









-
The escalating global demand for food has intensified agricultural production, making the application of nitrogen (N) fertilizers a cornerstone of modern agriculture[1]. While essential for achieving high crop yields and ensuring food security, the extensive use of synthetic N fertilizers has created significant environmental challenges[2]. A significant portion of applied N is lost to the environment, contributing to water pollution and the emission of nitrous oxide (N2O), a potent greenhouse gas with a global warming potential approximately 300 times that of CO2[3]. Agricultural soils are the largest anthropogenic source of N2O, placing the management of soil N cycling at the critical relation of food production and climate change mitigation[1].
Soil N2O emissions are determined by the delicate balance between microbial production and reduction processes, primarily nitrification and denitrification[4]. Nitrification, the oxidation of ammonia to nitrate, can produce N2O as a byproduct and is carried out by distinct microbial guilds, including archaeal (AOA) and bacterial (AOB) ammonia-oxidizers, as well as comammox bacteria[5]. Denitrification is a stepwise reduction of nitrate to dinitrogen gas (N2), with N2O as an intermediate. The final step, the reduction of N2O to N2, is catalyzed by the nitrous oxide reductase enzyme, encoded by the nosZ gene[6]. Consequently, the N2O emission potential of a soil is controlled by the relative abundances and diversities of microbial communities harboring genes for N2O production (e.g., amoA, nirK, nirS) vs N2O reduction (nosZ)[7,8].
The assembly of soil microbial communities is governed by the interplay between deterministic and stochastic processes. Deterministic processes, such as environmental filtering and species competition, involve niche-based selection where the 'fittest' organisms for a given environment are favored, leading to predictable community structures. In contrast, stochastic processes, including dispersal limitation, ecological drift, and random colonization events, result in community patterns that are largely governed by chance[9]. Anthropogenic activities, particularly long-term agricultural fertilization, represent a powerful environmental pressure that can significantly alter the balance between these two forces. It is widely hypothesized that by increasing the availability of limiting nutrients, fertilization imposes strong selective pressures that strengthen the role of determinism, as only a subset of taxa can tolerate or thrive in the altered soil chemistry, such as changes in pH and nutrient concentrations[9]. However, the nature of this shift can depend on the specific fertilization regime; while nutrient enrichment initially increases selection, extreme or unbalanced fertilization might lead to environmental homogenization, which could paradoxically increase the relative importance of stochasticity by weakening niche differentiation among surviving taxa[10,11]. Different agricultural management strategies, such as the use of synthetic versus organic fertilizers, are known to alter these microbial communities. Integrated nutrient management, which combines inorganic fertilizers with organic amendments like manure, has been proposed as a sustainable alternative that can improve soil health and nutrient use efficiency[12,13]. Phylogenetic null model analyses demonstrated that prolonged stover recycling mitigated the processes of dispersal limitation and homogeneous selection in rhizosphere fungal communities, with their formation and maintenance predominantly reliant on stochastic events rather than species' adaptive traits[14]. However, a clear understanding of how these practices specifically influence the key N cycling functional guilds and the ecological processes governing their community assembly is lacking. It is unclear whether the observed changes in N cycling are driven by deterministic selection or by stochastic processes like dispersal limitation, particularly in studies relying primarily on the 16S rDNA or ITS genes[15]. The relative importance of the deterministic and stochastic processes on soil microbial communities varies across environmental gradients, succession stages, and among different assemblages[16,17].
Therefore, understanding how different fertilization strategies modulate the relative influence of determinism and stochasticity is crucial for developing reliable strategies to engineer soil microbiomes for reduced N2O emissions. This study was designed to investigate the long-term impacts of different fertilizer regimes on soil properties, crop yield, N2O emissions, and the underlying assembly of N cycling microbial communities. It was hypothesized that: (1) an integrated strategy of balanced manure with synthetic N would maintain high crop yields while mitigating N2O emissions compared to conventional synthetic N application; (2) this mitigation would be mechanistically driven by a balance of the abundance and diversity of the N2O-producing and -reducing community; and (3) the assembly of these key N cycling functional guilds would be linked to N2O emissions. To test these hypotheses, a long-term field experiment consisting of four treatments was set: no fertilizer N applied; optimum rate of chemical N fertilizer; conventional rate of chemical N fertilizer; and balanced manure with synthetic N. This study employed a multifaceted approach combining measurements of soil physicochemical properties, N-transformation rates, and N2O fluxes with advanced molecular techniques (qPCR, high-throughput sequencing) and infer community assembly (iCAMP) mechanisms by phylogenetic-bin-based null model analysis to provide a comprehensive, mechanism-based assessment of sustainable nitrogen management in agriculture.
-
This study was conducted using a long-term field experiment located at the China Agricultural University Research Station in Shangzhuang, Beijing, China (39°48′ N, 116°28′ E, 40 m elevation). The annual mean air temperature and precipitation from 1981 to 2015 were 13.0 °C and 540 mm, respectively. The soil is a typical upland agricultural soil in the North China Plain, characterized by a bulk density of 1.3 g cm−3 and a clay loam texture (28.0% clay, 32.0% silt, 40.0% sand, according to the USDA standard). The studied winter wheat−summer maize rotation is the dominant cropping system in this region, in which wheat is sown in early October and harvested in early June of the following year, immediately followed by maize sowing, with harvest occurring in late September.
The long-term field experiment, initiated in October 2006, consisted of four N application rates. The N application treatments were: (1) zero N (N0), no fertilizer N applied; (2) optimum N (Nopt), optimum rates of chemical N fertilizer, determined via the mineral N test method to achieve synchronization of crop N demand with soil N supply; (3) conventional N (Ncon), conventional rates of chemical N fertilizer according to the local farming practice, with 300.0 and 260.0 kg N ha−1 during the wheat and maize seasons, respectively; (iv) balanced manure with synthetic N (Nbal + M), in which cattle manure was combined with supplementary chemical fertilizer, calculated to balance crop uptake and soil N surplus. The plot size was 8 × 8 m, and the number of replicates was three. Detailed information, including each N fertilization and irrigation, soil chemical properties for each treatment, and management activities, was described in the previously published papers[18]. Soil samples were collected on 22 July 2022, during the summer maize season, approximately 10 d after N fertilizer application (10 July) at the fourth-leaf growth stages and before the fertilization at the tenth-leaf growth stages (2 August). This sampling timing was deliberately chosen to minimize short-term microbial responses to N application and reflect the long- term effects of different treatments. Furthermore, the summer maize season accounts for up to 84% of annual N2O emission, as elevated temperatures combined with rainfall enhance microbial activity involved in key nitrogen cycle processes, particularly nitrification and denitrification. Sampling at this period enables assessment of the genetic potential of these active microbial populations. Soil properties in the top 0–20 cm layer from the experimental treatments are shown in Table 1. Soil samples were placed on ice and transported back to the laboratory, where visible grassroots and pebbles were subsequently removed. The soil samples were stored at –80 and 4 °C for DNA extraction and soil geochemical measurements, respectively[19]. The data on cumulative N2O emissions were cited from Wei et al.[18,20], who measured the N2O emissions from 2018 to 2019 in the same long-term field experiment. The methods for determining pH, SOC, TN, AK, NH4+–N, NO3––N, C/N ratio, PNR, and PDR are provided in Supplementary File 1 (S1.1–S1.3).
Table 1. Soil properties under different fertilizer treatments while sampling in 2022
Treatment pH SOC
(g kg–1)TN
(g kg–1)C/N Olsen P
(mg kg–1)AK
(mg kg–1)NH4+-N
(mg kg–1)NO3–-N
(mg kg–1)PNR
(mg N kg–1 h–1)PDR
(mg N kg–1 h–1)N0 7.96 ± 0.05 a 6.68 ± 0.19 c 0.72 ± 0.02 c 9.28 ± 0.08 a 28.26 ± 1.36 bc 93.94 ± 4.42 b 0.6 ± 0.06 b 3.03 ± 0.39 b 11.91 ± 1.58 b 0.55 ± 0.07 c Nopt 7.98 ± 0.03 a 7.6 ± 0.6 bc 0.88 ± 0.11 bc 8.74 ± 0.36 a 17.09 ± 1.23 c 116.04 ± 26.8 a 0.91 ± 0.19 a 8.81 ± 1.43 ab 39.58 ± 2.17 ab 0.70 ± 0.03 bc Ncon 7.94 ± 0.02 a 7.65 ± 0.47 bc 0.88 ± 0.02 bc 8.66 ± 0.52 a 19.72 ± 2.3 c 92.83 ± 3.32 b 0.94 ± 0.11 a 9.47 ± 1.99 a 41.44 ± 2.12 ab 0.86 ± 0.04 b Nbal + M 7.97 ± 0.01 a 10.33 ± 0.28 a 1.2 ± 0.05 a 8.62 ± 0.34 a 47.9 ± 1.74 a 113.83 ± 5.85 ab 0.85 ± 0.05 ab 8.75 ± 3.57 ab 47.25 ± 1.92 a 1.15 ± 0.06 a SOC, soil organic carbon; TN, total nitrogen content; AK, available K; NH4+–N, ammonium content; NO3––N, nitrate content; PNR, potential nitrification rate; PDR, potential denitrification rate. Different letters indicate the significance among treatments at p < 0.05 level by one-way ANOVA. Error bars represent standard deviation (n = 3). Soil quality index and crop yield
-
Soil quality is determined by various physical, chemical, and biological properties of soil. To assess soil quality consistently and accurately, systematic methods are needed for measuring and interpreting these properties. The soil quality index (SQI) was determined through the following steps[21]: (1) Indicator selection: The evaluation used a dataset including soil pH, SOC, TN, Olsen P, and available K. Principal component analysis (PCA) was applied to select factors from these soil attributes. (2) Indicator scoring and weighting: Each indicator was scored between 0.1 and 1.0 using the standard S equation[22]. The communality of each indicator was extracted by PCA, and weights were calculated as the quotient of each indicator's communality divided by the sum of all communalities. (3) Soil quality index (SQI) calculation: After scoring and weighting, the SQI was calculated using:
$ S QI={\sum }_{i=1}^{n}{W}_{i}{N}_{i} , $ where, Wi is the assigned weight, Ni is the indicator score, and n is the number of indicators.
At maturity, a 6 m2 (3 m × 2 m) area of winter wheat and a 10.8 m2 area (three rows, 6 m long per row) of summer maize were selected to harvest the aboveground crops, which were immediately separated into grains and straw. The samples were then dried separately at 70 °C and weighed to calculate the grain yield.
Sequencing and bioinformatics processing
-
For bacterial and archaeal 16S rRNA genes, the V4 region was amplified with the primer pair 515F (5'-GTGCCAGCMGCCGCGGTAA-3') and 806R (5'-GGACTACHVGGGTWTCTAAT-3')[23]; the primer pair Arch-amoAF (5'- STAATGGTCTGGCTTAGACG-3') and Arch-amoAR (5'-GCGGCCATCCATCTGTATGT-3') for archaeal amoA gene[24]; the primer pair amoA-1F (5'-GGGGTTTCTACTGGTGGT-3') and amoA-2R (5'-CCCCTCKGSAAAGCCTTCTTC-3') for bacterial amoA gene[25]; the primer pair comaA-244F (5'-TAYAAYTGGGTSAAYTA-3') and comaA-659R (5'-ARATCATSGTGCTRTG-3') for comammox amoA gene[26]; the primer pair cd3aF (5'- TSAACGTSAAGGARACSGG-3') and R3cd (5'-GASTTCGGRTGSGTCTTGA-3') for nirS gene[27]; the primer pair F1aCu (5'-ATCATGGTSCTGCCGCG-3') and R3Cu (5'-GCCTCGATCAGRTTGTGGTT-3') for nirK gene[28]; the primer pair nosZ-F (5'-CGYTGTTCMTCGACAGCCAG-3') and nosZ-R (5'-CGSACCTTSTTGCCSTYGCG-3') for nosZI gene[29]; and the primer pair nosZ-II-F (5'-CCTIGGICCIYTKCAYAC-3') and nosZ-II-R (5'-GCIGARCARAAITCBGTRC -3') for nosZII gene[30].
The details of qPCR, PCR amplification, amplicon purification, library preparation, Illumina NanoSeq sequencing, and sequence processing were described previously[31,32], and conducted by MAGIGENE company. Amplification conditions for qPCR were as follows: 95 °C for 3 min, 40 cycles of 10 s at 95 °C, 30 s at 52–65 °C, 30 s at 72 °C,and a final extension at 72 °C for 5 min, then held at 16 °C. qPCR amplification efficiencies of these eight genes were between 92% and 105%, and details of Melting Curves are shown in Supplementary File 1 (S1.4). Sequence processing was conducted on an in-house pipeline (
https://dmap.denglab.org.cn ) integrated with the necessary bioinformatics tools[31−33]. Unoise3[34] was used to remove chimeras and classify the sequences into Amplicon Sequence Variants (ASVs). FrameBot[35] was applied to check and correct the represented nucleotide sequences of ASVs from sequencing samples against the NcycFunGen database (https://ncycfungen.denglab.org.cn )[36] with protein sequences at a high level. The sequencing data are available in the National Microbiology Data Center (https://nmdc.cn/resource/genomics/project ) with project number NMDC10019933.Biodiversity and statistical analysis
-
Non-metric multidimensional scaling (NMDS) was used to show the differences among the four microbial communities, and the significance of the four fertilizer treatments' contributions to microbial community structure was analyzed by permutational multivariate analysis of variance (PERMANOVA). Further, the relationship between biodiversity and environmental variables was evaluated using the Mantel test. In addition, one-way ANOVA was used to test the differences among multiple groups. All biodiversity and statistical analyses were conducted with an online in-house pipeline (
https://dmap.denglab.org.cn )[32,33].To quantify the underlying mechanisms of microbial community assembly, the study utilised a newly developed method called infer community assembly mechanisms by phylogenetic-bin-based null model analysis (iCAMP)[15]. iCAMP assigns the microbial ASVs into phylogenetic tree-based groups and then quantifies the contribution of each ecological process to microbial community assembly, including homogeneous selection, heterogeneous selection, dispersal limitation, homogenizing dispersal, and undominated processes (drift and others). Null model analysis of the phylogenetic diversity was performed using the beta Net Relatedness Index (βNRI), and taxonomic β-diversities were assessed with the modified Raup–Crick metric (RC). Afterwards, the proportion of each ecological process between different samples was compared based on the bootstrapped methods (n = 1,000). The analyses were conducted with the online pipeline (
http://ieg3.rccc.ou.edu:8080 )[15], and the other parameters were set to pipeline defaults. -
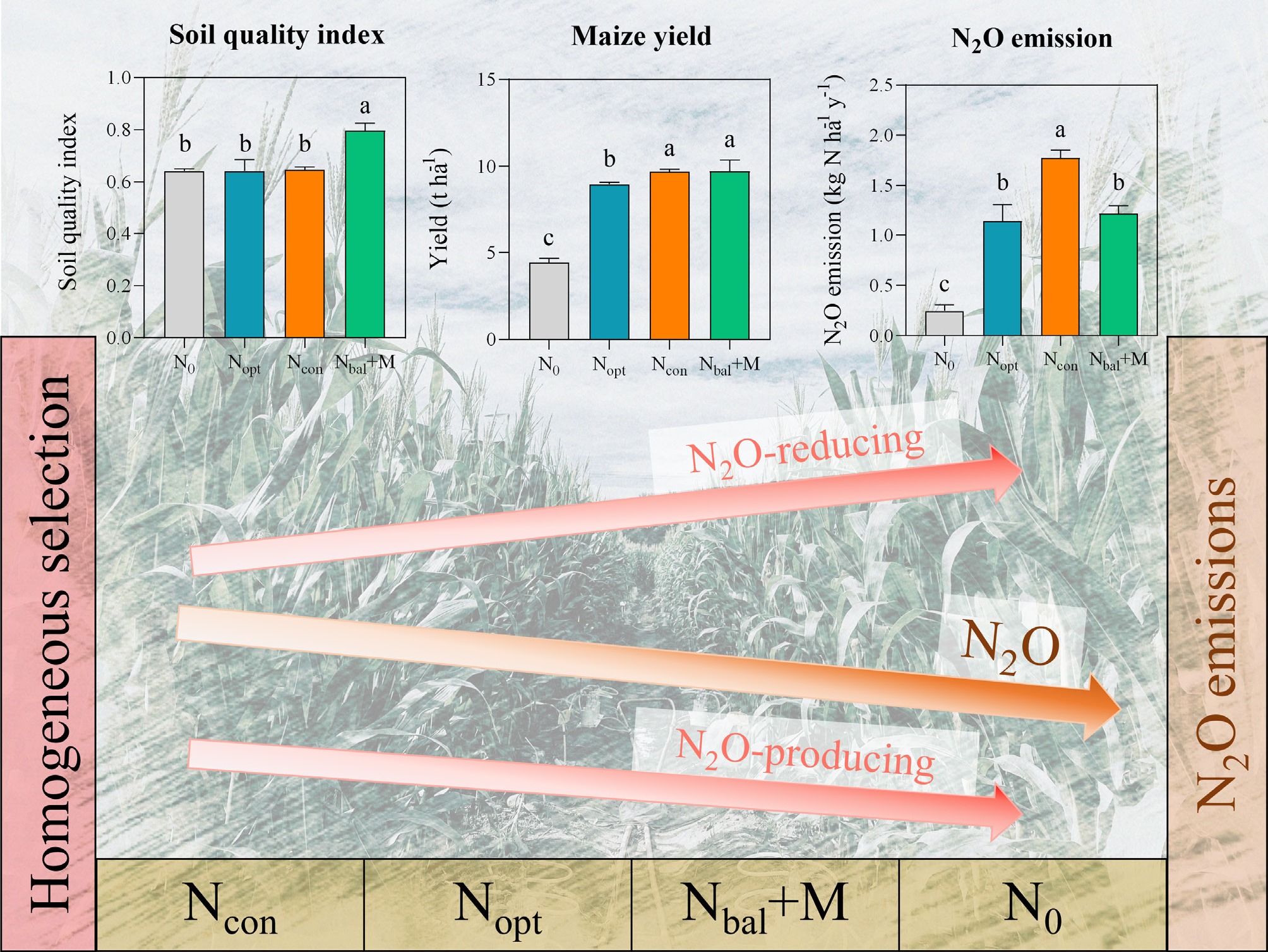
Different long-term fertilizer treatments affected key soil properties. There were no significant differences (p > 0.05; Table 1) in pH or soil C/N ratio observed among the four fertilizer treatments. In contrast, the Nbal + M treatment resulted in the highest concentrations of total N content (TN), soil organic carbon content (SOC), and Olsen P (p < 0.05; Table 1). For available K, the Nopt treatment recorded the highest value. All fertilizer applications (Nopt, Ncon, Nbal + M) significantly increased soil ammonium (NH4+–N) and nitrate content (NO3––N) compared to the N0 control (p < 0.05; Table 1). Potential nitrification (PNR) and denitrification rates (PDR) were significantly enhanced by fertilizer application (p < 0.05; Table 1). The soil quality index (SQI) was significantly enhanced by Nbal + M (p < 0.05; Fig. 1a). Crop yield was substantially improved by all fertilization strategies compared to N0, with the highest yield recorded in the Ncon and Nbal+M (p < 0.05; Fig. 1b). Nitrous oxide (N2O) emissions were significantly influenced by fertilizer application (p < 0.05; Fig. 1c). The lowest emission was recorded in the N0 control, and the highest was in Ncon.
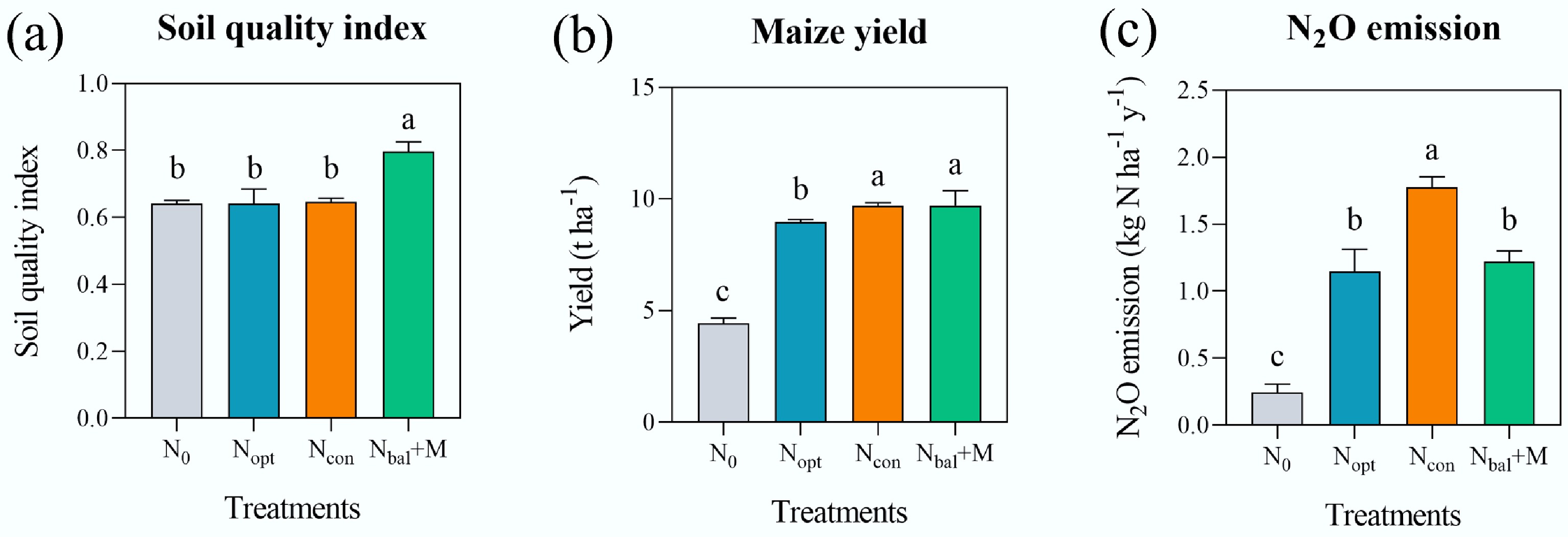
Figure 1.
(a) Soil quality index, (b) annual maize yield in 2022, and (c) average annual N2O emission from 2018 to 2019 under different fertilizer treatments. N0: no N, Ncon: conventional N, Nopt: optimal N, Nbal + M: balanced manure with synthetic N. Different letters indicate the significance among treatments at p < 0.05 level by one-way ANOVA. Error bars represent standard deviation (n = 3).
In summary, Nbal + M was the most effective strategy for improving soil quality, significantly increasing SOC, TN, Olsen P, PNR, PDR, SQI, and crop yield, with lower N2O emissions.
N cycling functional communities under different fertilizer treatments
-
To investigate the microbial mechanisms underlying the observed changes in nitrogen transformations, the abundances of key functional genes involved in the N cycling, as well as the total bacterial abundance, were quantified via qPCR (Fig. 2). There were no significant differences among the four treatments in 16S rDNA (Fig. 2a), archaeal amoA (Fig. 2b), comammox amoA (Fig. 2d), nirK (Fig. 2e), and nirS (Fig. 2f) gene abundance (p > 0.05).
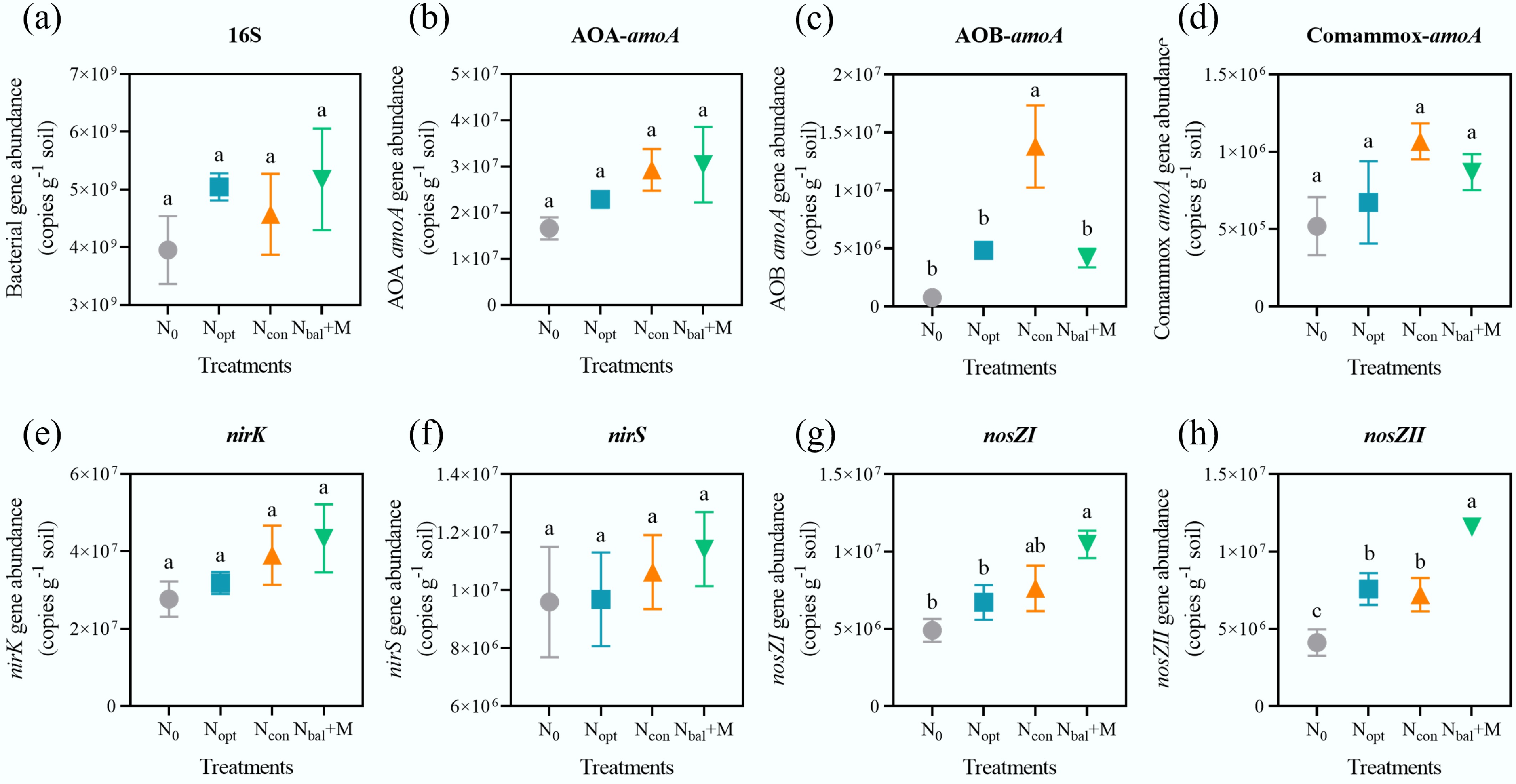
Figure 2.
Nitrogen cycling gene abundances under different fertilizer treatments. (a) 16S rDNA. (b) Archaeal amoA gene. (c) Bacterial amoA gene. (d) Comammox amoA gene. (e) nirK gene. (f) nirS gene. (g) nosZI gene. (h) nosZII gene. Different letters indicate the significance among treatments at p < 0.05 level by one-way ANOVA. Error bars represent standard deviation (n = 3).
Ncon significantly increased bacterial amoA gene abundance compared to the other three treatments (p < 0.05; Fig. 2c). However, the abundance of nosZ genes, responsible for the final step of denitrification (N2O reduction to N2), was significantly influenced by the fertilizer regimes. Ncon and Nbal + M showed the highest nosZI gene abundances (p < 0.05; Fig. 2g), while all three fertilizer treatments significantly increased the nosZII gene abundances, with Nbal + M being the highest (p < 0.05; Fig. 2h).
To assess the impact of different fertilizer treatments on the community structure of key N cycling genes, Non-metric Multidimensional Scaling (NMDS) analysis was performed based on Bray-Curtis dissimilarity. The significance of the community shifts was evaluated using a dissimilarity test (PERMANOVA). The overall bacterial community structure (p = 0.001; Fig. 3a), bacterial amoA (p = 0.049; Fig. 3c), comammox (p = 0.001; Fig. 3d), nirK (p = 0.011; Fig. 3f), and nosZII (p = 0.001; Fig. 3h) functional community structures were significantly altered by the fertilizer treatments, with the N0 samples separating from the fertilized treatments. In contrast, there were no significant differences in archaeal amoA (p > 0.05; Fig. 3b), nirS (p > 0.05; Fig. 3e), and nosZI (p > 0.05; Fig. 3g) functional community structures under different treatments.
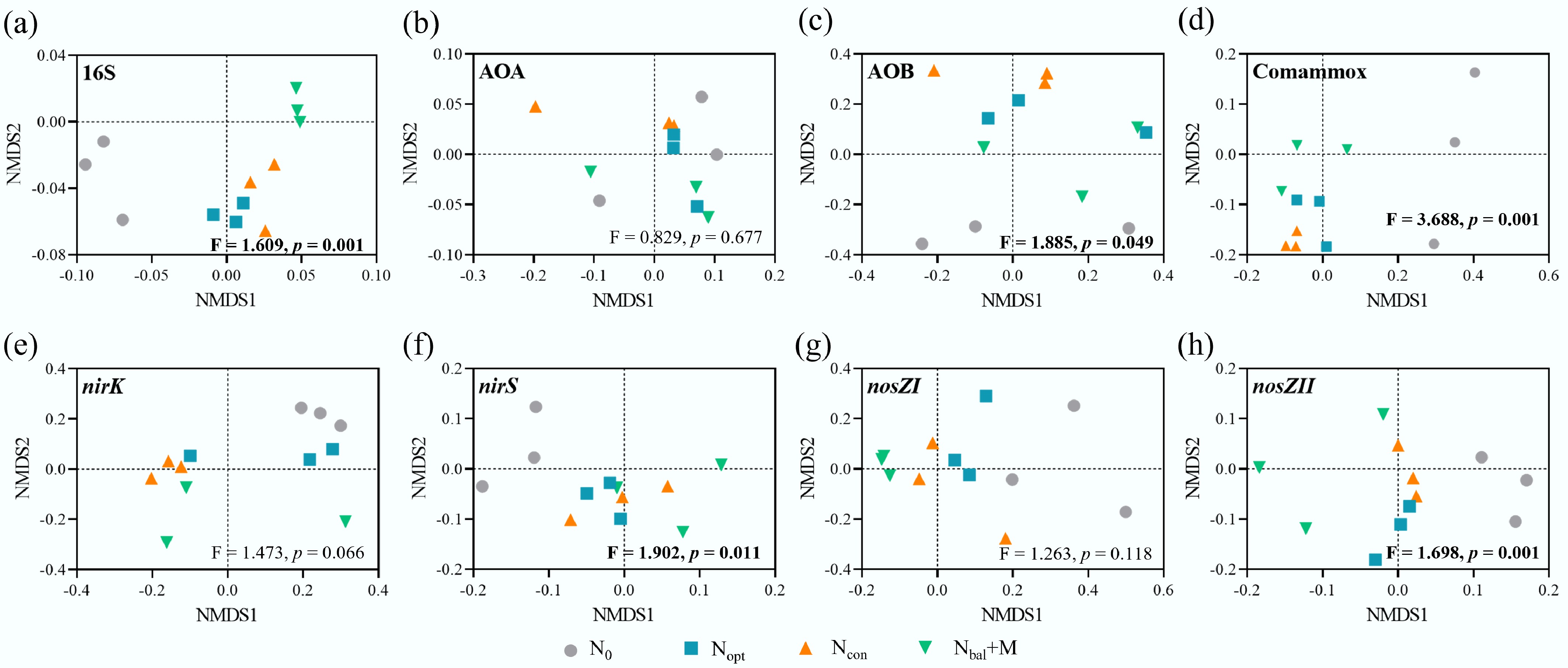
Figure 3.
Effects of different fertilizer treatments on nitrogen functional microbial communities based on NMDS. (a) 16S rDNA. (b) Archaeal amoA gene. (c) Bacterial amoA gene. (d) Comammox amoA gene. (e) nirK gene. (f) nirS gene. (g) nosZI gene. (h) nosZII gene. F- and p-values are based on dissimilarity test.
A Mantel test was performed to correlate soil properties and N2O emissions with the community composition of total bacteria and N functional guilds (Table 2). N2O emission, SOC, TN, Olsen P, PNR, and PDR were the significant variations correlated to N cycling functional community structures (p < 0.05; Table 2). In particular, PNR, PDR, and N2O emissions were significantly correlated to the denitrifying and comammox communities.
Table 2. Mantel test of N2O, soil properties with nitrogen functional microbial communities
16S AOA AOB Comammox nirK nirS nosZI nosZII r P r P r P r P r P r P r P r P N2O (kg N ha−1 y−1) 0.456 0.007 −0.004 0.439 0.343 0.017 0.733 0.001 0.134 0.194 0.438 0.004 0.293 0.044 0.199 0.118 pH 0.053 0.409 −0.122 0.743 −0.078 0.659 0.200 0.126 −0.080 0.606 0.007 0.461 0.462 0.008 −0.099 0.658 SOC (g kg−1) 0.434 0.008 0.089 0.273 −0.125 0.781 0.201 0.077 0.280 0.045 0.076 0.313 0.199 0.113 0.427 0.010 TN (g kg−1 0.477 0.001 0.014 0.374 −0.135 0.813 0.266 0.053 0.379 0.012 0.009 0.462 0.154 0.161 0.411 0.006 C/ N ratio −0.046 0.570 −0.248 0.992 −0.188 0.908 −0.003 0.457 −0.035 0.529 −0.193 0.880 −0.050 0.561 −0.107 0.710 Olsen P (mg kg−1) 0.336 0.036 0.109 0.223 −0.011 0.407 0.098 0.202 0.288 0.049 −0.023 0.565 −0.002 0.503 0.480 0.007 AK (mg kg−1) 0.017 0.530 −0.236 0.905 -0.314 0.999 −0.292 0.963 -0.187 0.787 -0.317 0.933 −0.321 0.975 −0.294 0.841 NH4+−N (mg kg−1) 0.100 0.253 −0.239 0.972 −0.067 0.636 0.113 0.216 −0.094 0.698 0.044 0.365 −0.054 0.582 −0.260 0.945 NO3−−N (mg kg−1) 0.183 0.141 −0.031 0.516 0.000 0.433 0.312 0.047 −0.008 0.506 0.223 0.101 0.021 0.408 −0.008 0.486 PNR (mg N kg−1 h−1) 0.602 0.004 0.060 0.303 0.129 0.228 0.795 0.003 0.318 0.042 0.459 0.005 0.471 0.008 0.320 0.045 PDR (mg N kg−1 h−1) 0.626 0.001 0.053 0.339 0.093 0.253 0.492 0.006 0.465 0.008 0.252 0.061 0.191 0.147 0.564 0.002 SOC, soil organic carbon; TN, total nitrogen content; AK, available K; NH4+−N, ammonium content; NO−−N, nitrate content; PNR, potential nitrification rate; PDR, potential denitrification rate. p-values shown in bold are significant at the 0.05 level. Ecological processes of N cycling functional guilds
-
The iCAMP analysis was used to quantify the relative influence of deterministic and stochastic processes on the assembly of N cycling functional microbial communities. The analysis partitions community turnover into contributions from homogeneous selection, heterogeneous selection, dispersal limitation, homogenizing dispersal, and drift processes (Fig. 4).
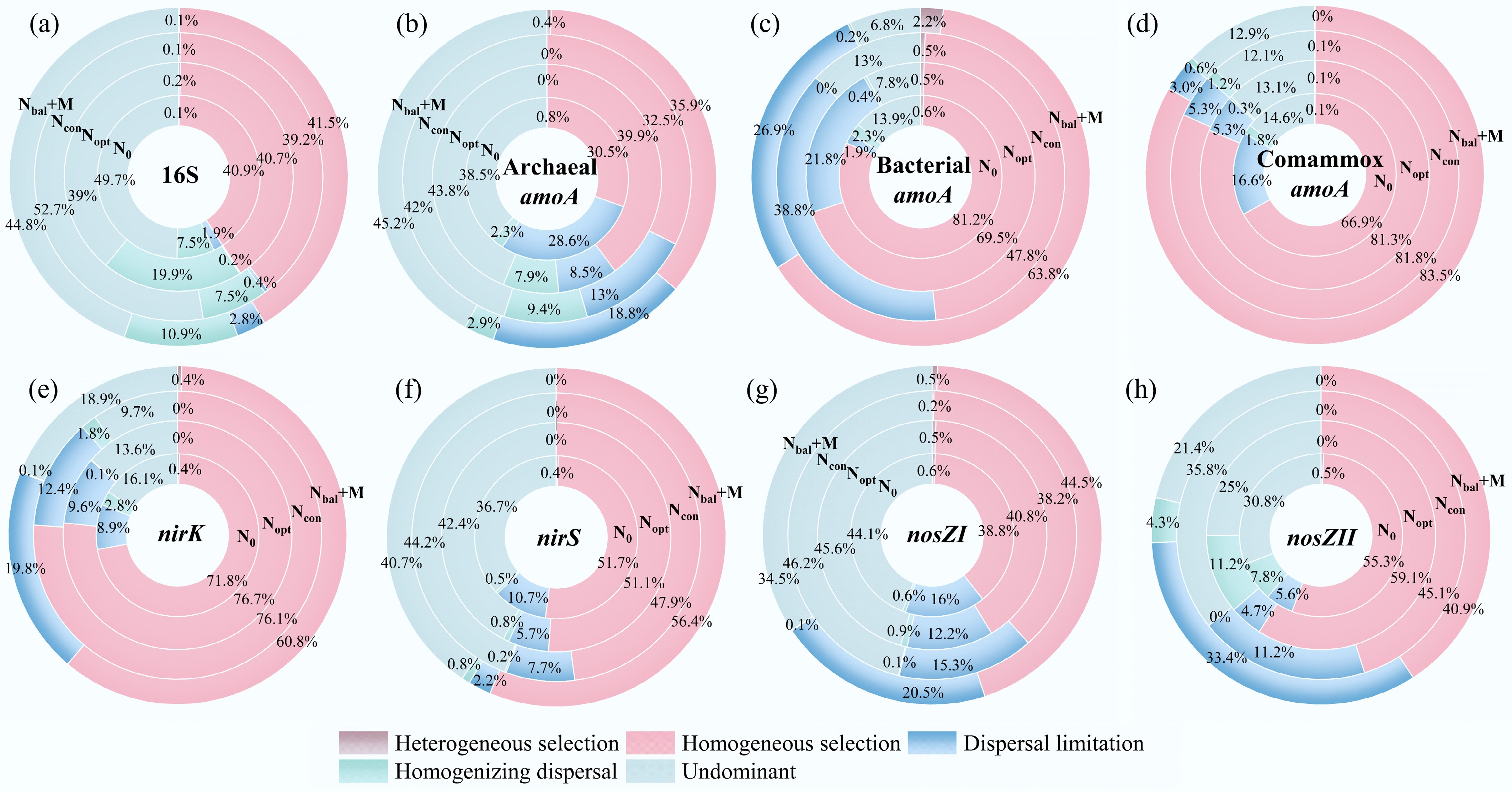
Figure 4.
Proportion of ecological processes for nitrogen functional microbial communities based on bootstrapping results (n = 1,000) from iCAMP. (a) 16S rDNA. (b) Archaeal amoA gene. (c) Bacterial amoA gene. (d) Comammox amoA gene. (e) nirK gene. (f) nirS gene. (g) nosZI gene. (h) nosZII gene. Undominant: drift and others.
For the total bacterial community (Fig. 4a), stochastic processes were the primary driver, accounting for over 50%, while the contribution of homogeneous selection was 39.2%–41.5%. Distinct assembly mechanisms were observed among the different ammonia-oxidizing guilds. The AOA (Fig. 4b) community assembly was dominated by stochastic processes (over 50%), with homogeneous selection accounting for 30.5%–39.9%. In contrast, the assembly of AOB and comammox communities was predominantly governed by deterministic processes. For the AOB community (Fig. 4c), homogeneous selection accounted for 47.8%–81.2% of the explained processes, with dispersal limitation also playing a notable role (1.9%–38.8%). The comammox community was almost entirely structured by homogeneous selection, which contributed a dominant 66.9%–85.5% to its assembly across all treatments (Fig. 4d). The denitrifier communities also displayed varied assembly patterns. The nirK-, nirS-, and nosZII-harboring communities were primarily shaped by deterministic processes. Homogeneous selection was the leading factor for these three guilds, accounting for 60.8%–76.7% for nirK (Fig. 4e), 41.9%–56.4% for nirS (Fig. 4f), and 40.9%–59.1% for nosZII (Fig. 4h). However, the assembly of the nosZI-harboring community (Fig. 4g) was driven by stochastic processes accounting for over 50%.
To further elucidate the influence of fertilization on community assembly, the relative contribution of stochastic processes (including dispersal limitation, homogenizing dispersal, and drift processes) and deterministic processes (heterogeneous and homogeneous selection) were quantified and compared across treatments for each microbial community (Fig. 5). Stochastic processes dominated in 16S DNA (Fig. 5a), AOA (Fig. 5b) and nosZI (Fig. 5g) communities, and deterministic processes dominated in AOB (Fig. 5c), comammox (Fig. 5d), nirK (Fig. 5e), nirS (Fig. 5f) and nosZII (Fig. 5h) communities.
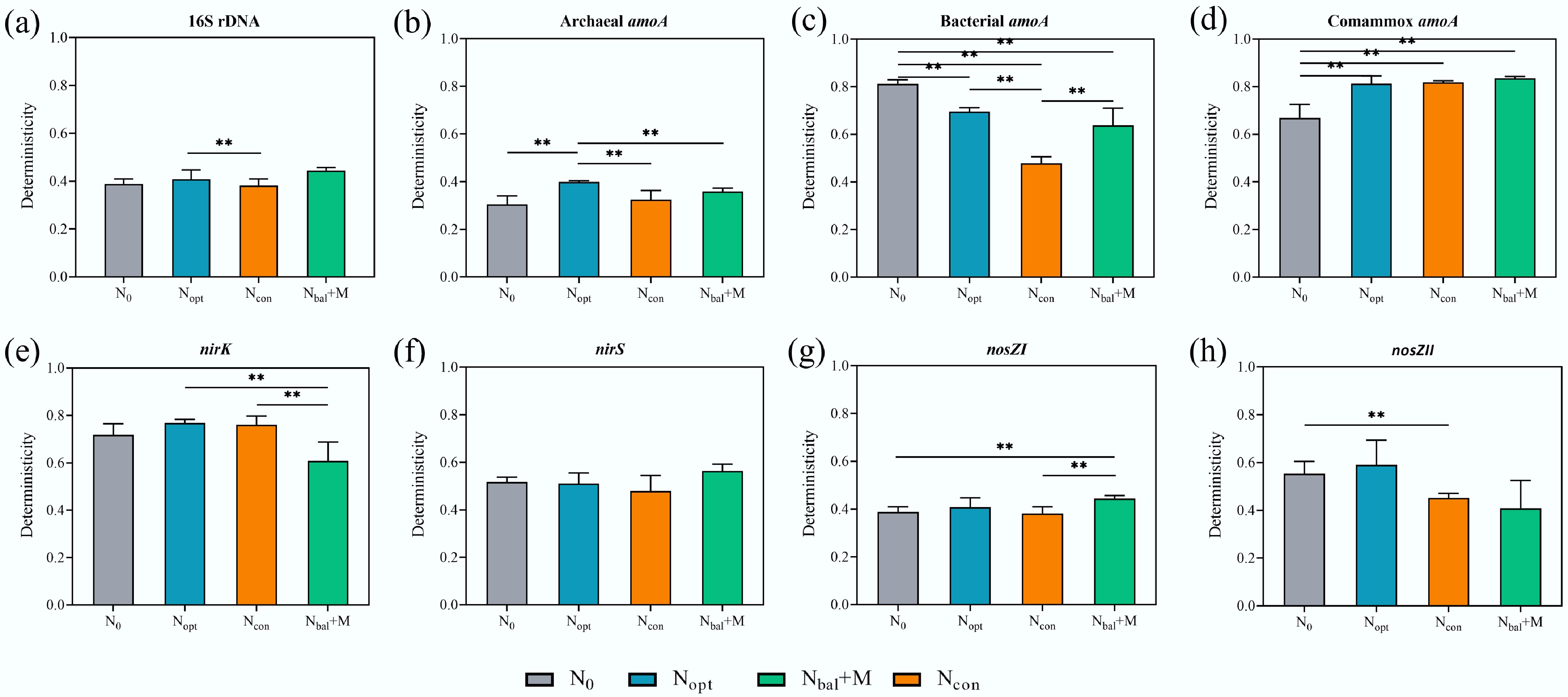
Figure 5.
Deterministicity (presented by homogeneous selection) portion comparison among different fertilizer treatments for nitrogen functional microbial communities based on iCAMP. (a) 16S rDNA. (b) Archaeal amoA gene. (c) Bacterial amoA gene. (d) Comammox amoA gene. (e) nirK gene. (f) nirS gene. (g) nosZI gene. (h) nosZII gene. ** p < 0.01, * p < 0.05. All comparisons show significant differences based on bootstrapping test (n = 1,000).
Deterministicity proportion was significantly different between Nopt and Ncon for the total bacterial community (p < 0.01; Fig. 5a). Fertilizer application had divergent and significant effects on the assembly processes of different ammonia-oxidizing guilds. For the deterministicity of the AOA community, the value in Nopt was significantly higher than in the other three treatments (p < 0.01; Fig. 5b). Conversely, for the AOB community (Fig. 5c), all fertilizer treatments significantly decreased the deterministicity proportion compared to the N0 control (p < 0.01). Ncon exhibited the lowest deterministicity (p < 0.01). For the comammox community (Fig. 5d), all fertilizer treatments (Nopt, Ncon, and Nbal + M) significantly increased the contribution of deterministicity compared to N0 (p < 0.01).
The assembly of denitrifier communities was also variably affected. For the nirK-harboring community, the Nbal + M had a significantly lower deterministic proportion than the Nopt and Ncon (p < 0.01; Fig. 5e). For the nosZI community, Nbal + M significantly increased stochasticity compared to N0 and Ncon (p < 0.01; Fig. 5g). For nosZII community, Ncon had a significantly higher determinsticity than N0 (p < 0.01; Fig. 5h). There was no significant difference in the determinsticity proportion observed across treatments for the nirS-harboring community.
-
The results revealed the distinct N cycling microbial mechanisms and ecological assembly processes that affected N transformations under different fertilizer regimes. The findings underscore that while conventional synthetic fertilizer application can sustain high yields, it does so at the cost of significant N2O emissions, driven by specific changes in the nitrifier community. In contrast, the manure-amended system achieves comparable productivity while actively mitigating these emissions by selectively enhancing the soil N2O reduction capacity.
Integrated nutrient management as a foundation for soil quality and yield productivity
-
The superiority of balanced manure with synthetic N (Nbal + M) treatment is reflected in its benefits for soil quality. The significant increases in SOC, TN, and Olsen P (Table 1) confirm the well-established role of organic amendments in improving long-term soil quality[12]. Unlike synthetic fertilizers that supply specific nutrients, manure provides a complex matrix of carbon compounds and a slow-release source of a wide array of nutrients. The resulting elevation of the soil quality index under Nbal + M (Fig. 1a) reflects this improvement. The enhanced soil quality translated into crop yields that were statistically similar to those under the highest-input synthetic fertilizer treatments Ncon and optimate fertilizer input Nopt (Fig. 1b). These results suggest that a portion of synthetic fertilizer can be substituted with organic amendments without compromising yield, at least in the context of this study system. The Nbal + M strategy thus represents a clear pathway toward sustainable intensification, in which productivity is maintained or enhanced while simultaneously enhancing soil quality.
Decoupling N cycling from N2O emissions
-
While all fertilizer treatments stimulated both potential nitrification rate (PNR) and potential denitrification rate (PDR), their ultimate impact on N2O flux varied, pointing to a critical shift in the balance between N2O-producing and N2O-reducing pathways. The high N2O flux observed under Ncon (Fig. 1c) is linked to AOB, whose abundance was highest in this treatment (Fig. 2c). Furthermore, the Mantel test revealed that PNR, PDR, and N2O emissions were significantly related to the comammox community structure (p < 0.05; Table 2). The unresponsiveness of the AOA to fertilization (Figs 2b and 3b), contrasted with the significantly varied AOB and comammox community structures among different treatments (p < 0.05; Fig. 3c, d), reinforces the niche differentiation among AOA, AOB, and comammox in dominating the high N nitrification process and its associated emissions[37].
Despite exhibiting the highest PDR, Nbal + M had significantly lower N2O emissions than Ncon (Fig. 1c). Nbal + M had significant increase in the abundance of nitrous oxide reductase (nosZ) genes (Fig. 2g, h). Furthermore, the Mantel test revealed that PNR and N2O emission were significantly related to the nosZ-harboring community structure, while PDR was related to the nosZII-harboring community (p < 0.05; Table 2). The nosZ-harboring community, known for its vast phylogenetic diversity and ecological versatility[6], was clearly stimulated by the manure application (Figs 2g, h, and 3g). The carbon and diverse nutrients in manure likely provided the necessary electron donors and cofactors to support a large and active population of these N2O-reducing heterotrophs[38]. This transforms the soil from a net source to a more efficient sink for N2O. Validated by the concept that managing agricultural systems to increase the nosZ / (nirS + nirK) gene ratio is a potent strategy for climate change mitigation[39], and the nosZII guild was identified in particular as the key biological lever for achieving this outcome in this study.
Long-term monitoring at our field site demonstrates consistent interannual patterns in N2O flux responses to different treatments, despite climate variability[18,40,41]. Furthermore, the N2O emission 'hot moments' are predominantly concentrated on the summer maize season, and the various treatment effects consistently manifested during this period across multiple years. This temporal stability suggests that the underlying microbial mechanisms governing N2O emissions remain relatively constant at this long-term experimental site. Consequently, by correlating 2022 soil data (reflecting long-term treatment effects on soil properties and microbial communities) with previous annual N2O emissions, the potential regulatory mechanisms of N-cycling microbes and N2O production could be explored. Although limited by single-time-point sampling, this approach effectively captures representative microbial drivers.
Deterministic assembly dominates N cycling function guilds
-
This study provides how deterministic ecological processes govern the assembly of N cycling functionally microbial guilds. While stochastic processes played a significant role in structuring the total bacterial community, the assembly of the most important players in nitrification (AOB, comammox) and denitrification (nirK, nosZII) was dominated by homogeneous selection (Fig. 4). This implies that their community structure, and thus their functional capacity, is not a random outcome but a predictable response to the environmental filters imposed by management, varied with the total bacterial community[7]. The assembly of AOA was dominated by stochastic processes, whereas that of AOB was governed by deterministic selection. AOB are classic copiotrophs, adapted to thrive in high-nutrient environments. Fertilization is a powerful and direct environmental filter that deterministically selects for specific, fast-growing AOB taxa[42], a conclusion supported by the significant increase in AOB abundance under the Ncon treatment (Fig. 2). In contrast, AOA are oligotrophs with a high-affinity ammonia oxidation system, which makes them superior competitors in low-N conditions. In the absence of a strong filter favoring their growth, the AOA community composition is more susceptible to stochastic processes like ecological drift and dispersal from the regional species pool[43].
Furthermore, N2O emission was significantly related to comammox, nirK, nosZI, and nosZII community structure (Table 2). Ncon treatment, which had the highest N2O emission, exhibited lower homogeneous selection in nosZI and nosZII communities, and higher homogeneous selection in comammox and nirK communities, along with significantly different homogeneous selection portions compared to Nbal + M in nirK and nosZI communities (Fig. 5). These findings indicate that the microbial assembly mechanisms underlying N2O emission involve increased homogeneous selection in N2O-producing communities and decreased homogeneous selection in N2O-reducing communities.
Moreover, the nosZI community was primarily shaped by stochastic processes, while the nosZII community was structured by deterministic selection. This can be attributed to their distinct phylogenetic diversity and ecological niches[6]. nosZII is known to be more phylogenetically diverse and ecologically versatile, found in a wider range of microorganisms (including non-denitrifiers) and habitats[44]. Crucially, many nosZII-containing organisms can reduce N2O under microaerobic conditions, which makes them highly adaptable to the fluctuating oxygen levels typical of agricultural topsoil. While the nosZI guild, often associated with 'canonical' denitrifiers in more strictly anaerobic niches, may find its activity limited to transient, spatially-patchy anaerobic microsites within the soil matrix[38]. Their assembly would therefore be less influenced by the overarching management strategy and more by unpredictable, localized conditions, appearing as a stochastic process at the scale of our investigation. This highlights that even within the same functional step, different clades can be governed by different ecological rules.
Limitations and future directions
-
Despite the mechanistic insights provided by this study, several limitations should be acknowledged to contextualize the findings and guide future work. First, the experiment was conducted at a single site within the North China Plain, characterized by a specific clay loam soil and an intensive winter wheat–maize cropping system. The magnitude of treatment effects on microbial community assembly and N2O fluxes is likely dependent on local soil properties and climate. Therefore, validating these findings across a broader range of agroecological zones is a critical next step to ascertain the universality of the observed mechanisms.
Second, although this study effectively demonstrates the potential of integrated manure management, the scalability of this practice faces practical and economic hurdles. The composition of manure varies significantly with animal source and storage method, which could alter its effect on the soil microbiome. Furthermore, widespread adoption depends on factors such as local manure availability, transportation costs, and the economic viability for farmers compared to synthetic fertilizers. Future research should integrate socioeconomic analysis and explore different types of organic amendments to develop strategies that are both ecologically sound and practically scalable. Finally, although the single-time-point microbial sampling was strategically timed to reflect long-term effects, it does not capture the intra-annual dynamics of the N-cycling communities. Future studies incorporating high-frequency temporal sampling would provide a more complete picture of how microbial assembly processes shift in response to fertilization, irrigation, and crop growth stages.
In conclusion, this investigation demonstrates that sustainable N management hinges on steering microbial ecological processes. A novel application of iCAMP to N-cycling functional guilds explains how N2O emissions are regulated by the balance of deterministic and stochastic forces shaping community assembly. Key guilds involved in nitrification (AOB, comammox) and denitrification (nirK, nosZII) were primarily structured by homogeneous selection, implying that their composition is a predictable response to environmental filters imposed by fertilizer management. The high N2O emission under conventional fertilization corresponds to stronger deterministic control over N2O-producing communities, making high emissions a predictable outcome. In contrast, integrated manure application mitigates N2O by shifting this balance: it weakens homogeneous selection on N2O-producers while strengthening it for N2O-reducers. This approach moves beyond simply supplying nutrients to actively curating a soil microbiome that is more efficient at N2O reduction. This study provides a mechanistic roadmap for this ecological engineering, demonstrating that by managing the underlying assembly processes of the N-cycle, the critical goal of high-yield agriculture can be achieved with a smaller environmental footprint.
-
It accompanies this paper at: https://doi.org/10.48130/nc-0025-0007.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: study conception and design, draft manuscript preparation: all authors; original research plan, all site experiments setup: Ju X; sampling implementation, the soil experiments, sequencing, and data analysis: Wang Z, Li Y. All authors reviewed the results and approved the final version of the manuscript.
-
The sequencing data are available in National Microbiology Data Center (https://nmdc.cn/resource/genomics/project) with project number NMDC10019933.
-
This work was supported by the National Natural Science Foundation of China (Grant No. U24A20625).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Zhujun Wang, Yue Li
Full list of author information is available at the end of the article. - Supplementary File 1 Supplementary materials and methods to this study.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Z, Li Y, Liu X, Ju X. 2025. Integrated manure application enhances soil quality and reduces nitrous oxide emissions by deterministically shaping N cycling guilds. Nitrogen Cycling 1: e007 doi: 10.48130/nc-0025-0007 

Integrated manure application enhances soil quality and reduces nitrous oxide emissions by deterministically shaping N cycling guilds
- Received: 28 July 2025
- Revised: 02 September 2025
- Accepted: 19 September 2025
- Published online: 17 October 2025
Abstract: A novel application of Infer Community Assembly Mechanisms (iCAMP) reveals how distinct ecological processes governing nitrogen (N) cycling guilds mediate agricultural nitrous oxide (N2O) emissions. In a long-term field experiment, the guild-specific explanation for how different N fertilizer strategies alter N2O emissions is provided. No N (N0), conventional N (Ncon), optimal N (Nopt), and balanced manure with synthetic N (Nbal + M) were also compared. The Nbal + M treatment significantly enhanced soil quality and maintained high crop yields comparable to the Ncon treatment. Crucially, Nbal + M achieved a lower N2O emission than Ncon, a reduction linked to increased abundance of nosZ genes. The novel application of iCAMP revealed that this outcome was governed by fundamental shifts in ecological selection. The high N2O emissions under Ncon were associated with increased homogeneous selection on N2O-producers, while the low emissions under Nbal + M were driven by decreased homogeneous selection of N2O-producers and increased homogeneous selection over N2O-reducers. This study demonstrates that integrated manure application decouples high crop yield from N2O emissions by predictably shaping the soil microbiome to enhance its N2O reduction capacity, providing a clear, mechanism-based pathway for developing sustainable agriculture.