









-
Oomycetes from the kingdom Straminipila[1] have adapted to a range of environmental conditions by evolving as either saprotrophs or pathogens. Members of this group are widely distributed in terrestrial and aquatic environments[2−5]. Oomycetes, often referred to as pseudo-fungi or water molds[1,5], exhibit several traits with similar characteristics as true fungi, such as mycelial growth, strategies of infection, and an osmotrophic mode of nutrition. Most studies on oomycetes are biased towards terrestrial plant pathogens[2,6−11]; however, there have been several studies reporting the occurrence and ecological roles of oomycetes in other biomes, such as freshwater[12−21], marine, and brackish environments[22−35]. This highlights the worth of exploration into this diverse group.
The freshwater biome serves as one of the most important habitats for oomycetes. Common aquatic species are congeners within the Peronosporales, Pythiales, and Saprolegniales (Table 1). They were isolated from diverse substrates, including algae, animals, plants, soil, and water of mountain streams, reservoirs, and rivers. Although some members of these groups are pathogens of fish[36−38], amphibians[39−41], and algae[42,43], the potential ecological roles of oomycetes in the freshwater biomes are not to be neglected (e.g., litter decomposition and recycling of organic materials)[44,45]. Saprotrophic microorganisms play an important role in biogeochemical cycles and food webs, and this functioning maintains the ecological processes[44−46].
Table 1. Oomycetes inhabiting freshwater ecosystems.
Taxon Substrate Distribution Ref. Order Species Algae Amphibian Crustaceans Fish Mosquito Nematoda Plant Soil Water Stream Reservoir River Myzocytiopsidales Resticularia marchaliana o o Czeczuga et al.[47] Leptomitales Apodachlya brachynema o o Czeczuga et al.[47] Peronosporales Halophytophthora fluviatilis o o o o Yang & Hong[48] Phytophthora ×alni o o o o o Jung & Blaschke[49], Bjelke et al.[50] Phytophthora taxon aquatilis o o Hong et al.[51] Phytophthora bilorbang o o Corcobado et al.[20], Christova[52] Phytophthora cactorum o o o o Jung et al.[53], Jung et al.[19], Reeser et al.[12] Phytophthora ×cambivora o o o o o o Hwang et al.[54], Remigi et al.[55], Reeser et al.[12], Jung et al.[17], Mullett et al.[56] Phytophthora capensis o o o o o Bezuidenhout et al.[57], Jung et al.[16], Bose et al.[58] Phytophthora chlamydospora* o o o o o o Remigi et al.[55], Reeser et al.[12], Jung et al.[59], Hong et al.[51], Jung et al.[10], Jung et al.[17], Hansen et al.[60], Jung et al.[16], Nam et al.[61], Corcobado et al.[20] Phytophthora cinnamomi o o o o o Hwang et al.[54], Hong et al.[51], Jung et al.[62], Stamler et al.[63], Jung et al.[16] Phytophthora citricola o o o o o Hwang et al.[54], Remigi et al.[55], Hulvey et al.[64] Phytophthora citrophthora o o o o o o Hwang et al.[54], Hulvey et al.[64], Hong et al.[51], Jung et al.[65], Jung et al.[66] Phytophthora cryptogea o o Hong et al.[51] Phytophthora drechsleri o o o o o Jung et al.[65], Jung et al.[16] Phytophthora europaea o o o o o Jung et al.[67], Remigi et al.[55], Hwang et al.[54], Reeser et al.[12] Phytophthora gallica o o Corcobado et al.[20] Phytophthora gibbosa o Jung et al.[59] Phytophthora gonapodyides o o o o o o Hwang et al.[54], Jung[68], Remigi et al.[55], Jung et al.[59], Belisario et al.[69], Corcobado et al.[20] Phytophthora gregata o o o o Jung et al.[59] Phytophthora heveae o o o o Erwin & Ribeiro[8], Hwang et al.[54], Jung et al.[19], Jung et al.[16] Phytophthora honggalleglyana o o Christova[52] Phytophthora hydropathica o o o o o Hulvey et al.[64], Hong et al.[70], Corcobado et al.[20] Phytophthora inundata* o o o o o o Brasier et al.[71], Jung et al.[59], Hüberli et al.[13], Christova[52] Phytophthora irrigata o o o o o Hong et al.[72], Hulvey et al.[64] Phytophthora lacustris o o Corcobado et al.[20], Christova[52] Phytophthora sp. 'lagoariana'* o o o Wallace[73], Nam et al.[61] Phytophthora litoralis o o o o o Hulvey et al.[64], Jung et al.[59] Phytophthora macrochlamydospora o o o o Jung et al.[16] Phytophthora megasperma o o o o o o Erwin & Ribeiro[8], Remigi et al.[55], Jung et al.[59], Reeser et al.[12], Jung et al.[65], Jung et al.[17] Phytophthora mississippiae o o o Hong et al.[51] Phytophthora nemorosa o o o o Reeser et al.[12] Phytophthora nicotianae o o o o Hulvey et al.[64] Phytophthora pini o o o o Reeser et al.[12], Jung et al.[65], Jung et al.[66], Milenković et al.[74] Phytophthora plurivora o o o o o Jung & Burgess[75], Reeser et al.[12], Jung et al.[10], Jung et al.[17], Jung et al.[18], Jung et al.[19], Jung et al.[65], Jung et al.[66] Phytophthora pseudosyringae o o o o o Hwang et al.[54], Jung[68], Reeser et al.[12], Jung et al.[10], Jung et al.[65] Phytophthora ramorum o o o o Werres et al.[76], Reeser et al.[12], Hong et al.[51], Eyre et al.[77], Jung et al.[16] Phytophthora riparia o o o Hong et al.[51], Corcobado et al.[20] Phytophthora siskiyouensis o o o o Reeser er al.[12], Reeser et al.[78] Phytophthora syringae o o o o o o Erwin & Ribeiro[8], Remigi et al.[55], Hulvey et al.[64], Reeser et al.[12], Hong et al.[51], Jung et al.[65], Corcobado et al.[20] Phytophthora thermophila o o o o o Jung et al.[17], Jung et al.[59] Phytophthora vesicula o o Nakagiri[26] Pythiales Elongisporangium anandrum o o Christova[52] Elongisporangium helicandrum o o Czeczuga et al.[47] Elongisporangium undulatum* o o o o o o Khulbe[79], Abdelzaher et al.[80], Czeczuga & Snarka[81] Globisporangium alternatum* o o Rahman et al.[82] Globisporangium attrantheridium* o o o o Nelson & Karp[83] Globisporangium carolinianum o o o Abdelzaher et al.[80], Czeczuga & Snarka[81] Globisporangium debaryanum o Poitras[84] Globisporangium echinulatum o o o o o o Robertson[85], Khulbe[79], Czeczuga[86], Czeczuga & Snarka[81] Globisporangium heterothallicum* o o Nam & Choi[87] Globisporangium intermedium* o o o o o o El-Hissy et al.[88], Czeczuga[86], Czeczuga & Snarka[81], Czeczuga & Muszynska[89], Nam & Choi[87] Globisporangium irregulare* o o o o Poitras[84], Abdelzaher et al.[80], Czeczuga & Snarka[81] Globisporangium lucens o o Czeczuga & Snarka[81] Globisporangium macrosporum* o o In the present study Globisporangium marsipium* o o o o Czeczuga[86], Czeczuga & Muszynska[89], Matsiakh et al.[90] Globisporangium megalacanthum o o o o Czeczuga & Snarka[81], Czeczuga & Muszynska[89] Globisporangium middletonii o o o o Karling[91], Czeczuga & Snarka[81] Globisporangium multisporum o o Poitras[84] Globisporangium nagaii* o o Vetukuri et al.[92] Globisporangium paddicum* o o In the present study Globisporangium pleroticum o o Abdelzaher et al.[80], Czeczuga & Muszynska[89] Globisporangium pulchrum o o o Poitras[84], Khulbe[79] Globisporangium rostratifingens* o o o o In the present study Globisporangium rostratum o o o o o Czeczuga[86], Czeczuga & Snarka[81], Czeczuga & Muszynska[89] Globisporangium selbyi* o o In the present study Globisporangium spinosum* o o o o o o Khulbe[79], Abdelzaher et al.[80], Czeczuga & Snarka[81] Globisporangium splendens o o Poitras[84] Globisporangium sylvaticum* o o o o o o o Abdelzaher et al.[80], Czeczuga[86], Czeczuga & Muszynska[89], Matsiakh et al.[90] Globisporangium terrestre * o o Matsiakh et al.[90] Globisporangium ultimum* o o o o o Poitras[84], El-Hissy et al.[88], Czeczuga[86], Czeczuga & Snarka[81], Czeczuga & Muszynska[89], Nam & Choi[87] Lagenidium giganteum o o Czeczuga et al.[47] Lagenidium microsporum o o Czeczuga et al.[47] Lagenidium oophilum o o Czeczuga et al.[47] Myzocytium megastomum o o Czeczuga et al.[47] Myzocytium rabenhorstii o o Czeczuga et al.[47] Myzocytium zoophthorum o o Czeczuga et al.[47] Phytopythium chamaehyphon* o o Nam & Choi[87] Phytopythium citrinum o o Christova[52] Phytopythium indigoferae o o Czeczuga et al.[47] Phytopythium iriomotense o o Baten et al.[93] Phytopythium litorale* o o o o o Nam & Choi[87], Vetukuri et al.[92], Christova[52] Phytopythium palingenes o o El-Hissy et al.[88], Czeczuga et al.[47] Phytopythium vexans* o o o o Poitras[84], Khulbe[79], Nam & Choi[87] Pythium acanthicum* o o o o Robertson[85], Czeczuga & Snarska[81] Pythium adhaerens o o o Sparrow[94], Abdelzaher & Kageyama[95] Pythium afertile o o o o Robertson[85], Czeczuga & Snarka[81] Pythium amaminum o o o Kikuchi et al.[96] Pythium angustatum o o o Matsiakh et al.[90], Christova[52] Pythium aphanidermatum* o o o o o o Poitras[84], Khulbe[79], Al-Sheikh & Abdelzaher[97], Lodhi et al.[98] Pythium apleroticum* o o In the present study Pythium aquatile* o o o o Czeczuga & Snarka[81], Czeczuga & Muszynska[89], Matsiakh et al.[90], Abdelzaher & Kageyama[95] Pythium aristosporum o o o o o Czeczuga & Snarka[81], Czeczuga[86], Czeczuga & Muszynska[89] Pythium arrhenomanes o o Czeczuga & Muszynska[89] Pythium biforme* o o o Uzuhashi et al.[99] Pythium brachiatum* o o Uzuhashi et al.[99] Pythium cactacearum o o Czeczuga et al.[47] Pythium catenulatum* o o o o Poitras[84], Abdelzaher et al.[80], Czeczuga & Snarka[81], Rahman & Sarowar[100] Pythium cedri* o o In the present study Pythium coloratum o o Robertson[85] Pythium debaryanum o o o o o o Robertson[85], Khulbe[79], Czeczuga et al.[47] Pythium deliense o o Abdelzaher et al.[80] Pythium diclinum* o o o o o Abdelzaher et al.[80], Al-Sheikh & Abdelzaher[97], Nam & Choi[87], Abdelzaher & Kageyama[95] Pythium dissimile o o Czeczuga & Snarka[81] Pythium dissotocum* o o o o o Poitras[84], Abdelzaher et al.[80], Czeczuga & Snarka[81], Al-Sheikh & Abdelzaher[97] Pythium elongatum o o Czeczuga et al.[47] Pythium gibbosum o o Czeczuga et al.[47] Pythium globosum o o Czeczuga et al.[47] Pythium gracile o o o Poitras[84], Czeczuga et al.[47] Pythium graminicola o o Czeczuga et al.[47] Pythium hydnosporum o o o o o Czeczuga et al.[47], Czeczuga & Snarka[81], Czeczuga & Muszynska[89] Pythium imperfectum o o Czeczuga et al.[47] Pythium inflatum o o o o o o o Poitras[84], Robertson[85], Czeczuga[86], Czeczuga & Snarka[81], Nam & Choi[87] Pythium insidiosum* o o Presser & Goss[101] Pythium jirovecii o o Czeczuga et al.[47] Pythium junctum o o Uzuhashi et al.[99] Pythium monospermum* o o o o o Poitras[84], Abdelzaher et al.[80], Czeczuga & Snarka[81], Matsiakh et al.[90] Pythium myriotylum o o o Czeczuga & Snarka[81] Pythium oligandrum o o Czeczuga et al.[47] Pythium oopapillum* o o o Matsiakh et al.[90], Nam & Choi[87] Pythium oryzicollum* o o o o Salmaninezhad & Mostowfizadeh-Ghalamfarsa[102] Pythium pachycaule* o o o o Czeczuga et al.[103], Abdelzaher & Kageyama[95] Pythium papillatum o o Czeczuga et al.[47] Pythium periilum o o Czeczuga et al.[47] Pythium periplocum* o o Czeczuga & Muszynska[89] Pythium phragmitis o o Nechwatal et al.[104] Pythium podbielkowskii o o Czeczuga et al.[47] Pythium polysporum o o Czeczuga et al.[47] Pythium pyrilobum o o Czeczuga & Snarka[81] Pythium rhizo-oryzae* o o o Uzuhashi et al.[99] Pythium rishiriense o o Rahman et al.[82] Pythium subutonaiense* o o Chen & Zheng[105] Pythium sukuiense o o Matsiakh et al.[90] Pythium tardicrescens o o Czeczuga et al.[47] Pythium tenue o o o Poitras[84], Czeczuga et al.[47] Pythium thalassium o o El-Hissy et al.[88] Pythium torulosum* o o o o Poitras[84], Czeczuga & Snarka[81], Czeczuga[86], Abdelzaher & Kageyama[95] Pythium utonaiense* o o Uzuhashi et al.[99] Pythium vanterpoolii o o Czeczuga et al.[47] Saprolegniales Achlya ambisexualis o o o o Czeczuga et al.[47], Robideau et al.[106], Hunjavanit et al.[107],
Vega-Ramírez et al.[108]Achlya americana o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Robideau et al.[106], Godlewska et al.[110], Choi et al.[111] Achlya apiculata o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Achlya bisexualis* o o o o Leclerc et al.[112], Robideau et al.[106], Choi et al.[111] Achlya caroliniana o o o o o Srivastava & Srivastava[113], Ogbonna & Alabi[109], Czeczuga et al.[47] Achlya colorata o o Czeczuga et al.[47] Achlya crenulata o o Czeczuga et al.[47] Achlya debaryana o o o Czeczuga et al.[47] Achlya diffusa o o Srivastava & Srivastava[113], Ogbonna & Alabi[109], Czeczuga et al.[47] Achlya dubia o o o Shanor & Saslow[114], Ogbonna & Alabi[109], Czeczuga et al.[47] Achlya flagellata o o Hoshina et al.[115], Shah et al.[116], El-Hissy et al.[88], Ogbonna & Alabi[109] Achlya heterosexualis o o Vega-Ramírez et al.[108] Achlya klebsiana o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Achlya megasperma o o Ogbonna & Alabi[109] Achlya oblongata o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Achlya oligocantha o o Czeczuga et al.[47] Achlya orion o o o o Ogbonna & Alabi[109], Czeczuga et al.[47] Achlya polyandra o o Czeczuga et al.[47], Godlewska et al.[110] Achlya prolifera o o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Achlya proliferoides o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Achlya racemosa o o o Ogbonna & Alabi[109], Czeczuga et al.[47] Achlya rodrigueziana o o Czeczuga et al.[47] Achlya spiralis o o Paul & Steciow[117] Aphanomyces acinetophagu o o Czeczuga et al.[47] Aphanomyces amphigynus o o o El-Hissy et al.[88], Czeczuga et al.[47], Godlewska et al.[110] Aphanomyces astaci o o Unestam & Nylund[118], Czeczuga et al.[47], Godlewska et al.[110], Hsieh et al.[119] Aphanomyces bosminae o o Czeczuga et al.[47] Aphanomyces daphniae o o o Czeczuga et al.[47] Aphanomyces frigidophilus o o o o Ballesteros et al.[120] Aphanomyces helicoides o o o o Srivastava & Srivastava[113], Czeczuga et al.[47] Aphanomyces invadans o o o Iberahim et al.[121] Aphanomyces irregularis o o o o Czeczuga et al.[47] Aphanomyces laevis* o o o o Shanor & Saslow[114], Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Aphanomyces ovidestruens o o Czeczuga et al.[47] Aphanomyces patersonii o o Czeczuga et al.[47] Aphanomyces stellatus o o o o Hoshina et al.[115], Shah et al.[116], Ogbonna & Alabi[109] Dictyuchus carpophorus o o El-Hissy et al.[88] Dictyuchus magnusii o o El-Hissy et al.[88] Dictyuchus monosporus o o El-Hissy et al.[88] Dictyuchus sterilis o o El-Hissy et al.[88], Godlewska et al.[110] Geolegnia helicoides* o o o o Steciow et al.[122], Nam et al.[123] Leptolegnia caudata* o o o o o o o o o El-Hissy et al.[88], Czeczuga et al.[47], Nam et al.[123] Newbya androgyna o o o El-Hissy et al.[88], Czeczuga et al.[47], Godlewska et al.[110] Protoachlya papillata o o o Ogbonna & Alabi[109], Czeczuga et al.[47] Pythiopsis cymosa o o El-Hissy et al.[88], Czeczuga et al.[124] Saprolegnia aenigmatica* o o o Sandoval-Sierra et al.[36], Nam et al.[123] Saprolegnia anisospora o o o o o Czeczuga et al.[47], Masigol et al.[125] Saprolegnia asterophora* o o o o o o Czeczuga et al.[47], Godlewska et al.[110], Nam et al.[123] Saprolegnia australis* o o o Vega-Ramírez et al.[108], Rezinciuc et al.[37], Pavić et al.[126], Nam et al.[123] Saprolegnia crustosa o o Czeczuga et al.[47] Saprolegnia delica* o o o o o o Czeczuga et al.[47], Godlewska et al.[110], Pavić et al.[126], Nam et al.[123] Saprolegnia diclina o o o o o o o Czeczuga et al.[47], Fernández-Benéitez et al.[39], Hunjavanit et al.[107], Godlewska et al.[110], Vega-Ramírez et al.[108], Martín-Torrijos et al.[127], Sadinski et al.[128], Nam et al.[123] Saprolegnia eccentrica o o Czeczuga et al.[47] Saprolegnia ferax* o o o o o o Blaustein et al.[129], Czeczuga et al.[47], Romansic et al.[41], Prada-Salcedo et al.[130], Godlewska et al.[110], Vega-Ramírez et al.[108], Groffen et al.[131], Nam et al.[123] Saprolegnia glomerata o o o o o Czeczuga et al.[47], Godlewska et al.[110], Vega-Ramírez et al.[108] Saprolegnia hypogna o o Godlewska et al.[110] Saprolegnia latvica o o Czeczuga et al.[47] Saprolegnia litoralis o o Ogbonna & Alabi[109], Czeczuga et al.[47] Saprolegnia megasperma o o o Nolan[132] Saprolegnia monilifera o o o o Czeczuga et al.[47], Godlewska et al.[110] Saprolegnia monoica o o o o Czeczuga et al.[47], Godlewska et al.[110] Saprolegnia parasitica* o o o o o o Czeczuga et al.[47], Godlewska et al.[110], Vega-Ramírez et al.[108],
Shin et al.[133], Pavić et al.[126], Nam et al.[123]Saprolegnia pseudocrustosa o o Czeczuga et al.[47] Saprolegnia salmonis* o o Hussein et al.[38], Nam et al.[123] Saprolegnia terrestris o o o Vega-Ramírez et al.[108] Saprolegnia torulosa* o o o o o o Godlewska et al.[110], Czeczuga et al.[47], Nam et al.[123] Saprolegnia turfosa o o Godlewska et al.[110] Saprolegnia uliginosa o o o o Czeczuga et al.[47], Vega-Ramírez et al.[108] Saprolegnia unispora o o o o o Czeczuga et al.[47], Godlewska et al.[110], Vega-Ramírez et al.[108] Thraustotheca clavata o o o Ogbonna & Alabi[109], Czeczuga et al.[47], Godlewska et al.[110] Species reported in Korea are shown in bold. Asterisk means oomycetes species isolated in the present study. Of the known members of freshwater oomycetes, saprotrophic oomycetes have been poorly studied in Korea and, therefore, underestimated. The exploration of their diversity and ecology is pivotal for a more profound comprehension of their function in aquatic ecosystems. To bridge this gap, an initial survey using a cultivation approach was deployed from 2016 to 2021. In this study, congeners of the Peronosporales (Phytophthora), Pythiales (Elongisporangium, Globisporangium, Phytopythium and Pythium), and Saprolegniales (Achlya, Aphanomyces, Geolegnia, Leptolegnia and Saprolegnia) are reported. Furthermore, this study identified and included eight novel species of Pythiales (Phytopythium and Pythium). This study contributes to our knowledge of the diversity and ecology of oomycetes in freshwater environments. Moreover, this study attempts to understand the substrate preference and spatial and temporal distribution of these oomycetes in Korea.
-
Between 2016 and 2021, samples of algae, animal debris, plant litter, soil sediment, and water were collected from different freshwater environments, including mountain streams, reservoirs, and lowland rivers in Korea (Fig. 1). The data on monthly mean water temperatures and average monthly air temperatures from each survey area were compiled utilizing two primary sources; the water environment information system of the National Institute of Environmental Research (
https://water.nier.go.kr ) and the weather data portal of the Korean Meteorological Administration (https://data.kma.go.kr ). Oomycete strains from soil sediment and water samples were isolated using a simple plating method on 5% V8 agar (V8A; 50 mL clarified V8 juice, 10 g CaCO3, 15 g agar, 950 mL deionized water). For substrates involving algal, animal, and plant debris, samples were rinsed with distilled water three times, cut into 2 mm2 sections, and then plated onto 5% V8A plates. To suppress bacterial and fungal growth, 15 ppm of rifampicin and 20 ppm of nystatin were added to the media. After the mycelia were formed, a hyphal tip was isolated and transferred onto a new 20% V8A plate, containing 200 mL clarified V8 juice, 10 g CaCO3, 15 g agar, and 800 mL deionized water. Representative strains were deposited in the Nakdonggang National Institute of Biological Resources (NNIBR, Sangju, Korea) or the Korean Agricultural Culture Collection (KACC).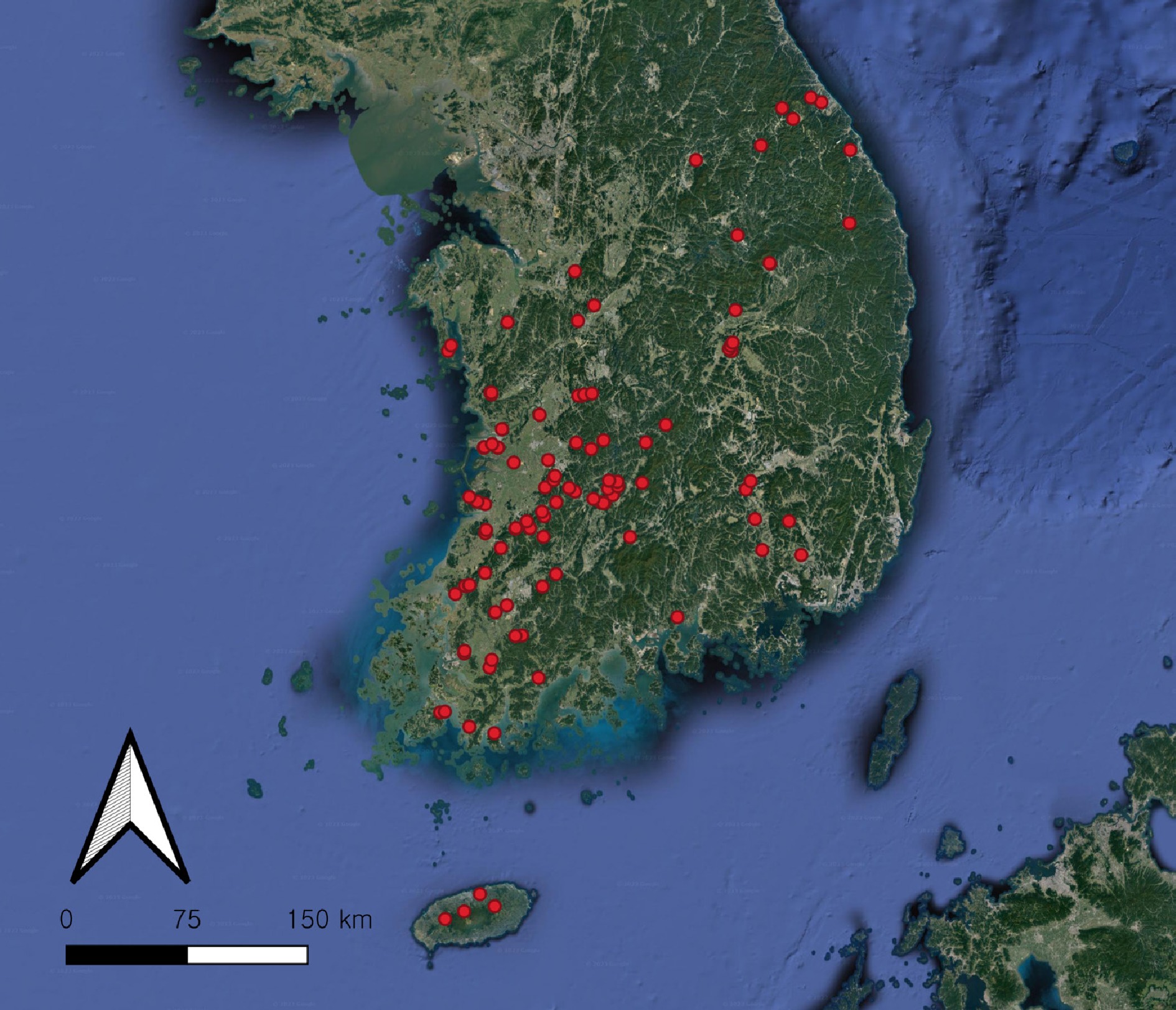
Figure 1.
Location of the 121 sampling sites (red dots) in South Korea. The map was created using QGIS v.3.24.
Cultural and morphological characterization
-
The strains were inoculated onto different agar media, including 20% V8A, potato dextrose agar (PDA; Difco, Detroit, MI, USA), and corn meal agar (CMA; Difco, Detroit, MI, USA). After the incubation for 2–3 d at 25 °C in the darkness, colony patterns were recorded. After 7–14 d, sporangia, gametangia, and chlamydospores on V8A[134] were observed using an Axio Imager A2 microscope (Carl Zeiss, Oberkochen, Germany) and Axiocam 512 color camera (Carl Zeiss, Oberkochen, Germany). The sterile distilled water was added to the surface of the colony on V8A, and the isolates were placed at 5 °C for 24 h[87], to induce the formation of sporangia or oospores. Alternatively, the morphology of zoosporangia, oogonia, oospores, and antheridia was investigated in a grass-leaf water culture[135] with Korean lawngrass (Zoysia sp.). Fifty measurements were conducted for each observed structure. The minimum, optimum, and maximum growth temperatures of the novel species were determined by conducting growth trials at various temperatures: 5, 10, 15, 20, 25, 30, 35, 40, and 45 °C, on V8A with five replicates.
DNA extraction, sequencing, and phylogenetic analysis
-
Genomic DNA was extracted from 7-day-old cultures using the MagListo 5M plant Genomic DNA Extraction Kit (Bioneer, Daejeon, Korea) following the manufacturer's instructions. The regions of internal transcribed spacer (ITS) rDNA and cytochrome c oxidase subunit I (cox1) were amplified with polymerase chain reaction (PCR), along with cytochrome c oxidase subunit II (cox2) mtDNA, for the isolates of the new species. The primer pairs used in PCR were ITS1/ITS4[136], OomCox1-levup/OomCox1-levlo[106], and cox2-F[137]/cox2-RC4[138], respectively. The amplicons were purified with the AccuPrep PCR Purification Kit (Bioneer, Daejeon, Korea) and sequenced by the Sanger method in Macrogen Inc. (Seoul, Korea). The sequences were edited using the DNAStar software package 5.05 (DNAStar, Inc., Madison, WI, USA) and subjected to an NCBI BLAST search to reveal the closest related sequences in the National Center for Biotechnology Information (NCBI) GenBank database. For phylogenetic analysis, previously published sequences of authentic oomycete strains were retrieved from GenBank. These reference sequences were included in the datasets created by aligning the sequences from the present study, using the G-INS-I algorithm[139] of MAFFT version 7 (
https://mafft.cbrc.jp/alignment/server/ )[140]. Phylogenetic trees were reconstructed using maximum likelihood (ML) and minimum evolution (ME) inferences in MEGA version 10.2[141]. The ML and ME analyses were carried out using the Tamura-Nei model at 1,000 bootstrap replicates (BS). Multilocus phylogenetic analysis for new species used a combined ITS, cox1, and cox2 dataset, which was produced by concatenating the sequences of the individual markers in SequenceMatrix v1.7.8[142]. The phylogenetic tree was reconstructed using the multi-locus sequence in MEGA as mentioned above. -
A total of 568 isolates were obtained from lowland rivers (ca. 25%), mountain streams (ca. 35%), and reservoirs (ca. 40%). These isolates were recovered during two distinct seasons: the cold to cool (CC) season (January–April), which accounted for ca. 46% of the total, and the warm to hot (WH) season (May–September), which comprised ca. 54%. During the CC season, the monthly mean air and water temperatures at the sampling sites were below 15 °C, while during the WH season, they crossed above 15 °C. These isolates originated from diverse freshwater substrates, including algae, animal debris, plant litter, soil sediment, and the water itself. Over half the strains (ca. 55%) were isolated from plant-derived materials like decaying leaves, stems, and twigs, suggesting that they served as the dominant isolation sources of the oomycete strains. The remaining strains were found in the soil (ca. 20%), water (ca. 12%), algae (ca. 5%), and animal sources (ca. 8%), such as fish, snail, or frog spawn. The strain information and collection details are outlined in Supplementary Table S1. The strains were identified based on cultural and morphological features, along with phylogenetic analyses. This process resulted in the recognition of 78 taxa, comprising 57 published species and 21 undescribed taxa of oomycetes in the orders Peronosporales (4%), Pythiales (77%), and Saprolegniales (19%).
Peronosporales – Phytophthora
-
In the phylogenetic tree based on cox1 gene sequences (Supplementary Fig. S1), the Korean strains formed a well-supported group with Phytophthora chlamydospora, Ph. inundata, and Ph. sp. 'lagoariana' with maximum support value in the ME and ML analyses. Based on a BLASTn search, the ITS and cox1 sequences of the Korean strains of Ph. chlamydospora were identical to those of the authentic isolate CPHST BL156 (MG865471 in ITS, MH136867 in cox1). Colony growth pattern and morphological characteristics of Ph. chlamydospora strains were identical to those in the original description by Hansen et al.[60]. Strains W655 and W675 were identified as Ph. sp. 'lagoariana', following the description provided by Wallace[73]. The cox1 sequences were identical to those of the isolate P8223 (HQ261441) and P11904 (GU594812) of Ph. sp. 'lagoariana', while ITS had 10–12 bp sequence differences (P8223; EF590256, P11904; FJ801644). Strain W1757 matched the strain CPHST BL 20 (MG865516 in ITS, MH136910 in cox1) of Ph. inundata with sequence similarities of 100% (847/847 bp) in ITS and 99.87% (747/748 bp) in cox1. This species has been previously unrecorded in Korea. The cultural and morphological characteristics of this strain were described in Supplementary Taxonomy S1, in comparison with the description by Brasier et al.[71]. Images of the colonies and taxonomically significant structures are shown in Figs 2 and 3.
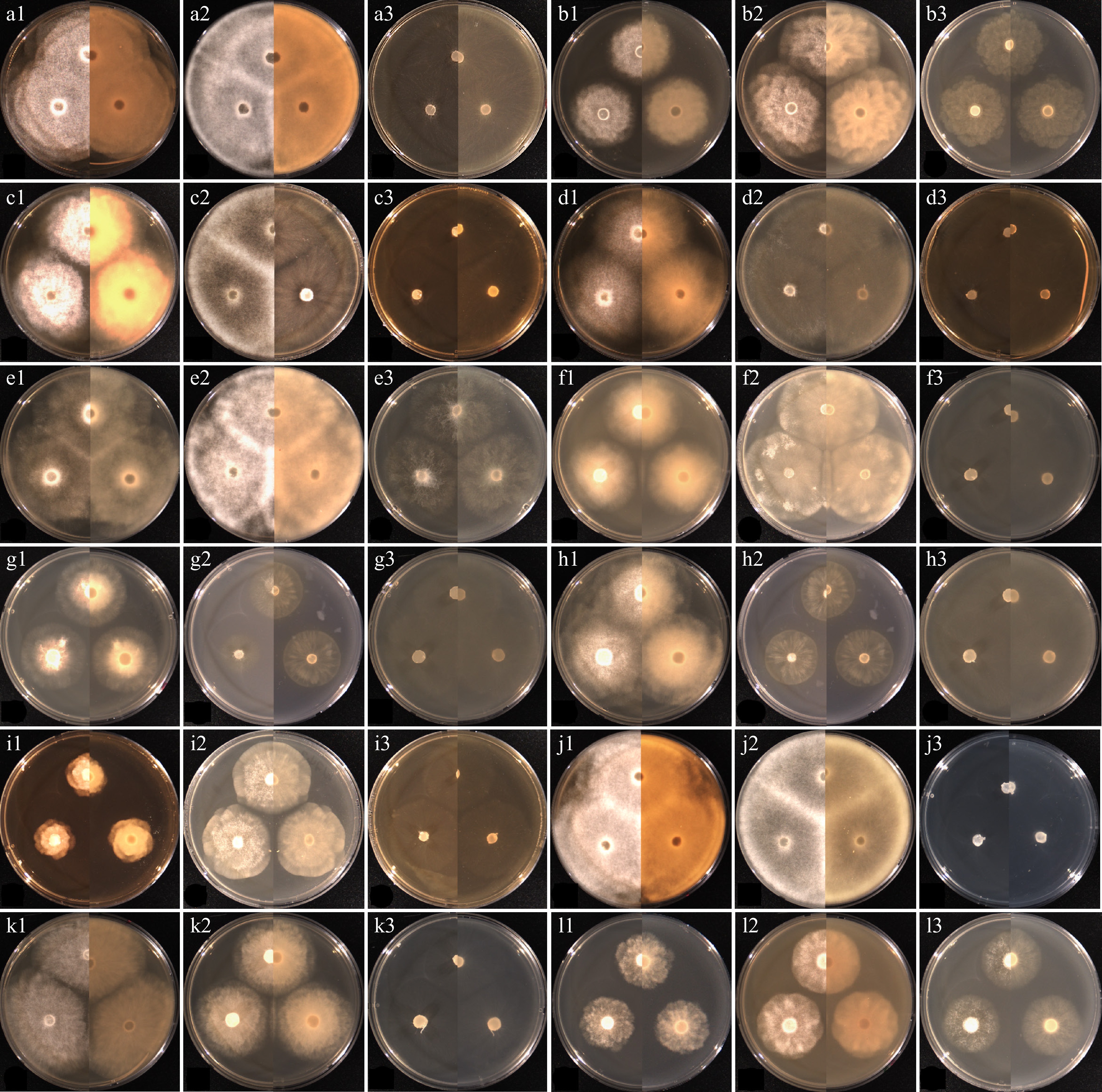
Figure 2.
Cultural characteristics of the isolates of Elongisporangium, Globisporangium, and Phytophthora species, which have previously been unrecorded in Korea, obtained from this study. (a) Elongisporangium undulatum W923, (b) Globisporangium alternatum W1167, (c) G. attrantheridium W786, (d) G. macrosporum W716, (e) G. marsipium W1296, (f) G. nagaii W1534, (g) G. paddicum W1603, (h) G. rostratifingens W1608, (i) G. selbyi W743, (j) G. sylvaticum W769, (k) G. terrestre W2010, and (l) Phytophthora inundata W1757 on (1) PDA, (2) V8A, (3) CMA, after 72 h at 25 °C (left: observed view and right: reverse view).
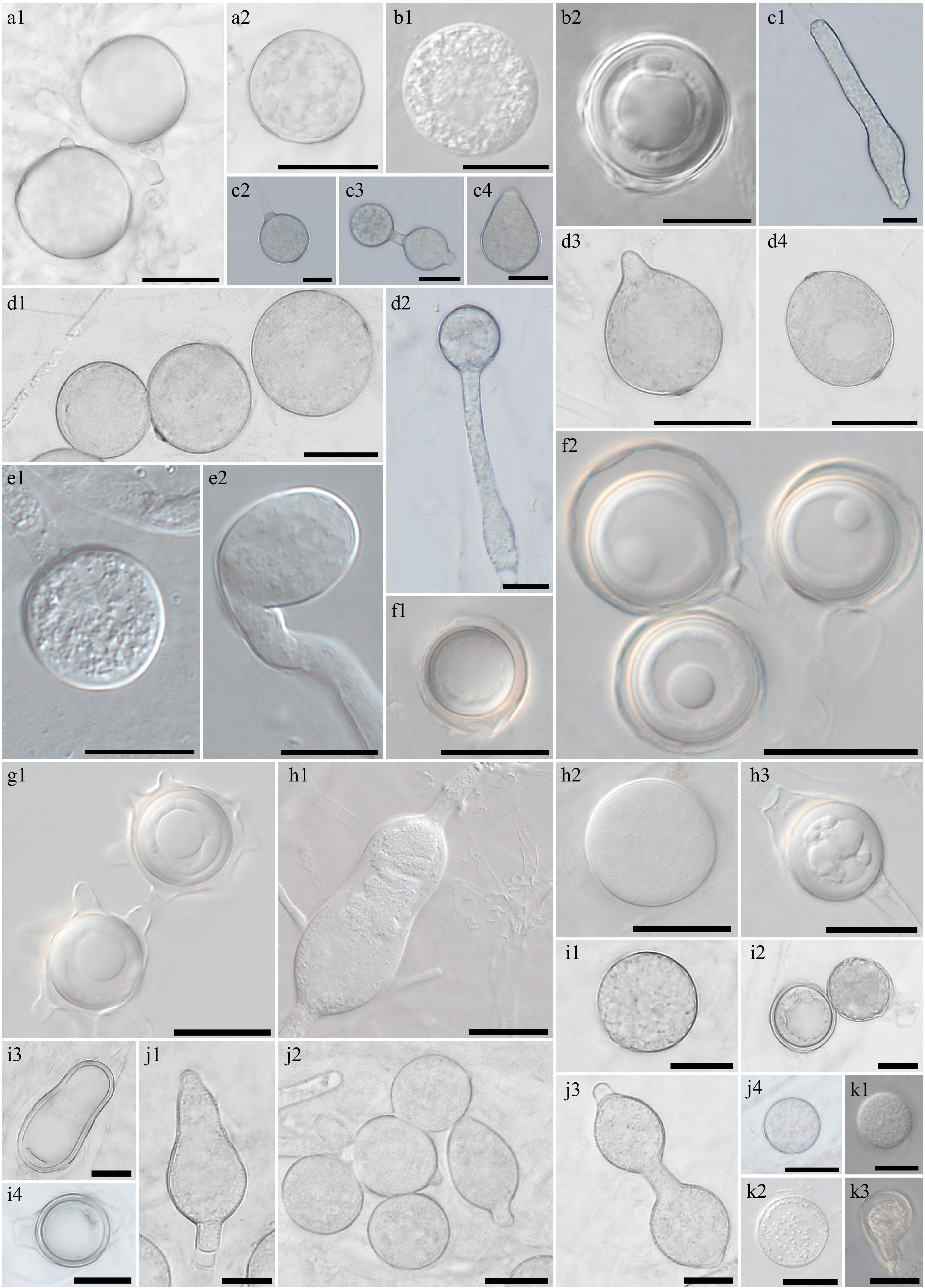
Figure 3.
Morphological characteristics of Elongisporangium, Globisporangium, and Phytophthora species, which have previously been unrecorded in Korea, obtained from this study. Microscopic structures observed under a microscope: (a1), (a2) Chlamydospores of Elongisporangium undulatum W923, (b1) sporangium, (b2) oogonium and oospore of Globisporangium alternatum W1167, (c1), (c3), (c4) hyphal swellings, and (c2) sporangium of (d1)–(d4) G. attrantheridium W786, sporangia of G. macrosporum W716, (e1) sporangium, (e2) hyphal swelling of G. marsipium W1296, (f1), (f2) oogonia and oospores of G. nagaii W1534, (g1) oogonia with ornamentation of G. paddicum W1603, (h1), (h2) sporangia, oogonium and (h3) oospore of G. rostratifingens W1608, (i1) sporangia and (i2)–(i4) oogonia of G. selbyi W743, (j1)–(j4) hyphal swellings of G. sylvaticum W769, (k1), (k2) sporangium, and (k3) oogonium of G. terrestre W2010 (scale bars: a1, a2, d1–k3 = 20 μm, b1–c4 = 10 μm).
Pythiales–Elongisporangium, Globisporangium, Phytopythium (= Ovatisporangium), and Pythium sensu stricto
-
In the present study, 82 of the Korean strains were classified under the genus Globisporangium, which consists of clades E–G, I, and J of Pythium Pringsheim (nom. cons.) sensu lato (s.l.) sensu Lévesque & de Cock[143]. The strains identified within this genus were Globisporangium alternatum, G. attrantheridium, G. heterothallicum, G. intermedium, G. irregulare, G. macrosporum, G. marsipium, G. nagaii, G. paddicum, G. rostratifingens, G. selbyi, G. spinosum, G. sylvaticum, G. terrestre, and G. ultimum sensu stricto (s.s.), all of which develop globose sporangia. In contrast, from the genus Elongisporangium, only E. undulatum was isolated. This strain, characterized by its elongated sporangia, was previously classified as clade H of Pythium s.l. sensu Lévesque & de Cock[143]. In the phylogenetic analyses, the groupings of Korean strains with the reference isolates for each species were supported by high bootstrap values ranging from 99% to 100% (Supplementary Fig. S1). Moreover, the BLASTn search results for the ITS and cox1 sequences of each Korean strain revealed high similarities of 99%–100% with the published sequences of authentic isolates. Cultural and morphological features of these species, which have previously not been recorded in Korea, were described in Supplementary Taxonomy S1. For selected known species, colonies and taxonomically important structures such as sporangia, oogonia, or oospores are shown in Figs 2 and 3.
Phytopythium (syn. Ovatisporangium) species were also found in the present study (64 strains). The Korean strains formed four well-supported groups, matching four distinct species, Phytopythium chamaehyphon, Pp. helicoides, Pp. litorale, and Pp. vexans, in the phylogenetic tree (Supplementary Fig. S1). The groupings of the Korean strains with each reference isolate were supported by high bootstrap values (99%–100%). Based on a BLASTn search of the ITS and cox1 sequences, 19 strains (e.g., W1307, W1308, W1322, W1389, and W1393) were identical to Pp. chamaehyphon CBS259.30 (HQ643374 in ITS, HQ708421 in cox1), 11 strains (e.g. W671, W682, W687, W688, and W1291) to Pp. helicoides CBS286.31 (HQ643383 in ITS, EF408875 in cox1), 14 strains (e.g. W595, W628, W629, W630, and W1193) to Pp. litorale CBS118360 (HQ643386 in ITS, HQ708433 in cox1), and ten strains (W707, W708, W714, W943, and W1000) to Pp. vexans CBS455.62 (HQ643386 in ITS, HQ708448 in cox1). Each group was morphologically identified, referencing the original descriptions of Pp. chamaehyphon (CBS259.30)[144], Pp. helicoides (CBS286.31)[145], Pp. litorale (CBS118360)[146], and Pp. vexans (CBS119.80)[147].
Two new species of Phytopythium were discovered in the present study. The strains W676, W1152, W1998, and W1999 formed a well-supported group in the multi-gene phylogenetic tree (Fig. 4), and this group is close to Pp. citrinum CBS119171, Pp. delawarense 382B, Pp. aichiense CBS137195 and Pp. litorale CBS118360. The cox1 sequences of the new strains were closest to those of Pp. citrinum CBS119171 (HQ708422), with a sequence similarity of 98.91% (635/642 bp) in the BLASTn search. The ITS and cox2 sequences were a 97% match to the strain CBS119171 (HQ643375.2 in ITS, AB690679 in cox2). The ITS and cox2 sequences were identical to those of an undescribed isolate UZ230 (AB468792 in ITS and AB468917 in cox2); however, the cox1 sequence of UZ230 is not available in the GenBank database. The strain W1435 occupied a unique branch in the phylogenetic trees, although it showed a close phylogenetic relation to Pp. iriomotense GUCC0025 with sequence similarities of 97% (625/642 bp) in cox1 (AB690652) and 97% (507/521 bp) in cox2 (AB690682). The cultural, morphological, and physiological characteristics of these novel species will be described in the following Taxonomy section.
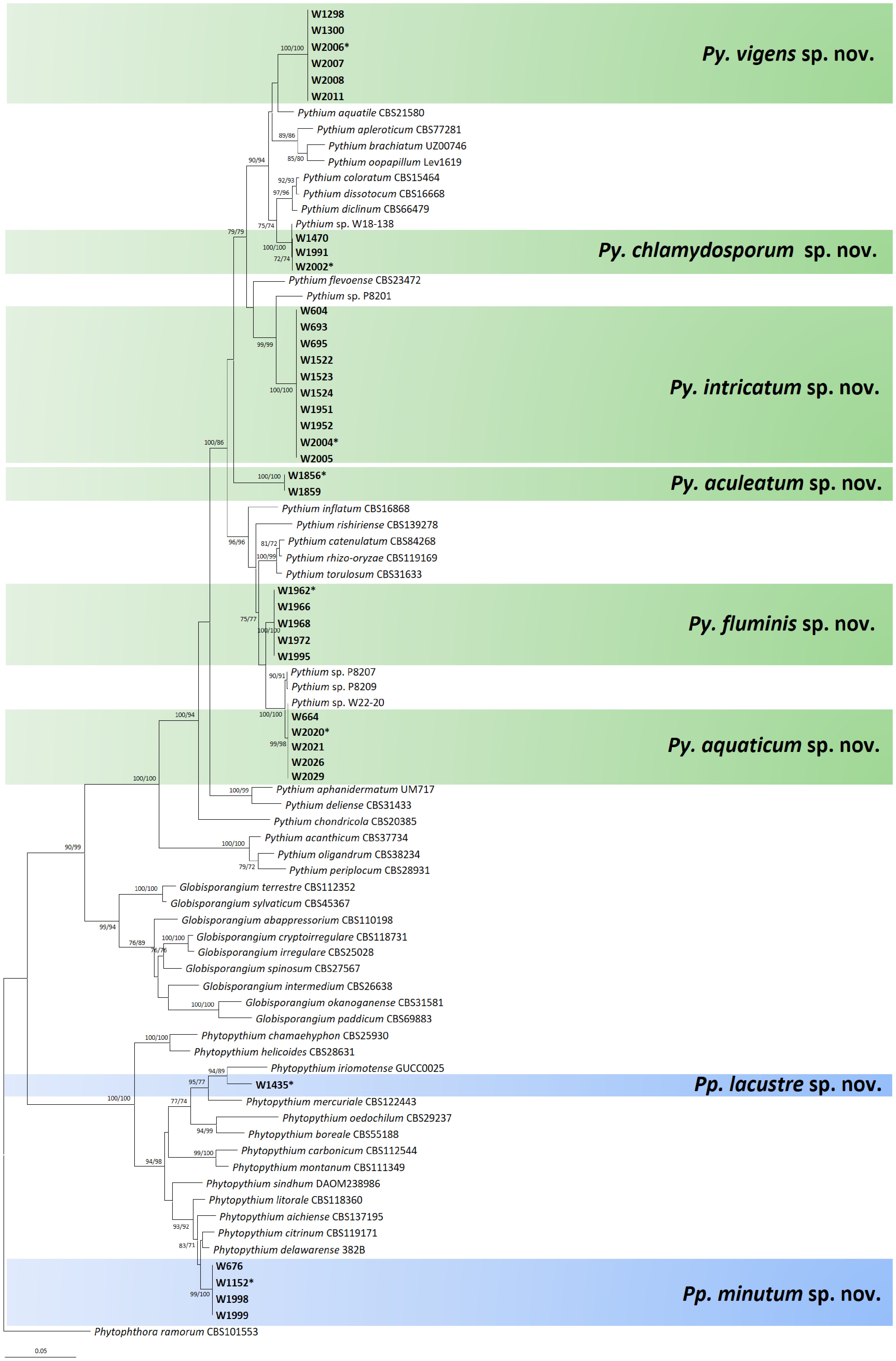
Figure 4.
Multigene phylogenetic tree of Phytopythium and Pythium species from the minimum evolution analysis based on internal transcribed spacer (ITS), cytochrome c oxidase subunit I (cox1), and II (cox2) mtDNA sequences. Bootstrapping values (minimum evolution BP/maximum likelihood BP) higher than 70% were given above or below the branches (1,000 replicates). The strains of new species obtained from this study are shown in bold (ex-type culture with asterisk). Phytophthora ramorum was used as an outgroup. The scale bar equals the number of nucleotide substitutions per site.
This study identified 21 species of Pythium sensu stricto (s.s.) sensu Uzuhashi et al.[148] and Nguyen et al.[149] (308 strains), namely, Py. acanthicum, Py. aphanidermatum, Py. apleroticum, Py. aquatile, Py. biforme, Py. brachiatum, Py. catenulatum, Py. cedri, Py. diclinum, Py. dissotocum, Py. insidiosum, Py. lutarium, Py. monospermum, Py. oopapillum, Py. oryzicollum, Py. pachycaule, Py. periplocum, Py. rhizo-oryzae, Py. subutonaiense, Py. torulosum, and Py. utonaiense. These species belonged to clades A–D of Pythium s.l. as classified in the phylogenetic studies by Lévesque & de Cock[143] and Uzuhashi et al.[148] and are characterized by their filamentous sporangia. In the cox1 tree (Supplementary Fig. S1), the Korean strains formed well-supported groups with each reference of Pythium s.s., which is supported with high bootstrap support. A BLASTn search revealed that the sequences of each Korean strain matched the published sequences of authentic isolates with high similarities of 99%–100%. The taxonomic characters of the previously unrecorded species in Korea were investigated in the present study. The cultural and morphological characteristics of these species were described in the supplementary data (Supplementary Taxonomy S1), with Figs 5 and 6 illustrating the colonies and taxonomically significant structures of representative strains. Morphological characteristics of Korean isolates were consistent with the descriptions of van der Plaats-Niterink[150].
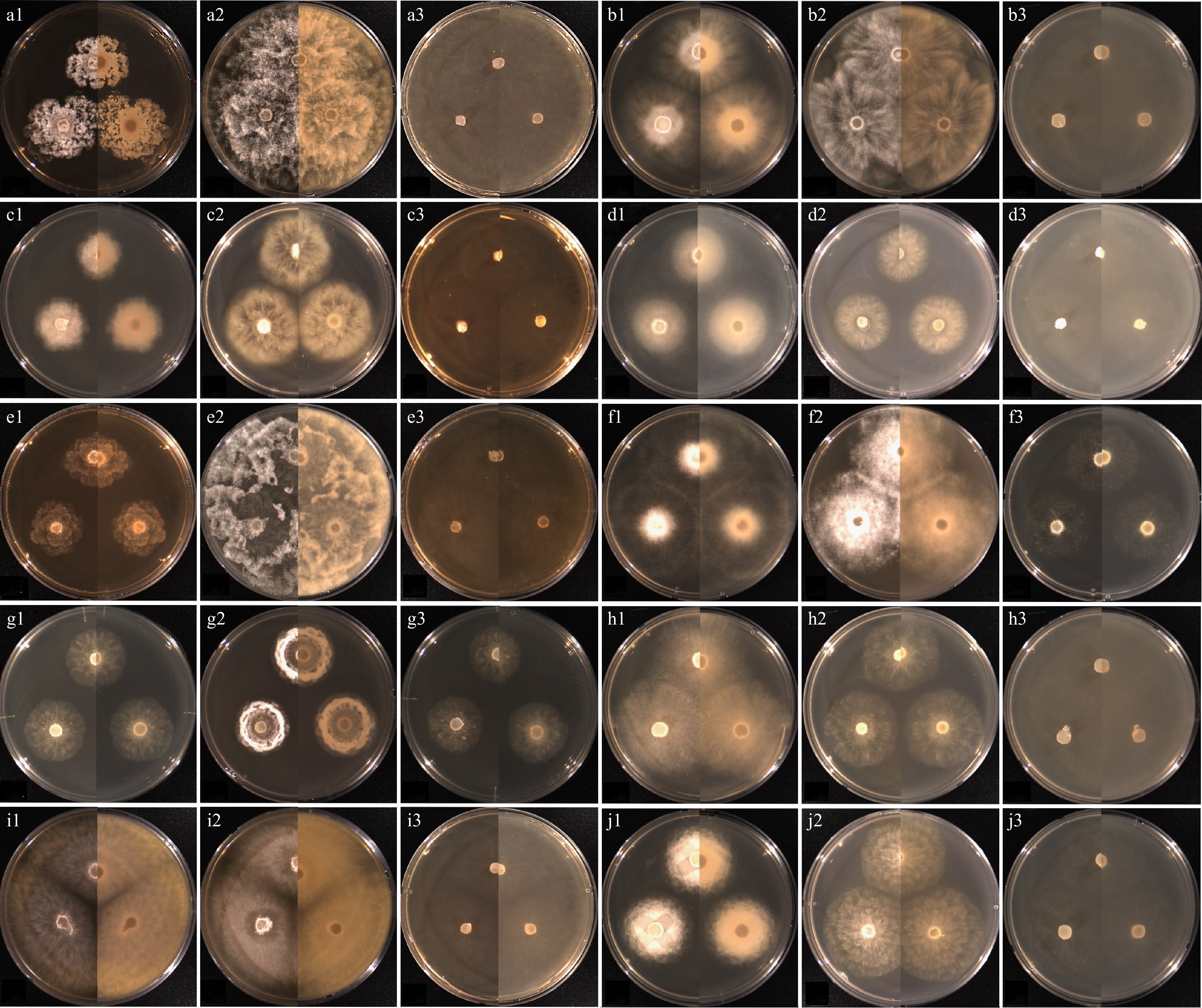
Figure 5.
Cultural characteristics of the isolates of Pythium species, which have previously been unrecorded in Korea, obtained from this study. (a) Pythium acanthicum W927, (b) Py. apleroticum W2032, (c) Py. biforme W813, (d) Py. brachiatum W1541, (e) Py. cedri W754, (f) Py. insidiosum W1285, (g) Py. oryzicollum W1211, (h) Py. pachycaule W1650, (i) Py. subutonaiense W929, and (j) Py. utonaiense W1684 on (1) PDA, (2) V8A, (3) CMA, after 72 h at 25 °C (left: observed view and right: reverse view).
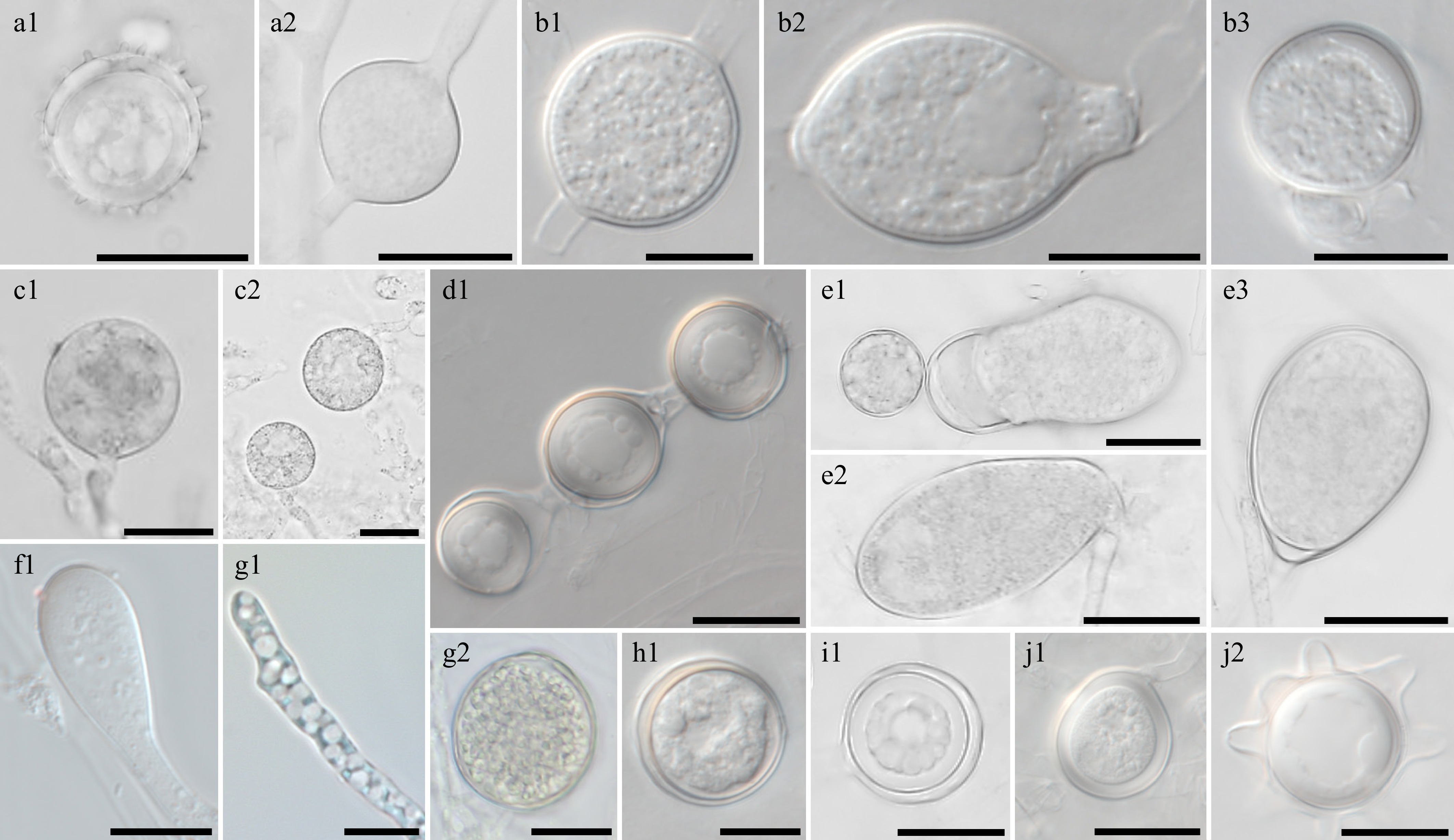
Figure 6.
Morphological characteristics of Pythium species, which have previously been unrecorded in Korea, obtained from this study. Microscopic structures observed under a microscope: (a1) oogonium and (a2) sporangium of Pythium acanthicum W927, (b1), (b3) oogonium and (b2) hyphal swellings of Py. apleroticum W2032, (c1), (c2) hyphal bodies of Py. biforme W813, (d1) oogonia in a chain of Py. brachiatum W1541, (e1)–(e3) sporangia of Py. cedri W754, (f1) Py. insidiosum W1285, (g1) Py. oryzicollum W1211, (g2) oogonium and oospore of Py. oryzicollum W1211, (h1) Py. pachycaule W1650, (i1) Py. subutonaiense W929, and (j1), ( j2) Py. utonaiense W1684 (scale bars: a1, a2, c1–e3, g2, i1–j2 = 20 μm, b1–b3, f1, g1, h1 = 10 μm).
We also identified six new species within Pythium s.s. The strains W1856 and W1859 formed a well-supported group in the phylogenetic tree (Fig. 4). The cox1 sequences of these strains were most closely related to P. flevoense isolate CBS23372 (HQ708583, 587/621 bp) with 94.5% similarity in the BLASTn search. Their ITS and cox2 sequences were identical to those of an undescribed isolate ZSF0011 (AB468820 in ITS and AB468939 in cox2), but the cox1 sequence of ZSF0011 is not available at GenBank.
Strains W604, W693, W695, W1522, W1523, W1524, W1951, W1952, W2004, and W2005 were related to an undescribed isolate P8201 with 96% similarity in the BLASTn search of the cox1 (HQ261483) and cox2 (GU222160) sequences (619/645 bp in cox1 and 489/508 in cox2). The ITS sequence was also a 96% match to isolate P8201 (GU258928, 748/777 bp). In the BLASTn search of ITS, the Korean strains were a 99.9% match to various strains of Pythium sp. 'group B' DP-2020[126], e.g., DG26 (MT555835) and UPG14 (MT555842); however, these isolates have not yet been formally described. Their cox1 and cox2 sequences are not available at GenBank.
Another new taxon included strains W1470, W1991, and W2002, which formed a well-supported cluster in the phylogenetic tree. ITS and cox1 sequences of these strains were identical with an undescribed strain CLE-2015d (KT247392 in ITS and KT247393 in cox1). The cox2 sequence CLE-2015d is not available at GenBank. The Korean strains had identical cox2 sequences as an undescribed strain W18-138 (PX437885). The ITS and cox1 sequences of Korean strains matched those of the strain W18-138 with 99% similarity (PX430644 in ITS and PX437881 in cox1).
The strains W1298, W1300, W2006, W2007, W2008, and W2011 constituted another novel lineage. Their ITS, cox1, and cox2 sequences exhibited a sequence similarity of 97% with Py. aquatile CBS21580 (HQ643445 in ITS, HQ708492 in cox1, and KJ595355 in cox2), Py. apleroticum CBS77281 (HQ643444 in ITS and HQ708491 in cox1), and Py. brachiatum UZ00746 (KJ995583 in ITS and KJ995594 in cox1).
The strains W664, W2020, W2021, W2026, and W2029 formed a well-supported cluster in the phylogenetic tree. These strains had identical ITS, cox1, and cox2 sequences as an undescribed isolate W22-20 (PX430645 in ITS, PX437883 in cox1, and PX437886 in cox2). The ITS of another undescribed isolate, P8207 (HQ261734), was identical to this new taxon, while cox1 (HQ261481) and cox2 (GU222161) sequences displayed similarities of 99% (634/637 bp in cox1 and 521/523 in cox2).
The next new group, strains W1962, W1966, W1968, W1972, and W1995, is close to a cluster containing the new taxa group mentioned above (W664, W2020, W2021, W2026, and W2029); however, their ITS and cox1 sequence similarities were 96.5%, and cox2 was 98.5%. The ITS sequences of the last new group displayed similarities of 99% to species such as Py. catenulatum CBS842.68 (AY598675.2) and Py. rhizo-oryzae CBS119169 (HQ643757.2). The cox1 and cox2 sequences showed the similarities of 96%–97% to Py. catenulatum CBS842.68 (HQ708540 in cox1 and KJ595404 in cox2) and Py. rhizo-oryzae CBS119169 (HQ708798 in cox1 and KJ595420 in cox2).
The cultural, morphological, and physiological characteristics of six novel taxa were described in the Taxonomy section.
Saprolegniales–Achlya, Aphanomyces, Geolegnia, Leptolegnia, and Saprolegnia
-
In the ITS tree of the Saprolegniales (Supplementary Fig. S2), 98 Korean strains grouped with multiple known species such as Achlya bisexualis, Aphanomyces laevis, Geolegnia helicoides, Leptolegnia caudata, Saprolegnia aenigmatica, S. asterophora, S. australis, S. diclina, S. ferax, S. parasitica, and S. torulosa. A BLASTn search revealed that the Korean strains shared high sequence similarities with previously published sequences of authentic isolates. Prior studies have documented both morphological and phylogenetic descriptions of the Korean strains of A. bisexualis[111] and Saprolegniaceae species[123].
Substrate, season, and habitat-based distribution of oomycetes
-
Identification of oomycete strains isolated from various sources revealed their preference for plant-derived sources, including decaying leaves, stems, and twigs. Of the 78 identified taxa, 60 were isolated from plant-derived substrates. These taxa were assigned to the Aphanomyces, Elongisporangium, Geolegnia, Globisporangium, Leptolegnia, Phytophthora, Phytopythium, Pythium s.s., or Saprolegnia. In contrast, other taxa were isolated from soil sediments and/or water but not found in the plant sources in the present study; A. laevis, E. undulatum, G. macrosporum, G. selbyi, G. sylvaticum, G. ultimum, Py. acanthicum, Py. apleroticum, Py. monospermum, Py. oryzicollum, Py. torulosum, and S. parasitica. Two Saprolegniales species (Achlya bisexualis and Saprolegnia australis) were isolated only from animal sources, such as fish and marsh snails. Pythiales was isolated mainly from the plant sources (ca. 56%); however, Phytopythium vexans, Pythium catenulatum, Py. diclinum, Py. oopapillum, Py. rhizo-oryzae, Py. subutonaiense, and Py. torulosum were also isolated from marsh snails or frog eggs (ca. 5%). Phytopythium litorale, Pythium apleroticum, Py. diclinum, and Py. oopapillum were also isolated from the algae (4%). From water, Elongisporangium undulatum, Globisporangium attrantheridium, G. spinosum, Phytopythium chamaehyphon, Pythium apleroticum, Py. diclinum, Py. lutarium, Py. oopapillum, Py. oryzicollum, Py. pachycaule, Py. periplocum, Py. subutonaiense were found (ca. 10%). The genus Globisporangium was mainly isolated from soil (ca. 54%), and diverse species were identified: Globisporangium attrantheridium, G. heterothallicum, G. macrosporum. G. marsipium, G. rostratifingens, G. selbyi, G. spinosum, G. sylvaticum, G. terrestre, G. ultimum, and two unknown new taxa. Elongisporangium, Phytopythium, and Pythium s.s. were also isolated from soil; Elongisporangium undulatum, Phytopythium chamaehyphon, Pp. helicoides, Pp. vexans, Pythium acanthicum, Py. aphanidermatum, Py. aquatile, Py. biforme, Py. catenulatum, Py. diclinum, Py. dissotocum, Py. lutarium, Py. monospermum, Py. oopapillum, Py. rhizo-oryzae, Py. subutonaiense, and Py. torulosum. 24% of the total strains of Pythiales were found in soil sediments.
Across both seasons, the Pythiales were frequently isolated, accounting for ca. 86.4% of the isolates obtained in WH and ca. 73.4% of those obtained in the CC season. Among congeners of Pythium s.s., 12 taxa were observed in both WH and CC seasons; Py. aquatile, Py. catenulatum, Py. diclinum, Py. dissotocum, Py. lutarium, Py. oopapillum, Py. pachycaule, Py. subutonaiense, and four undescribed taxa. However, 12 taxa, Py. acanthicum, Py. aphanidermatum, Py. apleroticum, Py. insidiosum, Py. monospermum, Py. oryzicollum, Py. periplocum, Py. rhizo-oryzae, Py. torulosum and three undescribed taxa were exclusively discovered in the WH season, while ten taxa, Py. biforme, Py. brachiatum, Py. cedri, Py. utonaiense and six undescribed taxa were unique to the CC season. In the case of Globisporangium, more taxa were found in the CC season (14 taxa) than in the WH season (ten taxa). Several species, including G. alternatum, G. irregulare, G. nagai, G. paddicum, G. rostratifingens, G. selbyi, and G. sylvaticum, were isolated only in the CC season, while G. marsupium and G. ultimum were exclusive to the WH season. In Phytopythium, which was increased sharply in the WH season, a total of seven taxa were found in the present study. In the CC season, only Pp. vexans and an unknown taxon were found, while diverse species were isolated in the WH season; Pp. chamaehyphon, Pp. helicoides, Pp. litorale, Pp. vexans, and three unknown taxa.
In the case of Saprolegniales, the proportion among isolates was ca. 12.6% in WH and ca. 22.8% in the CC season. Species richness was eight taxa in the WH and nine taxa in the CC seasons. However, the proportions of individual taxa were different between the two seasons; Geolegnia helicoides, Saprolegnia parasitica, S. salmonis, S. torulosa, and two undescribed taxa were exclusively found in the CC season. Conversely, Achlya bisexualis, Aphanomyces laevis, Leptolegnia caudata, S. aenigmatica, S. asterophora, and S. australis were found only in the WH season. S. diclina and S. ferax were found in both seasons, but accounting for ca. 81% of the isolates in the CC and ca. 28% in the WH season. Finally, order Peronosporales represented a small proportion; ca. 1% in WH and ca. 4% CC season.
The Pythiales was frequently isolated across the different types of freshwater ecosystems (ca. 89% in reservoir, 74% in lowland river, and 76% in mountain stream). At the genus level, Pythium s.s. was the most isolated genus in all types (ca. 59% in reservoir, 60% in lowland river, and 45% in mountain stream). The proportion of Globisporangium among isolates was ca. 17% in reservoir, 6% in lowland river, and 17% in mountain stream. Similarly, Phytopythium accounted for ca. 13%, 7%, and 13% of the isolates in these habitats, respectively. Saprolegniales was isolated at a proportion of ca. 10% in reservoir, 22% in lowland river, and 22% in mountain stream. The species richness of Saprolegnia was higher in mountain stream (seven taxa) and lowland river (eight taxa), whereas its presence was relatively limited in reservoirs with only four taxa. Lastly, Peronosporales was detected at proportions of a few percent across all freshwater ecosystem types (ca. 1% in reservoir, 4% in lowland river, and 2% in mountain stream).
Taxonomy
-
Phytopythium minutum B. Nam & Y.J. Choi, sp. nov. Fig. 7
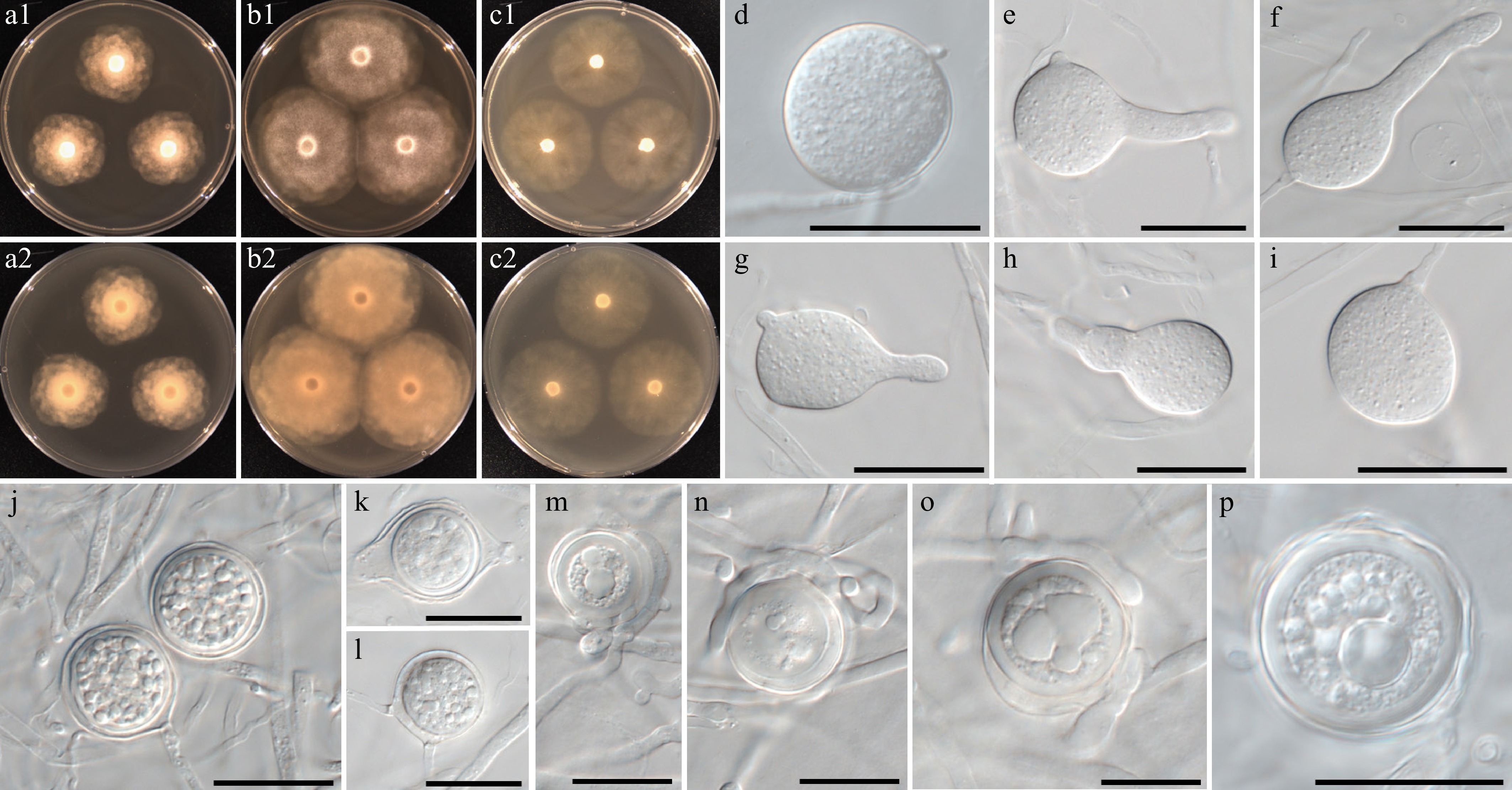
Figure 7.
Colony patterns and morphology of Phytopythium minutum W1152. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(p) Microscopic structures observed under a microscope: (d)–(i) (sub-)globose to (ob-)pyriform sporangia with or without discharge tube, (j)–(p) globose oogonia with one or multiple antheridia per oogonium, sometimes encircling the oogonium (scale bars: 20 μm).
MycoBank number: MB861107; Facesoffungi number: FoF18859
Etymology – Name refers to the small size of sporangia and oogonia (minutus Latin = very small).
Main hyphae up to 5 µm wide. Sporangia were globose to sub-globose, with an average size of 18 ± 3 μm in diameter, with a total range of 13–23 μm, rarely pyriform to obpyriform or ovoid, 16–20 × 24–27 μm (average 18 ± 2 × 26 ± 1 μm). Sporangia were terminal or intercalary. Hyphal swellings and chlamydospores were not observed. Oogonia were globose to sub-globose or limoniform and had smooth walls. Oogonial diameter averaged 21.5 ± 4 µm with an overall range of 15.5–29.3 µm. Oospores were aplerotic or plerotic, globose, ranging in size from 13–26 µm (av. 18.5 ± 3 µm) in diameter with a wall 0.5–3 (av. 1.8) µm thick. Antheridia were monoclinous or diclinous, paragynous, and showing undefined shapes or encircled the oogonium.
Culture characteristics – Colonies showed a chrysanthemum pattern on PDA, a faint chrysanthemum on V8A, and faintly striate on CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 8, 12, and 9.5 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 4, 30, and 35 °C, respectively.
Material examined – Korea, Gyeongsangbuk-do, Sangju, isolated from leaf litter in a mountain stream, Apr. 2020, B. Nam & Y.J. Choi, W1152 (holotype NNIBRFG31634, ex-type culture NNIBRF31694); Jeollabuk-do, Namwon, isolated from leaf litter in a river, Sept. 2018, ditto, W676. Jeollanam-do, Muan, isolated from leaf litter in a river, June 2021, ditto, W1998, W1999.
GenBank numbers – W1152: ITS = PP812166, cox1 = PP798551, cox2 = PP834383; W676: ITS = PP812163, cox1 = PP798444, cox2 = PP834380; W1998: ITS = PP812183, cox1 = PP798888, cox2 = PP834397; W1999: ITS = PP812184, cox1 = PP798889, cox2 = PP834398.
Notes – Morphological and physiological characters of Phytopythium minutum are listed in Table 2, for comparison, with those of already known and phylogenetically close species, Pp. aichiense, Pp. citrinum, Pp. delawarense, and Pp. litorale. Sporangia and oogonia of Pp. minutum are on average smaller than those of the related species. Optimum growth temperature is higher (30 °C) than in other species (25–28 °C), like Pp. litorale.
Table 2. Comparison of Phytopythium minutum with morphologically and phylogenetically related species (n.a. = not available).
Character Phytopythium minutum Phytopythium citrinum Phytopythium delawarense Phytopythium aichiense Phytopythium litorale Colony Chrysanthemum pattern on PDA, faintly chrysanthemum on V8A, and faintly striate on CMA Submerged and mixed chrysanthemum-rosette pattern on PCA Radiate pattern on PCA Stellate pattern on V8A Radiate to chrysanthemum pattern on PCA and V8A Growth rate 8 mm per day at 25 °C
on PDA
12 mm per day at 25 °C
on V8A
9.5 mm per day at 25 °C
on CMA9 mm per day at 25 °C on CMA
11 mm per day at 25 °C
on PCA10 mm per day at
25 °C on CMA11.6 mm per day at
25 °C on CMA13.3 mm per day at 25 °C on CMA
12.6 mm per day at 25 °C on PCAProliferation Internal extended Internal extended Internal nested and internal extended Internal nested and internal extended Internal nested and internal extended Hyphal swellings Absent Absent Present Absent Present Sporangia (Sub-)globose, 13–23 (av. 18 ± 3) μm diameter, or ovoid to (ob-)pyriform, 16–20 × 24–27 (av. 18 ± 2 × 26 ± 1) μm, terminal or intercalary (Sub-)globose or pyriform to lemoniform, terminal
or intercalary, 15–35 (av. 24.2) μm diameterGlobose to slightly elongate, limonform or (ob-)void, terminal or intercalary, 25–40 (av. 32.5) μm diameter Sub-globose, ovoid or limonoform, 25–39 × 19–27 (31.4 × 24.9) μm (av. 31.4 μm) diameter (Sub-)globose, broad ovoid or obpyriform, terminal or intercalary, 22.8–32.2 × 20.8–29.1 (av. 28.5 × 26.7) μm Oogonia Smooth, globose to sub-globose or limoniform, 15.5–29.3 (av. 21.5 ± 4) μm diameter Smooth, globose, 20–36 (av. 27.6) μm diameter Smooth, globose, 21–34 (av. 27.2) μm diameter Smooth, globose, 29–40 (av. 34.3) μm diameter Absent Antheridia Monoclinous or diclinous, showing undefined shapes or encircling the oogonium Strictly hypogynous 1–2 per oogonium, diclinous, broad lengthwise contact 1–2 per oogonium, mostly diclinous, and rarely monoclinous, crook-necked or filamentous inflated Absent Oospores Aplerotic or plerotic, globose, 13–26 (av. 18.5 ± 3) μm diameter Plerotic, 18–34 (av. 24.9) μm diameter Aplerotic, 21–26 (av. 24.2) μm diameter Aplerotic, av. 24.2 μm diameter Absent Cardinal growth temperature (minimum/optimum/
maximum temperature)4/30/35 °C n.a. 10/28/34 °C 4/28/34 °C 4/30/35 °C Ref. This study Paul[151] Broders et al.[152] Baten et al.[93] Nechwatal & Mendgen[146] Phytopythium lacustre B. Nam & Y.J. Choi, sp. nov. Fig. 8
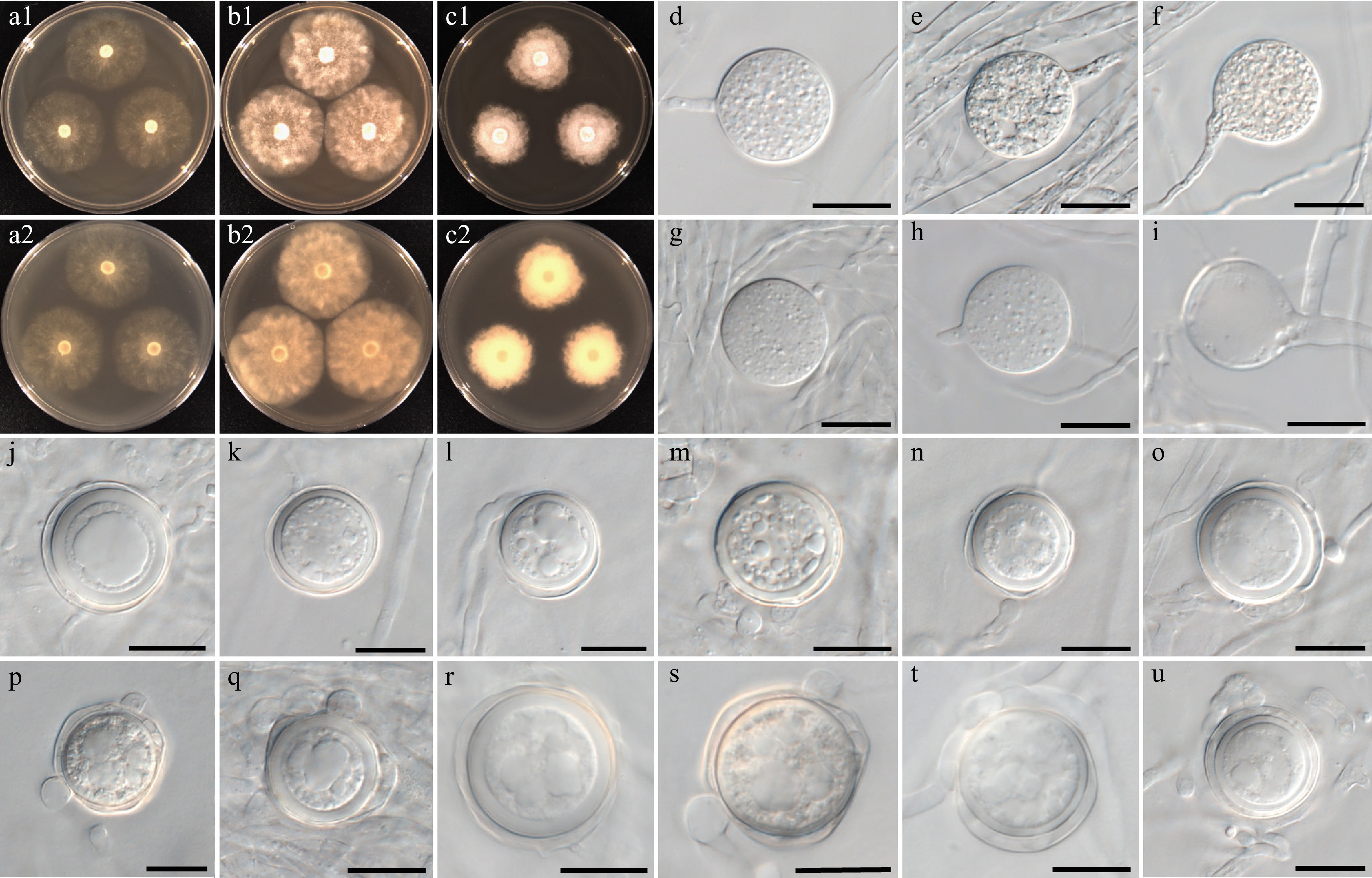
Figure 8.
Colony patterns and morphology of Phytopythium lacustre W1435. Cultural characteristics of (a) PDA, (b) V8A, and (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(u) Microscopic structures observed under a microscope: (d)–(i) (sub-)globose sporangia with (h) growing release tube, and (i) empty sporangium, (j)–(u) (sub-)globose oogonia with one to two club-shaped or filamentous inflated antheridia per oogonium (scale bars: 20 μm).
MycoBank number: MB861116; Facesoffungi number: FoF18860
Etymology – Name refers to the isolation of the type strain from a lake (lacustis Latin = lake).
Main hyphae up to 6 µm wide. Sporangia were globose to sub-globose. The size of the sporangia averaged 31 ± 4 μm in diameter with a total range of 24–38 μm. Sporangia were terminal. Hyphal swellings and chlamydospores were not observed. Oogonia were globose to subglobose and had smooth walls. Oogonial diameter averaged 32.5 ± 2 µm with an overall range of 29–36 µm. Oospores were aplerotic or plerotic, globose, ranging in size from 23–35 µm (av. 29.5 ± 4 µm) in diameter with a wall 1–3 (av. 1.7) µm thick. Antheridia were monoclinous or diclinous, paragynous, and produced one to two per oogonium. Antheridial cells were club-shaped or inflated clavate.
Culture characteristics – Colonies showed radiate to faint chrysanthemum patterns on PDA, V8A, and CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 12, 14, and 10 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 10, 30, and 35 °C, respectively.
Material examined – Korea, Chungcheongnam-do, Boryeong, isolated from leaf litter in an artificial reservoir, Sept. 2020, B. Nam & Y.J. Choi, W1435 (holotype NNIBRFG31641, ex-type culture NNIBRF31701).
GenBank numbers – W1435: ITS = PP812169, cox1 = PP798688, cox2 = PP834405
Notes – Morphological and physiological characters of Phytopythium lacustre are compared with those of already known and phylogenetically close species, Pp. boreale, Pp. iriomotense, Pp. mercuriale, and Pp. oedochilum (Table 3). The growth rate at 25 °C on the CMA of Pp. lacustre and Pp. mercuriale is lower (9–10 mm) than in other closely related species (20–28.5 mm). Pp. lacustre was obtained during the warm season, which may be related to its minimum (10 °C), optimum (30 °C), and maximum (35 °C) growth temperatures. Interestingly, the related species Pp. iriomotense and Pp. oedochilum exhibit the same cardinal temperatures. This species is currently represented by a single isolate, which limits assessment of intraspecific variation. However, the species is clearly distinguished from its closest relatives based on diagnostic morphological characters and robust multilocus phylogenetic evidence. Additional isolates may further refine the characterization of this species in future studies.
Table 3. Comparison of Phytopythium lacustre with morphologically and phylogenetically related species (n.a. = not available).
Character Phytopythium lacustre Phytopythium iriomotense Phytopythium mercuriale Phytopythium oedochilum Phytopythium boreale Colony Radiate to faint chrysanthemum patterns on PDA, V8A and CMA Chrysanthemum pattern on V8A Tightly chrysanthemum on PDA, rosaceous on MEA, slightly stellate on CMA Submerged with some aerial mycelium on CMA, without a special pattern on PCA n.a. Growth rate 12 mm per day at 25 °C
on PDA
14 mm per day at 25 °C
on V8A
10 mm per day at 25 °C
on CMA24.3 mm per day at 25 °C on CMA 9 mm per day at
25 °C on CMA20 mm per day at 25 °C on CMA 20 mm per day at
25 °C on CMAProliferation Internal extended Internal or internal nested Internal or internal nested External, internal and internal nested Internal Hyphal swellings Absent Present Present Absent Present Sporangia Globose to sub-globose, terminal, 24–38
(av. 31 ± 4) μm
diameterGlobose to sub-globose, papillated and terminal
av. 32.7 μmSub-globose to obovoid, papillated and terminal
av. 22 μmSub-globose, lemoniform, obovoid
or ovoid, papillated
av. 32.8 μmAbsent Oogonia Smooth, (sub-)globose, terminal or intercalary, 29–36 (av. 32.5 ± 2) μm diameter Smooth, globose av.
27.3 μm diameter, oogonia with double oospores presentSmooth, globose, av. 29.8 μm diameter Smooth and globose av. 32.8 μm diameter Smooth, globose, av. 32.8 μm diameter Antheridia Monoclinous or diclinous, 1−2 per oogonium, club-shaped or inflated clavate Diclinous, 1−2 per oogonium, crook-necked or occasionally amphigynous antheridial cells are present Mostly diclinous, multiple knotted around the oogonia Diclinous, occasionally monoclinous, 1−2(4) per oogonium, cells were large, curved, and broadly laterally attached to the oogonium Monoclinous, rarely diclinous, 1−2 per oogonium, antheridial cells are amphigynous
or sessilOospores Aplerotic or plerotic, globose, 23–35(av. 29.5 ± 4) μm diameter Aplerotic or plerotic av. 24.4 μm diameter Absent Aplerotic av. 30.3 μm diameter Plerotic av. 22.2 μm diameter Cardinal growth temperature (minimum/optimum/
maximum temperature)10/30/35 °C 10/30/35 °C 5/25–30/43 °C 10/30/35 °C 4/25–30/43 °C Ref. This study Baten et al.[93] Belbahri et al.[153] Van der Plaats-Niterink[150] Duan[154] Pythium aculeatum B. Nam & Y.J. Choi, sp. nov. Fig. 9
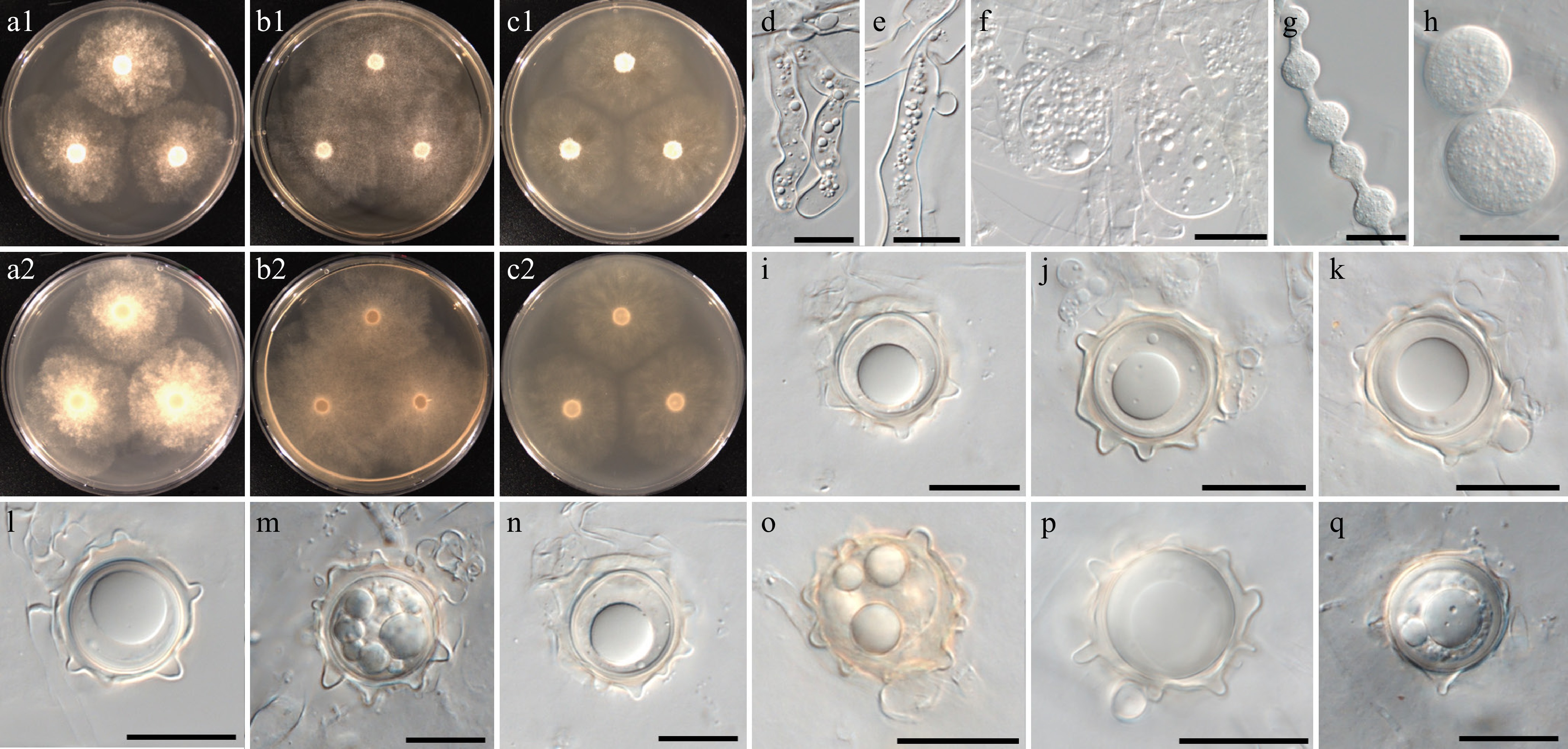
Figure 9.
Colony patterns and morphology of Pythium aculeatum W1856. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(q) Microscopic structures observed under a microscope, (d)–(f) inflated filamentous sporangia, (g), (h) hyphal swellings, (i)–(q) globose oogonia with ornamented wall and globose or clavate antheridia (scale bars: d–h = 10 μm, i–q = 20 μm).
MycoBank number: MB861117; Facesoffungi number: FoF18892
Etymology – Name refers to the ornamented walls of the oogonia (aculeatus Latin = spiny).
Main hyphae up to 6 µm wide. Sporangia were filamentous and inflated. Hyphal swellings were globose to subglobose, limoniform or catenulate, terminal or intercalary. Oogonia were globose and had ornamented walls with blunt projections. Oogonial diameter averaged 21 ± 2 µm with an overall range of 19–27 µm. Oospores were mostly aplerotic or plerotic, globose, often yellowish, ranging in size from 16–24 µm (av. 20 ± 2 µm) in diameter with a wall 0.3–2.5 (av. 1.5) µm thick. Antheridia were produced one or two per oogonium, monoclinous or diclinous, paragynous. Antheridial cells were globose or clavate.
Culture characteristics – Colonies showed faint chrysanthemum patterns on PDA and V8A, and a radiate pattern on CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 13.5, 15.5, and 13.5 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 20, and 30 °C, respectively.
Material examined – Korea, Jeju-do, Jeju, isolated from water in a mountain stream, Apr. 2021, B. Nam & Y.J. Choi, W1856 (holotype NNIBRFG35173, ex-type culture NNIBRF35147); Jeju-do, Jeju, isolated from water in a mountain stream, Apr. 2021, ditto, W1859.
GenBank numbers – W1856: ITS = PP812174, cox1 = PP798833, cox2 = PP834389; W1859: ITS = PP812175, cox1 = PP798835, cox2 = PP834390.
Notes – Morphological and physiological characters of Pythium aculeatum are listed in Table 4 for comparison with those of the phylogenetically close new taxon Py. intricatum and already known species, namely Py. flevoense, Py. inflatun, Py. pyrilobum, and Py. rishiriense. Oogonial walls of Py. aculeatum are ornamented with blunt projections, while those of related species are smooth. The optimum growth temperature of the related species is between 25 and 35 °C, but Py. aculeatum grows optimally at 20 °C, and the maximum temperature is 30 °C. ITS and cox2 sequences of Py. aculeatum were identical to those of isolate ZSF0011 reported by Uzuhashi et al.[148], but its taxonomic characteristics are still undescribed.
Table 4. Comparison of Pythium aculeatum with morphologically and phylogenetically related species (n.a. = not available).
Character Pythium aculeatum Pythium flevoense Pythium rishiriense Pythium pyrilobum Pythium inflatum Colony Faint chrysanthemum pattern on PDA and V8, radiate pattern on CMA Chrysanthemum pattern on PDA and CMA Rosette petaloid on PDA, vaguely petaloid on V8A Radiate pattern without a special pattern on PCA Radiate pattern on PCA, slightly radiate pattern on CMA Growth rate 13.5 mm per day at 25 °C on PDA
15.5 mm per day at 25 °C on V8A
13.5 mm per day at 25 °C on CMA10 mm per day at 25 °C
on CMA16 mm per day at
25 °C on PCA22 mm per day at
25 °C on PCA20 mm per day at 25 °C on CMA Hyphal swellings Present. Hyphal bodies, terminal or intercalary, (sub-)globose, limoniform or catenulate Absent Absent n.a. n.a. Sporangia Filamentous, inflated, irregularly swollen Filamentous, not-inflated Filamentous, inflated Filamentous, inflated, irregularly swollen Filamentous, inflated, forming irregular or globose outgrowths Oogonia Ornamented with blunt projections, globose, terminal, 19–27 (av. 21 ± 2) μm diameter Smooth, mostly terminal on short side branches of feather-like hyphae, 17–30 (av. 19) µm diameter Smooth, globose, terminal or intercalary, sometimes in a chain, av. 24.3 μm diameter Smooth, globose or pyriform, terminal or intercalary, 23–33 (av. 27) μm diameter Smooth, globose, terminal or intercalary, 19–27 (av. 22.5) μm diameter Antheridia Monoclinous or diclinous, 1−2 per oogonium, antheridial cells globose or clavate Diclinous, one to several per oogonium, antheridial stalks mostly bifurcate or differently branched near the oogonium 1−2(–5) per oogonium, monoclinous or diclinous, terminal on branched stalk, rarely intercalary, clavate or
crook-necked, hypogynousMonoclinous or diclinous, mostly two or three cells on one long stalk which encircles the oogonium, mostly crook-necked 1−2 per oogonium, diclinous Oospores Aplerotic or plerotic, globose, often yellowish, 16–24 (av. 20 ± 2) µm diameter Aplerotic, occasionally nearly plerotic, 14–24
(av. 17.7) µm diameterMostly plerotic, av. 21.8 μm diameter Aplerotic or plerotic, globose, yellowish, 20–32 (av. 23.5) µm diameter Plerotic, 18–25 (av. 21.5) µm diameter Cardinal growth temperature (minimum/optimum/
maximum temperature)5/20/30 °C 5/25/35 °C 5/35/42 °C 4/25/37 °C 5/30/37 °C Ref. This study Van der Plaats-Niterink[155] Rahman et al.[82] Vaartaja[156] Van der Plaats-Niterink[150] Pythium aquaticum B. Nam & Y.J. Choi, sp. nov. Fig. 10
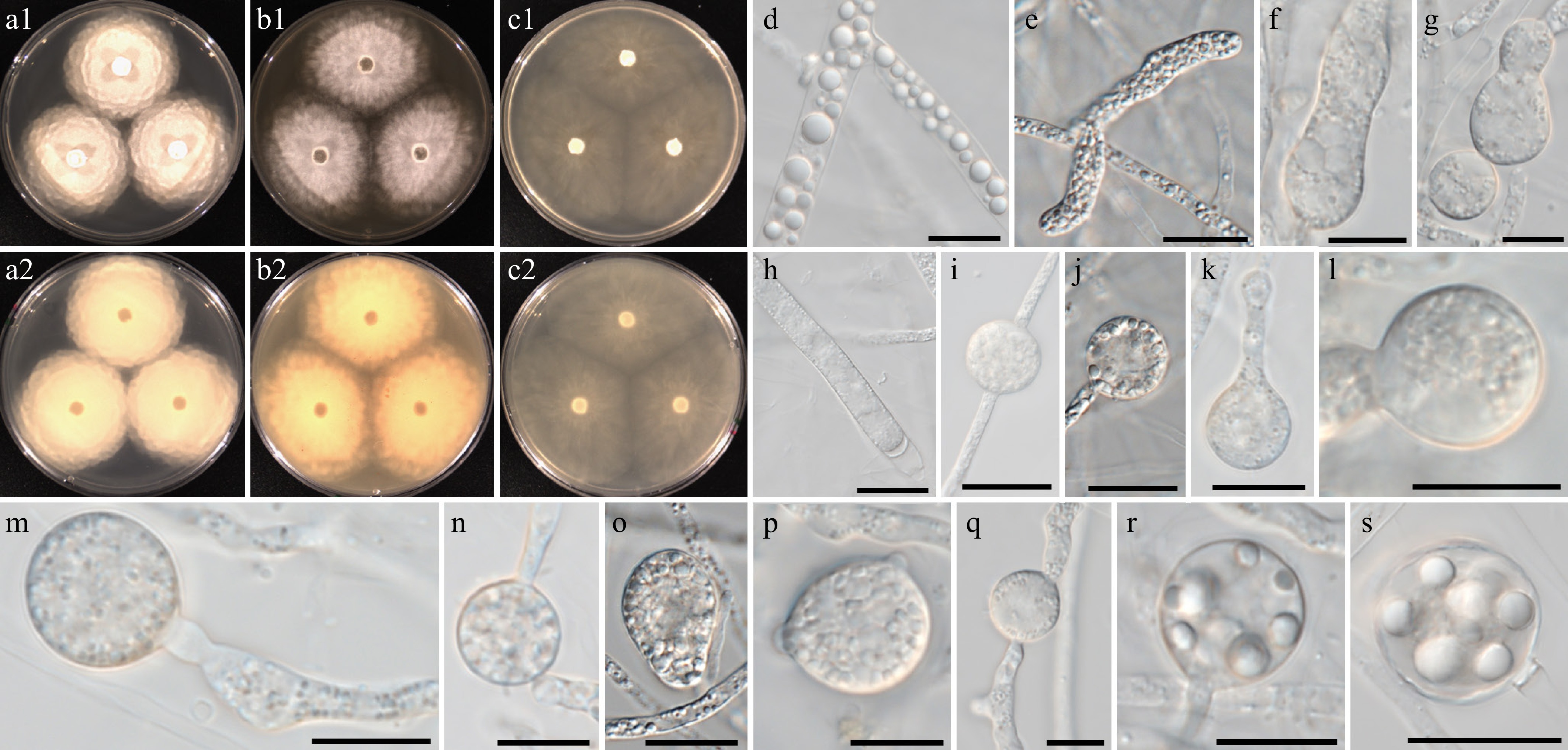
Figure 10.
Colony patterns and morphology of Pythium aquaticum W2020. Cultural characteristics on (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(s) Microscopic structures observed under a microscope, (d), (r), (s) lipid globules inside the hyphae, (e) filamentous, inflated sporangia, (f)–(q) cylindrical, clavate, (sub-)globose, ovoid or peanut-shaped hyphal swellings (scale bars: 10 μm).
MycoBank number: MB861131; Facesoffungi number: FoF18861
Etymology – Name refers to the wide distribution in freshwater ecosystems (aquaticus Latin = living in water).
Main hyphae up to 7 µm wide. Sporangia were filamentous and slightly inflated. Cylindrical, clavate, (sub-)globose, ovoid, or peanut-shaped hyphal swellings were produced terminally or intercalary. Chlamydospores and sexual structures were not observed.
Culture characteristics – Colonies showed a rosaceous pattern on PDA and radiate patterns on V8A and CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 12.5, 9.5, and 10.5 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 30, and 40 °C, respectively.
Material examined – Korea, Gyeongsangbuk-do, Mungyeong, isolated from water in a river, June 2021, B. Nam & Y.J. Choi, W2020 (holotype NNIBRFG48680, ex-type culture NNIBRF48682); Chungcheongbuk-do, Cheongwon, isolated from soil sediments in a river, June 2018, ditto, W664; Gyeongsangbuk-do, Mungyeong, isolated from water in a river, June 2021, ditto, W2021. Gyeongsangbuk-do, Mungyeong, isolated from soil sediments in a river, June 2021, ditto, W2026; Gyeongsangbuk-do, Mungyeong, isolated from marsh snail in a river, June 2021; ditto, W2029.
GenBank numbers – W2020: ITS = PP812192, cox1 = PP798909, cox2 = PP842635; W664: ITS = PP812162, cox1 = PP798442, cox2 = PP842631; W2021: ITS = PP812193, cox1 = PP798910, cox2 = PP842636; W2026: ITS = PP812194, cox1 = PP798915, cox2 = PP842637; W2029: ITS = PP812195, cox1 = PP798918, cox2 = PP842638.
Notes – Py. aquaticum has higher optimum (30 °C) and maximum (40 °C) growth temperatures, like related species, including the new taxon Py. fluminis (Table 5). Py. aquaticum produces diverse shapes of hyphal swellings: (sub-)globose, ovoid, and peanut-shaped. The ITS sequence of Py. aquaticum is identical to unclassified Pythium sp. AL-2010 (NCBI taxonomy ID 907948; specimen-voucher P8207), but the cox1 and cox2 sequences displayed similarities of 99%.
Table 5. Comparison of Pythium aquaticum and Py. fluminis with morphologically and phylogenetically related species (n.a. = not available).
Character Pythium aquaticum Pythium fluminis Pythium catenulatum Pythium torulosum Pythium rhizo-oryzae Pythium rishiriense Colony Rosaceous pattern on PDA, radiate pattern on V8 and CMA Rosaceous pattern on PDA, chrysanthemum pattern on V8A, irregular pattern on CMA Narrow chrysanthemum pattern on PCA, rosette pattern on CMA Rosette pattern on PCA Narrow chrysanthemum pattern on PCA, rosette pattern on CMA Rosette petaloid on PDA, vaguely petaloid on V8A Growth rate 12.5 mm per day at 25 °C on PDA
9.5 mm per day at
25 °C on V8A
10.5 mm per day at 25 °C on CMA12.5 mm per day at
25 °C on PDA
14.5 mm per day at
25 °C on V8A
10.5 mm per day at
25 °C on CMA14 mm per day at
25 °C on PCA15 mm per day at
25 °C on PCA12 mm per day at 25 °C on PCA 16 mm per day at
25 °C on PCAHyphal swellings Present. Hyphal bodies, terminal or intercalary, cylindrical, clavate, (sub-)globose, ovoid or peanut-shaped Present. Hyphal bodies, terminal or intercalary, clavate, (sub-)globose, catenulate, ovoid or peanut-shaped Present Absent Present. Hyphal bodies, catenulate and terminal Absent Sporangia Filamentous, slightly inflated Filamentous, inflated Filamentous, inflated, irregularly swollen, branched parts of the mycelium Filamentous, inflated Not observed Filamentous, inflated Oogonia Not observed Smooth, globose, terminal or intercalary, 11–22 (av. 15 ± 3) μm diameter Smooth, globose, terminal or intercalary, mostly heterothallic,
av. 22.8 μm diameterSmooth, terminal or intercalary, 11–22
(av. 15) μm diameterSmooth, globose, terminal or intercalary, av.
19.7 μm diameterSmooth, globose, terminal or intercalary, sometimes in a chain, av. 24.3 μm diameter Antheridia Not observed Monoclinous, antheridial cells clavate, bell-shaped,
or with wide contact with oogonium5–(–12) per oogonium, diclinous, occasionally monoclinous in homothallic isolates, clavate or crook-necked, stalk branched 1–2(–3) per oogonium, monoclinous or occasionally diclinous, borne on rarely branched antheridial stalks Wrapping around, making a knot 1–2(–5) per oogonium, monoclinous or diclinous, terminal on branched stalk, rarely intercalary, clavate or
crook-necked, hypogynousOospores Not observed Aplerotic or plerotic, globose, 9.5–20 (av.
14 ± 3) μm diameterMostly plerotic, occasionally aplerotic Plerotic, 10–21 μm diameter Plerotic and aplerotic, 14–
20 μm diameterMostly plerotic, av. 21.8 μm diameter Cardinal growth temperature (minimum/
optimum/
maximum temperature)5/30/40 °C 5/30/40 °C 10/30–35/40 °C 5/25–30/35 °C n.a. 5/35/42 °C Ref. This study This study Van der Plaats-Niterink[150] Van der Plaats-Niterink[150] Bala[157] Rahman et al.[82] Pythium chlamydosporum B. Nam & Y.J. Choi, sp. nov. Fig. 11
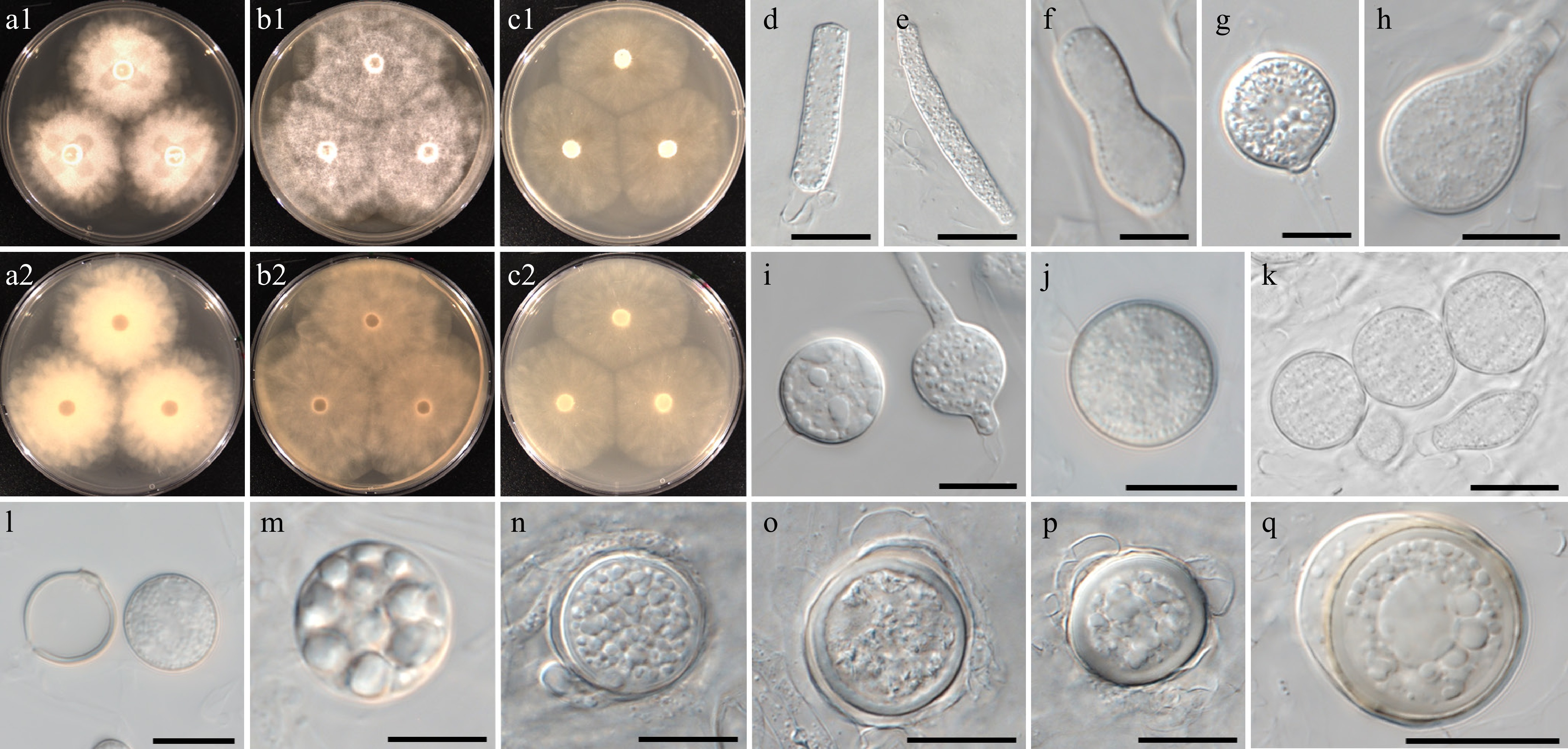
Figure 11.
Colony patterns and morphology of Pythium chlamydosporum W2002. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(q) Microscopic structures observed under a microscope: (d), (e) filamentous slightly inflated structures, (f)–(i) cylindrical, clavate, (sub-)globose, ovoid or peanut-shaped hyphal swellings, (j), (k) chlamydospores, (l) empty and mature chlamydospore, (m)–(q) globose oogonia with antheridia (scale bars: 10 μm).
MycoBank number: MB861143; Facesoffungi number: FoF18862
Etymology – Name refers to the frequent formation of chlamydospores.
Main hyphae up to 7 µm wide. Sporangia and zoospores not observed. Hyphal inflated structures filamentous slightly inflated. Hyphal swellings and chlamydospores were observed. Hyphal swellings were globose, rarely ovoid or peanut-shaped, and produced terminally or intercalary. Chlamydospores formed in V8A agar media, abundant at 25 °C. Chlamydospores were globose, terminal, and intercalary, with an average diameter of 15 ± 2 µm. Oogonia had smooth walls, were globose, and produced terminal or intercalary. Oogonial diameter averaged 18 ± 4 µm with an overall range of 10–24 µm. Oospores were plerotic, globose, ranging in size from 9–22 µm (av. 16 ± 3 µm) in diameter with a wall 0.5–2.5 (av. 1.6) µm thick. Antheridia were produced one or two per oogonium, monoclinous or diclinous, paragynous, globose, or encircling an oogonium.
Culture characteristics – Colonies showed a radial pattern on PDA, V8A, and CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 16, 18, and 16 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 25, and 30 °C, respectively.
Material examined – Korea, Jeollanam-do, Jindo, isolated from soil sediments in a reservoir, June 2021, B. Nam & Y.J. Choi, W2002 (holotype NNIBRFG35175, ex-type culture NNIBRFG35149); Jeollabuk-do, Jeonju, isolated from leaf litter in a reservoir, Sept. 2020, ditto, W1470; Jeollanam-do, Muan, isolated from soil sediments in a river, June 2021, ditto, W1991.
GenBank numbers – W2002: ITS = PP812185, cox1 = PP798892, cox2 = PP842634; W1470: ITS = PP812170, cox1 = PP798704, cox2 = PP842632; W1991: ITS = PP812181, cox1 = PP798883, cox2 = PP842633.
Notes – Morphological and physiological characters of Pythium chlamydosporum are listed in Table 6 for comparison with those of phylogenetically related species, i.e., Py. apleroticum, Py. aquatile, Py. brachiatum, Py. diclinum and the new species Py. vigens. Py. chlamydosporum produces chlamydospores abundantly at 25 °C. Hyphal swellings were formed in diverse shapes. The ITS and cox1 sequences of Py. chlamydosporum are identical to an undescribed Pythium species, CLE-2015d (NCBI taxonomy ID 1695211). Another undescribed Pythium strain, W18-138 (NCBI taxonomy ID 1862977), formed a cluster with Py. chlamydosporum in the phylogenetic tree. This strain was also isolated from freshwater ecosystems of Cocalico Creek in Pennsylvania, USA, according to Professor Blair (pers. comm.).
Table 6. Comparison of Pythium chlamydosporum and Py. vigens with morphologically and phylogenetically related species (n.a. = not available)
Character Pythium chlamydosporum Pythium vigens Pythium aquatile Pythium apleroticum Pythium brachiatum Pythium oopapillum Pythium diclinum Colony Radiate pattern on PDA, V8A, and CMA Faint striate to radiate pattern on PDA, V8A, and CMA Rosette pattern on PCA and CMA n.a. Radiate pattern Vague chrysanthemum pattern on PCA Radiate pattern on CMA and PCA Growth rate 16 mm per day at 25 °C on PDA
18 mm per day at 25 °C on V8A
16 mm per day at 25 °C on CMA22 mm per day at 25 °C
on PDA
28.5 mm per day at 25 °C on V8A
20 mm per day at 25 °C
on CMA17 mm per day at
25 °C on PCA19 mm per day at
25 °C on PCA14 mm per day at 25 °C on PCA 17 mm per day at
25 °C on PCA19 mm per day at
25 °C on PCAHyphal swellings Present. Hyphal bodies, terminal or intercalary, (sub-)globose, rarely ovoid or peanut shaped Present. Hyphal bodies, terminal or intercalary, (sub-)globose, ovoid or peanut-shaped, rarely catenulate Absent Absent Filamentous slightly inflated n.a. n.a. Sporangia Not observed Not observed Filamentous or slightly inflated, forming rectangular, dendroid structures Filamentous, not-inflated Not observed Filamentous, inflated, lobate elements Filamentous, not-inflated Oogonia Smooth, globose, terminal or intercalary, 10–24 (av. 18 ± 4) μm diameter Smooth, globose, terminal or intercalary, 21–26 (av. 23 ± 1) μm diameter Oogonia globose, smooth, terminal or less often intercalary, but frequently clustering in small groups, 19–23 (av. 21 µm) diameter Smooth, globose, terminal or intercalary, 11–20 μm Intercalary or terminal, sometimes in chain; smooth- walled or occasionally with a projection, 13.3–34.4 (av. 22.7) Globose, sub-terminal or intercalary, av.
21.5 μmSmooth, spherical or ovoid, mostly terminal or subterminal, occasionally intercalary, 18–23 μm (av. 20.5) Antheridia Monoclinous or diclinous, 1–2 per oogonium, antheridial cells globose or encircling an oogonium Lacking One, rarely two per oogonium, monoclinous, originating from the oogonial stalk at 2–10 µm or more below the oogonium, or from the parent hypha Diclinous, 1–2 per oogonium. Diclinous and monoclinous; arm-like shaped on unbranched stalks; 1–2 per oogonium Rarely hypogynous, antheridial stalks branched Diclinous, 1–2 per oogonium Oospores Aplerotic or plerotic, globose, 9–22 (av. 16 ± 3) µm diameter Aplerotic, globose, 13–16 (av. 15) µm diameter Aplerotic, 16–19 (av. 17.9) µm diameter Aplerotic, 9–17 µm diameter Plerotic, occasionally aplerotic, 9.4–27.2
(av. 16.4) diameterav. 16.4 μm diameter av. 17.5 μm diameter Cardinal growth temperature (minimum/optimum/
maximum temperature)5/25/30 °C 5/25/35 °C 5/25/35 °C 5/30/40 °C 4/25/30 °C n.a./30/30 °C 5/30/40 °C Ref. This study This study Höhnk[158] Ito & Tokunaga[159] Uzuhashi et al.[99] Bala et al.[160] Ito & Tokunaga[159] Pythium intricatum B. Nam & Y.J. Choi, sp. nov. Fig. 12
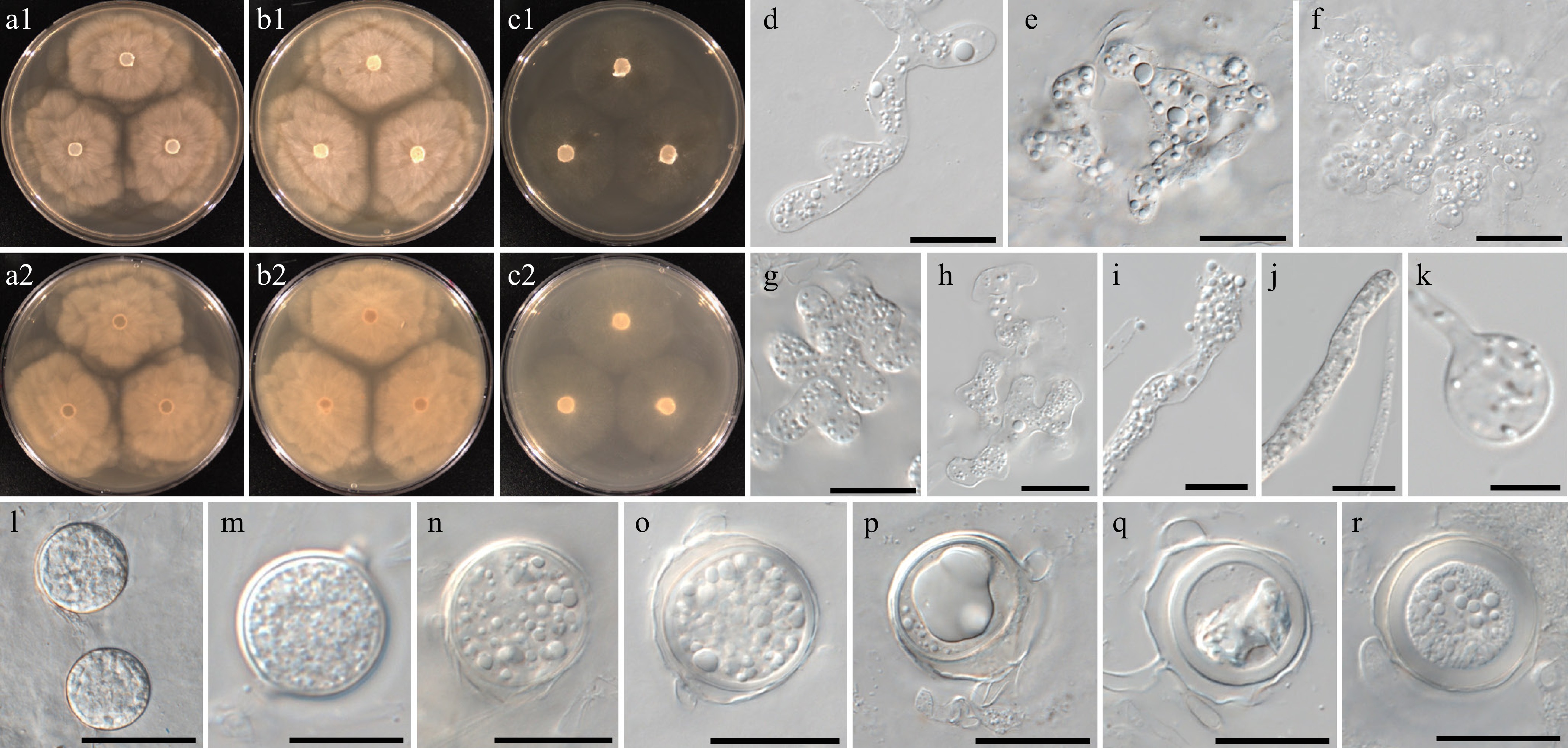
Figure 12.
Colony patterns and morphology of Pythium intricatum W2004. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(r) Microscopic structures observed under a microscope: (d) filamentous inflated sporangia, (e)–(h) lobulated sporangia forming toruloid complexes or dendroid structures, (i)–(k) filamentous sporangia and swelling, (l), (m) chlamydospores, (n)–(r) globose oogonia with antheridial cells (scale bars: d–k = 10 μm, l–r = 20 μm).
MycoBank number: MB861145; Facesoffungi number: FoF18863
Etymology – Name refers to the shape of sporangia; strongly inflated, irregular toruloid complexes or dendroid structures (intricatus Latin = complex).
Main hyphae up to 7 µm wide. Sporangia were filamentous, strongly inflated, and formed toruloid complexes or dendroid structures. Terminal hyphal swellings were abundantly formed. Globose chlamydospores were rarely observed. Oogonia had smooth walls, were globose with oogonial diameter averaging 22 ± 3 µm and an overall range of 17–27 µm. Oospores were aplerotic or plerotic, globose, ranging in size from 15–26 µm (av. 20.5 ± 3 µm) in diameter with a wall 1–3 (av. 2.2) µm thick. Antheridia were produced one or two per oogonium, monoclinous or diclinous, paragynous. Antheridia were globose, clavate, or encircling an oogonium.
Culture characteristics – Colonies showed chrysanthemum patterns on PDA and V8A, and a very faint striate pattern on CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 10.5, 10.5, and 8.5 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 25 and 30 °C, respectively.
Material examined – Korea, Jeollanam-do, Jindo, isolated from soil sediments in a reservoir, June 2021, B. Nam & Y.J. Choi, W2004 (holotype NNIBRFG35176, ex-type culture NNIBRFG35150); Jeollabuk-do, Wanju, isolated from leaf litter in a stream, May 2018, ditto, W604; Jeollabuk-do, Imsil, isolated from water in a stream, Sept. 2018, ditto, W693 & W695; Jeollanam-do, Damyang, isolated from the algae in a stream, Jan. 2021, ditto, W1522, W1523 and W1524; Gyeongsangbuk-do, Yeongju, isolated from leaf litter in a stream, May 2021, ditto, W1952; Jeollanam-do, Jindo, isolated from leaf litter in a reservoir, June 2021, ditto, W2005.
GenBank numbers – W2004: ITS = PP812186, cox1 = PP798894, cox2 = PP834399; W604: ITS = PP812161, cox1 = PP798437, cox2 = PP834379; W693: ITS = PP812164, cox1 = PP798451, cox2 = PP834381; W695: ITS = PP812165, cox1 = PP798452, cox2 = PP834382; W1522: ITS = PP812171, cox1 = PP798741, cox2 = PP834386; W1523: ITS = PP812172, cox1 = PP798742, cox2 = PP834387; W1524: ITS = PP812173, cox1 = PP798743, cox2 = PP834388; W1952: ITS = PP812176, cox1 = PP798858, cox2 = PP834391; W2005: ITS = PP812187, cox1 = PP798895, cox2 = PP834400.
Notes – Morphological and physiological characters of Pythium intricatum are listed in Table 7 for comparison with those of phylogenetically related species. Filamentous sporangia of Py. intricatum are strongly inflated or lobulated. They often produce toruloid complexes or dendroid structures. The hyphal swellings of Py. intricatum were formed only on the terminal portion of hyphae. The ITS, cox1, and cox2 sequences of W2004 are close to the sequences of unclassified Pythium sp. AL-2010 (NCBI taxonomy ID 907948; specimen-voucher P8201, P8204) and MNS-2013 (NCBI taxonomy ID 1383453; strain ABDN_66 and isolate VN851a) with 96%–98% similarity in the BLASTn search.
Table 7. Comparison of Py. intricatum with morphologically and phylogenetically related species (n.a. = not available).
Character Pythium intricatum Pythium flevoense Pythium rishiriense Pythium inflatum Pythium aquatile Colony Chrysanthemum patterns on PDA and V8A, very faint striate pattern on CMA Chrysanthemum pattern on PDA and CMA Rosette petaloid on PDA, vaguely petaloid on V8A Radiate pattern on PCA, slightly radiate pattern on CMA Rosette pattern on PCA and CMA Growth rate 10.5 mm per day at 25 °C on PDA
10.5 mm per day at 25 °C on V8A
8.5 mm per day at 25 °C
on CMA10 mm per day at 25 °C
on CMA16 mm per day at 25 °C on PCA 20 mm per day at 25 °C on CMA 17 mm per day at 25 °C on PCA Hyphal swellings Present. Hyphal bodies, terminal, (sub-)globose Absent Absent n.a. Absent Sporangia Filamentous, strongly inflated, lobulated, forming toruloid complexes or dendroid structures. Filamentous, not-inflated Filamentous, inflated Filamentous, inflated, forming irregular or globose outgrowths Filamentous or slightly inflated, forming rectangular, dendroid structures Oogonia Smooth, globose, terminal or intercalary, 17–27 (av. 22 ± 3) μm diameter Smooth, mostly terminal on short side branches of feather-like hyphae, 17–30 (av. 19) µm diameter Smooth, globose, terminal or intercalary, sometimes in a chain, av. 24.3 μm diameter Smooth, globose, terminal or intercalary, 19–27 (av. 22.5) μm diameter Oogonia globose, smooth, terminal or less often intercalary, but frequently clustering in small groups, 19–23 (av. 21 µm) diameter Antheridia Monoclinous or diclinous, 1–2 per oogonium, antheridial cells globose, clavate or encircling an oogonium Diclinous, one to several per oogonium, antheridial stalks mostly bifurcate or differently branched near the oogonium 1–2(–5) per oogonium, monoclinous or diclinous, terminal on branched stalk, rarely intercalary, clavate or
crook-necked, hypogynous1–2 per oogonium, diclinous One, rarely two per oogonium, monoclinous, originating from the oogonial stalk at 2–10 µm or more below the oogonium, or from the parent hypha Oospores Aplerotic or plerotic, globose, 15–26 (av. 20.5 ± 3) µm diameter Aplerotic, occasionally nearly plerotic, 14–24
(av. 17.7) µm diameterMostly plerotic, av. 21.8 μm diameter Plerotic, 18–25 (av. 21.5) µm diameter Aplerotic, 16–19 (av. 17.9) µm diameter Cardinal growth temperature (minimum/optimum/
maximum temperature)5/25/30 °C 5/25/35 °C 5/35/42 °C 5/30/37 °C 5/25/35 °C Ref. This study Van der Plaats-Niterink[155] Rahman et al.[82] Van der Plaats-Niterink[150] Höhnk[158] Pythium fluminis B. Nam & Y.J. Choi, sp. nov. Fig. 13
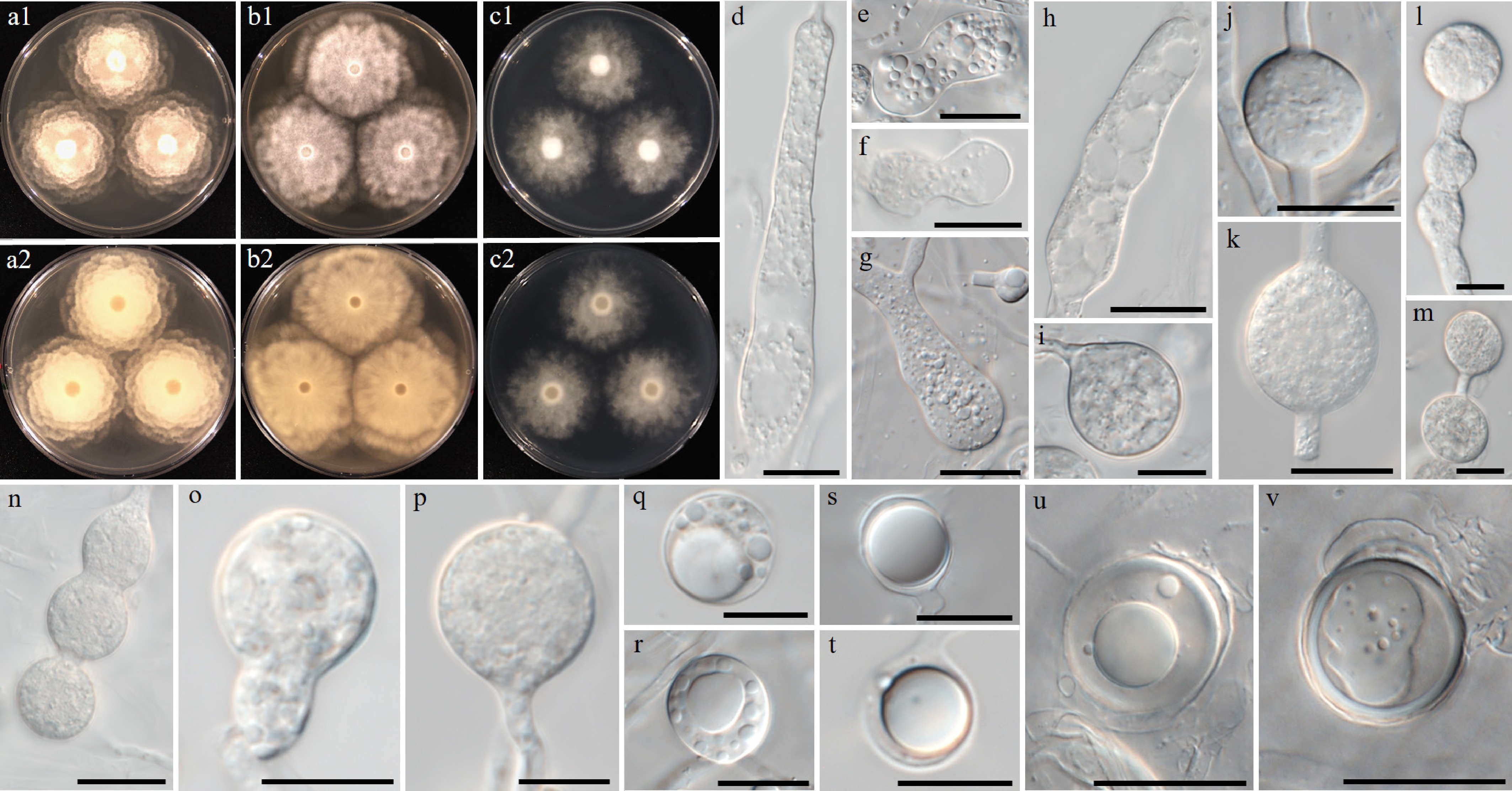
Figure 13.
Colony patterns and morphology of Pythium fluminis W1962. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(v) Microscopic structures observed under a microscope: (d) filamentous, inflated sporangium, (e)–(r) clavate, (sub-)globose, ovoid or peanut-shaped, sometimes catenulate hyphal swellings, (s), (t) globose oogonia (u), (v) with antheridia (scale bars: d–p = 10 μm, q–v = 20 μm).
MycoBank number: MB861146; Facesoffungi number: FoF18864
Etymology – Name refers to the isolation of the type strain from a lowland river (flumen Latin = river).
Main hyphae up to 7 µm wide. Sporangia were filamentous and inflated. The hyphal swellings were observed to be irregularly swollen. Hyphal swellings were clavate, (sub-)globose, rarely ovoid or peanut-shaped, and produced terminally or intercalary. Chlamydospores were not observed. Oogonia were globose and had a smooth wall. Oogonial diameter averaged 15 ± 3 µm with an overall range of 11–22 µm. Oospores were aplerotic or plerotic, and globose, ranging in size from 9.5–20 µm (av. 14 ± 3 µm) in diameter with a wall 0.5–2.5 (av. 1.7) µm thick. Antheridia were monoclinous and paragynous. Antheridial cells were clavate, bell-shaped, or with a wide contact with the oogonium.
Culture characteristics – Colonies showed a rosaceous pattern on PDA, a chrysanthemum pattern on V8A, and an irregular pattern on CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 12.5, 14.5, and 10.5 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 30, and 40 °C, respectively.
Material examined – Korea, Jeollanam-do, Haenam, isolated from soil sediments in a river, June 2021, B. Nam & Y.J. Choi, W1962 (holotype NNIBRFG35174, ex-type culture NNIBRFG35148); Jeollanam-do, Haenam, isolated from soil sediments in a river, June 2021, ditto, W1966; Jeollanam-do, Haenam, isolated from marsh snail in a river, June 2021, ditto, W1968 & W1972; Jeollanam-do, Muan, isolated from leaf litter in a river, June 2021, ditto, W1995.
GenBank numbers – W1962: ITS = PP812177, cox1 = PP798860, cox2 = PP834392; W1966: ITS = PP812178, cox1 = PP798863, cox2 = PP834393; W1968: ITS = PP812179, cox1 = PP798865, cox2 = PP834394; W1972: ITS = PP812180, cox1 = PP798869, cox2 = PP834395; W1995: ITS = PP812182, cox1 = PP798886, cox2 = PP834396.
Notes – Morphological and physiological characters of Pythium fluminis are compared with those of a phylogenetically close new taxon, Py. aquaticum, but also related to Py. catenulatum, Py. rhizo-oryzae, Py. rishiriense, and Py. torulosum (Table 5). Optimum and maximum growth temperatures of these species, including Py. fluminis and Py. aquaticum are 30 and 40 °C, respectively. Py. fluminis produces diverse shapes of hyphal bodies: (sub-)globose, ovoid, peanut-shaped, and catenulate. In cox1 Py. rishiriense is closest to Py. fluminis with a 97% similarity in the BLASTn search; however, it does not produce hyphal swellings. The ITS sequences of Py. fluminis displayed similarities of 98%–99% to phylogenetically related species, but cox1 sequences exhibited a 4%–5% genetic distance.
Pythium vigens B. Nam & Y.J. Choi, sp. nov. Fig. 14
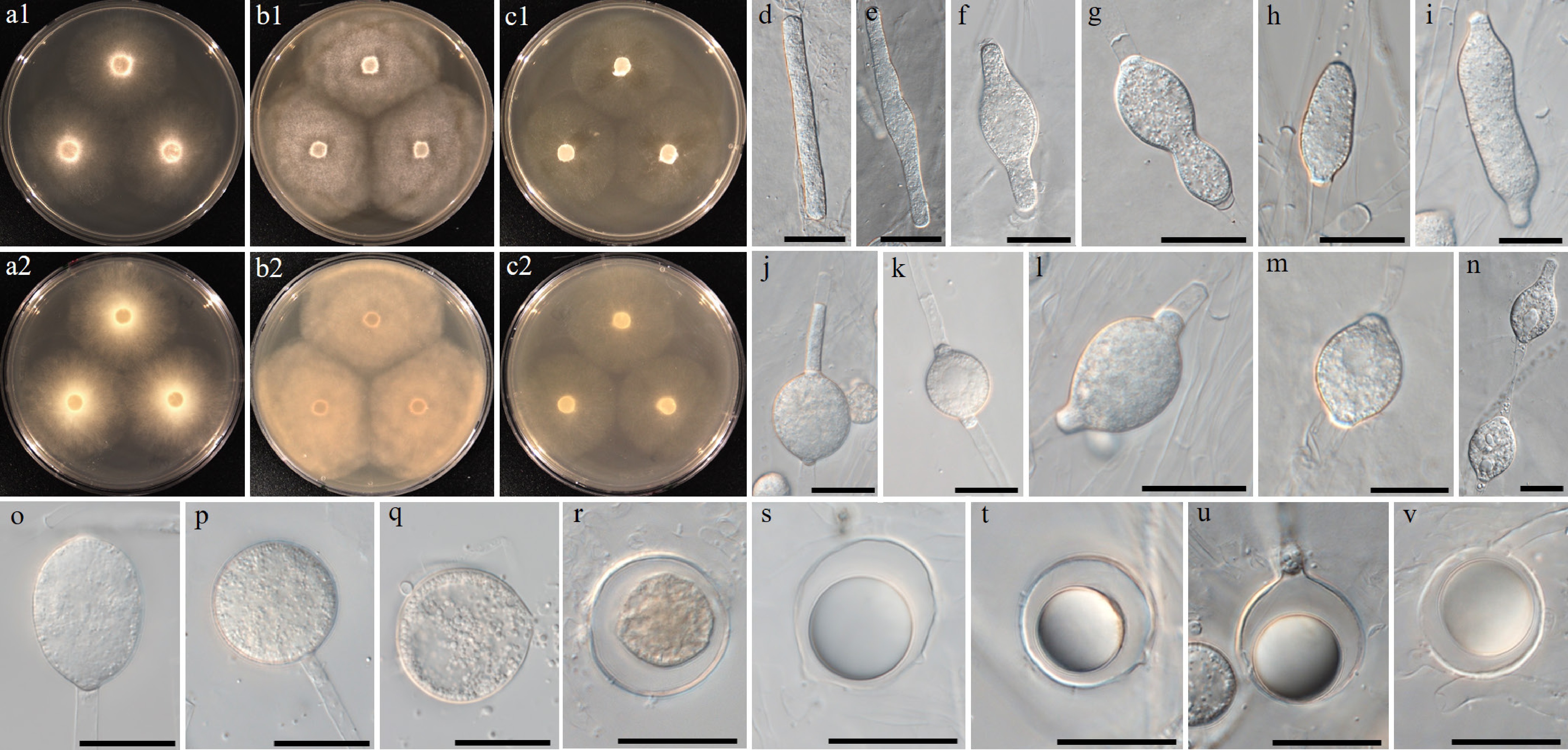
Figure 14.
Colony patterns and morphology of Pythium vigens W2006. Cultural characteristics of (a) PDA, (b) V8A, (c) CMA, after 72 h at 25 °C (1: observed view and 2: reverse view). (d)–(v) Microscopic structures observed under a microscope: (d), (e) filamentous slightly inflated structures, (f)–(q) intercalary, cylindrical, (sub-)globose, ovoid or peanut-shaped hyphal swellings, (n) rarely catenulate, (r)–(v) globose oogonia (scale bars: d–q = 10 μm, r–v = 20 μm).
MycoBank number: MB861147; Facesoffungi number: FoF18865
Etymology – Name refers to the rapid growth of the colonies (vigens Latin = being strong, vital).
Main hyphae up to 6 µm wide. Hyphal filamentous inflated structures. (Sub-)globose, ovoid, or peanut-shaped, rarely, catenulate hyphal swellings were produced terminally or intercalary. Sporangia and chlamydospores were not observed. Oogonia had smooth walls, were globose, and produced terminally or intercalary. Oogonial diameter averaged 23 ± 1 µm with an overall range of 21–26 µm. Oospores were aplerotic and globose, ranging in size from 13–16 µm (av. 15 ± 1 µm) in diameter with a wall 0.5–1.2 (av. 1) µm thick. Antheridia were absent or encircling an oogonium, showing undefined shapes.
Culture characteristics – Colonies showed a faint striate to radiate patterns on PDA, V8A, and CMA. Radial growth rates on PDA, V8A, and CMA at 25 °C were 22, 28.5, and 20 mm/d, respectively. On V8A, minimum, optimum, and maximum growth temperatures were 5, 25, and 35 °C, respectively.
Material examined – Korea, Jeollanam-do, Jindo, isolated from leaf litter in a reservoir, June 2021, B. Nam & Y.J. Choi, W2006 (holotype NNIBRFG35177 ex-type culture NNIBRFG 35151); Jeollabuk-do, Iksan, isolated from the weed in a reservoir, June 2020, ditto, W1298 & W1300; Jeollanam-do, Jindo, isolated from the leaf litter in a reservoir, June 2021, ditto, W2007, W2008, and W2011.
GenBank numbers – W2006: ITS = PP812188, cox1 = PP798896, cox2 = PP834401; W1298: ITS = PP812167, cox1 = PP798625, cox2 = PP834384; W1300: ITS = PP812168, cox1 = PP798626, cox2 = PP834385; W2007: ITS = PP812189, cox1 = PP798897, cox2 = PP834402; W2008: ITS = PP812190, cox1 = PP798898, cox2 = PP834403; W2011: ITS = PP812191, cox1 = PP798901, cox2 = PP834404.
Notes – Morphological and physiological characters of Pythium vigens are listed in Table 6 for comparison with those of phylogenetically close species, including Py. chlamydosporum. Hyphal swellings with diverse shapes were abundantly observed. Growth rate of Py. vigens is higher than phylogenetically close species, and fastest on V8A at 25 °C (28.5 mm/d). The ITS, cox1, and cox2 sequences of Py. vigens differed from those of the closest related species with sequence similarities of 95%–97%.
-
A wide and diverse range of species from the genus Pythium Pringsheim (nom. cons.) sensu lato (s.l.) were found in the present study. Recently, a phylogenomic investigation conducted by Nguyen et al.[149] supported splitting up the genus of Pythium s.l. into five distinct genera, namely Elongisporangium, Globisporangium, Phytopythium (= Ovatisporangium), Pilasporangium, and Pythium sensu stricto (s.s.), proposed by Uzuhashi[148] based on an earlier phylogenetic analysis. The division of Pythium s.l. was validated by the cox1 tree of the present study (Supplementary Fig. S1), in which the phylogenetic relationship among the newly proposed genera was unraveled. Of Pythium s.l., Globisporangium, Phytopythium, and Pythium s.s. were widely distributed in freshwater habitats of Korea. These genera are probably ubiquitous in freshwater ecosystems; they are underrepresented so far. In particular, Pythium s.s. was the most frequently isolated genus from diverse substrates such as algae, animals, plants, soil, as well as water, and was found in all seasons in the present study. Some species in this genus have been well-known due to their pathogenicity, e.g., Py. aphanidermatum, Py. catenulatum, Py. deliense, and Py. dissotocum to crops[161−163], Py. chondricola and Py. porphyrae to algae[43,164], and Py. insidiosum to mammals[165]. However, most strains of Pythium s.s. obtained in the present study inhabited plant litter, and their pathogenicity remains unknown. Previous research also reported that a diverse range of Pythium s.s. spp. inhabits plants, soil, and water in freshwater ecosystems[95,99,166]. In conclusion, it is likely that the species from this group perform a role as saprophytes in freshwater ecosystems. Masigol et al.[125,167] determined the enzymatic capacity of aquatic Saprolegniales as well as their involvement in humic substances mineralization and processing, but the role of other groups remain obscure. As shown in Table 1, pathogenic oomycetes of fishes, amphibians, and algae have been investigated primarily so far, leading to an underestimation of saprotrophic oomycetes. Various species were isolated and reported also from plants, soil, and water (Table 1), but their ecological features have not been studied. Further investigation is imperative for future research to determine the role of oomycete saprophytes in biogeochemical and ecological processes in freshwater ecosystems.
Previously, clades A–D of Pythium s.l. have merged into Pythium s.s.[143,148,149], and the presence of filamentous sporangia is their common trait. This group appears to be associated with aquatic habitats. For instance, Py. adhaerens, Py. chondricola, and Py. porphyrae in clade A originate from marine algae, Py. biforme, Py. brachiatum, Py. junctum, Py. subutonaiense, and Py. utonaiense in clade B from freshwater environment[99,105], and Py. grandisporangium and Py. insidiosum in clade C from reed and seawater[143]. In the present study, Clade B showed the highest number of isolates and the largest number of taxa: ca. 95% of total strains of Pythium s.s. and 40 taxa. Moreover, we described six newly encountered species belonging to clade B of Pythium s.s., i.e., Py. aculeatum, Py. aquaticum, Py. chlamydosporum, Py. intricatum, Py. fluminis and Py. vigens. In addition, the other seven taxa are potential new species of Pythium s.s. also belonged to clade B. These taxa were isolated mainly from plant sources. Considering these factors, it is conceivable that freshwater plants or organic materials may harbor a greater diversity of clade B of Pythium s.s.
Genus Phytopythium was also prominently identified in the current study. All eight Phytopythium species were detected in the sampled plant sources, which mainly consisted of plant litter. The findings validate freshwater environments as suitable habitats for these organisms, along with Pythium s.s.[87,168]. Two novel species of Phytopythium were also described in the present study: Pp. minutum and Pp. lacustre. Phytopythium was originally classified in clade K of Pythium s.l.[143], but introduced to separate it from other groups of Pythium s.l. by Bala et al.[169] and de Cock et al.[170], and by Uzuhashi et al.[148] under the synonymous name Ovatisporangium. The members of Phytopythium produce globose to ovoid papillate sporangia, unlike Pythium s.s.[169], and further subdivided into three phylogenetic clades[171]. Our novel species, Pp. minutum and Pp. lacustre, belongs to the clade 1 sensu Baten et al.[171] and de Cock[170]. Pp. minutum formed a monophyletic group with Pp. citrinum and Pp. delawarense. Pp. lacustre is close to Pp. iriomotense. Their sister species have also been previously reported from freshwater ecosystems[52,93].
Various Globisporangium species were also found in this study; however, they were absent from the plant samples. For instance, G. macrosporum, G. sylvaticum, and G. ultimum (= Pythium ultimum var. ultimum), known for their aggressiveness to certain plant species[150,172,173], were exclusively recovered from soil and/or water samples in the present study. The greater part of the genus Globisporangium was found in soil sediments. The absence of Globisporangium species in the plant sources prompts further investigation. It is plausible that these oomycete species exhibit a preference for young or actively growing plants[150], in contrast to the plant litter used as the plant substrate in this study.
Our study found congeners of Saprolegniales: Achlya, Aphanomyces, Geolegnia, Leptolegnia, and Saprolegnia. This group preferred to inhabit plant-derived sources, although they were found from diverse substrates, including algae, animals, plants, soil, and water. However, two species, namely Achlya bisexualis and Saprolegnia australis, were isolated only from animal sources, such as fishes and marsh snails. Overall, Saprolegniales is strongly associated with animal sources such as amphibians, crustaceans, fishes, and mosquitoes, which is in line with previous research. (Table 1). Previous studies have frequently reported that the Saprolegniales group, including, Achlya, Aphanomyces, as well as Saprolegnia, are commonly isolated from fish and crustaceans and are often considered potential pathogens[39,41,133,174,175]. Notorious pathogens belonging to Saprolegniales are responsible for high-profile declines in aquatic animals and cause economic damage to aquaculture. As our research primarily involved sampling from plants, soil, and water, where non-pathogenic strains are more likely to occur, Saprolegniales taxa frequently reported in host-associated studies were rarely detected in the present study. However, in our survey, which included amphibian and fish samples, the pathogenic species frequently reported in previous studies were indeed found to be more frequently isolated from these animal-associated samples: Achlya bisexualis, Saprolegnia australis, S. diclina, and S. ferax. Based on the findings of this study, future research should aim to include a greater diversity of host organisms, particularly animal-associated samples, to comprehensively compare oomycete communities across host types. This will contribute to a more complete understanding of host-oomycete interactions and ecology in freshwater ecosystems.
In general, the saprotrophic oomycetes inhabiting freshwater have, until recently, received less attention. However, diverse plant pathogenic species of the genus Phytophthora have been reported from freshwater ecosystems[12−19]. In the present study, only three Phytophthora taxa have been found, which is much lower than the diversity found in previous freshwater surveys. All three taxa are mainly aquatic saprophytes, and primarily plant pathogenic species were missing. Previous studies obtained various species of Phytophthora from the streams and rivers using leaf baiting approaches or selective agar with hymexazol, which restricts the quick-growing Pythiales group. The study of Perez-Sierra et al.[176] and Jung et al.[16,66] established the methods for sampling and isolation of Phytophthora. For Phytophthora isolations from sediment and water samples, a baiting approach along with selective agar with hymexazol would have been needed. Given the broad scope of this study, the methodology was designed to capture overall oomycete diversity rather than to maximize the detection of specific groups. Consequently, some taxa, especially those requiring specialized isolation or detection techniques, may have been underrepresented. Previous metagenomic studies have revealed an exceptionally high Phytophthora diversity in freshwater environments[21,177], suggesting that many Phytophthora taxa remain undiscovered in Korea. This reinforces the need for follow-up studies that employ more targeted and diverse isolation strategies, which are likely to uncover additional Phytophthora groups beyond those identified in the present study.
Our extensive and long-term surveys have garnered noteworthy insights into an unexplored area of freshwater oomycetes that have gained less attention so far than terrestrial oomycete groups. This study contributes to our knowledge of the diversity of freshwater oomycetes and attempts to understand ecological features. In addition, we suggested that the saprotrophic oomycetes have been underestimated and poorly studied, despite their crucial role in decomposing and recycling organic materials in aquatic ecosystems. Our results highlight the need for targeted research into the specific ecological roles of oomycetes in freshwater environments. Employing selective isolation techniques and integrating culture-independent approaches such as metagenomics may help uncover functionally significant or currently unculturable oomycete taxa. These approaches could contribute to a more complete understanding of host-oomycete interactions and ecological roles in freshwater ecosystems.
This study was supported by the Nakdonggang National Institute of Biological Resources (NNIBR) (Grant No. NNIBR20261103). The authors thank Professor Jaime Blair (Department of Biology, Franklin & Marshall College) for providing oomycete strain data and related background information.
-
The authors confirm contributions to the paper as follows: study conception and design: Nam B, Choi YJ; data collection: Nam B, Na DH, Lee JS; analysis and interpretation of results: Nam B, Bennett RM, Tojo M; writing − original draft manuscript preparation: Nam B; writing − review and editing: Bennett RM, Tojo M, Choi YJ, Lee HB, Mun HY, Kang KH; supervision: Choi YJ, Lee HB; project administration, funding acquisition: Lee HB, Mun HY. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/mycosphere-0026-0006.
- Supplementary Table S1 Strain information and collection details.
- Supplementary Fig. S1 Phylogenetic tree of Peronosporales and Pythiales species from the minimum evolution analysis based on cytochrome c oxidase subunit I (cox1) mtDNA sequences.
- Supplementary Fig. S2 Phylogenetic tree of Saprolegniales species from the minimum evolution analysis based on internal transcribed spacer (ITS) rDNA sequences.
- Supplementary Taxonomy S1 Descriptions of Oomycota species previously unrecorded in Korea Freshwater samples, including algae, animal debris, plant litter, soil sediment, and water, were collected from various freshwater environments in Korea. B. Nam and Y.J. Choi collected samples, from which B. Nam isolated pure oomycete cultures.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Nam B, Bennett RM, Tojo M, Na DH, Lee JS, et al. 2026. Uncovering the hidden diversity of oomycetes (Straminipila) in freshwater environments. Mycosphere 17: e006 doi: 10.48130/mycosphere-0026-0006 

Uncovering the hidden diversity of oomycetes (Straminipila) in freshwater environments
- Received: 11 November 2025
- Revised: 22 December 2025
- Accepted: 17 March 2026
- Published online: 12 May 2026
Abstract: Oomycetes play a key role in decomposing and recycling organic materials in freshwater ecosystems. However, their diversity and ecological study gained less attention compared with other organisms. Based on extensive, long-term surveys for freshwater oomycetes, the present study undertakes a comprehensive examination of oomycete diversity and ecology in the Korean freshwater environments. We successfully identified a total of 78 taxa of oomycetes (568 isolates), which are classified into the orders Peronosporales (4%), Pythiales (77%), and Saprolegniales (19%), based on their cultural, morphological, and molecular characteristics. These taxa consist of 57 previously described species, including 22 species new to Korea, and 21 previously unknown lineages. Eight of these taxa were described as new species under the Pythiales, with detailed taxonomic descriptions in this study. Our findings revealed that the distributions of freshwater oomycetes varied depending on substrates and that oomycetes were primarily found in plant-derived sources, such as decaying leaves, stems, and twigs. Across all seasons and freshwater ecosystem types, Pythiales were more frequently isolated than Peronosporales and Saprolegniales. However, isolation frequencies among taxa varied across seasons and habitat types. This study provides extensive strain resources and establishes a fundamental reference for future taxonomic and ecological research on freshwater oomycetes, expanding knowledge of their diversity and functional roles in freshwater ecosystems.
-
Key words:
- Phytopythium /
- Pythium /
- Saprophytes /
- Taxonomy /
- Water mold