









-
Nitrogen (N) is an essential element for all life forms[1,2]. However, since the 1950s, excessive N (primarily in inorganic and bioavailable forms, with NO3–-N being the dominant inorganic N) from intensive agriculture and industrial activities has accumulated in rivers, leading to global environmental issues such as eutrophication, algal blooms, and biodiversity loss[3,4]. Denitrification (autotrophic denitrification and heterotrophic denitrification) and anaerobic ammonium oxidation (anammox) are recognized as the key net N removal processes that mitigate excessive N loadings in riverine ecosystems[5]. Consequently, a comprehensive understanding of the mechanisms governing N removal in river ecosystems is critical for developing effective watershed management strategies and advancing knowledge of the global riverine N cycle.
Rivers receive substantial N inputs and function as critical filters, converting approximately 40% of terrestrial N runoff (ca. 47 Tg N yr−1) into dinitrogen gas (N2)[6,7]. Denitrification is widely acknowledged as the dominant pathway for eliminating reactive N in riverine sediments[8−10], accounting for 80.4% ± 0.4% of global sedimentary N removal[4]. In comparison, anammox is an alternative pathway that utilizes nitrite (NO2−) as an electron acceptor to oxidize ammonium (NH4+) to N2 under anoxic conditions[11,12]. The relative contribution of anammox (1%–20%) varies considerably at the local scale, yet it plays a more prominent role in aquatic ecosystems than in terrestrial ones globally[4]. Although previous studies have focused on quantifying N removal rates and contributions across diverse ecosystems[7,13−15], a systematic understanding of how abiotic and biotic factors synergistically drive these processes remains elusive across temporal and spatial scales.
Previous studies have demonstrated that denitrification and anammox in river sediments are closely governed by micro-environmental abiotic and biotic factors[10,16,17]. Substrate concentrations (NH4+/NO2−/NO3−), moisture, temperature, organic carbon availability, and C/N ratio are the key factors influencing N removal and the competition and synergy between denitrification and anammox in river sediments[7,8,18]. Furthermore, the N removal processes are largely mediated by functional microorganisms[12]. For instance, functional genes such as nosZ, nirS, and nirK are key markers for denitrification[7,10], whereas hzsA and hzsB act as diagnostic markers for anammox bacteria[19]. Concurrently, these physicochemical factors also play a crucial role in shaping the biogeographic distribution of microbial communities and their metabolic activities[20]. For instance, previous studies have demonstrated that microbial and enzymatic activities are tightly constrained by temperature and moisture conditions[10,21].
Furthermore, sediment physicochemical parameters in river systems are fundamentally governed by macroscale factors such as altitude, climate, and land use[7,22]. Altitude, for instance, indirectly regulates sediment environments by influencing temperature regimes, precipitation patterns, and land-cover distribution. Climatic factors such as precipitation and temperature modulate the transport and deposition of nutrients from soils to river networks through soil leaching and surface runoff, as well as the transformation of N species in the catchment[2,23]. Recent studies have shown that land use patterns not only determine the nutrient loading and physical environments, but also influence the composition of sediment microbial communities involved in N cycling[24−26]. For example, anthropogenic land-use types (such as agricultural and urbanized land) can result in simple microbial community structures and stimulate microbe-mediated denitrification in rivers[26]. While existing studies have examined the roles of geographical (e.g., altitude and land uses) or micro-environmental drivers (e.g., NH4+, NO2−, and NO3−) in regulating N removal mechanisms in rivers[27,28], the lack of an integrated cross-scale perspective prevented a systematic understanding of riverine N removal dynamics and their implications for riverine N budgets. Therefore, we hypothesized that both geographical factors (e.g., land use types) and micro-environmental biotic and abiotic factors jointly exert significant effects on N removal processes[29].
In this study, we analyzed a total of 36 sediment, and 36 water samples (n = 18 per season) from the Jinshui River, a forested tributary in central China, during summer and winter. Remote sensing,15N isotope labeling, and qPCR were integrated to systematically investigate N removal processes in the river, focusing on drivers at both macroscopic (e.g., altitude, climate, and land-use type) and micro-environmental scales (e.g., water properties, sediment properties, and functional gene abundances). The objectives of this study were to: (1) quantify the relative contributions of denitrification and anammox to total NO3− removal in summer and winter; and (2) elucidate the respective roles and interactions of macroscale and microscale factors in regulating N removal rates. By integrating interdisciplinary and multi-scale perspectives, this study advances mechanistic understanding of seasonal N removal in river ecosystems and offers a conceptual framework for explaining spatial heterogeneity in riverine nitrogen cycling.
-
The Jinshui River (33°16' N–33°45' N, 107°40' E–108°10' E), located in central China (Fig. 1), spans an altitude range of 416–2,904 m and drains an area of 730 km2. This catchment lies in a subtropical monsoon zone, where the mean yearly temperature ranges from 12 to 15 °C[30]. Surface runoff within the watershed is primarily driven by precipitation. According to the rainfall characteristics, the river experiences distinct wet (July to October), dry (December to March), and normal (April, May, and November) seasons.
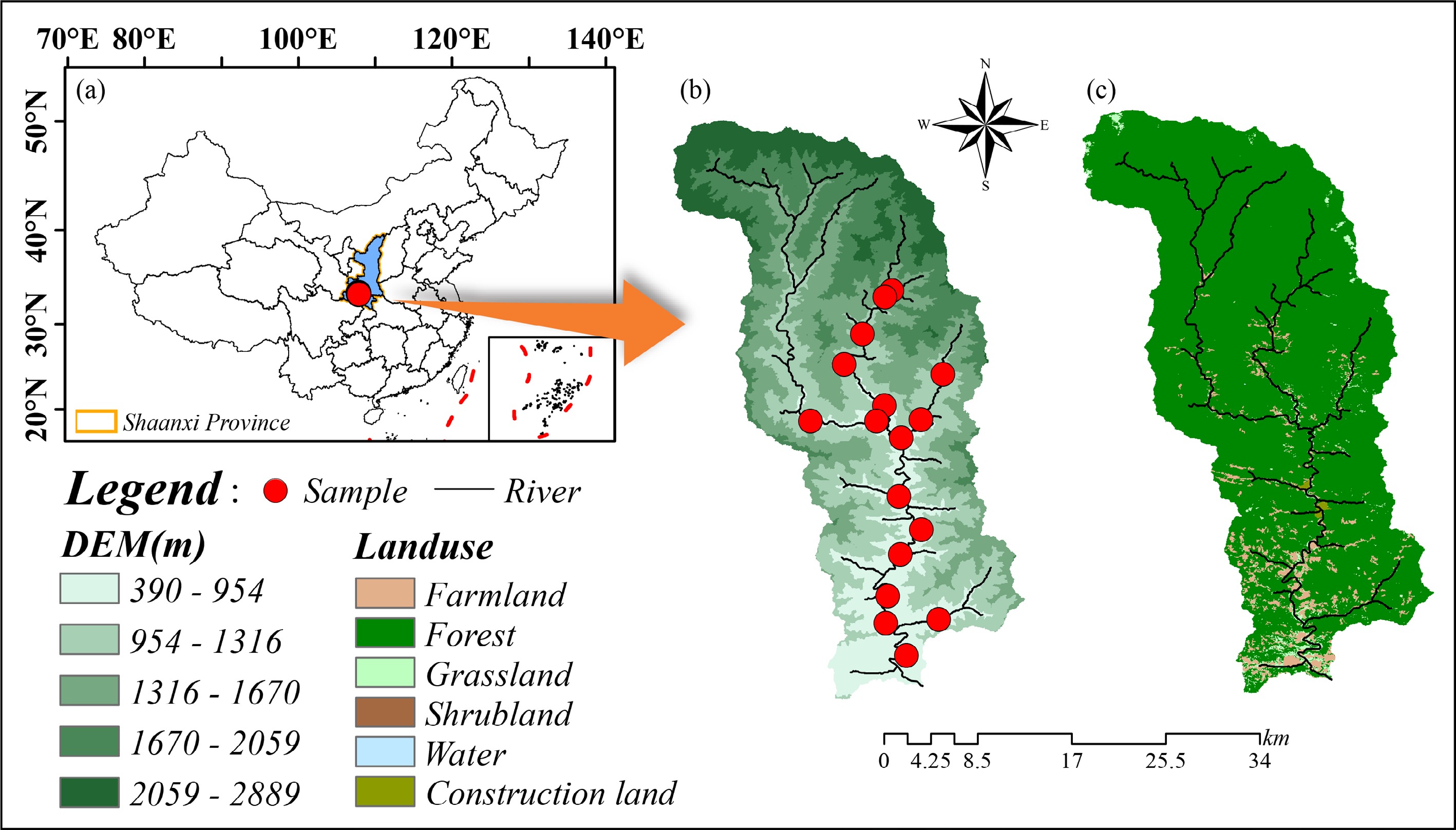
Figure 1.
Locations of the study sites across the Jinshui River. (a) Location of the catchment; (b) the topography of the catchment and the distribution of the sampling sites; (c) the land use types of the catchment.
The catchment is characterized by low anthropogenic disturbance, particularly in the upstream regions (JS1–10), and maintains a forest coverage exceeding 95% (Supplementary Table S1)[30]. Influenced by altitude, the vegetation within the watershed exhibits a pronounced vertical distribution pattern. Nitrogen-fixing tree species are sparsely distributed throughout the catchment. Granite constitutes the dominant bedrock, and the overlying thin topsoil exhibits high susceptibility to erosion. The catchment is devoid of industrial activities, and agriculture consists primarily of small-scale farming concentrated in downstream areas (JS11–18), with maize and rice as the dominant crops. Urea serves as the primary fertilizer, with a typical application rate of approximately 800 kg N hm−2[30].
Sample collection
-
Field sampling campaigns were conducted in July (summer) and December (winter) 2021. A total of 18 sampling sites were selected along the Jinshui River (Fig. 1), with selection based on site accessibility, channel morphology, and the gradient of anthropogenic disturbance.
Paired water and surface sediment samples were collected at each site. Surface water samples were obtained at depths ranging from approximately 10 to 20 cm using pre-cleaned polyethylene water bags. These samples were immediately filtered through fiberglass membranes (Whatman GF/F, 0.7 µm pore size) and transferred into 50 mL polyethylene bottles. Sediment samples (0–10 cm depth) were collected from the mid-channel after removal of aquatic macrophytes and coarse organic debris[30]. Within a 5 m × 5 m area at each site, multiple subsamples were collected and thoroughly mixed to form a composite sample. Water samples and aliquots of the sediment samples were immediately frozen for subsequent analysis of physicochemical parameters. Additionally, separate sediment samples were stored in liquid nitrogen prior to N transformation analysis and qPCR assays[31].
Measurements of environmental factors
-
Water physicochemical parameters, including pH, temperature (T), dissolved oxygen (DO), and oxidation-reduction potential (ORP), were measured in situ using a calibrated YSI EXO3 multi-parameter sonde. Dissolved inorganic N species (NO2−, NO3−, and NH4+) were quantified using ion chromatography (ICS-1100, Dionex), with measurement precision maintained within ± 5%.
Sediment physicochemical characteristics were determined using standard protocols. Sediment NH4+ and NO3− were extracted from fresh sediments with a 2 M KCl solution, and their concentrations were subsequently determined using a UV–1800 UV spectrophotometer (Shimadzu, Kyoto, Japan) with a detection limit of 0.01 mg N L−1 in the laboratory[32]. Total nitrogen (TN) and total organic carbon (TOC) were determined with a Vario EL cube elemental analyzer, while total phosphorus (TP) was determined by digesting samples with a persulfate-sulfuric acid solution and colorimetrically measuring the resulting orthophosphate[33]. Sediment moisture content was determined gravimetrically by measuring the mass loss after drying fresh sediments at 105 °C for 24 h[34]. For quality assurance and quality control, all measurements were performed in triplicate, with the analytical error maintained below 5%.
Geographic coordinates and altitude were recorded at each site using a handheld GPS unit (Rino 650, Garmin). Land use types within the catchment were classified and mapped using ArcMap 10.8, based on the 30-m resolution GlobalLand30 dataset (2021). Climatic data with a spatial resolution of 1.0 km were retrieved from the WorldClim database (
www.worldclim.org ).Measurements of N transformation rates and gene abundances
-
Potential rates of denitrification, anammox, and dissimilatory NO3− reduction to ammonium (DNRA) in the sediments were measured using 15N isotope pairing techniques (Supplementary Text S1)[13]. Net nitrification rates were determined using the aerobic incubation method (Supplementary Text S1)[35]. While nitrification and DNRA does not remove N, the rates were also determined to illustrate how the coupling between N transformation processes influences denitrification and anammox. Quantification of functional gene abundances was conducted using qPCR, targeting genes associated with the key N cycling processes, including amoA-AOA, amoA-AOB, nirK, nirS, nosZ, hzsA, and nrfA (Supplementary Text S2). Detailed experimental protocols are provided in the Supplementary Information, and in our recently published studies[30,36].
Data analyses
-
In this study, statistical analyses and data visualization were performed using OriginPro 2025 and R software (v4.3.2). Data normality was assessed with the Shapiro-Wilk test. The non-parametric Kruskal–Wallis test was employed to evaluate the spatiotemporal differences in N transformation rates. Spearman's rank correlation analysis was conducted to examine the correlations among N-cycling genes, N transformation rates, and environmental factors. Partial least squares path modeling (PLS-PM) was conducted to assess the direct and indirect effects of N-cycling genes, physicochemical properties (water and sediment), land use, and climate on denitrification and anammox rates. The model estimated path coefficients (indicating the direction and strength of linear relationships between the variables) and explained variance (R2). The analysis was implemented using the 'plspm' package in R, with overall model performance evaluated using the Goodness of Fit (GoF) index.
-
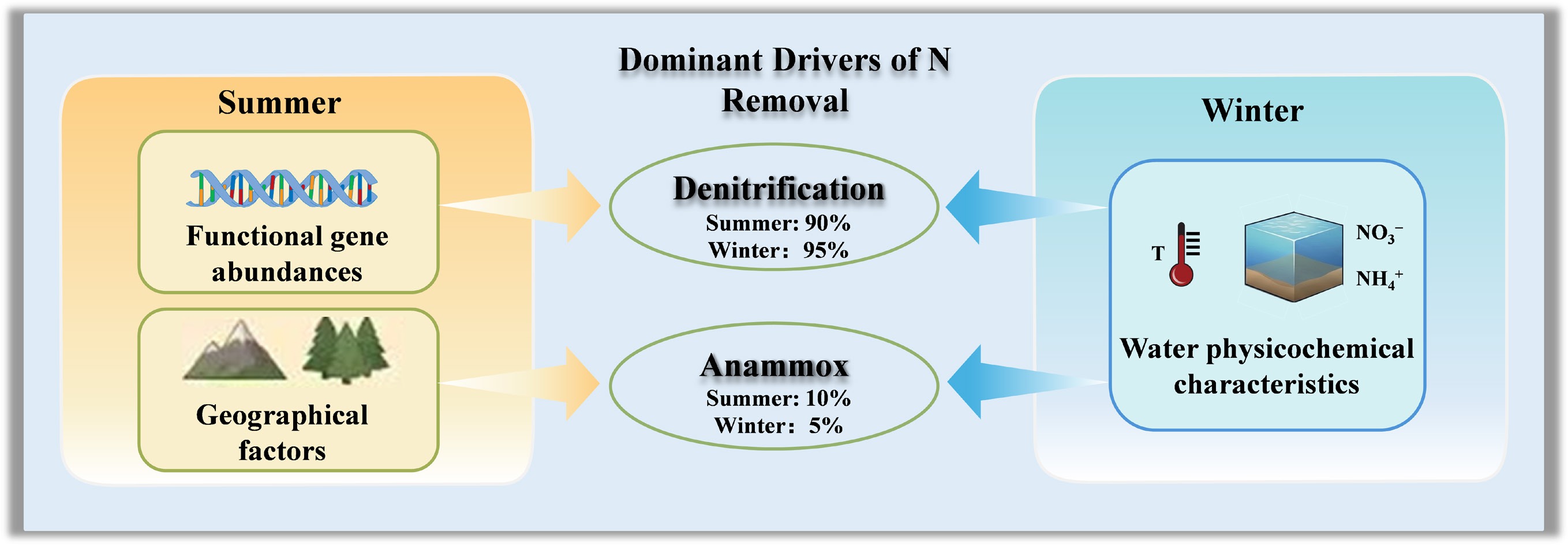
In summer, the mean denitrification rate (mean ± SD, 1.68 ± 2.22 mg N kg−1 d−1) in sediments was significantly higher than that of anammox (0.21 ± 0.23 mg N kg−1 d−1) (Fig. 2a). In winter, denitrification and anammox rates declined to 0.67 ± 1.52 mg N kg−1 d−1 and 0.07 ± 0.12 mg N kg−1 d−1, respectively (Fig. 2a). Denitrification rates decreased from summer to winter, whereas anammox rates exhibited relatively limited seasonal variation (Fig. 2a). This distinct thermal response suggests that anammox bacteria demonstrate greater resilience to low temperatures than denitrifiers[4,12]. The summer temperatures (~19.5 °C) were closer to the reported optimal range for denitrification activity (25–37 °C), thereby promoting its microbial activity[10]. In contrast, the low temperature in winter may reduce organic matter mineralization and electron donor availability[37], consequently exerting a more pronounced suppression on heterotrophic denitrification than on the anammox process[38]. In both seasons, denitrification-dominated biological NO3− reduction accounted for more than 90% of total N removal (Fig. 2b), which is comparable to reported global estimates for sedimentary N removal[4].
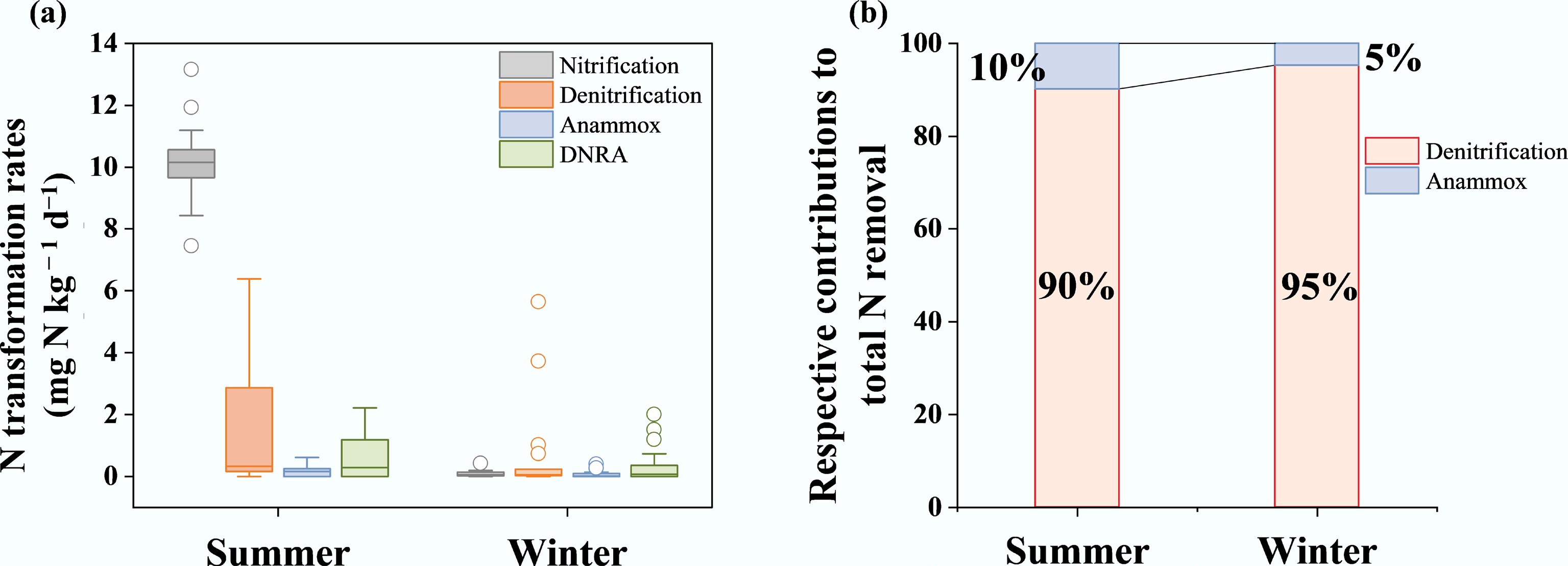
Figure 2.
(a) A box plot displaying the nitrification, denitrification, anammox, and DNRA rates of the sediments in summer and winter. The boxes and whiskers indicate the 5th, 25th, 50th, 75th, and 95th percentiles. The circles are outliers. (b) A bar chart showing the respective contributions of denitrification and anammox to total N removal. The relative contributions of denitrification and anammox were calculated based on the average rates during each season.
The mean net nitrification rate in summer (10.17 ± 1.28 mg N kg−1 d−1) was the highest among the N transformation rates for two seasons (p < 0.05; Fig. 2a), but declined sharply to 0.10 ± 0.10 mg N kg−1 d−1 in winter (Fig. 2a). The higher nitrification rate in summer suggests that vigorous nitrification likely provided abundant substrates (NO3–) to fuel coupled nitrification-denitrification[3]. The DNRA rate in summer (0.62 ± 0.74 mg N kg−1 d−1) was higher than in winter (0.37 ± 0.60 mg N kg−1 d−1), but remained lower than the global ecosystem average[22].
The linkages between N removal processes and potential factors
-
N removal rates exhibited significant seasonal heterogeneity (Fig. 2a), generally underscoring the broad metabolic constraints imposed by temperature. However, denitrification showed no significant correlation with altitude (an effective proxy for temperature) (p > 0.05; Fig. 3a), indicating that temperature was not the sole regulator. Meanwhile, anammox rates were positively correlated with altitude during summer (p < 0.05; Fig. 3), despite the inverse relationship between altitude and temperature (p < 0.001; Supplementary Fig. S1). These observations suggest that the direct impact of temperature on N removal may be confounded or modulated by other regulating mechanisms, such as variations in substrate availability and microorganisms.
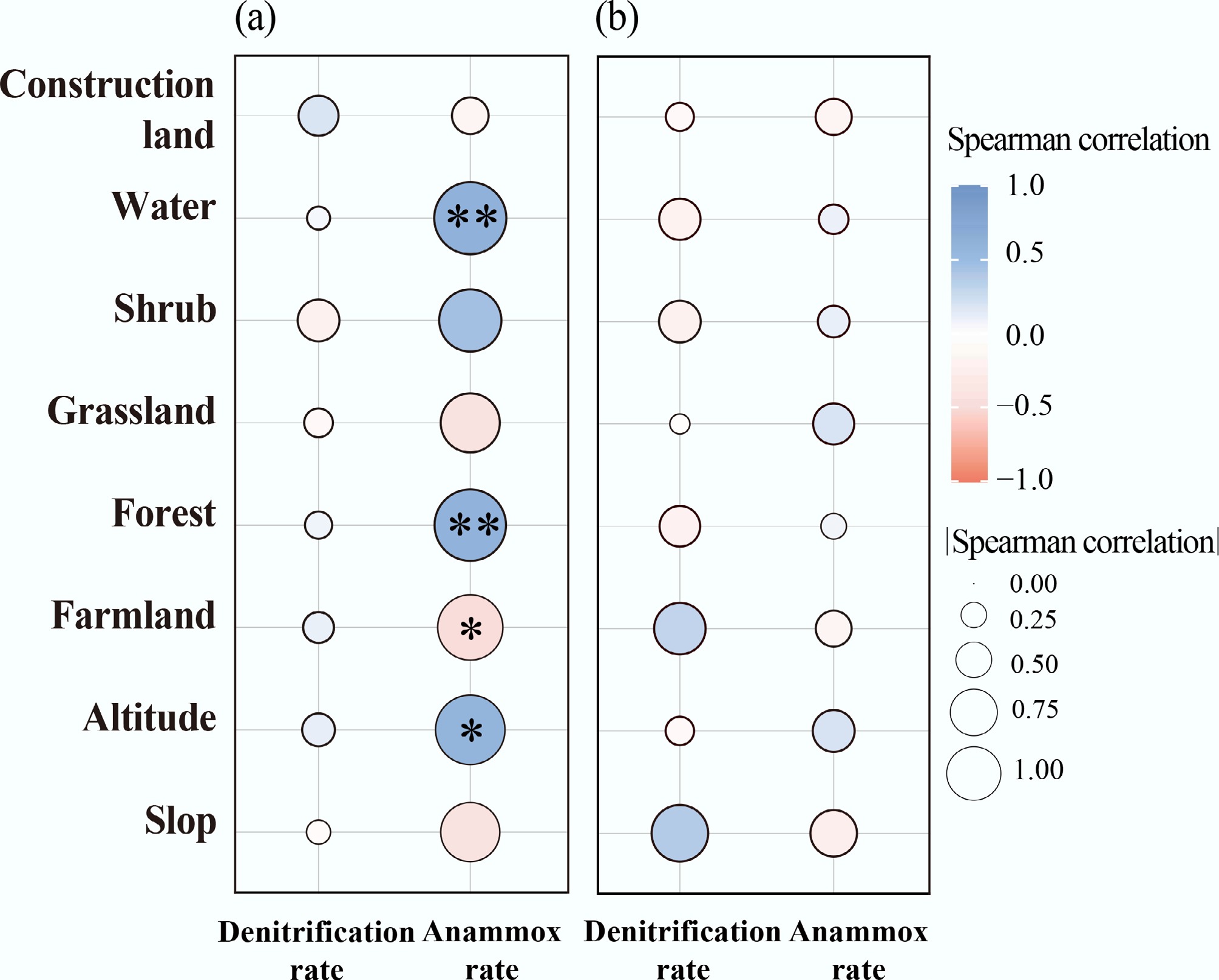
Figure 3.
Correlations of denitrification and anammox rates with geographic factors during (a) summer and (b) winter. * p < 0.05; ** p < 0.01.
Notably, altitude acts as a key determinant of land use patterns within this catchment (p < 0.05; Supplementary Fig. S2). Shifts in land use are typically accompanied by alterations in substrate characteristics, physicochemical properties, and microbial community structures[39,40]. In summer, increased forest cover was positively correlated with anammox rates (p < 0.05; Fig. 3), and this spatial preference might be driven by the intense substrate competition between anammox and denitrification[12,19]. While anammox and denitrification often occur under similar redox conditions, denitrifying bacteria typically exhibit faster growth and stronger competitiveness under nitrogen- and carbon-limited environments[41]. Previous studies have shown that abundant organic carbon generally promotes denitrification, whereas the relative contribution of anammox to N loss decreases with increasing organic carbon availability[41]. In forest-dominated watersheds, the typically lower organic carbon availability and nutrient inputs severely constrain the growth of denitrifiers[42]. This carbon limitation diminishes their competitive suppression over nitrogen oxides (e.g., NO2− and NO3−), thereby creating a more favorable environment for anammox processes. Conversely, fertilization and sewage discharge in the downstream regions led to higher nutrient availability in the river, potentially inhibiting sedimentary anammox (Fig. 3)[1]. Furthermore, regarding the interaction between denitrification and anammox, the warmer downstream environment may intensify the competitive suppression of anammox by denitrifiers, thereby diminishing anammox activity while promoting denitrification[10].
Climate and land use are likely to interact, and thus have integrated effects on N removal in local ecosystems[43]. In this study, although most geographic factors exhibited no direct relationship with N removal processes in the river (except for anammox in summer), they may still indirectly influence denitrification by shaping the micro-environment (Supplementary Fig. S2). In summer, the macroscale factors (altitude, slope, and land use) were significantly correlated with the physicochemical properties of water (e.g., temperature, pH) and sediment (e.g., NH4+, NO3−, TN), as well as key denitrification genes (nirS, nirK, and nosZII) (p < 0.05; Supplementary Fig. S2). Altitude and slope regulate hydrological processes and sediment transport, while land use determines nutrient inputs and organic matter supply to river systems[41,44]. Such changes in physicochemical conditions alter substrate availability and redox environments, thereby influencing the abundance and activity of denitrifying microorganisms[22]. Consequently, these micro-environmental factors act as proximal drivers regulating denitrification and overall N removal processes in river sediments.
During the warm season, denitrification was primarily driven by the abundances of nirS and nosZII genes, TN, TOC, C : N ratios, and sediment moisture (p < 0.05; Fig. 4a). Specifically, TN provides abundant substrates for denitrification, and TOC serves as an essential electron donor and energy source for denitrifiers[10]. Furthermore, the anaerobic conditions fostered by high sediment moisture were conducive to denitrification[45]. In contrast, no significant correlation was observed between microscale nutrient parameters and anammox rates during summer (p > 0.05; Fig. 4a). This decoupling likely reflects competitive suppression of anammox under carbon-rich and warm conditions. Elevated C : N ratios and high temperatures can stimulate heterotrophic denitrification, accelerating the consumption of NO2− and NO3− and thereby limiting the availability of NO2− required for anammox metabolism[10,18]. As a result, anammox activity becomes less responsive to local nutrient variability, despite the presence of suitable substrates.
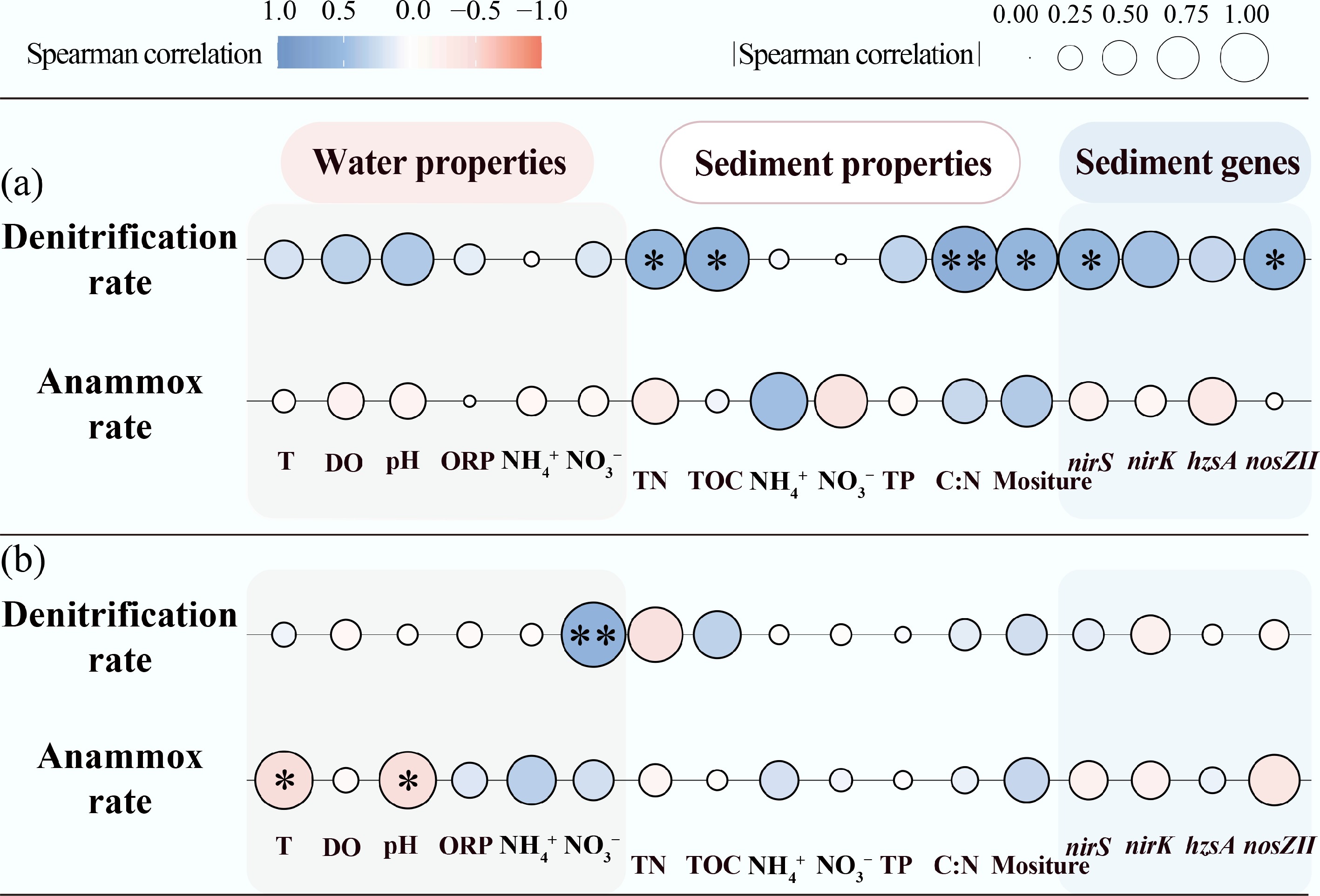
Figure 4.
Correlation of water properties, sediment properties, and abundance of key genes with denitrification and anammox: (a) summer; (b) winter. * p < 0.05; ** p < 0.01.
The regulatory drivers of N removal processes exhibited distinct patterns in winter (Fig. 4b; Supplementary Fig. S2). During this season, the supply NO3−-N from overlying water emerged as the primary direct driver of sediment denitrification (p < 0.05; Fig. 4b). Low temperatures likely suppressed organic matter decomposition and N mineralization[11], thereby reducing the availability of NO3− derived from internal sedimentary recycling. Concurrently, prolonged water residence times facilitated the diffusion of NO3− from the overlying water into surface sediments, serving as a major substrate source for denitrifying bacteria[11,46]. Anammox was also predominantly regulated by water physicochemical properties, particularly temperature and pH (p < 0.05; Fig. 4b). Lower temperatures are known to suppress the growth, metabolic activity, and enzymatic function of anammox bacteria[7,10]. Furthermore, pH is a critical factor driving spatial variation in biogeochemical N cycling[15], as it directly affects bacterial growth and enzyme activity[47,48]. Additionally, pH variations may modulate anammox rates by affecting the availability of NO2− and NH4+[31,48].
In summary, the rates and relative contributions of denitrification and anammox were governed by a complex interplay of direct and indirect geographic and micro-environmental drivers. To fully elucidate the mechanisms driving riverine N removal, it is essential to quantify the roles of these microscale factors (water, sediment, and microbial communities) across varying geographic conditions.
The roles of geographic and micro-environmental factors in regulating N removal processes
-
PLS-PM was employed to evaluate the direct and indirect effects of geographic and micro-environmental factors on N removal processes[4,49]. We assumed that micro-environment factors (e.g., microbial abundance and physicochemical parameters) directly influenced the N removal process in the river, and were inevitably regulated by the geographical factors. Furthermore, the geographical factors themselves also had a direct impact on N removal. In summer, the PLS-PM revealed that geographic factors (altitude, farmland, grassland, and forest) and micro-environmental variables (water properties, sediment properties, and sediment-associated functional genes) collectively explained 67% of the total variation in denitrification rates (Fig. 5a, b). Specifically, water physicochemical properties (DO, pH) and functional gene abundance (nirS/nirK/nosZ) were identified as the most significant drivers of denitrification (Fig. 5b). Furthermore, sediment properties (TOC, TN, C : N ratio, and moisture) and water properties exerted indirect effects on denitrification rates by regulating the abundance of N-transformation genes (Fig. 5a)[50].
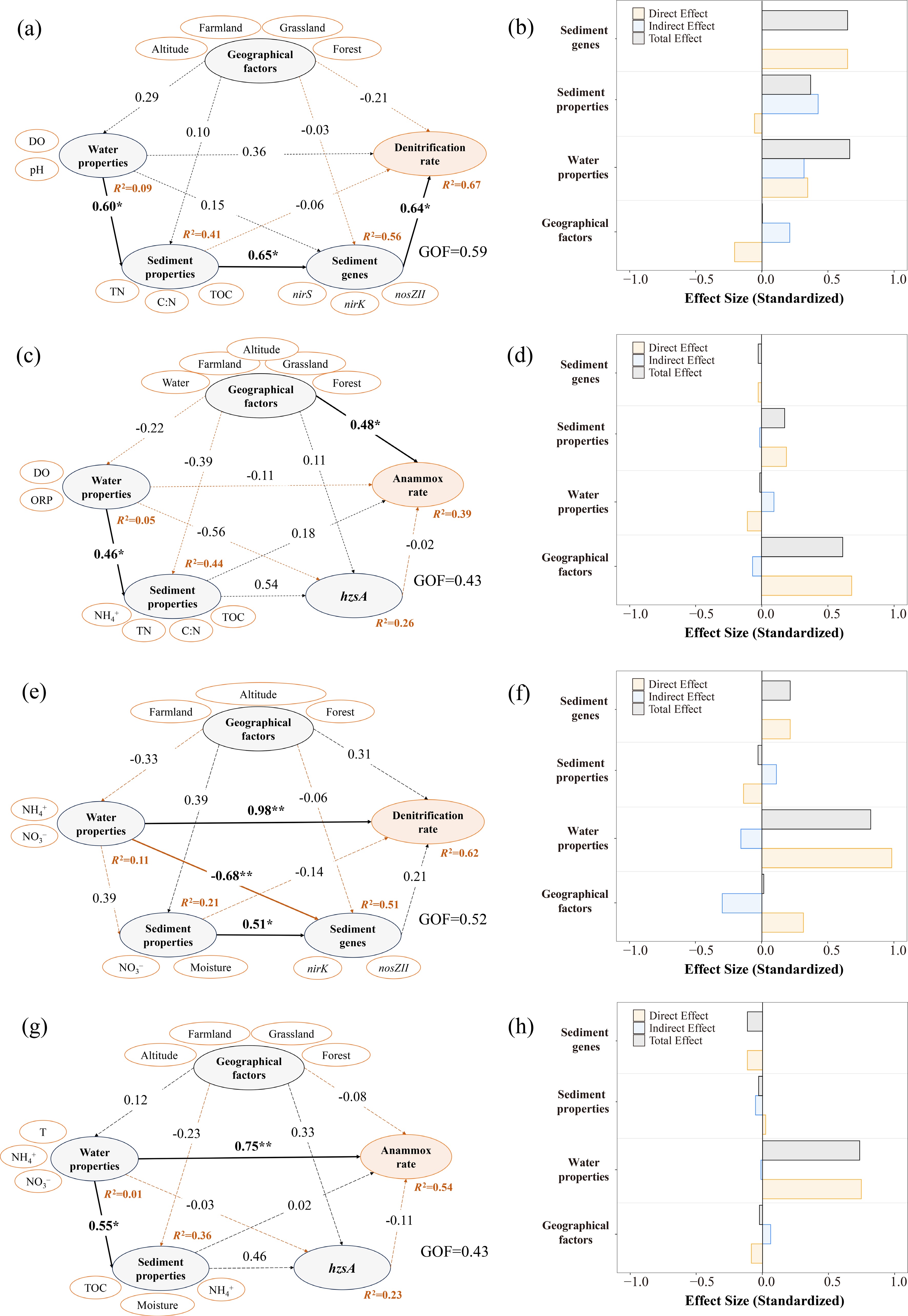
Figure 5.
PLS-PM demonstrates the impacts of geographic and micro-environmental factors on the denitrification and anammox rates in (a)–(d) summer, and (e)–(h) winter. Bold lines indicate significant effects, and dashed lines indicate non-significant effects. Positive and negative pathways are depicted with black and orange lines, respectively. Numbers adjacent to arrows are standardized path coefficients (* p < 0.05; ** p < 0.01; *** p < 0.001). The bar charts represented the normalized direct, indirect, and total effects of factors obtained from PLS-PM.
In contrast, the geographic and micro-environmental factors explained 39% of the total variation in the anammox process during summer (Fig. 5c, d). Geographic factors, particularly altitude and land use, exerted the strongest direct influence on the anammox process (Fig. 5d; Supplementary Fig. S2a). Furthermore, geographic factors also primarily influenced physicochemical properties (e.g., W-T, W-pH, S-TN, S-NH4+, and S-TP), thereby indirectly regulating anammox activity (p < 0.05; Supplementary Fig. S2a). The influence of water properties, sediment properties, and gene abundance on the anammox process was relatively weak (Fig. 5d). Notably, water physicochemical properties (DO and ORP) significantly influenced sediment chemical characteristics (Fig. 5c). The abundance of hzsA was decoupled from anammox rates (Fig. 5c), suggesting that microbial abundance did not appear to be the primary limiting factor for this process, consistent with findings reported in previous studies[15,20].
In winter, the PLS-PM revealed that geographic and micro-environmental factors jointly explained 62% of the total variation in denitrification (Fig. 5e), and 54% in anammox rates (Fig. 5g). Water physicochemical properties (temperature, NH4+, and NO3−) were strongly associated with both denitrification and anammox rates (Fig. 5e, g). Low temperatures significantly constrained sedimentary denitrification rates[51]. Meanwhile, prolonged water residence times and increased inputs of municipal sewage transformed the overlying water into a substantial N source for sediments, highlighting its critical role in winter N removal[30]. Low winter temperatures suppressed the abundance and expression of functional genes (nirK, nirS, nosZ, and hzsA) in sediments[11,50], leading to the decoupling between transformation rates and the gene abundances (Fig. 5e, g)[30,36]. Consequently, micro-environmental factors (primarily water properties) emerged as the dominant regulators of N removal processes during winter. In contrast, the pathways linking geographic factors to micro-environmental variables were not statistically significant (Fig. 5e, f). In winter, lower temperatures and reduced precipitation weakened hydrologic connectivity along the terrestrial–fluvial continuum[2,52], resulting in a functional decoupling between geographic drivers and the riverine micro-environment.
In summary, geographic factors exhibited regulatory effects on denitrification and anammox during summer, with a particularly strong direct influence on anammox (Fig. 5d). Simultaneously, higher summer temperatures exerted a strong moderating influence on internal biogeochemical conditions, such as by enhancing microbial activity. Conversely, reduced precipitation, lower temperatures, and weakened hydrologic connectivity in winter attenuated the direct impacts of geographic features on riverine N removal processes (Fig. 5d, h). Under conditions of weak internal turnover and long water residence time, the nutrient-replete overlying water served as a critical substrate source for sediments, thereby serving as the key regulator of winter N removal.
Interactions between N removal and transformation processes
-
While PLS-PM elucidated the direct and indirect linkages between geographical and micro-environmental factors and riverine N removal, the efficiency of N removal is also strongly mediated by interactions among multiple N-cycling pathways[19]. Within sedimentary environments, nitrification, denitrification, anammox, and DNRA do not operate in isolation; rather, they are inextricably coupled and collectively regulate N transformation and fate through exchanging intermediates (e.g., NO2−), modulating electron donor/acceptor availability, and competing within shared micro-environments[53,54]. For instance, while nitrification does not directly contribute to net N loss, it functions as an internal N transformation pathway that regulates substrate availability (e.g., NO2– and NO3–) for denitrification[17]. DNRA could also provide NH4+ for anammox[55].
Nitrite (NO2−) acts as a pivotal intermediate in nitrification, denitrification, and DNRA, while serving as an obligatory substrate for anammox[11]. In summer, denitrification was significantly correlated with nitrification (p < 0.05; Fig. 6), suggesting a strong coupling between nitrification and denitrification in sediments, where NO2− produced from nitrification may serve as a substrate for denitrification. Liu et al.[56] identified coupled nitrification–denitrification as a primary mechanism for sedimentary N removal. In contrast, no significant correlations were observed between anammox and either denitrification or nitrification (p > 0.05; Fig. 6). This lack of correlation may be attributed to intense competition for shared substrates (e.g., NO2− or NH4+).
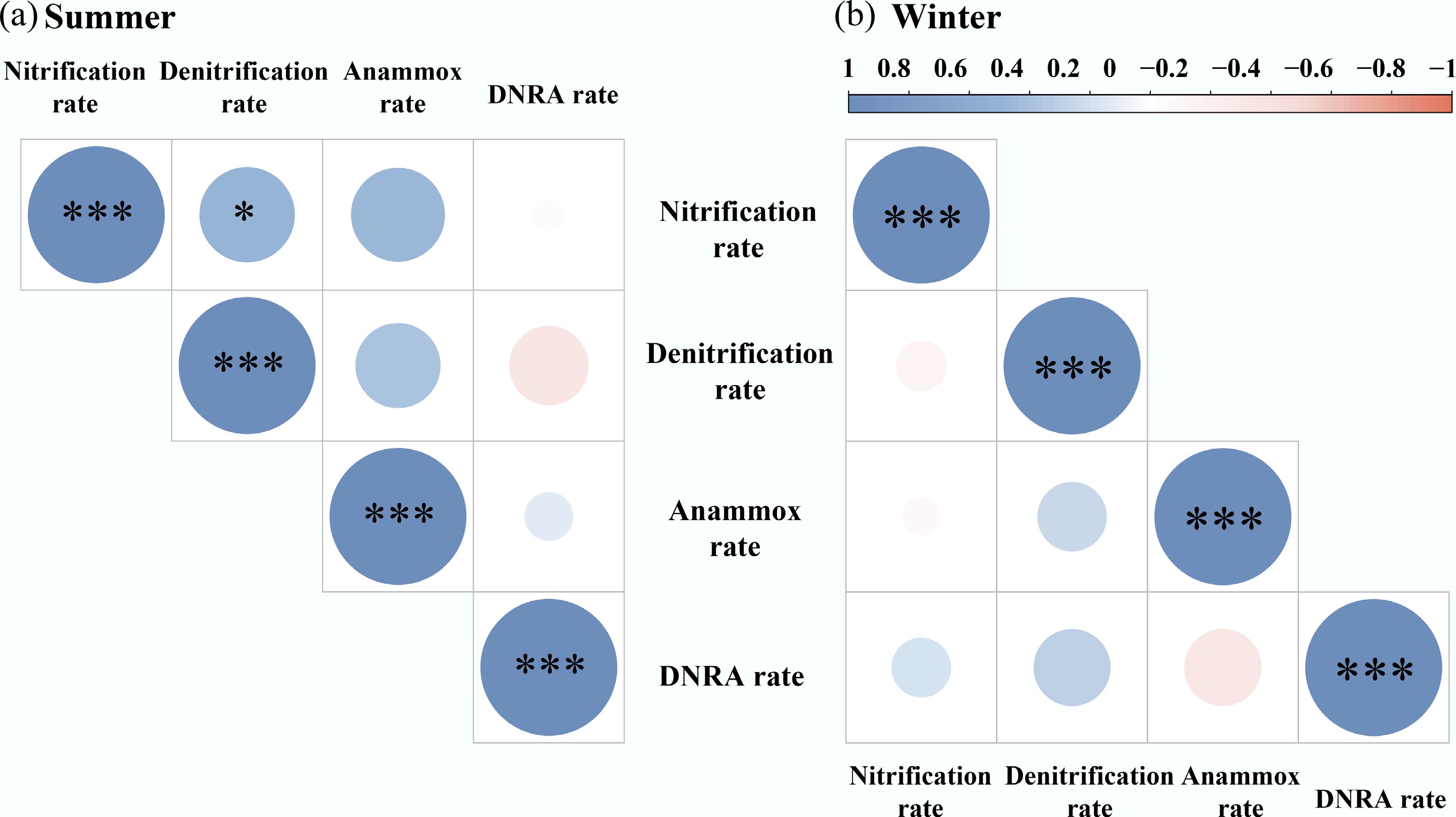
Figure 6.
Correlation coefficients among the N transformation processes in sediments, including net nitrification, denitrification, anammox, and DNRA in (a) summer, and (b) winter. * p < 0.05; *** p < 0.001.
However, during winter, coupling among the various N transformation processes was not evident (p > 0.05; Fig. 6). Low temperatures significantly suppressed microbial N-cycling activity (Fig. 2; Supplementary Fig. S2b) and the production of the intermediate substrates such as NO2− and NO3−[57], which may have weakened the coupling between N transformation pathways (e.g., nitrification–denitrification and denitrification–anammox). Because microbial communities responsible for nitrification, denitrification, anammox, and DNRA possess distinct temperature optima, cold weather can disproportionately inhibit specific steps within the N cascade, disrupting their metabolic synchronization[3]. In addition, the reduced river discharge in winter weakens hydrodynamic mixing, restricting substrate diffusion across oxic-anoxic interfaces[58]. Consequently, these processes appeared more independent or sporadic, reflecting a decoupled state.
Implications for N removal processes in rivers
-
Rivers function as integrators of materials produced in the draining catchment. In-stream components, such as microorganisms, nutrients, and physicochemical parameters are profoundly influenced by both catchment-scale anthropogenic activities and natural conditions[41]. For instance, land use (e.g., agriculture and urbanization) and geographical conditions (e.g., slope and precipitation) are closely linked with land-to-river nutrient fluxes and riverine geochemistry, thereby regulating microbial communities and N removal processes[59,60] (Fig. 5). Therefore, elucidating the mechanisms governing in-stream N removal requires a holistic approach that considers the catchment as an integrated system. This study clearly demonstrates that integrating catchment-scale geographical factors with microscale biotic and abiotic properties (water and sediment) offers a comprehensive understanding of the drivers regulating riverine N removal. Such a holistic perspective is critical for identifying effective strategies to control N pollution in river ecosystems, and for advancing our understanding of the global N cycle.
-
This study systematically identifies and quantifies N removal processes and their driving mechanisms in the Jinshui River by integrating remote sensing, 15N pairing techniques, and microbial molecular methods. This research indicates that N removal in forest-dominated riverine sediments is driven by distinct seasonal regulatory mechanisms (geographic-factor dominated/micro-factor dominated). The results demonstrated that denitrification constituted the primary N removal pathway and was predominantly regulated by micro-environmental factors (sediment properties, water properties, and the abundance of functional genes) in summer, whereas anammox was primarily constrained by geographic factors (altitude and land use). Conversely, in winter, water physicochemical parameters emerged as the primary drivers regulating N removal processes. These findings provide mechanistic insights into how hierarchical controls and seasonal context jointly regulate riverine N removal, thereby improving our ability to interpret spatial heterogeneity in N removal and to anticipate riverine N responses to climate change and land-use alteration. Furthermore, this study underscores the necessity of integrating interdisciplinary and cross-scale approaches to elucidate the mechanisms governing N removal.
-
It accompanies this paper at: https://doi.org/10.48130/nc-0026-0007.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: Wenshi Zhang: data curation, formal analysis, investigation, writing – original draft; Xiaodong Li: funding acquisition, methodology, writing – review & editing; Hao Jiang: conceptualization, funding acquisition, investigation, methodology, writing – review & editing; Quanfa Zhang: conceptualization, funding acquisition, writing – review & editing. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets used or analyzed during the current study are available from the corresponding author upon reasonable request.
-
This work was supported by the National Natural Science Foundation of China (32030069, U23A2048), the Key R&D Program of Guangxi Province (GUIKE AB24010128), and the Arts and Science Excellence and Innovation Award Program for Graduate Students of Tianjin University (No. B1-2024-015).
-
The authors declare that they have no conflict of interest.
-
Full list of author information is available at the end of the article.
- The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang W, Li X, Jiang H, Zhang Q. 2026. Geographical and micro-environmental factors regulate nitrogen removal in a forest-dominated river. Nitrogen Cycling 2: e020 doi: 10.48130/nc-0026-0007 

Geographical and micro-environmental factors regulate nitrogen removal in a forest-dominated river
- Received: 09 December 2025
- Revised: 14 March 2026
- Accepted: 16 April 2026
- Published online: 01 May 2026
Abstract: Nitrogen (N) removal processes (denitrification and anammox) are critical for mitigating N pollution in aquatic ecosystems. However, how macroscale geographical settings interact with micro-environmental factors to regulate these processes across seasons remains poorly resolved. Here, we investigated the seasonal drivers of N removal in a forest-dominated river by combining remote sensing, 15N-labeling, and molecular techniques (qPCR). Denitrification dominated sedimentary N removal, contributing approximately 90% of the total N removal in summer (1.68 ± 2.22 mg N kg−1 d−1) and 95% in winter (0.67 ± 1.52 mg N kg−1 d−1). In summer, Partial Least Squares Path Modeling (PLS-PM) revealed that denitrification was directly influenced by the abundances of functional genes, which were in turn regulated by water and sediment physicochemical properties. Anammox was significantly governed by geographical factors (e.g., altitude and land use). In winter, water properties (temperature, NH4+, NO3–) were the primary drivers directly regulating N removal. This study underscores the efficacy of incorporating cross-disciplinary and cross-scale methodologies to systematically elucidate the mechanisms and seasonal factors influencing N removal in forested rivers. These findings could improve current conceptual models of the global riverine N cycle and offer insights for sustainable N pollution management in forested catchments.
-
Key words:
- Sediment /
- River /
- N removal /
- 15N-labeling /
- Mechanisms