









-
Known as the 'Queen of forages', alfalfa (Medicago sativa L.) is a vital perennial legume that provides affordable, nutritious, and easily digested fodder, with alfalfa hay containing over 20% of crude protein[1,2]. However, alfalfa yields are significantly affected by environmental stresses such as drought, alkalinity, and salinity. Enhancing the stress tolerance of alfalfa and mining its adversity-related genes are crucial steps for increasing yield. Recent advancements in sequencing technologies and declining sequencing costs have led to continuous refinement of the alfalfa genome, laying a solid foundation for genome-wide gene family research[3−5].
The vast range of secondary metabolites produced by plants is impressive, yet many terpenes, phenols, fatty acid derivatives, and nitrogenous compounds are synthesized through a limited number of main pathways[6]. These metabolites gain their structural diversity through enzymatic modifications such as catalytic decarboxylation, oxidation/reduction, hydroxylation, glycosylation, methylation, and acylation[7]. Among these, acylation processes play a pivotal role in the formation of esters and amides, which are common modifications in secondary metabolites, by modifying oxygen- and nitrogen-containing substrates, respectively. Enzymes catalyzing these acyl activation are typically found in large protein families[8]. Acylation involves the introduction of alkyl (RCO-) or aromatic (ArCO-) acyl groups onto nitrogen, oxygen, carbon, or sulfur atoms in organic molecules[9]. Acyltransferases catalyze these reactions in organisms, influencing cellular functions and metabolism[10]. For example, SCPL genes are involved in the synthesis of galloylated catechins in tea plants[11] and chicoric acid in purple coneflower[12]. These enzymes are widespread in plants, animals, and microorganisms, transferring acyl groups from donors to acceptors[13−17]. SCPL enzymes, part of the S10 subfamily, share structural and functional similarities with serine carboxypeptidases and possess conserved regions critical for catalysis[15−18]. SCPLs are distinct in their significant role in plant defense mechanisms[18,19]. The type I SCPL gene in tomatoes is a 'ate trauma-induced gene', meaning it is strongly induced by mechanical injury and exposure to methyl jasmonate (MeJA)[20]. Defense-related genes such as OsPR1, OsPR2, OsPR5, and OsPDF1.2 were consistently up-regulated in transgenic rice plants, with those overexpressing OsBISCPL1 displaying notably higher resistance to Pseudomonas syringae compared to wild-type rice. Moreover, plants overexpressing OsBISCPL1 exhibited enhanced oxidative stress tolerance along with a significant increase in the expression of genes associated with oxidative stress[21].
As an important forage crop, alfalfa is often cultivated in marginal environments, which are typically characterized by multiple abiotic stresses such as drought, salinization, and low temperatures. These stress conditions severely affect the growth and yield of alfalfa. In recent years, studies have shown that SCPL proteins play a key role in plant stress responses. For example, SCPL genes in tea plants are involved in the synthesis of galloylated catechins, enhancing the plants' tolerance to environmental stresses[11]. In purple coneflower (Echinacea purpurea), SCPL enzymes catalyze the production of chicoric acid, which helps plants resist pathogens and adverse conditions[12]. Arabidopsis plants overexpressing TaSCPL184-6D reportedly showed elevated proline levels and reduced malondialdehyde content, which contributed to improved tolerance to salt and drought stresses[22]. Given the unique stress challenges faced by alfalfa in marginal environments, it is hypothesized that the SCPL gene family may play an important role in alfalfa's stress adaptation. Therefore, this study aims to comprehensively analyze the SCPL gene family in alfalfa, including the number of genes, their structure, chromosomal distribution, and phylogenetic relationships. Moreover, the detailed analysis of the SCPL gene family will help elucidate its functions in alfalfa and other closely related species. These efforts will enable researchers to gain a deeper understanding of the evolution, diversity, and physiological roles of the SCPL gene family.
-
The reference genome sequence and protein datasets for alfalfa (Zhongmu No. 1) were sourced from Figshare (
https://doi.org/10.6084/m9.figshare.12623960.v1 )[3], while SCPL protein sequence data for 54 Arabidopsis thaliana entries was retrieved from the TAIR database (www.arabidopsis.org ). The genes in the alfalfa genome were subsequently co-identified using BLAST with an E-value threshold of > 1e-10. The HMM profile (PF00450) of the SCPL structural domain was obtained from the InterPro database (www.ebi.ac.uk/interpro ) and analyzed using the Hidden Markov Model (HMM) method. Sequences were then entered into the NCBI Conserved Structural Domain Database (www.ncbi.nlm.nih.gov/cdd ) to confirm the presence of functional domains.Gene information and phylogenetic analysis
-
TBtools was used to examine and depict the chromosomal distribution of the identified SCPL family genes. The nomenclature of MsSCPLs was determined according to their chromosomal positions. The Expasy website (
https://web.expasy.org/compute_pi/ ) was utilized to evaluate the physicochemical properties of the identified genes. Subcellular localization predictions were performed using the WolfPSort online tool (https://wolfpsort.hgc.jp/ ). Phylogenetic trees were constructed using MEGA software[23]. Multiple amino acid sequence alignments were conducted using ClusterX2.0 software with default settings. The neighbour-joining (NJ) method was applied to create a phylogenetic tree, and 1,000 bootstrap replicates were performed for validation.Gene structure, motif identification, and conserved domains
-
The MEME Sequence Analysis Tool (
https://meme-suite.org/meme ) was employed to predict conserved motifs in the SCPLs, with a limit of 24 motifs specified. Additionally, the gene structures of the genes were analyzed using the advanced gene structure view function in TBtools, and the results were visualized through TBtools.Gene duplication and synteny analysis
-
MCScanX was run to locate duplication points within genes and to identify regions of collinearity with A. thaliana and M. truncatula[24]. The results were subsequently visualized using TBtools.
Promoter analysis
-
A 2,000 bp sequence upstream of the start codon for each identified gene was extracted and analyzed using the PlantCARE (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html ) database to predict cis-acting elements[25].Transcriptomic data analysis
-
The NCBI database (PRJNA276155, SRP055547) provided transcriptomic data for six alfalfa tissues: leaves, nodules, elongated stems, flowers, pre-elongated stems and roots[26]. Additionally, transcriptomic information for genes under drought, salt, and cold stress conditions was retrieved from the NCBI database (PRJNA454564, SRR7091780−SRR7091794 and PRJNA450305, SRR7160313−SRR7160357)[27]. The alfalfa reference genome (cv. Zhongmu No.1) was aligned using HISAT2[28], resulting in the generation of SAM files. Subsequently, the count values of the transcriptome data that corresponded to the genomic data were determined via the featureCounts software (v2.0.1)[29]. The gene expression levels were then estimated based on the FPKM values derived from the R package DESeq2 (v1.40)[30].
RT-qPCR analysis of genes
-
Total RNA was isolated using TRIzol Reagent (Vazyme, Nanjing, China) and single-stranded cDNA was synthesised using a First Strand cDNA Synthesis Kit (Vazyme, Nanjing, China). RT-qPCR was conducted using a SYBR Premix Ex Taq (Takara, Japan) and a 7,500 real-time fluorescence quantitative PCR system. Primers for three MsSCPL genes were designed using NCBI's Primer-BLAST (Supplementary Table S1), and Actin from alfalfa was used as an internal reference gene. Gene expression levels were calculated using the 2−ΔΔCᴛ method[31]. Each experiment was conducted with three technical replicates. Analysis of variance (ANOVA) was performed, followed by Duncan's multiple range test for post hoc multiple comparisons, and different lowercase letters indicate significance at p < 0.05.
Plant growth and treatments
-
Zhongmu No.1 alfalfa seeds were cold-primed at 4 °C for three days and then hydroponically grown in a greenhouse under a 16/8 h light/dark photoperiod at 24/20 °C day/night temperatures and 70%–80% relative humidity for two weeks. During cold treatment, leaves were maintained at 4 °C and sampled at 0, 2, 6, 12, and 24 h. To simulate salt stress, NaCl was applied at a concentration of 250 mM, and root tips were collected at six time points: 0, 1, 3, 6, 12, and 24 h post-treatment. For drought stress, mannitol (400 mM) was used to treat the root tips, with samples collected at six intervals: 0, 1, 3, 6, 12, and 24 h after treatment.
-
The 39 MsSCPL genes identified in the alfalfa genome are catalogued with their gene IDs, genomic positions, CDS lengths, protein sizes, molecular weights, isoelectric points, and predicted subcellular localizations (Supplementary Table S2). Among these 39 MsSCPL members, MsSCPL26 has the shortest CDS sequence at 636 bp, while MsSCPL19 has the longest, measuring 3,432 bp. The molecular weights of the proteins for these MsSCPL members vary from 15.24 kDa (MsSCPL27) to 128.54 kDa (MsSCPL19), and the pI values range from 4.74 (MsSCPL11) to 9.18 (MsSCPL3). According to the subcellular localization results, six members are found in the extracellular space, 13 in the chloroplast, seven in the vacuole, six in the cytoplasm, five in the nucleus, and two in the mitochondria.
The 39 genes are distributed across eight chromosomes, primarily concentrated on chromosomes 1, 2, 3, and 4 (Fig. 1). Chromosome 4 had the most MsSCPL genes, totaling 10. Interestingly, only one gene, MsSCPL39, was discovered on chromosome 8. A phylogenetic tree was built using sequences from 39 alfalfa proteins and 54 Arabidopsis thaliana SCPL gene family proteins to improve understanding of the taxonomic and genetic relationships between members of the alfalfa gene family (Fig. 2). The SCPL gene family can be categorized into three subgroups: Group I, II, and III. Group I contains 12 members, Group III has 11 members, and Group IIII, has the largest number, with 16 members. It is possible to predict the functions of proteins that occur in the same group because they generally have similar characteristics.
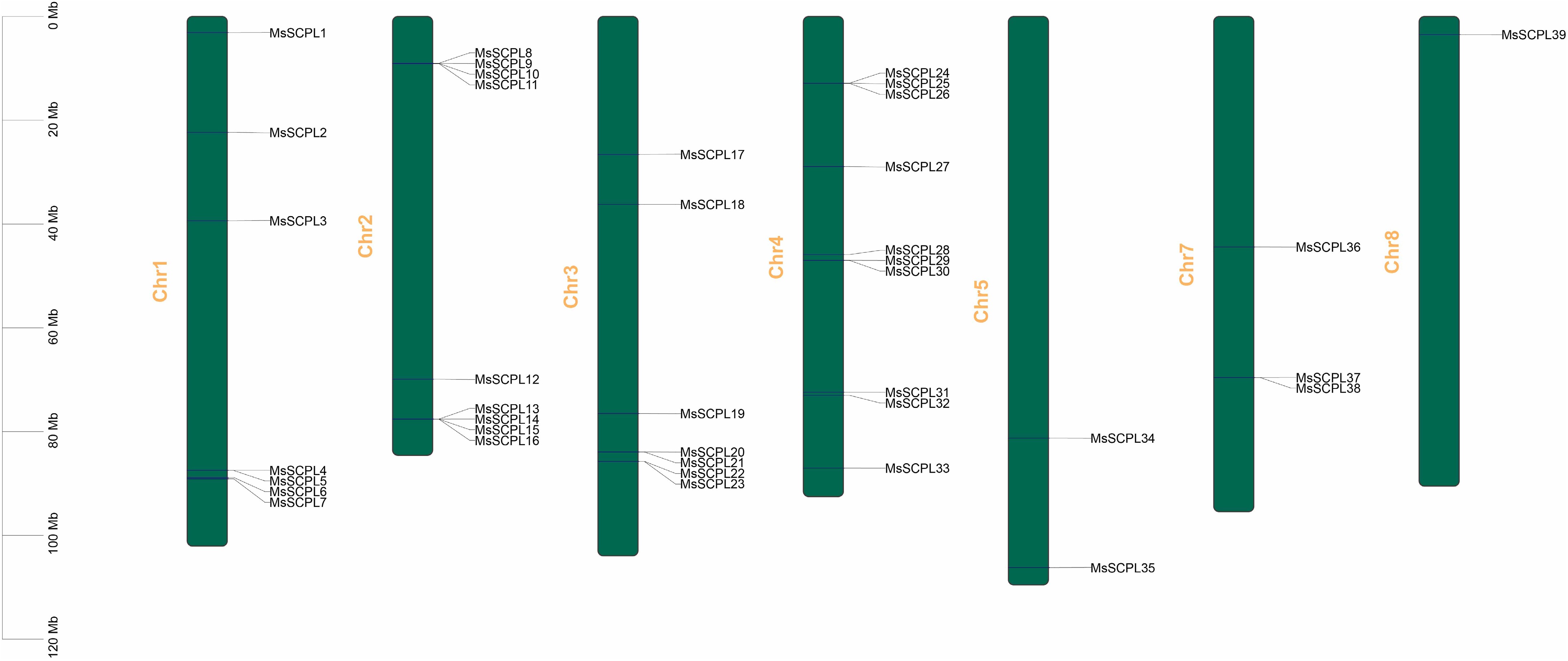
Figure 1.
Chromosomal positions of members of the SCPL gene family in alfalfa.
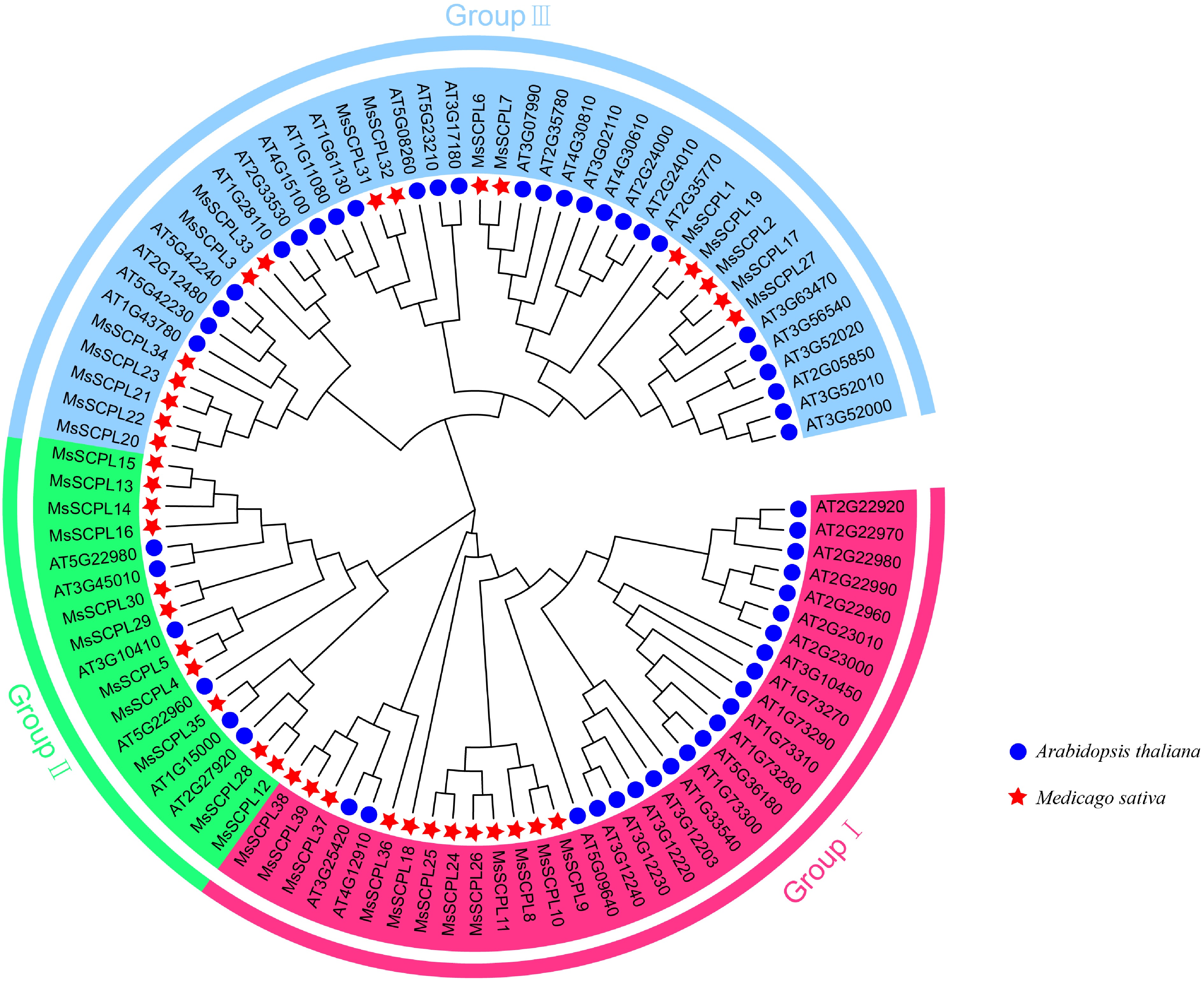
Figure 2.
Phylogenetic tree of the SCPL gene family in A. thaliana and M. sativa.
MsSCPL gene structure and conserved motif analysis
-
The conserved motifs of SCPL proteins were analyzed using the MEME online software to enhance understanding of the range and overlap of motifs within the MsSCPL gene family. Among the 39 MsSCPL members, 10 conserved motifs (motif1–motif10) were identified (Fig. 3). Notably, motifs 3 and 4 were found exclusively in MsSCPL26 of Group I, while the majority of members contained 7 to 8 motifs. The motifs of SCPL proteins within the same subgroup are identical or similar. For instance, motif 5 is widespread in the SCPL proteins of Group II, whereas motifs 1 and 4 are predominantly present in the majority of Group II members. Although only one protein in Group III lacks motif 8, all Group III proteins possess the conserved motif 10.
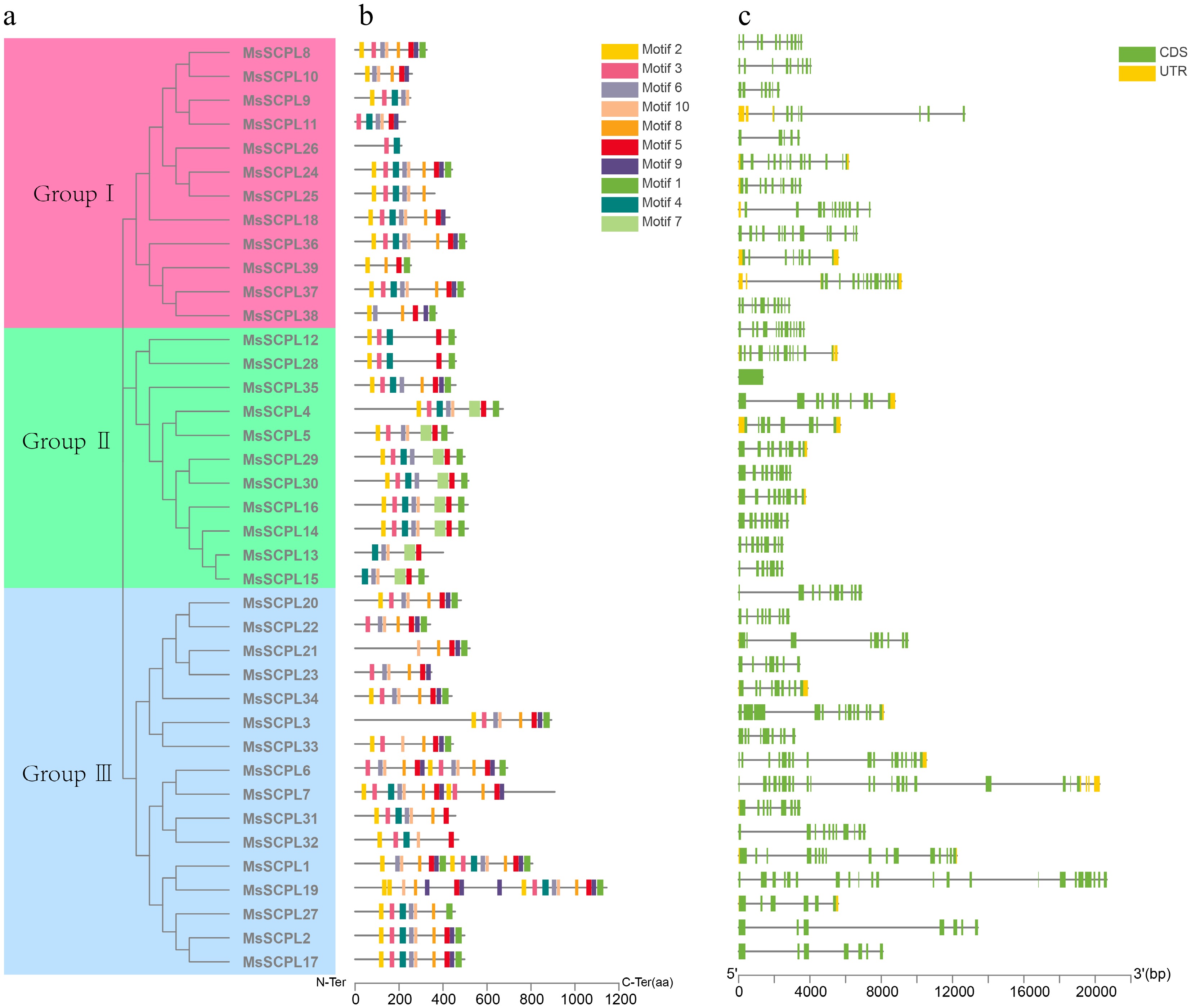
Figure 3.
Structural characterization of SCPL in alfalfa. (a) Phylogenetic analysis of proteins. (b) Predicted motifs of proteins. (c) Exon-intron structure of the gene.
Analysis of gene synteny and gene duplication events in MsSCPL genes
-
In alfalfa, tandem and segmental duplication events of MsSCPL were investigated to clarify the evolutionary processes that contributed to the expansion of the MsSCPL gene family (Fig. 4). Seven pairs of tandem duplication events were identified in the MsSCPL genes of alfalfa, and the data on these duplication events shed light on the diversity and complexity of the alfalfa genome, indicating a significant number of duplication events within the MsSCPL family. A covariance map was created to illustrate potential evolutionary events influencing the MsSCPL gene family across M. sativa, A. thaliana, and M. truncatula (Fig. 5). A total of 28 co-lineage pairs of SCPL genes were identified between alfalfa and M. truncatula, while seven co-lineage pairs were found between alfalfa and A. thaliana. Notably, the number of co-lineage pairs of SCPL genes between M. truncatula and alfalfa was four times greater than that between A. thaliana and alfalfa.
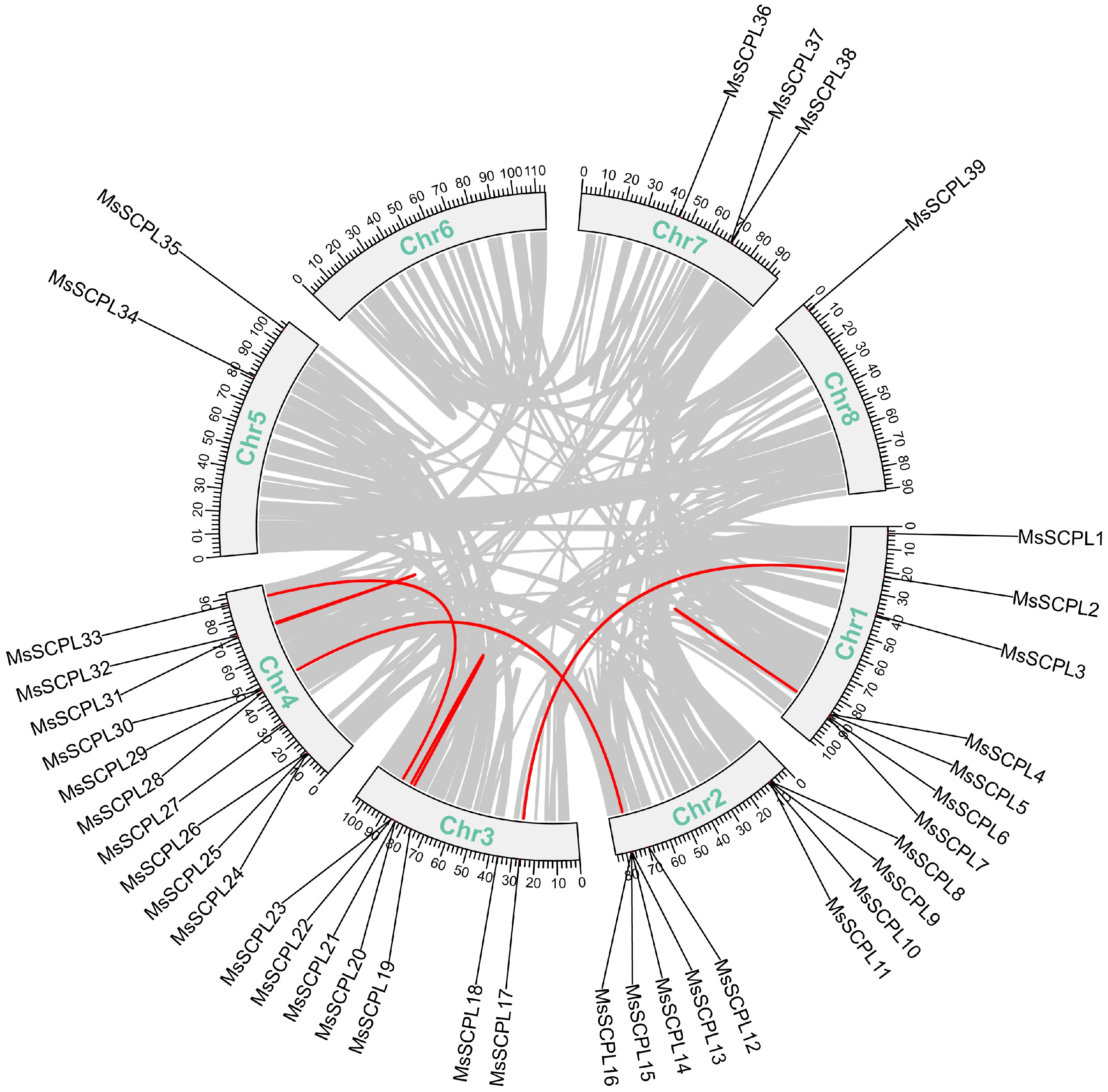
Figure 4.
Schematic representation of collinear connections of the gene in alfalfa. Segmental duplication events in the alfalfa genome are indicated in red, while collinear blocks are represented by grey lines.
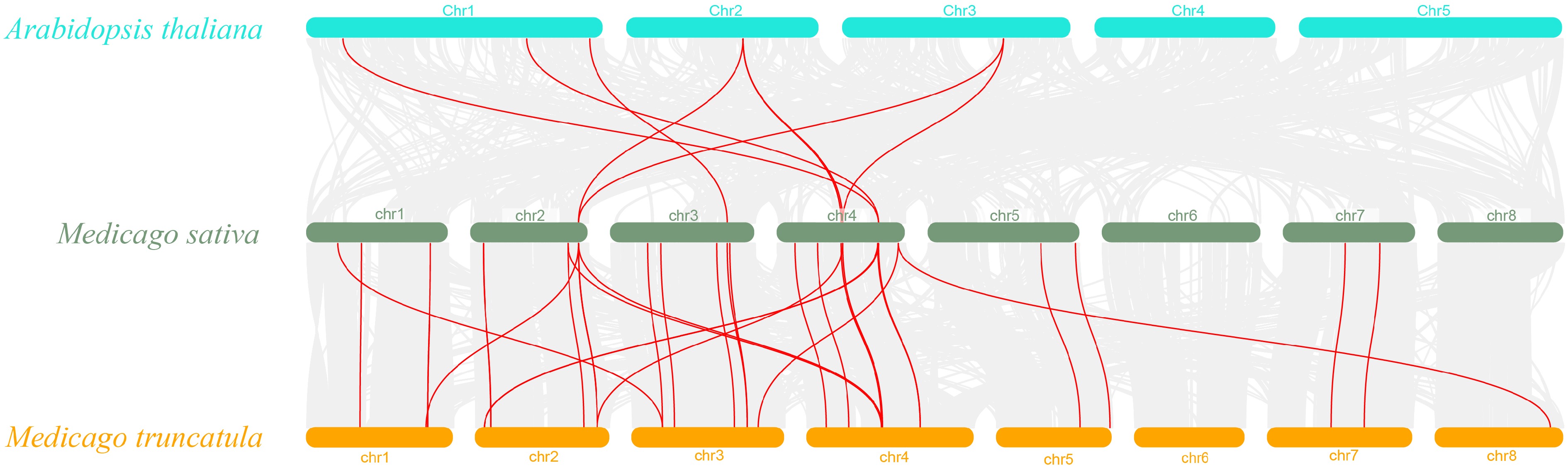
Figure 5.
Synteny analysis of the SCPL gene between M. sativa and two representative plant species. Red lines highlight co-linear SCPL gene pairs, while grey lines in the background illustrate co-linear blocks between M. sativa and the selected plant species.
Examination of cis-elements in MsSCPL gene promoter regions
-
To better understand the biological roles of MsSCPL genes in alfalfa, an analysis was conducted of cis-elements in the promoter sequences of the genes, located around 2 kb upstream of the start codon (ATG). Thirteen distinct cis-elements related to hormone-responsive, plant development regulatory, and stress were identified in promoter regions (Fig. 6). Among these, hormone-responsive elements are the most prevalent, comprising approximately 57%, while stress-related elements account for 30%, and development-related elements make up the remaining 13%. Within the hormone-related elements, those associated with jasmonic acid and abscisic acid responses (such as CGTCA-motif and ABRE) are more numerous, and some genes also contain auxin and gibberellin response elements (e.g., TGA-box, P-box, etc.). Stress-related elements primarily include anaerobic ARE elements, drought-related MBS elements, and low temperature-related LTR elements. It is noteworthy that most of the MsSCPL genes have ARE, ABRE, CGTCA, and TGACG elements, with ARE being present in all members of Group I.
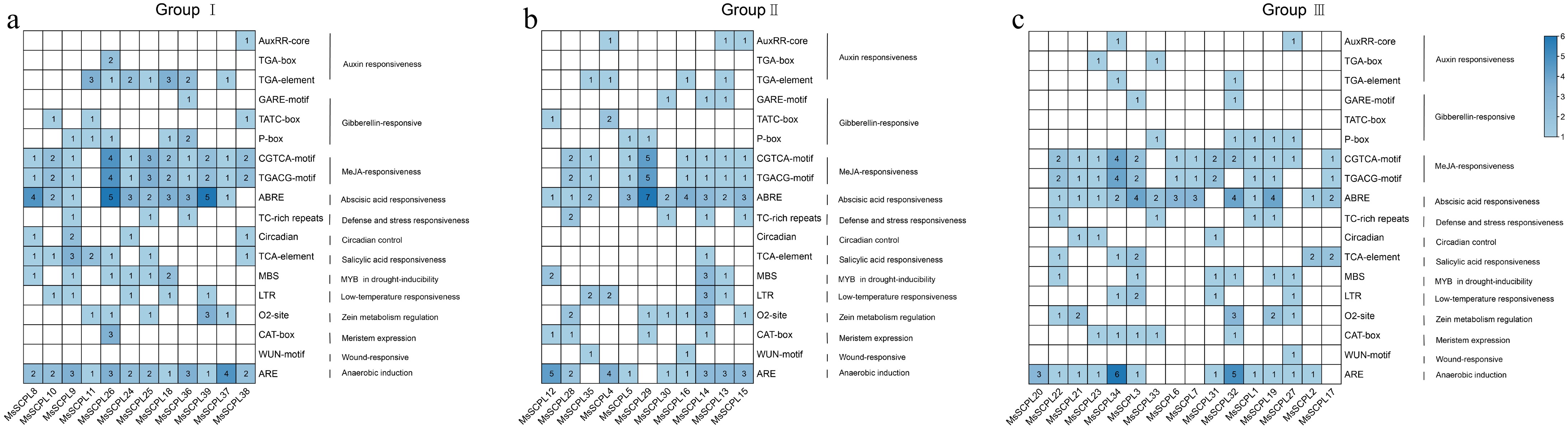
Figure 6.
Analysis of cis-acting elements in alfalfa SCPLs. (a) Panel represents the cis-acting elements of gene family members from Group I. (b) Panel represents the cis-acting elements of gene family members from Group II. (c) Panel represents the cis-acting elements of gene family members from Group III.
MsSCPL gene expression trends in alfalfa tissues
-
Transcriptome data for six distinct alfalfa tissues were sourced from a freely available database to analyze the expression patterns of the MsSCPL genes in these various tissues. Thirty-three MsSCPL genes were expressed in six tissues, and the remaining six genes were partially expressed in six tissues (Supplementary Table S3). Specifically, only MsSCPL36, MsSCPL20, and MsSCPL22 are expressed in flowers and nodules, while MsSCPL12 is not expressed in post-elongated stems and roots. Additionally, MsSCPL1 and MsSCPL30 are each absent in only one tissue: MsSCPL1 is not expressed in roots, and MsSCPL30 is not expressed in post-elongated stems. In addition, the expression abundance of these MsSCPL genes differed significantly between tissues. For instance, MsSCPL20/22/36 was found to be co-expressed in flowers and roots, but its expression level was significantly higher in flowers than in roots. Although 33 genes were expressed in all six tissues, MsSCPL6/7/9/18/24 were predominantly expressed in leaves while MsSCPL2/17/19/25/26/29/35/37/38 were highly expressed in roots.
Analysis of MsSCPL gene expression in abiotic stresses
-
A public database was utilized to collect and analyze transcriptome data of alfalfa seedlings subjected to three stresses, aiming to elucidate the diverse expression levels of the MsSCPL genes under these three abiotic stresses. Expression profiling showed that 11, 16, and 14 MsSCPL genes were responsive to drought, salt and cold stresses, respectively (Supplementary Tables S4–S6; Fig. 7). Notably, MsSCPL6/7/29 responded to both salt and cold stress, while nine genes—MsSCPL17/19/27/28/32/33/34/37—were responsive to both salt and drought stress. Remarkably, MsSCPL31/35 showed responses to all three stresses; therefore, these two genes were selected for RT-qPCR validation analysis.
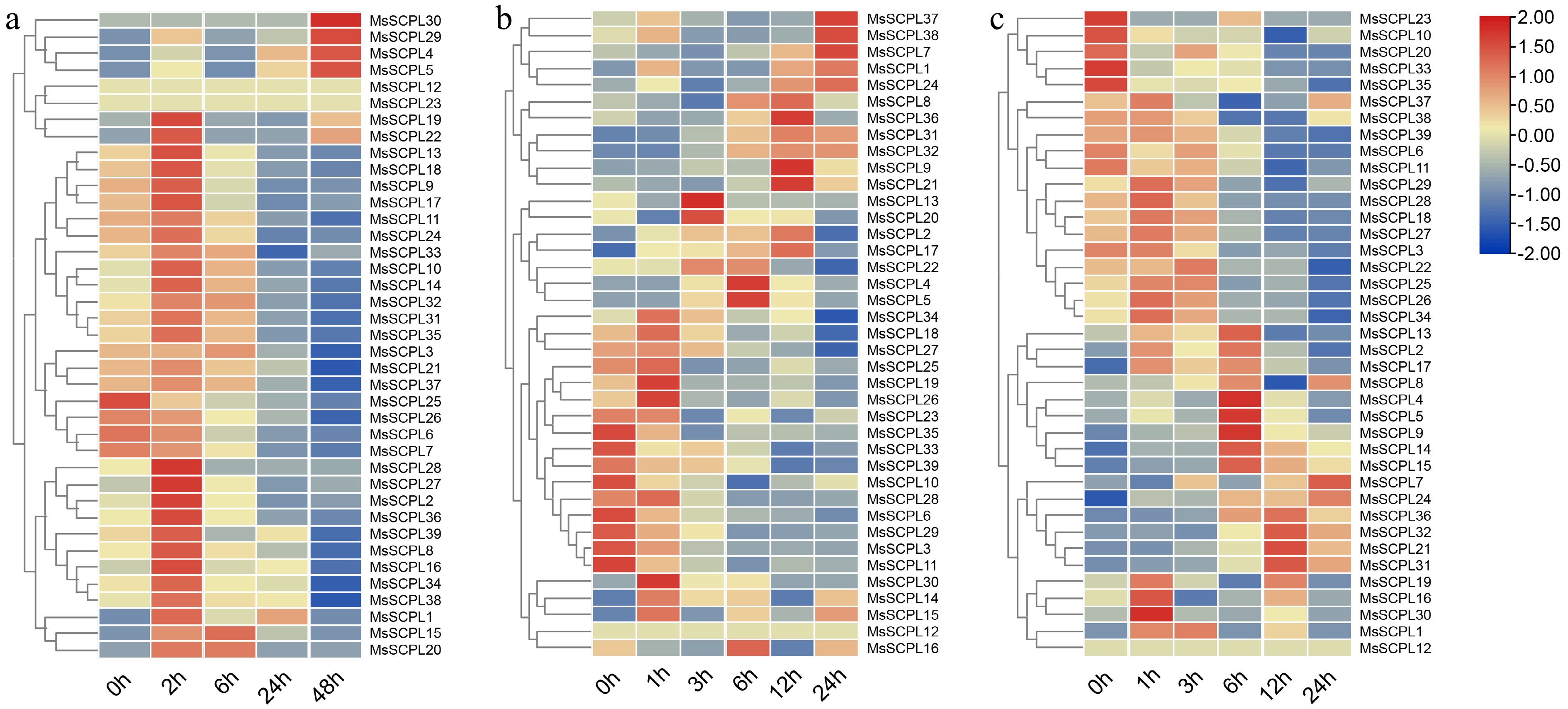
Figure 7.
Expression of genes in response to cold, salt, and drought conditions. (a) Expression of genes under cold stress. (b) Expression of genes under salt stress. (c) Expression of genes under drought stress.
To verify the transcriptome data findings, RT-qPCR tests were conducted on two selected genes (MsSCPL31/35). Under the three stress conditions, the expression patterns of MsSCPL31/35 varied over time (Fig. 8). Interestingly, under drought stress, the expression of MsSCPL31 showed an upward and then downward trend, and the expression was considerably increased compared to the control at 12 h. The expression of MsSCPL31 increased to a peak at 3 h of salt stress treatment, followed by a downward trend. Under cold stress, the expression of MsSCPL31 was up-regulated at 2 h of the cold stress treatment, and then showed a downward trend. Under drought stress, the expression of MsSCPL35 was first up-regulated at 0–1 h, then down-regulated at 3 h, then up-regulated to the maximum at 6 h, and finally showed a continuous down-regulation trend. After just 1 h of salt stress treatment, the level of expression of MsSCPL35 was considerably above that observed in the control, and then declined as a whole with the prolongation of the time of salt stress; and the expression of MsSCPL35 first rose and then fell in response to cold stress. These genes should be further investigated for their roles under abiotic stress.
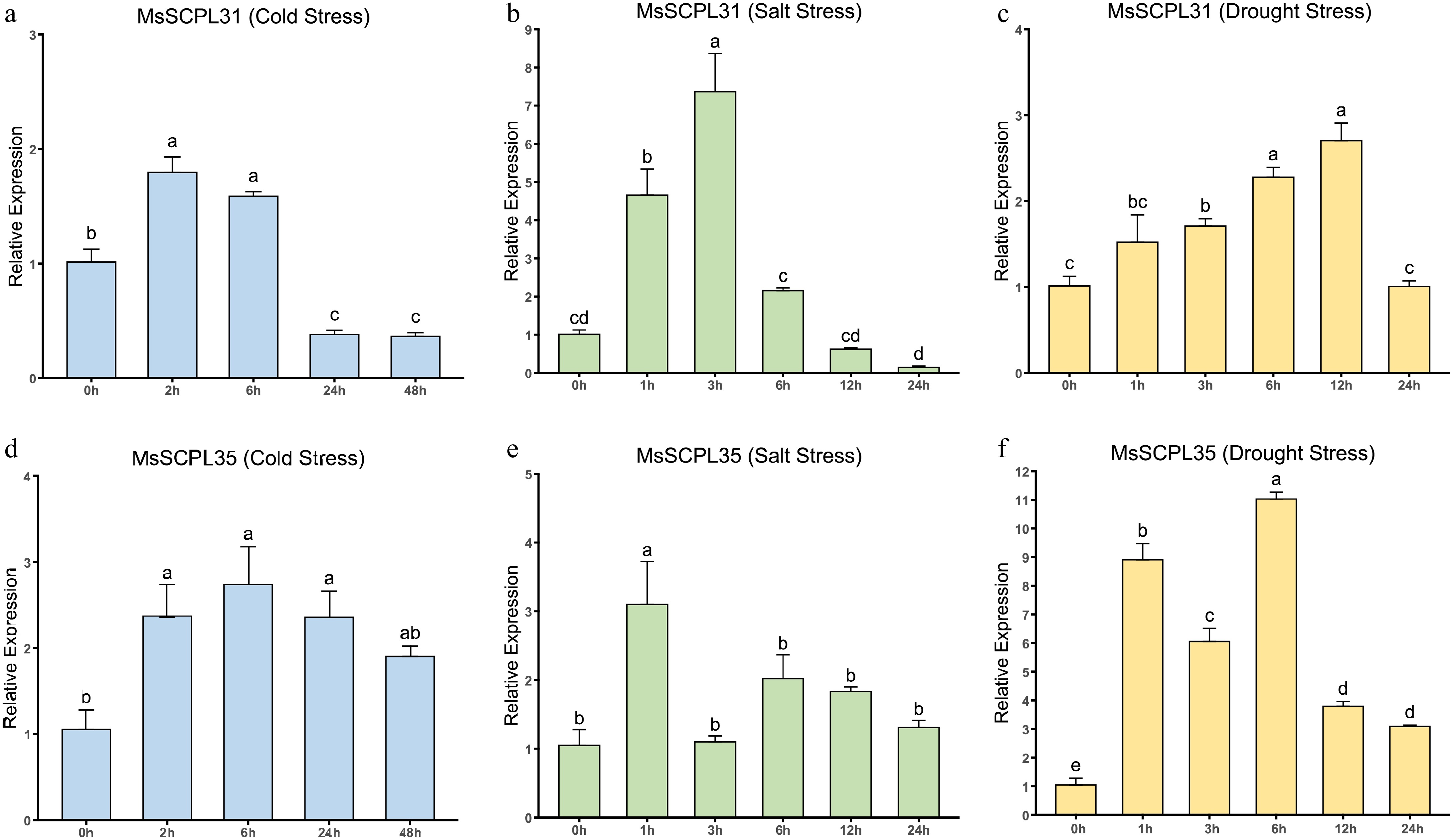
Figure 8.
Expression of genes under drought, salt and cold stress conditions as determined by RT-qPCR. (a) Expression levels of MsSCPL31 under cold stress. (b) Expression levels of 31 under salt stress. (c) Expression levels of MsSCPL31 under drought stress. (d) Expression levels of MsSCPL35 under cold stress. (e) Expression levels of MsSCPL35 under salt stress. (f) Expression levels of MsSCPL35 under drought stress. Different lowercase letters indicate significance at p < 0.05.
-
The SCPL gene, originating from an ancestral serine carboxypeptidase family, is significant in plant growth, maturation, and stress adaptation[21,32,33]. Genome-wide analyses of the SCPL gene family have been conducted in several plants, identifying 54, 210, 71, 41, and 96 SCPL genes in Arabidopsis[34], wheat[22], rice[35], soybean[36], and cotton[37], respectively. Comparing the number of SCPL genes across species shows that the MsSCPL count in alfalfa is relatively low. For instance, rice has 71 SCPL genes, soybean has 73, and wheat contains as many as 210. This high number in wheat is attributed to its complex heterozygous hexaploid nature, which underwent two whole-genome duplication events. In contrast, the alfalfa genome analyzed in this study is based on the haploid genome of 'Zhongmu No.1', leading to the identification of only 39 SCPL genes. Additionally, variations in threshold settings and screening strategies may also contribute to these differences in gene identification.
In this study, RNA-seq data from various alfalfa tissues (leaves, elongated stems, roots, pre-elongated stems, nodules, and flowers) revealed that MsSCPL genes exhibit high expression levels in roots and flowers, suggesting their potential roles in plant development and formation processes. The SCPL acyltransferase is involved in various metabolic pathways, which are likely strongly linked to the plant's response to abiotic stress[36]. Moreover, the analysis showed that six genes are located extracellularly, while seven genes are situated in vesicles, indicating that MsSCPL genes may participate in diverse secondary metabolic pathways associated with abiotic stress responses. Promoter analysis of MsSCPLs revealed elements associated with developmental processes, resistance to environmental stress and reaction to hormones (Fig. 6). These results suggest that MsSCPLs may play critical roles in alfalfa's responses to different types of stress. Previous studies have shown that the SCPL gene family is relevant to how plants respond to salt and drought stress. For example, following three hours of ABA treatment and 68 h of salt and drought treatments, TaSCPL1-24D showed the highest expression levels in wheat[22]. It was reduced under cold, salt stress, and drought stress. This suggests that the CsSCPL gene may play a regulatory role in the response of tea tree to these stresses[11]. In potatoes, StSCPL genes showed significant expression under salt stress, heat stress, drought stress, and hormone treatments[38]. The RT-qPCR results of this study also demonstrated that the expression patterns of MsSCPL31 and MsSCPL35 under different abiotic stresses showed significant time dependence and stress specificity. MsSCPL31 is significantly up-regulated at 12 h under drought stress, suggesting its involvement in early stress responses; it reaches a peak at 3 h under salt stress, indicating a potential role in early defense mechanisms; and it is up-regulated at 2 h under cold stress, likely participating in rapid signal transduction. The peak expression of MsSCPL35 at 6 h under drought stress may reflect its involvement in the early stress response rather than sustained stress acclimation; it is significantly up-regulated at 1 h under salt stress, followed by a decline, suggesting its role in early stress responses; and it displays an initial increase followed by a decrease under cold stress, likely contributing to early defense mechanisms. These distinct expression profiles are presumably closely related to their subcellular localization, protein interactions, and mechanisms of stress signal transduction. Future research could employ approaches such as gene knockout, overexpression, and protein interaction analysis to further elucidate the specific molecular mechanisms underlying these genes.
Alfalfa, as a high-quality forage crop, is rich in protein and has always held a significant position in the forage industry. However, most alfalfa cultivation areas are located on marginal lands, often facing stresses such as drought and salinity, making large-scale cultivation in these challenging environments difficult. In recent years, the rapid advancement of transgenic and gene editing technologies has facilitated significant progress in the development of stress-resistant alfalfa varieties. This research study identified 39 SCPL gene family members in alfalfa, and some were found to respond to drought, salt, and cold stresses. These responses can be functionally validated using molecular techniques, paving the way for the development of new resilient alfalfa varieties. Additionally, this enhances understanding of the biological functions of genes in alfalfa and other plants, as well as the molecular mechanisms underlying abiotic stress regulation.
-
This study comprehensively identified and analyzed the members of the alfalfa SCPL gene family. Phylogenetic analysis categorized the family members into Groups I, II, and III based on the distribution of the 39 gene loci across four chromosomes. Phylogenetic analysis showed that the MsSCPL gene family exhibited significant homology to the gene family found in Arabidopsis, and the expansion of the gene family in alfalfa was primarily driven by segmental duplications rather than tandem duplications. According to the gene expression data from different types of tissue, 33 MsSCPL genes were expressed in six tissues, while the remaining 6 genes were expressed in some tissues. MsSCPL genes are evidently crucial for plant growth and development. Moreover, the analysis of cis-acting elements has revealed that SCPL genes encompass several elements associated with abiotic stresses. The transcriptome data under different stresses showed that 11, 16, and 14 genes responded to cold, salt, and drought stresses, respectively, while MsSCPL31/35 surprisingly responded to all three stresses simultaneously, and the transcriptome data were further validated by RT-qPCR. This study provides useful information about MsSCPL genes, but lacks further functional validation. In the future, techniques such as overexpression, gene editing, and molecular interactions may be used to validate and analyze the functions of the MsSCPL gene family. In summary, this study deepens the understanding of MsSCPL genes in response to abiotic stresses and provides candidate genes for stress-resistant alfalfa breeding.
This work was supported by the Central Public-Interest Scientific Institution Basal Research Fund (Grant No. Y2025YC44), the China Agriculture Research System of MOF and MARA (Grant No. CARS-34), and the Agricultural Science and Technology Innovation Program of CAAS (Grant No. ASTIPIAS14).
-
The authors confirm their contributions to the paper as follows: methodology: Zhu H, Liu H, Wu Y; investigation: Zhu H, Liu Q; software: Zhu H, Liu H, Xu Y; validation: Liu Q, Xu Y, Qu X, Chen Y; data curation & resources: Wu Y; original draft: Zhu H; funding acquisition, supervision & project administration: Yang Q, Long R; writing - review & editing: Liu Q, Qu X, Chen Y, Yang Q, Long R. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers of MsSCPL genes used for RT-qPCR.
- Supplementary Table S2 Basic information on the MsSCPL family genes in alfalfa.
- Supplementary Table S3 The expression of MsSCPLs in different tissues.
- Supplementary Table S4 Candidate MsSCPL genes under drought stress.
- Supplementary Table S5 Candidate MsSCPL genes under salt stress.
- Supplementary Table S6 Candidate MsSCPL genes under cold stress.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhu H, Liu H, Liu Q, Wu Y, Xu Y, et al. 2025. Identification and response to abiotic stress of the alfalfa serine carboxypeptidase-like protein gene family. Grass Research 5: e022 doi: 10.48130/grares-0025-0019 

Identification and response to abiotic stress of the alfalfa serine carboxypeptidase-like protein gene family
- Received: 18 May 2025
- Revised: 27 June 2025
- Accepted: 03 July 2025
- Published online: 02 September 2025
Abstract: A class of acyltransferase enzymes known as serine carboxypeptidase-like (SCPL) proteins play a pivotal role in plant growth, development, and responses to stress. While SCPL proteins have been extensively studied in various plants, their biological functions in alfalfa remain largely unexplored. In this study, 39 SCPL gene family members were identified in alfalfa and classified into three subgroups (Groups I–III). Their chromosomal structure and homology were also analyzed. Gene transcription levels were analyzed, revealing that six genes were expressed in a few specific tissues, while the remaining 33 MsSCPL were expressed throughout the tissues. Additionally, analysis of cis-acting elements revealed that the SCPL gene family contains components linked to abiotic stress. Meanwhile, transcriptome analysis revealed that 11, 16, and 14 members of MsSCPL gene family were found to respond to three stresses, respectively (cold, salt, and drought), in addition to MsSCPL31/35, which responded to all stresses at the same time. The transcriptome results were further validated by RT-qPCR. This highlights the role of MsSCPL31/35 in abiotic stress response, suggesting their importance for plant adaptation mechanism. These results could be crucial for understanding the mechanisms behind alfalfa SCPL gene function.
-
Key words:
- Medicago sativa /
- Serine carboxypeptidase /
- Gene family /
- Abiotic stress