antibody (TransGen Biotech, Beijing, China) was pre-incubated with Protein A/G Magnetic Beads (Selleckchem, Houston, TX, USA) at 4 °C for 2 h with gentle rotation. Chromatin was pre-cleared by incubation with the Protein A/G Magnetic Beads before adding antibody-bound beads, and then incubated overnight at 4 °C. The beads were successively washed with low-salt washing buffer (150 mM NaCl, 0.1% [w/v] SDS, 1% [v/v] Triton X-10, 2 mM EDTA [pH 8.0], and 20 mM Tris-HCl [pH 8.0]), high-salt washing buffer (500 mM NaCl, 0.1% [w/v] SDS, 1% [v/v] Triton X-10, 2 mM EDTA [pH 8.0], and 20 mM Tris-HCl [pH 8.0]), LiCl washing buffer (0.25 M LiCl, 1% [v/v] NP-40, 1% [w/v] sodium deoxycholate, 1 mM EDTA [pH 8.0], and 10 mM Tris-HCl [pH 8.0]), and TE buffer (1 mM EDTA [pH 8.0] and 10 mM Tris-HCl [pH 8.0]). Finally, the immunoprecipitated chromatin was eluted with elution buffer (1% [w/v] SDS and 0.1 M NaHCO3). The immunoprecipitation complex was dissociated by adding 5 M NaCl and stirring using a Thermo Mixer at 700 rpm for at least 8 h or overnight. The DNA was recovered using ChIP DNA Clean & Concentrator (Zymo Research, Irvine, CA, USA) and analyzed by qRT-PCR as described previously[23,53]. The sequences of all primers are listed in Supplementary Table S1.
qRT-PCR
-
Total RNA was extracted from shoot tips and explants using the cetyl trimethyl ammonium bromide (CTAB) method. HiScriptII qRT Super Mix (Vazyme, Nanjing, China) was used to obtain cDNA, and 1–2 μg total RNA was used for cDNA construction. qRT-PCR was performed on a LightCycler 96 system (Roche, Basel, Switzerland) using Cham Q SYBR qPCR Master Mix (Vazyme) with gene-specific primers. Data are mean ± sd of three independent biological repeats. Data were analyzed by two-tailed Student’s t-tests, and *** indicates significant difference at p < 0.001. The housekeeping gene TUBULIN was employed as an internal reference. All primers are listed in Supplementary Table S1.
Shoot regeneration analysis
-
Poplar saplings were grown under sterile conditions at 25 °C with a 16-h light/8-h photoperiod for 4–6 weeks. The 3rd to 5th leaves from the shoot tip were excised and cultured on differentiation medium (Murashige and Skoog medium [MS] + 30 g/L sucrose + 0.5 mg/L 6-benzyl aminopurine [6-BA] + 0.1 mg/L naphthaleneacetic acid [NAA] + 0.002 mg/L thidiazuron [TDZ] + 8 g/L agar) under light intensity of 100 μmol/m2/s. The number of regenerated shoots was counted after 30 d of culture. Arabidopsis seeds were germinated on ½ MS medium at 22 °C under a 16-h light/8-h dark photoperiod, and then grown for 7 d before excising cotyledons for culture. Cotyledons were cultured on callus induction medium (CIM) (MS + 0.5 g/L 2-(N-morpholino) ethane sulfonic acid hydrate [MES] + 10 g/L sucrose + 0.5 mg/L 2,4-dichlorophenoxyacetic acid [2,4-D] + 0.05 mg/L kinetin [KT] + 8 g/L agar) for 6 d before transfer onto shoot induction medium (SIM) (MS + 0.5 g/L MES + 10 g/L sucrose + 2.5 μM 2-isopentenyladenine [2-IP] + 0.9 μM indoleacetic acid [IAA] + 8 g/L agar) for shoot induction under continuous light at 100 μmol/m2/s. The number of regenerated shoots was counted after 21 d of culture on SIM (SIM21)[54]. The data are presented as mean ± sd from independent biological replicates. For each analysis, n > 20.
PagCLV3-2 peptide in vitro treatment
-
The PagCLV3-2 peptide (ELRAVPSGPDPLHH) was artificially synthesized (Sangon Biotech, Shanghai, China)[55]. The shoots of the CRISPR-PagCLV3-2 plants showing fasciation and phyllotaxis phenotypes were cultured on differentiation medium (MS + 30 g/L sucrose + 0.5 mg/L 6-BA + 0.1 mg/L NAA + 0.002 mg/L TDZ + 50 mg/L kanamycin + 200 mg/L timentin + 8 g/L agar) containing different concentrations of the PagCLV3-2 peptide. Complementation of the fasciation phenotype was analyzed after 30 d of culture.
Subcellular localization analyses
-
The 35S::PagCLV3-1-GFP, 35S::PagCLV3-2-GFP, 35S::PagWUS1-GFP, 35S::PagWUS2-GFP, or 35S::CBL1-RFP (kindly provided by Sha Li, Shandong Agricultural University) vector was transformed into fully expanded leaves of N. benthamiana via Agrobacterium tumefaciens GV3101[56]. The 35S::PagCLV3-1-GFP and 35S::PagCLV3-2-GFP vectors were co-infiltrated with the plasma membrane marker 35S::CBL1-RFP. After incubation in the dark for 48 h, fluorescence was viewed and imaged under a Zeiss LSM 880 NLO confocal microscope (Carl Zeiss, Jena, Germany). The GFP signal was detected at 488 nm, and the DAPI signal at 405 nm[57].
GUS staining
-
Shoot tips of pPagCLV3-1::GUS saplings were fixed in 90% [v/v] acetone on ice for 20 min. After washing twice with a buffer containing 50 mM NaPO4 (pH 7.2), 0.5 mM K3Fe (CN)6, and 0.5 mM K4Fe (CN)6, the tissue was immersed in β-glucuronidase (GUS) staining buffer containing 2 mM X-Gluc and vacuum-infiltrated for 10 min. After incubation at 37 °C for 12 h, the stained material was washed with 70% ethanol solution for 12 h, then embedded in low-melting-point agarose and sectioned. The sections were viewed and imaged under an Olympus BX-51 microscope.
Confocal microscopy
-
Shoot tips were fixed in 2.5% (v/v) paraformaldehyde on ice and vacuum-infiltrated for 30 min. After fixing at 4 °C overnight, the tissue was successively washed with 10%, 20%, and 30% sucrose solutions (dissolved in 1% [v/v] paraformaldehyde). The tissues were then embedded in low-melting-point agarose, sectioned, and viewed and imaged under a Zeiss LSM 880 NLO confocal microscope (Carl Zeiss). The GFP signal was detected with excitation at 488 nm and emission at 505–550 nm. The cell outline was stained with Fluorescent Brightener, and its signal was detected with excitation at 405 nm, and emission at 425–475 nm[58].
-
To identify the putative orthologs of Arabidopsis WUS and CLV3 in poplar, we used the protein sequences of AtWUS and AtCLV3 as queries in Blast searches against the genome of Poplar 84K (Populus alba × Populus tremula var. glandulosa) (Supplementary Table S2). Consistent with a previous report, two sequences demonstrated the highest conservation with AtWUS, and were thus annotated as PagWUS1 and PagWUS2 (Supplementary Fig. S2)[59]. Two CLAVATA3/Embryo Surrounding Region-related (CLE) family proteins containing putative N-terminal signal peptides showed the highest similarity to AtCLV3, and were annotated as PagCLV3-1 and PagCLV3-2 (Supplementary Fig. S3, Supplementary Table S3)[60−62].
We expressed PagWUS1 under the control of the AtWUS promoter in the Arabidopsis wus-1 mutant[63]. The shoot development phenotype of wus-1 seedlings was largely rescued (Supplementary Fig. S4a, S4b), and the defects in the floral organs of wus-1 were also substantially complemented (Supplementary Fig. S4c). Introduction of the pAtCLV3::PagCLV3-2 vector into the clv3-2 mutant partially rescued its defective seedling, silique, and flower development (Supplementary Fig. S5). The above results indicate that PagWUS1 and PagCLV3-2 have similar functions to AtWUS and AtCLV3, respectively.
PagWUS1/2 and PagCLV3-1/2 are specifically expressed in the shoot meristem
-
To examine the distribution of PagWUS1/2 and PagCLV3-1/2 transcripts, in situ hybridization was performed using cross sections and longitudinal sections of the meristem, internodes, and leaves. Transcripts of both PagWUS1 and PagWUS2 were present in the OC of the shoot meristem, but not in the cross section of the third internode and the longitudinal section of the young leaf (Fig. 1a–c). The PagCLV3-1 and PagCLV3-2 transcript signals were specifically localized in the CZ of the shoot meristem, but were not visible in the fifth internode or the cross section of the young leaf (Fig. 1d–f).
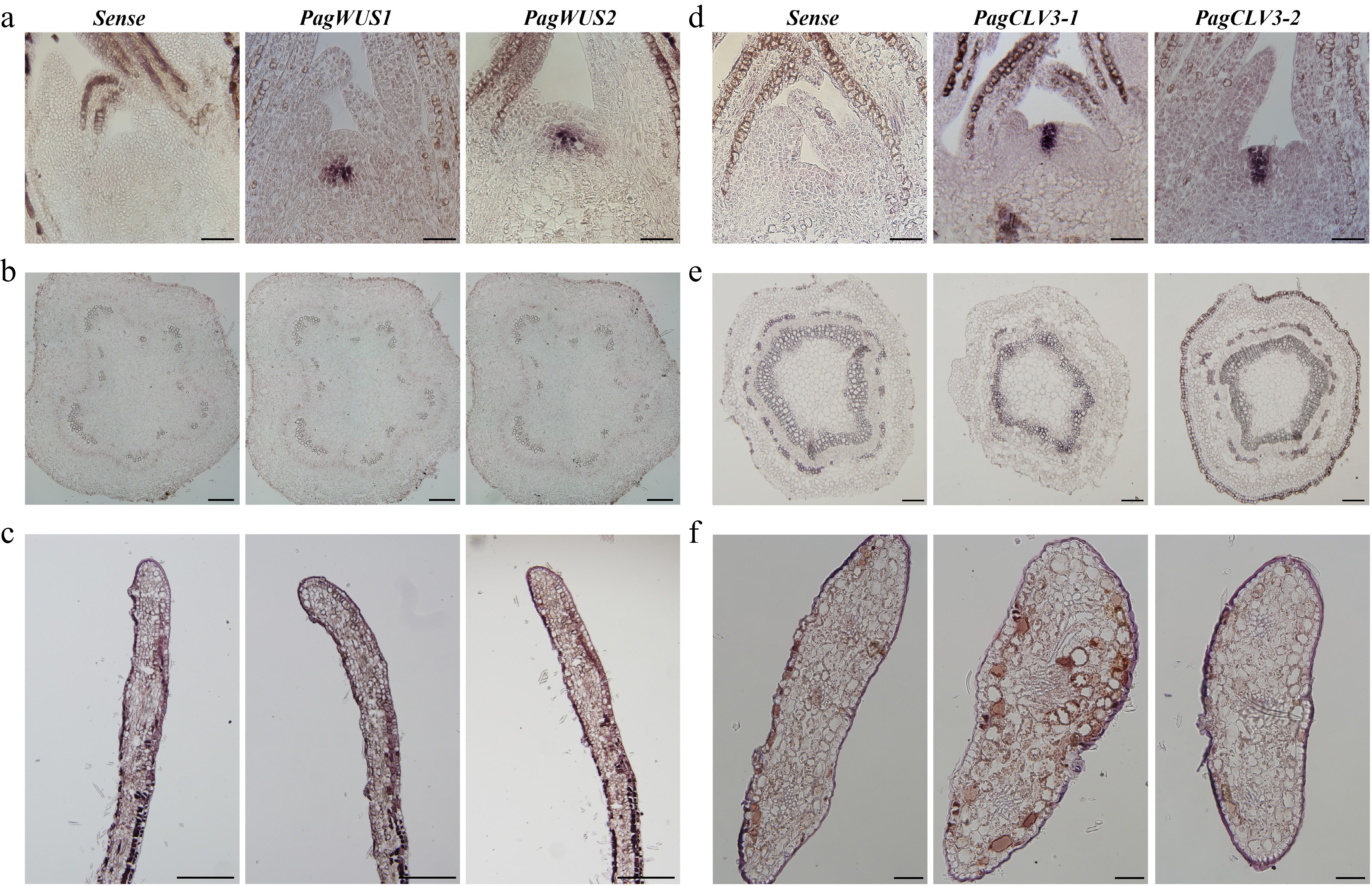
Figure 1.
Expression patterns of PagWUS1/2 and PagCLV3-1/2 as detected by in situ hybridization. Signals of PagWUS1 and PagWUS2 transcripts were (a) present in the organizing center (OC) of shoot meristem, but (b) absent from the cross section of third internode, and (c) longitudinal section of young leaf. Signals of PagCLV3-1 and PagCLV3-2 transcripts were (d) localized in the central zone (CZ) of shoot meristem, but (e) absent from the cross section of fifth internode, and (f) cross section of young leaf. Sense probe controls are shown in the left column. Bar = 100 μm.
In the Arabidopsis shoot meristem, AtCLV3 is expressed in the center of the outermost three cell layers, while AtWUS is expressed in the three-layered OC beneath[6,10]. However, the PagCLV3-1/2 signals were present in six cell layers, and PagWUS1/2 signals were present in four to five cell layers (Supplementary Fig. S6a–S6d). Cells expressing AtCLV3 and AtWUS, which constitute the stem cell niche in Arabidopsis, accounted for 10.1% of the total cell number of the shoot meristem (Supplementary Fig. S6e), whereas this proportion was much higher (20.9%) in poplar. Moreover, the ratios of height, width, and area of the PagCLV3-1/PagWUS-expressing domain to those of the poplar meristem were significantly higher than those of the AtCLV3/AtWUS-expressing domain to the Arabidopsis meristem (Supplementary Fig. S6f, S6g). Estimates based on analyses of the 2D sections indicate that, during vegetative growth under normal conditions, the stem cell niche accounts for a larger proportion of the meristem in poplar than in Arabidopsis.
We then tried to visualize the expression patterns of PagWUS and PagCLV3 using reporter lines. The 2,408-bp or 6,186-bp sequences immediately upstream of the ATG start codon of PagWUS2 were used as promoters to generate the PagWUS2::GFP3 and PagWUS2::GFP3-L lines, respectively. The results show that GFP signals were distributed beyond the OC into the PZ or leaf primordium (Supplementary Fig. S7a, S7b). The 4,707-bp upstream sequence of PagCLV3-1 gave rise to expression signals in the OC (Supplementary Fig. S7c). After adding the 1,791-bp sequence downstream of the translational stop codon, GFP signals were still mainly distributed in the OC (Supplementary Fig. S7d). These results suggest that the tested regulatory fragments of PagWUS2 and PagCLV3-1 may be insufficient. Additional approaches are required to capture the authentic promoter regions.
To determine the subcellular localization of PagWUS1 and PagWUS2, 35S::PagWUS1-GFP and 35S::PagWUS2-GFP vectors were separately transformed into tobacco leaf epidermal cells. Consistent with their proposed roles as transcription factors, the PagWUS1-GFP and PagWUS2-GFP signals overlapped with those of DAPI, confirming their localization in the nucleus (Supplementary Fig. S8a). We next co-infiltrated the 35S::PagCLV3-1-GFP and 35S::PagCLV3-2-GFP vectors with the plasma membrane marker 35S::CBL1-RFP, respectively[56]. Both the GFP and RFP signals were localized along the cellular outline (Supplementary Fig. S8b). To determine whether PagCLV3-1/2 were localized in the plasma membrane or exported into the extracellular space, tobacco leaves were treated with a sucrose solution to induce plasmolysis. The PagCLV3-1-GFP and PagCLV3-2-GFP signals were partially separated from CBL1-RFP and localized in the apoplast, indicating that PagCLV3-1 and PagCLV3-2 are secreted proteins.
PagWUS1/2 and PagCLV3-1/2 regulate the maintenance of the shoot meristem by forming a feedback loop
-
To study the biological functions of PagWUS1 and PagWUS2, we first overexpressed these two genes under the control of the cauliflower mosaic virus 35S promoter. There were two phenotypes among the 35S::PagWUS1/2 lines (Fig. 2a, b). In total, we obtained four Type I and three Type II 35S::PagWUS1 lines, and four Type I and four Type II 35S::PagWUS2 lines. Different lines from the same type exhibited consistent and stable phenotypes. We selected two lines with different phenotypic types of 35S::PagWUS1 and 35S::PagWUS2, respectively, for further analysis. Compared with the wild type, the Type I lines with relatively lower PagWUS1/2 transcript levels grew slowly with reduced plant height (Fig. 2a, c). The shoot meristem was fasciated, which generated a fasciated stem with disordered phyllotaxis, indicating the production of supernumerary meristematic cells (Fig. 2d). The Type II lines demonstrated stronger phenotypes in terms of growth retardation and stem fasciation, consistent with the relatively higher PagWUS1/2 transcript levels.
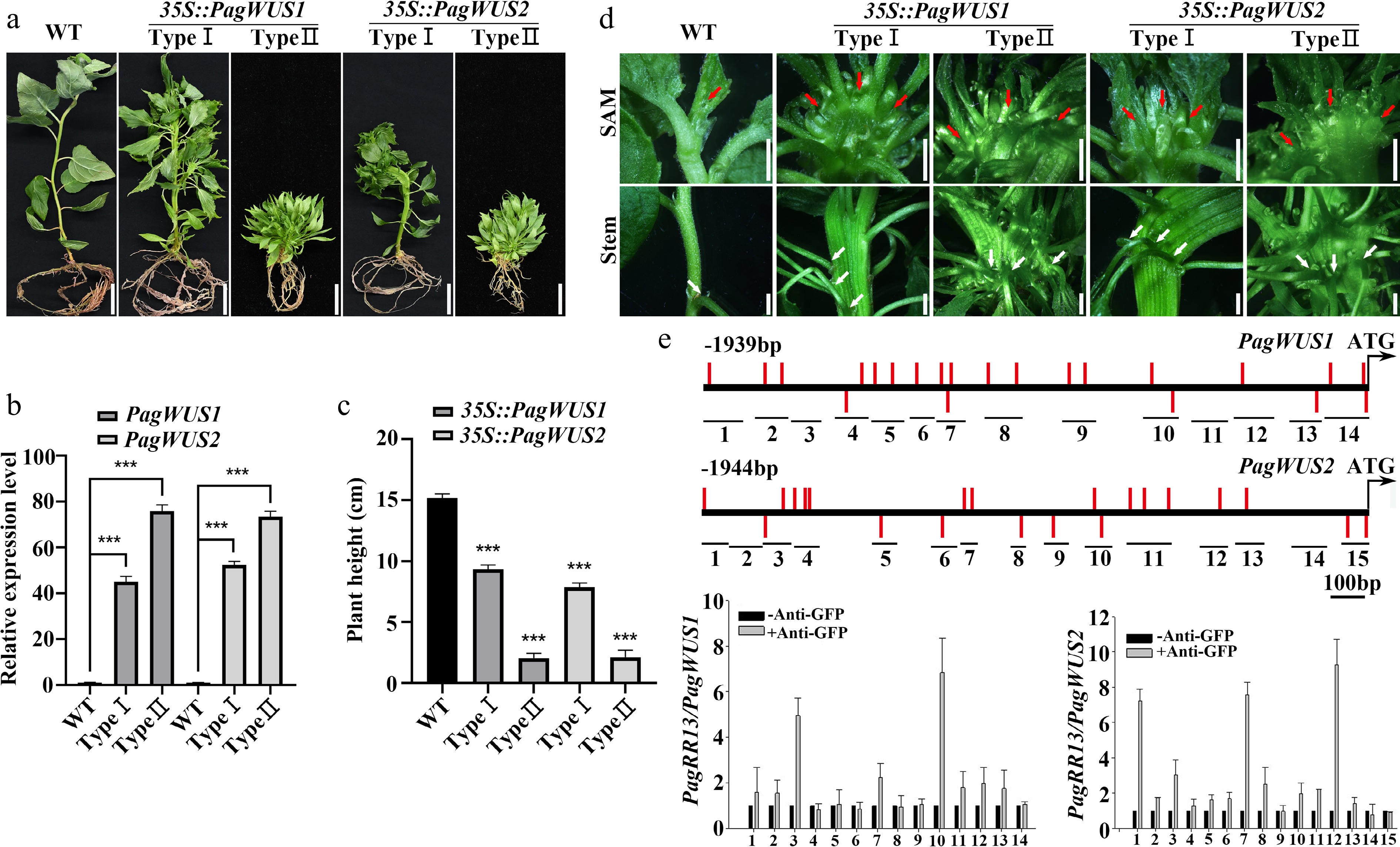
Figure 2.
PagWUS1/2 positively regulate shoot meristem activity. (a) Phenotype of 35S::PagWUS1 and 35S::PagWUS2 seedlings. Bar = 2 cm. (b) Correlation between relative transcript levels of PagWUS1/2 and phenotype severity of overexpressing lines. (c) Plant height of 35S::PagWUS1 and 35S::PagWUS2 saplings. (d) Fasciated shoot meristem and stem with disordered phyllotaxis in 35S::PagWUS1 and 35S::PagWUS2 lines. Red arrows indicate fasciated shoot meristem. White arrows denote disordered phyllotaxis. Bar = 2 mm. (e) ChIP-qPCR results showing association between PagRR13 and PagWUS1/2 promoter fragments. Scheme of promoter regions of PagWUS1 and PagWUS2 is shown above. Black short lines with numbers indicate positions of corresponding fragments used for ChIP-qPCR. Red bars indicate type-B RR binding elements GAT(T/C). Data are mean ± sd of three independent biological repeats. *** Indicates significant difference at p < 0.001 (two-tailed Student’s t-tests).
Neither the CRISPR-PagWUS1 line nor the CRISPR-PagWUS2 line showed any visible phenotype, possibly because of functional redundancy. We were unable to obtain CRISPR-PagWUS1-2 saplings, suggesting that disruption of both genes may compromise shoot regeneration (Supplementary Fig. S1).
Additionally, ChIP-qPCR analysis revealed that PagRR13 (a homolog of the AtARR1 transcription factor) associated with the promoter regions of both PagWUS1 and PagWUS2 (Fig. 2e). The results of the dual-luciferase assays showed that PagRR13 activated the expression of PagWUS1/2, indicative of direct regulation between cytokinin signaling and PagWUS1/2 transcription (Supplementary Fig. S9).
We further explored the functions of PagCLV3-1/2 through overexpression and knock-out (CRISPR/Cas9) strategies. The 35S::PagCLV3-1/2-overexpressing lines exhibited two types of phenotypes (we obtained five Type I and three Type II 35S::PagCLV3-1 lines, and six Type I and three Type II 35S::PagCLV3-2 lines) (Fig. 3a, b; Supplementary Fig. S10). For each type, two independent lines were selected for phenotypic analysis. The saplings with relatively lower PagCLV3-1/2 transcript levels grew in a 'stop-go' manner (Type I). After a period of growth, the shoot meristem vanished, and the meristematic cells were consumed (Fig. 3a; Supplementary Figs S11 and S12). About 10 d later, the shoot meristem was reformed. During this process, a number of axial buds grew out and formed branches, indicating compromised apical dominance resulting from the cessation of meristem activity. In saplings with relatively higher PagCLV3-1/2 transcript levels (Type II), shoot meristem activity ceased at early stages, resulting in a disorganized bunch of leaves and no stem elongation (Fig. 3a, b). Therefore, overexpression of PagCLV3-1/2 repressed shoot meristem activity.

Figure 3.
PagCLV3 negatively regulates shoot meristem maintenance. (a) Phenotype of 35S::PagCLV3-1 seedings. White arrow denotes position of vanished shoot meristem. Yellow arrows indicate axial bud outgrowths. (b) Correlation between relative transcript levels of PagCLV3-1 in 35S::PagCLV3-1 lines and phenotype severity. Data are mean ± sd of three independent biological repeats. (c) Phenotype of CRISPR-PagCLV3-2 seedlings. (d) Fasciation phenotypes of saplings in (c). (e) Phenotype of CRISPR-PagCLV3-1-2 (f) Fasciation phenotypes of saplings in (e). In (d) and (f), red arrows indicate shoot meristem. For (a), bar = 2 cm. For (c) and (e), bar = 2 cm. For (d) and (f), bar = 2 mm. Data are mean ± sd of three independent biological repeats. *** indicates significant difference at p < 0.001 (two-tailed Student’s t-tests).
In contrast, the CRISPR-PagCLV3-2 saplings (eight Type I and three Type II lines) generated a fasciated shoot meristem, leading to a fasciated stem and random phyllotaxis (Fig. 3c, d; Supplementary Figs S1 and S13). To confirm that the fasciation and phyllotaxis defects in CRISPR-PagCLV3-2 mutants resulted from the loss of PagCLV3-2, we applied a synthetic PagCLV3-2 peptide (0.1, 1, and 10 μM) to mutate shoot apices in vitro. This largely rescued the fasciation and phyllotaxis phenotypes (Supplementary Fig. S14). Mutation of both PagCLV3-1 and PagCLV3-2 in the CRISPR-PagCLV3-1-2 lines (nine Type I and four Type II lines) resulted in more severe phenotypic defects than those of single mutants (Fig. 3e, f). Thus, similar to overexpression of PagWUS1/2, defects in PagCLV3-1/2 led to supernumerary meristematic cells.
To further determine the regulatory relationship between PagWUS1/2 and PagCLV3-1/2 transcription, we examined the transcript signals of PagWUS1/2 in the 35S::PagCLV3-2 shoot meristem by in situ hybridization. The results show that PagWUS1/2 transcripts were present at considerably reduced levels in the active meristem and undetectable in the inactive meristem (Fig. 4a). In the fasciated CRISPR-PagCLV3-2 shoot meristem, PagWUS1/2 were ectopically expressed in a band-like region beneath the outermost two cell layers (Fig. 4a). These results show that PagCLV3-1/2 negatively regulate the expression of PagWUS1/2. As determined by qRT-PCR analyses, the transcript levels of PagCLV3-1/2 were significantly increased in the shoot tips of both 35S::PagWUS1 and 35S::PagWUS2 saplings, indicating that PagWUS1/2 activate the transcription of PagCLV3-1/2 (Fig. 4b). Together, these results show that PagWUS1/2 and PagCLV3-1/2 maintain the shoot meristem by forming a feedback loop.
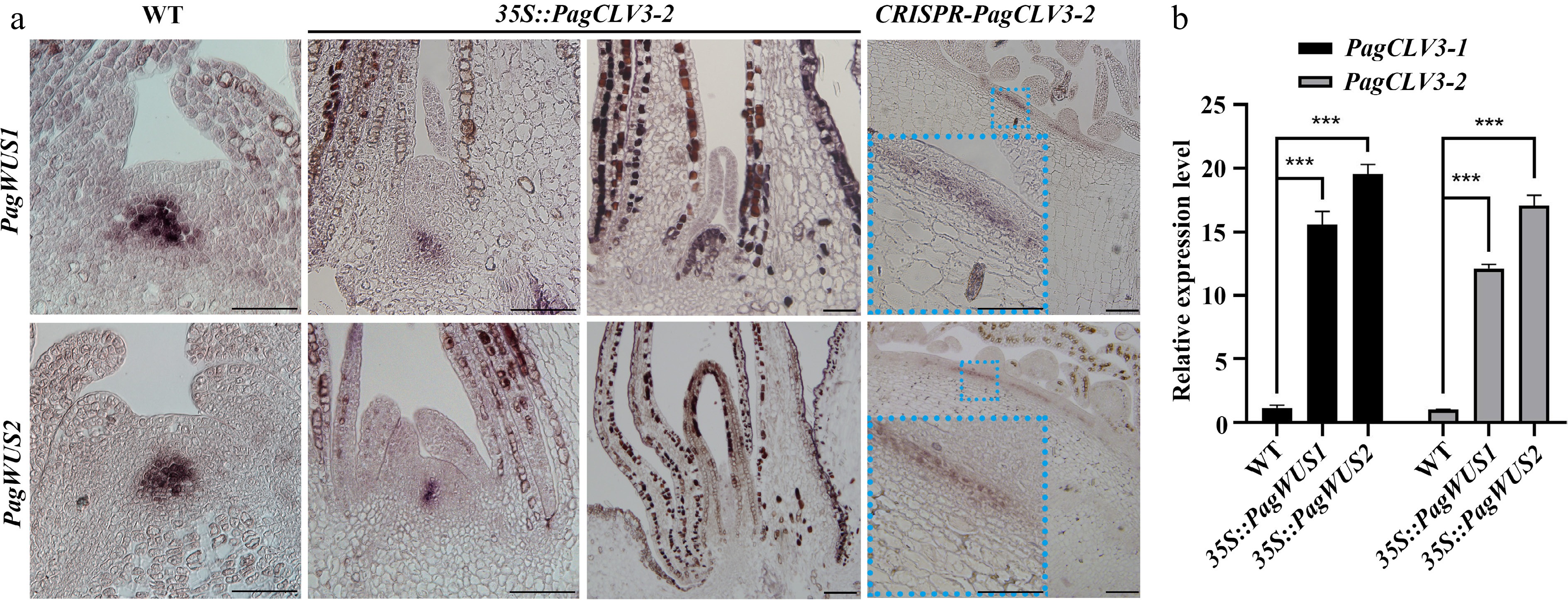
Figure 4.
Negative feedback regulation between transcription of PagWUS and PagCLV3. (a) In situ hybridization of PagWUS1 and PagWUS2 transcripts in shoot meristem of wild-type, 35S::PagCLV3-2, and CRISPR-PagCLV3-2 saplings. Lower left insets in the right column are magnified images of blue dotted rectangles. Bar = 100 μm. (b) qRT-PCR analyses showing increased PagCLV3-1/2 transcript levels in shoot tips of 35S::PagWUS1 and 35S::PagWUS2 saplings. Data are mean ± sd of three independent biological repeats. *** Indicates significant difference at p < 0.001 (two-tailed Student’s t-tests).
Involvement of the PagWUS-PagCLV3 module in shoot regeneration
-
Next, we examined whether PagWUS1/2 and PagCLV3-1/2 participate in shoot regeneration. When wild-type leaves were used as explants, an average of 141 shoots were produced from one leaf after 30 d of incubation (Fig. 5a, d). In the 35S::PagWUS1 and 35S::PagWUS2 lines, this number was increased to 285 and 293, respectively. By contrast, overexpression of PagCLV3-1 or PagCLV3-2 significantly reduced shoot regeneration (Fig. 5b, d).
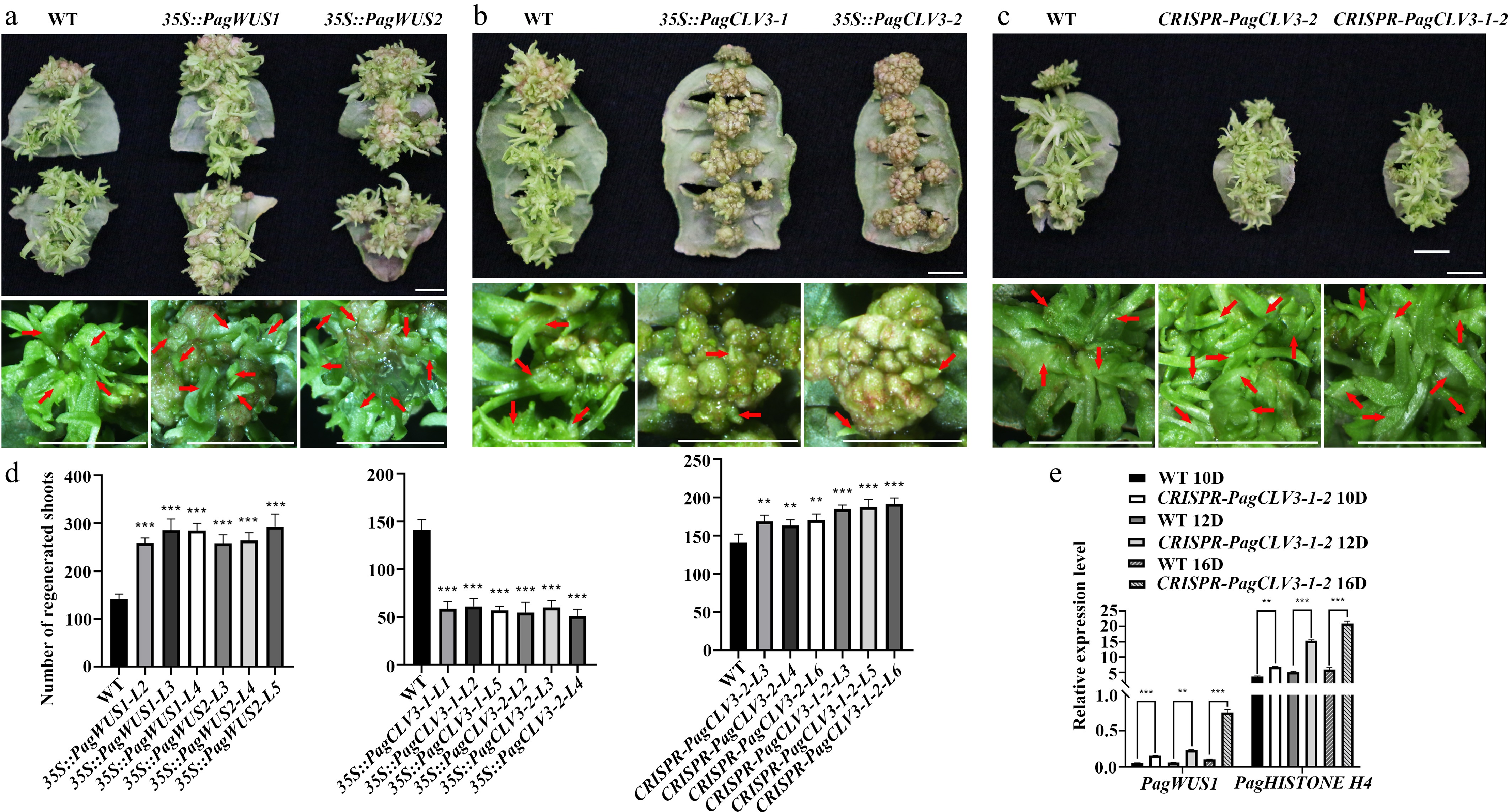
Figure 5.
Participation of PagWUS-PagCLV3 module in shoot regeneration. Shoot regeneration was promoted in (a) 35S::PagWUS, repressed in (b) 35S::PagCLV3, and enhanced in the (c) CRISPR-PagCLV3 explants. Bars = 5 mm. For (a)–(c), upper panels show shoots regenerated from leaf explants. Lower panels show formation of regenerated shoots. Red arrows indicate position of regenerated shoots. (d) Numbers of regenerated shoots per leaf explant. For 35S::PagWUS1, n = 25; 35S::PagWUS2, n = 25; 35S::PagCLV3-1, n = 25; 35S::PagCLV3-2, n = 25; CRISPR-PagCLV3-2, n = 25; CRISPR-PagCLV3-1-2, n = 25. Error bars show mean ± sd. (e) qRT-PCR analyses showing increased PagWUS1 and Histone H4 transcript levels in CRISPR-PagCLV3-1-2 explants, as compared with wild type. D indicates days of culture. Data are mean ± sd of three independent biological repeats. Asterisks indicate significant differences: ** 0.001 < p < 0.01 and *** p < 0.001 (two-tailed Student’s t-tests).
Unexpectedly, the CRISPR-PagCLV3-2 and CRISPR-PagCLV3-1-2 lines also showed enhanced shoot regeneration capacity (Fig. 5c, d). By contrast, the Arabidopsis clv3 mutant did not show a similar phenotype (Supplementary Fig. S15). Previous studies revealed that CLV3 negatively regulates WUS expression and restricts cell division[64]. We thus postulate that during the reestablishment of the shoot meristem, the defective PagCLV3-1/2 released the constraints on PagWUS1/2 expression and cell proliferation, and thus promoted shoot meristem formation. To test this hypothesis, we examined the transcript levels of PagWUS1 and Histone H4 (a marker for cell division) by qRT-PCR (Fig. 5e; Supplementary Fig. S16). The results show that, compared with the wild type, the transcript levels of PagWUS1 and H4 were significantly increased at day 10 of culture on SIM (SIM10), SIM12, and SIM16.
PagWUS-PagCLV3 module controls shoot meristem cessation
-
Next, we investigated whether PagWUS1/2 expression is responsive to seasonal changes. For this purpose, in situ hybridization analyses were performed on shoot meristems collected in different seasons. The results show that both PagWUS1 and PagWUS2 were actively expressed during the growth season (Fig. 6a–c). However, the transcript signals were obviously weaker at the onset of growth cessation and bud set, and completely absent after bud scale maturation. Thus, the transcript levels of PagWUS1 and PagWUS2 were correlated with the shift in meristem activity toward dormancy.

Figure 6.
The PagWUS-PagCLV3 module regulates the cessation of shoot meristem activity. In situ hybridization analyses showing abundance of PagWUS1 and PagWUS2 transcripts: (a) abundant during growth season, (b) reduced at the onset of cessation, and (c) absent after bud scale maturation. Bar = 100 μm. After short day treatment for 4 weeks, shoot meristem became inactive in (d) wild-type, but remained active in (e) 35S::PagWUS1, (f) 35S::PagWUS2, (g) CRISPR-PagCLV3-2, and (h) CRISPR-PagCLV3-1-2 saplings. Red arrows indicate shoot meristem. Bar = 1 cm.
We wondered whether the PagWUS-PagCLV3 module is implicated in regulating the cessation of shoot meristem activity. To address this question, we first determined whether growth cessation of the shoot is controlled by day length or temperature. Consistent with previous studies, we found that short-day (8-h light/16-h dark photoperiod) but not low temperature (4 °C) conditions for 7 weeks led to growth cessation in wild-type saplings (Supplementary Fig. S17)[65]. Next, we tested whether overexpression of PagWUS1/2 or disruption of PagCLV3-1-2 affected the cessation of meristem activity. Saplings of the 35S::PagWUS1/2, CRISPR-PagCLV3-2, and CRISPR-PagCLV3-1-2 lines were cultured under short-day conditions. After 4 weeks, meristem activity had ceased in the wild-type but persisted in all the transgenic lines, which continually formed new leaves (Fig. 6d–h; Supplementary Fig. S18). These results indicate that PagWUS1/2 positively regulates meristem activity, whereas PagCLV3-1/2 has the opposite role.
PagWUS-PagCLV3 module mediates proper pattern of secondary growth
-
Because mutation of PagCLV3-1/2 or overexpression of PagWUS1/2 gave rise to a fasciated stem, we wondered whether secondary growth was affected in these lines. To address this question, we compared the histological structure of the stem between the wild-type and CRISPR-PagCLV3-1-2 and 35S::PagWUS1 lines. In the cross sections of the wild-type stem, vascular bundles were evenly distributed in the fourth internode, while the secondary phloem and xylem were radially thickened in the tenth internode (Supplementary Fig. S19a, S19b). In the CRISPR-PagCLV3-1-2 stem, it was difficult to distinguish two adjacent internodes because of the disrupted phyllotaxis. Therefore, we observed cross sections of the upper (5 mm from the shoot tip) and lower (5 cm from the shoot tip) parts of the stem. In the upper part, vascular bundles were evenly distributed in the fasciated stem (Supplementary Fig. S19c). However, in contrast to the wild-type’s stem, that of the CRISPR-PagCLV3-1-2 mutant showed uneven thickening in the lower part, i.e., the secondary phloem and xylem were clearly formed at one end, but the other end was still in the primary growth stage (Supplementary Fig. S19d). The 35S::PagWUS1 stem demonstrated a similar histological structure to that of CRISPR-PagCLV3-1-2 (Supplementary Fig. S19e, S19f). These results show that the PagWUS-PagCLV3 module mediates the proper pattern of secondary growth by maintaining the shoot meristem.
-
More than two decades ago, the WUS-CLV3 module was established as the pivotal circuit regulating the balance between stem cell homeostasis and differentiation of the shoot meristem[6,10]. Since then, WUS and CLV3 orthologs have been characterized and analyzed in a number of herbaceous species. Their biological roles have been found to be largely conserved in dicots, including Arabidopsis, Brassica rapa, tomato (Solanum lycopersicum), Medicago truncatula, Antirrhinum majus, and petunia (Petunia hybrida)[9,66−71]. However, the regulatory roles of the WUS-CLV3 module in perennial woody plants remain unknown.
In this study, we investigated the functions of the PagWUS-PagCLV3 module. The results demonstrate conserved as well as deviated functions compared with those of their orthologs in herbaceous species. PagWUS and PagCLV3 maintain the shoot meristem by forming a feedback loop, similar to the case in herbaceous species[72].
Different from its herbaceous orthologs, the PagWUS-PagCLV3 module is consistent with the developmental characteristics of perennial trees by regulating bud dormancy and secondary growth. In perennial plants, bud dormancy is a strategy to cope with seasonal changes[39]. The induction and signal transduction mechanisms related to dormancy have been studied in detail[48]. Despite these studies, how shoot meristem activity is regulated during the induction of dormancy remains elusive. We found that the transcript levels of PagWUS1/2 were correlated with meristem activity in poplar. Their transcript signals were abundant during the growth season, reduced at the onset of growth cessation, and completely absent after bud scale maturation (Fig. 6). Furthermore, overexpression of PagWUS1/2 or disruption of PagCLV3-1/2 delayed bud dormancy. These findings suggest that the PagWUS-PagCLV3 module acts as the mediator between dormancy regulators and shoot meristem activity.
Moreover, disrupting the PagWUS-PagCLV3 module gave rise to a fasciated stem in which the progress of secondary growth was uneven (Supplementary Fig. S19). We speculate that this phenotype was caused by the disruption of the radial distribution of hormones such as cytokinin and auxin, which are essential for cambium activity[73]. Whether the distribution patterns of these hormones and their signaling transduction are directly regulated by PagWUS or PagCLV3 is unknown. However, our results show that the PagWUS-PagCLV3 module influences the proper pattern of secondary growth in poplar through maintaining the appropriate structure and function of the shoot meristem.
Previous studies reported that overexpression of WUS facilitated the formation of somatic embryo and shoot structure[74−76]. In contrast, whether and how CLV3 is involved in shoot regeneration remained unknown. Consistently, our results show that shoot regeneration in poplar was significantly enhanced in the 35S::PagWUS lines. However, whereas the Arabidopsis clv3 single mutant did not show any changes in shoot regeneration phenotype, the CRISPR-PagCLV3 lines showed a clearly enhanced regeneration capacity. Because PagCLV3 and AtCLV3 have similar molecular functions (Supplementary Figs S5, S15), we speculate that the regulatory network of shoot regeneration differs between poplar and Arabidopsis. The results imply that silencing of CLV3 may be a strategy for establishing a high-efficiency shoot regeneration system in trees.
To our knowledge, this is the first report on the function of the WUS-CLV3 module in a perennial woody plant. The results reveal that the PagWUS-PagCLV3 module plays a basic role in regulating homeostasis of the shoot meristem in a manner conserved among dicots, and mediates the proper pattern of secondary growth and bud dormancy consistent with the typical characteristics of perennial trees. Our findings provide new information about the function and regulatory mechanisms of the shoot meristem in perennial woody species.
-
We investigated the functions of the poplar genes PagWUS and PagCLV3, homologs of Arabidopsis WUS and CLV3, respectively. Similar to their orthologs in dicotyledonous herbs, PagWUS and PagCLV3 form a feedback loop that regulates shoot meristem maintenance. Overexpression of PagWUS promoted shoot regeneration. Compared with herbaceous species, poplar has a much larger stem cell niche. Disruption of PagCLV3 enhanced the shoot regeneration capacity. Our results show that the PagWUS-PagCLV3 module controls the cessation of shoot meristem activity and mediates the proper pattern of secondary growth. These findings provide insights into the mechanisms of shoot meristem regulation in trees.
-
The authors confirm their contributions to the paper as follows: conceived and designed the experiments: Sang YL, Cheng ZJ, Zhang XS; performed the experiments: Dong WC; data analysis: Wang FL, Li XT, Wu XX, Yu LF, Luan LL, Wang Y; written manuscript: Sang YL, Zhang XS. All authors reviewed the results and approved the final version of the manuscript.
-
All the data supporting the findings of this study are available in the paper and supplementary data.
-
We thank Dr. EM Meyerowitz (California Institute of Technology), Dr. T Laux (University of Freiburg), Dr. Yao-Guang Liu (South China Agricultural University) and Dr. Zhong Zhao (University of Scienceand Technology of China) for providing materials. This work wassupported by the Key R&D Program of Shandong Province, China (2024LZGC025), National Excellent Young Talents Support Program, Taishan Scholar Youth Expert Project, National Natural Sciences Foundation of China (31870178 and 32270333) and Provincial Key Research and Development Program of Shandong (2021LZGC021). We thank Jennifer Smith, PhD, from Liwen Bianji (Edanz) (www.liwenbianji.cn) for editing the English text of drafts of this manuscript.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in this study.
- Supplementary Table S2 Protein sequences of AtWUS and AtCLV3.
- Supplementary Table S3 Signal peptides predicted by SignalP (v6.0).
- Supplementary Fig. S1 Targets of CRISPR-PagCLV3 and CRISPR-PagWUS lines.
- Supplementary Fig. S2 Structural and phylogenetic analysis of PagWUS1 and PagWUS2.
- Supplementary Fig. S3 Structural and phylogenetic analysis of PagCLV3-1 and PagCLV3-2.
- Supplementary Fig. S4 Expressing pAtWUS::PagWUS1-GFP largely rescues the phenotype of the wus1 mutant.
- Supplementary Fig. S5 Introducing the pAtCLV3::PagCLV3-GFP vector partially rescues the clv3-2 mutant phenotype.
- Supplementary Fig. S6 Proportion of cells constituting the stem cell niche in the total number of shoot meristem cell is larger in poplar than in Arabidopsis.
- Supplementary Fig. S7 Expression patterns of reporters with different lengths of regulatory sequences.
- Supplementary Fig. S8 Subcellular localization of PagWUS1/2 and PagCLV3-1/2.
- Supplementary Fig. S9 PagRR13 activates PagWUS1/2 transcription.
- Supplementary Fig. S10 PagCLV3-2 negatively regulated shoot meristem homeostasis.
- Supplementary Fig. S11 The 'stop-and-go' growth manner of Type Ⅰ 35S::PagCLV3-1 saplings.
- Supplementary Fig. S12 The process of shoot meristem consumption in Type Ⅰ 35S::PagCLV3-1 saplings.
- Supplementary Fig. S13 Morphology of fasciated shoot meristem in CRISPR-PagCLV3 saplings.
- Supplementary Fig. S14 Exogenous PagCLV3-2 rescued the phenotype of CRISPR-PagCLV3-2 shoots.
- Supplementary Fig. S15 Shoot regeneration of Arabidopsis clv3 mutant did not show obvious difference from the wild-type control.
- Supplementary Fig. S16 Shoot regeneration of wild-type and CRISPR-PagCLV3-1-2 explants.
- Supplementary Fig. S17 Shoot meristem activity was controlled by day-length.
- Supplementary Fig. S18 PagWUS-PagCLV3 module is involved in regulating shoot meristem cessation.
- Supplementary Fig. S19 The PagWUS-PagCLV3 module mediated proper secondary growth of the stem.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Dong WC, Wang FL, Li XT, Wu XX, Yu LF, et al. 2026. The PagWUS-PagCLV3 module regulates shoot meristem maintenance and activity in poplar. Forestry Research 6: e007 doi: 10.48130/forres-0026-0007 

The PagWUS-PagCLV3 module regulates shoot meristem maintenance and activity in poplar
- Received: 01 August 2025
- Revised: 04 December 2025
- Accepted: 22 January 2026
- Published online: 26 March 2026
Abstract: The shoot apical meristem of vascular plants generates all the aboveground organs. During this process, the structure and function of the meristem are maintained by a group of regulatory genes, among which the WUSCHEL (WUS)-CLAVATA3 (CLV3) module plays the core role. To date, all of the insights into shoot meristem homeostasis have been derived from studies on herbaceous plants. The mechanism by which the shoot meristem is maintained in trees remains unknown. In this study, we analyzed the functions of the poplar genes PagWUS and PagCLV3, homologs of Arabidopsis WUS and CLV3, respectively, in the maintenance and regeneration of the shoot meristem. Our results reveal both conserved and divergent functions compared to those of their orthologs in herbaceous species. Similar to their herbaceous counterparts, PagWUS and PagCLV3 are specifically expressed in the organizing center and stem cells, respectively, and form a feedback loop that regulates shoot meristem maintenance. Overexpression of PagWUS promoted shoot regeneration. Compared with herbaceous species, poplar possesses a much larger stem cell niche. The function of the PagWUS-PagCLV3 module is consistent with the developmental characteristics of perennial trees in that it regulates the cessation of the shoot meristem and mediates the proper pattern of secondary growth. Disruption of PagCLV3 enhanced shoot regeneration capacity. Our results shed light on shoot meristem regulation in trees and pave the way for understanding the mechanisms of meristem activity and plant development.
-
Key words:
- CLV3 /
- Poplar /
- Shoot meristem /
- Shoot regeneration /
- WUS