








-
Ganoderma lucidum (Lingzhi) has been used in traditional medicine for over 6,800 years[1]. Its spores, mycelia, and fruiting bodies produce a diverse array of bioactive compounds, including polysaccharides, triterpenoids, sterols, nucleosides, and amino acids. These constituents confer a broad spectrum of pharmacological activities, including immunomodulation, antitumor activity, anti-atherosclerotic effects, and lipid-lowering properties[2].
Among these, triterpenoids exhibit remarkable structural diversity, with over 300 compounds identified[3], including more than 100 ganoderic acids (GAs)[4]. As shown in Fig. 1, GAs can be broadly categorized into C30, C27, and C24 types based on carbon skeleton, or into acids, alcohols, and lactones based on functional groups. Despite their promising bioactivities, the therapeutic application of GAs is hindered by their extremely low abundance (often < 0.1% of dry weight[5]) and significant fluctuations across developmental stages. Furthermore, their complex chemical structures make large-scale isolation or chemical synthesis impractical. For the functional food and nutraceutical industries, these limitations pose major challenges in securing a stable supply, achieving standardized product profiles with consistent bioactivity, and controlling cost. Overcoming these constraints requires routes that decouple GA production from mycelial/fruiting-body physiology while maintaining food-grade quality and batch-to-batch consistency.
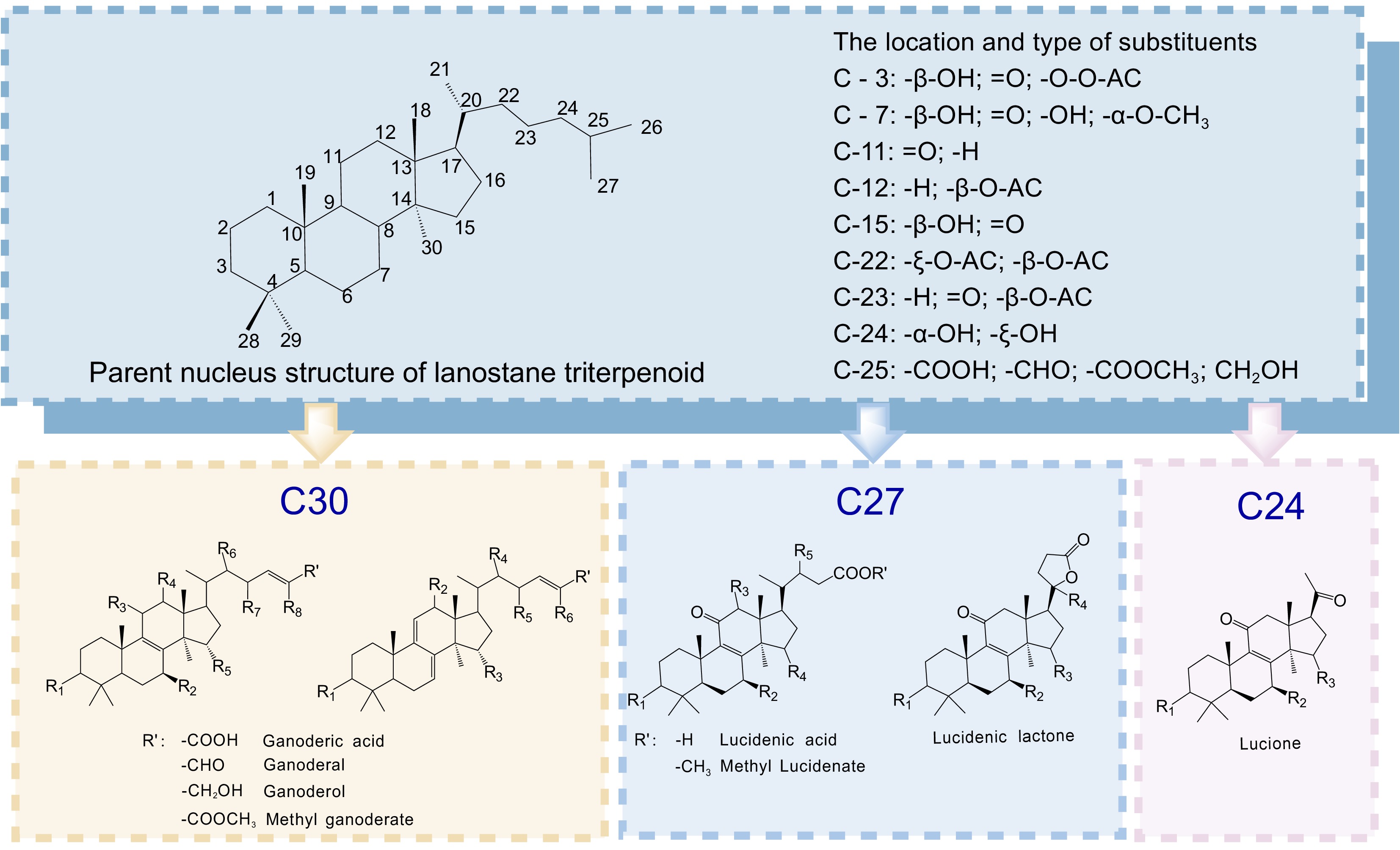
Figure 1.
Structural classification of key Ganoderma lucidum triterpenoids. The triterpenoids are categorized as C30, C27, and C24, based on their carbon-skeleton size. Variable substituents [Rn (n = 1–8) and R′] at key positions modulate their bioactivity and physicochemical properties.
Synthetic biology offers a powerful alternative to overcome these barriers by engineering microbial systems for efficient GA production. G. lucidum is an ideal model organism for such efforts due to its characterized genome, established transformation protocols, and active secondary metabolism[6]. Current strategies focus on two aspects: enhancing GA biosynthesis within Ganoderma itself and constructing heterologous GA biosynthetic pathways in food-safe microbial chassis, such as yeast or filamentous fungi (e.g., Aspergillus spp.).
Several previous reviews have summarized the structural diversity, bioactivities, or biosynthetic genes of Ganoderma triterpenoids[7−9]. In contrast, the present review places greater emphasis on integrating pharmacology, biosynthetic regulation, synthetic biology strategies, and efficacy-driven breeding into a continuous translational framework. We also incorporate recent advances reported up to early 2026 and highlight current bottlenecks and future opportunities for functional-food and nutraceutical applications. This review consolidates recent advances in understanding GA pharmacology and biosynthetic network, from precursor supply via the mevalonate (MVA) pathway to post-modification by cytochrome P450s. We critically evaluate synthetic biology approaches aimed at boosting GA production and discuss unresolved challenges, such as incomplete pathway elucidation and low heterologous titers. We also propose future directions in integrating multi-omics, CRISPR/Cas-based genome editing systems, and metabolic engineering to advance toward industrial-scale and economically feasible GA biomanufacturing.
-
GAs are highly oxidized lanostane-type tetracyclic triterpenoids commonly grouped into two structural classes: Type I GAs feature a single endocyclic double bond between C-8 and C-9 (Δ8(9)), and are typically decorated with C-3, C-11, and C-23 carbonyls, a C-7 hydroxyl, and a C-21 methyl group; Type II GAs are characterized by a conjugated diene system (Δ7(8),9(11)) with fewer ring substituents, and their functional diversity is introduced mainly at C-3 (acetoxyl, hydroxyl or carbonyl) and C-21 (methyl)[10]. Type I GAs have a single double bond, whereas Type II GAs possess conjugated double bonds. The structural modifications of Type II GAs often exhibit stronger or more diverse bioactivities in reported studies, including anti-tumor, immunomodulatory, and hepatoprotective effects. Classifying GAs based on their structures helps analyze the pharmacological activity differences among various GA components and is significant for screening and developing compounds with higher medicinal value.
This scaffold diversity underpins a broad spectrum of pharmacological activities that have been validated in vitro and in vivo. The representative structures of I/II GAs and their important biological activities are highlighted in Fig. 2, and the following sections summarize the key therapeutic effects, highlighting conserved mechanisms and structure-activity insights.
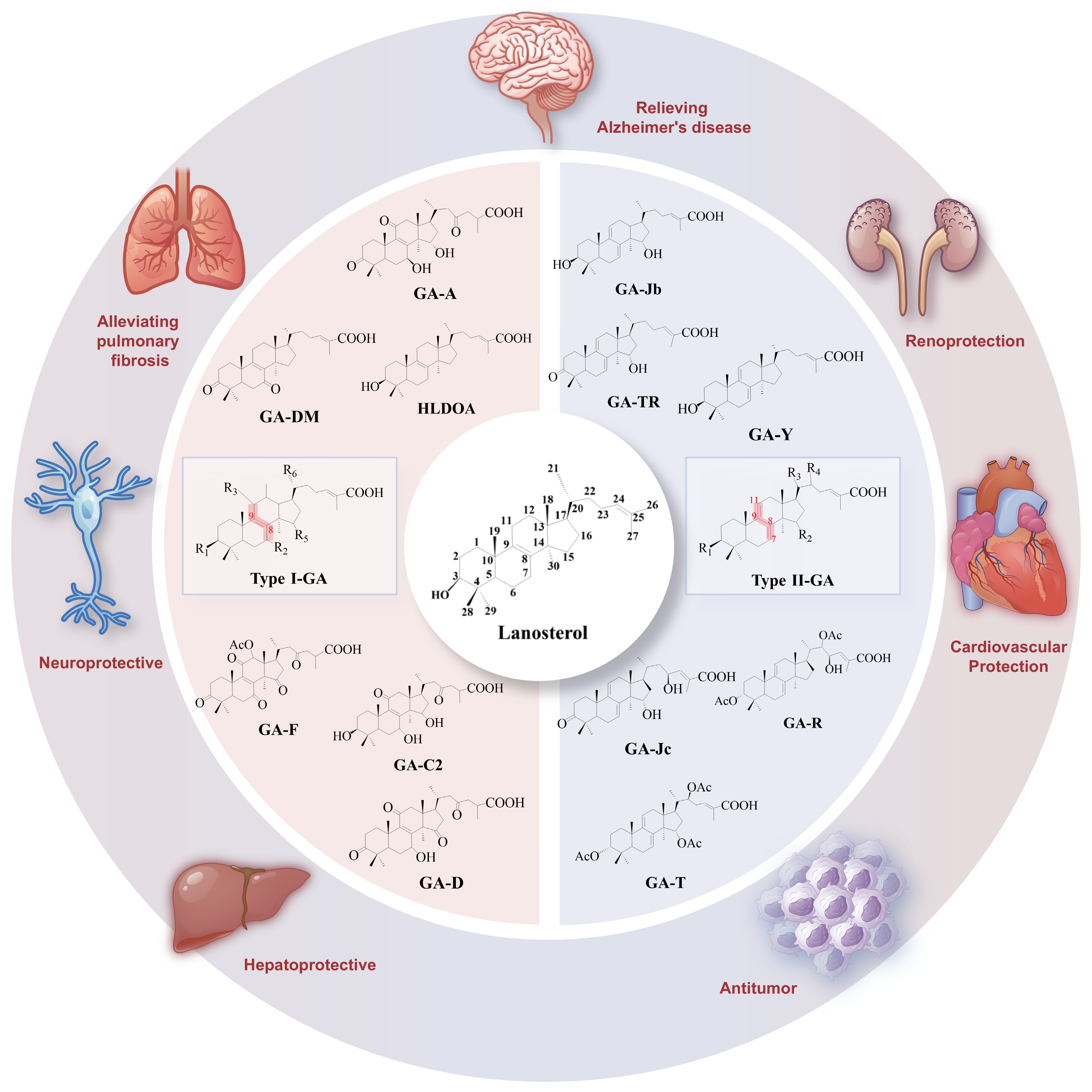
Figure 2.
Type I vs Type II GAs and representative bioactivities. The left and right fan charts summarize typical Type I and Type II scaffolds, respectively, highlighting their roles as Ganoderma actives with antioxidant, anti-inflammatory, immunomodulatory, and metabolic benefits.
Antitumor activity
-
Multiple GAs—including A, C2, D, DM, F, X, and Me—exhibit potent anti-cancer effects via complementary mechanisms, targeting hallmarks such as proliferation, cell death, angiogenesis, and metastasis. Ganoderic acid A (GA-A) demonstrates broad-spectrum activity[11]. In hepatocellular carcinoma, it re-programs tumor-associated macrophages through CSF1R inhibition, downregulates cell-cycle proteins[12], and alters enhancer-associated lncRNAs[13]; in prostate cancer, it blocks STAT3 signaling[14]; in neuroblastoma it suppresses Notch-1[15]; in breast cancer, it triggers mitochondrial apoptosis via the JAK2/STAT3 axis[16]; in osteosarcoma, it activates p38-NF-κB-mediated cell death[17]; and in lymphoma, it induces caspase-3/9 activity and modulates Bcl-2/Bax[18]. Ganoderic acid T (GA-T) enhances DNA damage and switches radiation-induced apoptosis to necroptosis in HeLa cells, markedly increasing radiosensitivity[19]. In an orthotopic ovarian cancer model, GA-T promotes ubiquitin-dependent degradation of galectin-1, reduces α-SMA-positive stromal cells, and increases immune-cell infiltration[20]. GA-T also inhibits liver cancer by activating pyruvate carboxylase. Ganoderic acids Mf and S (GA-Mf and GA-S) collapse mitochondrial membrane potential, release cytochrome c and activate caspase-3/9, leading to cell-cycle arrest, and apoptosis in cervical carcinoma cells[21]. Ganoderic acids A, D, DM, F, and H (GA-A, GA-D, GA-DM, GA-F) exert chemopreventive and therapeutic activity against breast cancer by orchestrating autophagy, triggering cell death, suppressing proliferation, and blocking angiogenesis[22]. Network-pharmacology analyses further nominate PIK3CA, EGFR, STAT1, and CTNNB1 as high-confidence targets of GA-DM[23,24]. Ganoderic acid X (GA-X) curbs hepatoblastoma growth and proliferation via autophagy induction, reflected by elevated Beclin-1, ATG5, and LC3-II levels and concomitant downregulation of p62[25]. Collectively, these studies establish GAs as multi-target anti-cancer agents capable of modulating immunity, apoptosis, angiogenesis, and metastasis.
Hepatoprotective effects
-
Lanostane-type triterpenoids broadly counteract liver injury induced by toxins, alcohol, or high-fat diets. At the molecular level, GA-A limits hepatocyte damage by repressing the Txnip/Trx/NF-κB and JAK2/STAT3 axes while upregulating cytoprotective ERp57, p-AKT, and p-MAPK signalling pathways[26]. In vivo, oral GA-A reverses alcohol-induced steatosis and rebalances the gut microbiota in mice[27]. Beyond metabolic insults, GA-A lowers serum ALT and AST, attenuates CCl4-mediated necrosis, and suppresses hepatitis B virus replication in HepG2.2.15 cells[28]. It also dampens neutrophil extracellular trap formation via p38 MAPK modulation, conferring protection in concanavalin A-induced autoimmune hepatitis[29]. Finally, GA-A rectifies the NAFLD-associated lipid signature—restoring ALT, AST, total bilirubin, triglycerides, and total cholesterol—while curbing free-fatty-acid influx, oxidative stress, and hepatic inflammation in rats[30].
Neuroprotective effects
-
GA-A attenuates neuroinflammation in models of Alzheimer's disease, epilepsy, depression[31], Parkinson's disease, and multiple sclerosis by suppressing pro-inflammatory cytokines[2]. GA-DM inhibits Aβ42-induced neuronal apoptosis and oxidative stress via NRF2 activation, alleviating cognitive decline[32]. Deacetylated GA-F (DeGA-F) reduces lipopolysaccharide (LPS)-triggered neuroinflammation by blocking NF-κB signaling in mouse brains[33].
Antioxidant effects
-
GA-A protects against oxidative injury by preserving glutathione (GSH) and glutathione peroxidase (GPX)[34], scavenging reactive oxygen species (ROS) and activating the NRF2/ARE pathway[35]. GA-D delays oxidative stress-induced senescence in human mesenchymal stem cells by targeting 14-3-3ε to activate CaM/CaMKII/NRF2 signaling[36]. These findings position GAs as potent natural antioxidants that counteract cellular ageing and ROS-mediated pathologies.
Cardiovascular protection
-
GAs exert anti-atherosclerotic effects by inhibiting macrophage M1 polarization through the TLR4/MyD88/NF-κB axis and stabilizing atherosclerotic plaques[37]. In a myocardial infarction model, GA-A downregulates TNF-α, IL-6, and IL-1β, reduces cardiac injury markers (CK-MB, LDH), and decreases infarct size[38]. GA-A, GA-B, GA-C6, GA-G, and ganodermanontriol from sporoderm-removed Ganoderma lucidum spore powder effectively suppress foam-cell formation. Specifically, GA-A and GA-G markedly upregulate ATP-binding cassette transporter A1 (ABCA1), whereas GA-A, GA-B, GA-G, and ganodermanontriol significantly increase ATP-binding cassette transporter G1 (ABCG1) expression in macrophages, driving macrophage cholesterol efflux and dampening lipid-driven calcification[39]. Concomitantly, these compounds repress IL-1β and TNF-α and reciprocally induce the anti-inflammatory armamentarium (eNOS, TGF-β, Arg1, and IL-10), thereby quelling plaque inflammation. Beyond lipid handling, GA-A, GA-B, GA-G, GA-C6, and ganodermanontriol activate Sirt7–Nrf2 signaling, boost NQO1 and HMOX1, and accelerate aortic mitophagy. This orchestrated response alleviates oxidative stress, decelerates vascular ageing, and provides sustained protection against atherosclerosis[40].
Additional therapeutic effects
-
Beyond the above, GAs demonstrate protective effects in several other contexts:
• Renal protection: GA-A activates the Trx/TrxR system against nephrotoxicity[41].
• Anti-pulmonary fibrosis: GA-A inhibits Rho/ROCK/NF-κB signaling and TGF-β/Smad/MAPK cascades.[42].
• Bone health: GA-A prevents osteoporosis via the PIK3CA/p-Akt/TWIST1 pathway[43].
• Anti-cellular senescence: GA-A binds TCOF1 to maintain ribosomal homeostasis[44].
• Anti-arthritis: GA-A reduces IL-6/TNF-α expression[45].
• Anti-inflammatory bowel disease: GA-A modulates tryptophan metabolism via gut microbiota, enhancing AhR activity[46].
• Anti-fatigue: GA-A can counteract exercise-induced fatigue by improving glucose metabolism through the AMPK/PGC-1α/GLUT4 pathway[47].
In summary, GAs deliver broad, multi-target pharmacological activities, with the therapeutic scope spanning cancer, hepatic disorders, neurodegenerative disorders, and cardiovascular and metabolic diseases, all with an encouraging safety margin. Unlocking their clinical potential critically depends on overcoming the supply bottleneck. The following sections delve into the biosynthetic pathways and synthetic biology strategies poised to enable scalable production of these valuable compounds.
-
The biosynthetic route to GAs originates in the cytosolic MVA pathway, which begins with the condensation of two acetyl-CoA molecules to acetoacetyl-CoA, catalyzed by acetyl-CoA acetyltransferase (AACT). Acetoacetyl-CoA is then converted to 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) by HMG-CoA synthase (HMGS). The committed and rate-limiting step is catalyzed by HMG-CoA reductase (HMGR), yielding mevalonate (MVA). Sequential phosphorylation by mevalonate kinase (MK) and phosphomevalonate kinase (PMK) generates mevalonate-5-pyrophosphate (MVA-PP), which is decarboxylated by mevalonate-5-pyrophosphate decarboxylase (MVD) to form isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP). Under the catalysis of isopentenyl diphosphate isomerase (IDI), IPP, and DMAPP can be interconverted through isomerization. These C5 units serve as universal precursors for all terpenoids.
Downstream condensation reactions produce geranyl pyrophosphate (GPP; C10), farnesyl pyrophosphate (FPP; C15), and squalene (SQ; C30), catalyzed by farnesyl diphosphate synthase (FPS) and squalene synthase (SQS), respectively. Squalene (SQ) is converted into 2,3-oxidosqualene under the catalysis of squalene monooxygenase (SE). Oxidosqualene cyclase (lanosterol synthase, LS) then converts 2,3-oxidosqualene to lanosterol[48], the first committed sterol/triterpenoid intermediate. The G. lucidum genome encodes two copies each of AACT and FPS, plus single copies of all other core MVA-pathway genes. Functional validation has confirmed the critical roles of AACT, HMGS, HMGR, MVD, IDI, FPS, SQS, SE, and LS in GA biosynthesis.
Cytochrome P450 post-modification pathway
-
The structural diversity of GAs arises from the extensive tailoring of lanosterol, primarily mediated by cytochrome P450 monooxygenases (CYPs). These enzymes introduce hydroxylations, oxidations, and other modifications at specific carbon positions, giving rise to the various GA subtypes. Key characterized reactions and their associated CYPs are summarized in Table 1. In Fig. 3, the biosynthetic pathways of GAs and their derivatives are summarized based on the substrates acted on by CYP enzymes and the products they catalyze, clarifying the localization of products and enzymes in the GAs biosynthesis pathway.
Table 1. The functions of CYP enzymes and regulatory factors in the synthesis of GAs in G. lucidum.
Gene name Mode of action Catalytic site/target gene Associated product Ref. CYP5150L8 CYP enzyme C-26 oxidation Lanosterol → HLDOA [49] CYP512U6 CYP enzyme C-23 hydroxylation GA-DM → Hainanic acid A; GA-TR → GA-Jc;
7-Oxo-GA-Z → GA-ZXYL[50] CYP5139G1
CYP FUM15A2CYP enzyme C-28 oxidation HLDOA → DHLDOA [51,52] CYP505D13
CYP5150W17CYP enzyme Oxidation 2, 3-Oxidosqualene → ST-3 [52,53] CYP512W2 CYP enzyme C-7/C-11/C-15 hydroxylation HLDOA → GA-Y; GA-Y → GA-Jb;
HLDOA → 15-/(15, 30)-hydroxy-GA-HLODA[52] CYP512V2 CYP enzyme -- GA-T [54] CYP512A3 CYP enzyme C-3/C-11/C-15 oxidation;
Δ24 (25) reductionHLDOA → Ganolucidic acid E;
Ganolucidic acid E → Ganolucidic acid F[55] CYP512A13 CYP enzyme C-7 oxidation
C-12 hydroxylation
C-15 oxidationGA-Jb → 7-Oxo-GA-Z3
GA-Jb → THLTOA
THLTOA → DHOLTOA[56] CYP512W6 CYP enzyme C-22 hydroxylation GA-Jb → 3β-TLTOA; GA-Ja → TLTOA;
GA-Me → GA-T1; GA-Mf → GA-T2;
HLODA → DLDOA[49] CsSDR Short-chain dehydrogenase C3 epimerization GA-Jb → GA-TR [49] AKR1C4 Ketone reductase C3 ketoreduction GA-TR → GA-Ja [49] GlAT Acyltransferase C15/C22 acetylation GA-Mf → GA-Me [49] BsAT Acyltransferase C3 acetylation GA-Ja → GA-Mf [49] SREBP bHLH-zip TF HMGR, MK GAs, Lanosterol, GA-C2 [57] GlbHLH1 bHLH TF HMGR, SQS, LS GAs [58] GlbHLH5 bHLH TF LS GAs [59] GlbHLH7 bHLH TF SQS, SE GAs [60] GlSwi6 APSES TF ROS GAs [61] CRZ1 Calcineurin-responsive TF Ca2+ GAs [62] PacC pH-responsive TF SQS, LS GAs [63] AreA GATA TF NO GAs [64] GlMADS1 Mads-box TF ROS [65] LaeA Methyltransferase SQS, LS GA-T, Me [66] VHb Homodimeric oxygen binding protein HMGR, SQS, LS, CYP512A2, CYP512V2, CYP512A13 GA-O, Mk, T, S, Me [67] GlSkn7 Stress-responsive TF HMGR, SQS, LS GAs [68] WC-2 Blue light photoreceptor Gl-25098, HMGR, SQS, LS GA-Mk, T, S, Me [69] Glsirt1 Lysine deacetylase ROS GAs [70] GlSlt2 mitogen-activated protein kinases ROS GAs [71] PKA protein kinase ROS GAs, GA-Mk, T, S, Me [72] Nox NADPH oxidase ROS GAs [73] GPx Glutathione peroxidase ROS GAs [74] AOX Alternative oxidase ROS GAs, Lanosterol, SQ [75] ODC Ornithine decarboxylase ROS GAs [76] PRMT5 Type II arginine methyltransferase GlPP2C1 GAs [77] GlPP2C1 Protein phosphatase SQS GAs [77] Gl-25098 Spore formation-specific genes HMGR, SQS, LS GA-T, Mk, Me [78] 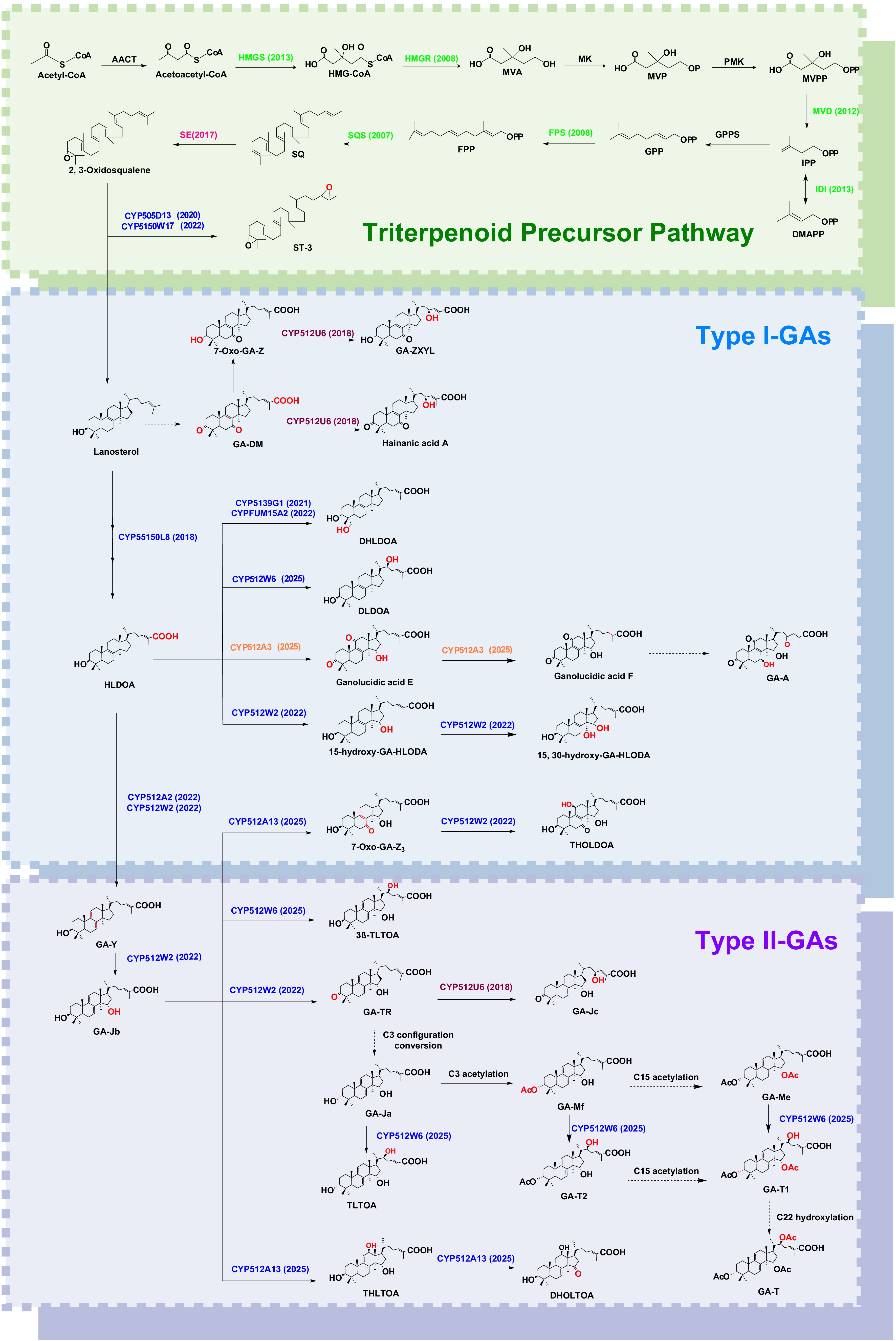
Figure 3.
Biosynthetic route to GAs and derivatives with implications for fermentation and metabolic engineering. '→' indicates a step catalysed by a defined enzyme; '$\twoheadrightarrow $' denotes a single enzyme catalysing multiple consecutive steps; '$\dashrightarrow $' marks unresolved steps or configuration interconversions without an assigned enzyme. Enzyme validation year is noted in parentheses after each enzyme; enzyme-modified sites are highlighted in red. Abbreviations: HMGR, SQS, OSC, CYP, SDR, UGT.
In addition to CYP enzymes, GA production is also regulated by various transcription factors and signaling components. These regulators modulate the expression of pathway genes in response to developmental or environmental cues. Their modes of action, molecular targets, and effects on GA biosynthesis are also summarized in Table 1, and Fig. 4 illustrates the modes of action of these transcription factors to briefly explain the regulatory mechanisms of GA synthesis.
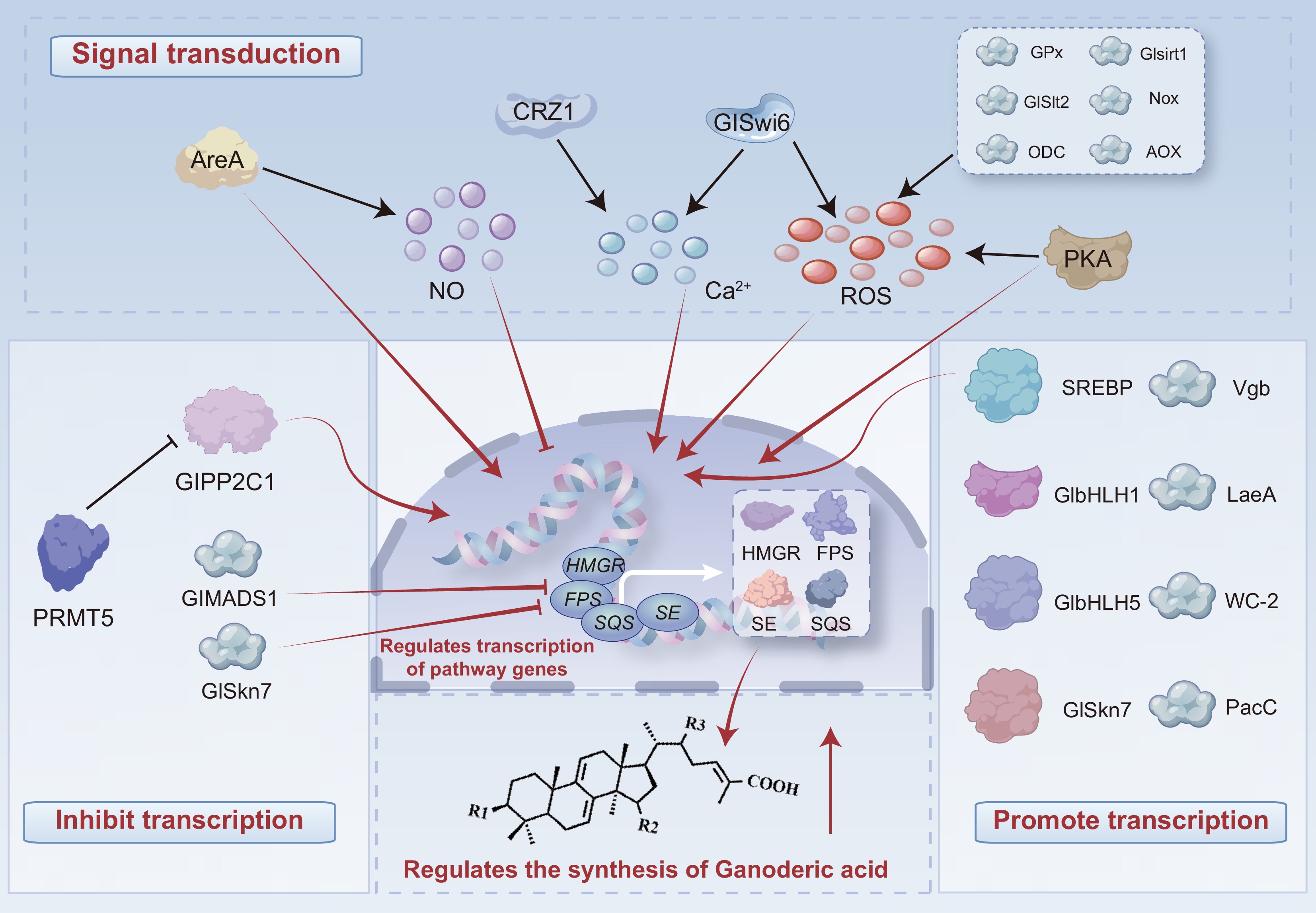
Figure 4.
Regulatory control of GA biosynthesis. Signal transduction: regulators modulate pathway-gene transcription via signaling cascades. Transcriptional repression: direct/indirect suppression of pathway genes. Transcriptional activation: direct enhancement of pathway genes. Shown targets are illustrative; regulators can influence additional nodes not depicted, informing strategies to boost yields in food-grade production.
-
Synthetic biology offers a powerful platform for the scalable production of high-value natural products, including triterpenoids from Ganoderma lucidum. A range of microbial chassis, such as bacteria (e.g., Escherichia coli), yeasts (e.g., Saccharomyces cerevisiae), and filamentous fungi (e.g., Aspergillus spp.), have been employed to reconstruct or enhance triterpenoid biosynthesis. Among these, filamentous fungi are particularly promising due to their innate capacity for secondary metabolism, robust protein secretion, and compatibility with CYP450 enzymes.
Compared to bacterial and yeast systems, filamentous fungi offer several advantages: they possess cellular compartments that support CYP450 activity, have well-developed post-translational modification machinery, and can correctly process intron-containing fungal genes. Additionally, their rapid growth, genetic tractability, and suitability for industrial-scale fermentation make them ideal hosts for the heterologous production of complex fungal metabolites such as GAs[79].
Fungal chassis systems
-
Several filamentous fungi have been explored as chassis for triterpenoid production.
Aspergillus niger: A well-established industrial strain known for high-yield citric acid production. It offers robust genetic tools and strong secretion capacity, though it may produce undesirable byproducts and can be genetically unstable[80].
Aspergillus oryzae: Traditionally used in food fermentation, this species is recognized as safe and has a well-characterized genome. It supports efficient protein expression and secondary metabolite production, though its slower growth rate can be a limitation[81].
Trichoderma reesei: Known for its exceptional cellulase production, it is increasingly used as a chassis for heterologous protein and metabolite production. Its strong inducible promoters and efficient secretion systems are advantageous, though product inhibition and stringent culture conditions can pose challenges[82].
Cordyceps militaris: An emerging fungal platform for the production of bioactive compounds such as cordycepin. While promising, it lacks well-developed genetic tools and industrial fermentation protocols[83].
Ganoderma lucidum: The native producer of GAs, G. lucidum is a logical chassis for metabolic engineering. It possesses the complete biosynthetic machinery for triterpenoid synthesis and is amenable to genetic transformation via protoplast-mediated methods, Agrobacterium tumefaciens-mediated transformation (ATMT), and electroporation. Limitations include slow growth and limited sporulation efficiency, which hinder large-scale cultivation[84].
Each chassis presents unique strengths and limitations. Ganoderma species, owing to their metabolic specialization for triterpenoid biosynthesis, represent particularly promising candidates for both native pathway optimization and synthetic pathway engineering.
Promoters and terminators
-
Efficient gene expression in synthetic biology depends heavily on the choice of regulatory elements:
• Constitutive promoters drive continuous expression and are commonly used to maintain pathway flux. Examples include gpdA from Aspergillus nidulans, pki1 from Trichoderma reesei[85], tef1 from Saccharomyces cerevisiae[86], paf from Penicillium chrysogenum[87], cdna1[88], and trpC from Aspergillus nidulans[89].
• Inducible promoters allow temporal control of gene expression, useful for balancing growth and production phases. Examples include glaA (glucoamylase), pcbAB (α-aminoadipyl-cysteinyl-valine synthetase), pcbC (isopenicillin N synthase) from Penicillium chrysogenum[90], and cbh1 (cellobiohydrolase I) from Trichoderma reesei[85].
• Terminators ensure proper transcription termination and enhance mRNA stability. Examples include trpC from Aspergillus nidulans, Anid_AN4594 and Anid_AN7354 from Aspergillus nidulans[91], and NOS from Agrobacterium tumefaciens.
Additionally, synthetic hybrid promoters and terminators are also being developed to fine-tune expression levels and improve metabolic flux control.
Selectable markers
-
Selectable markers are essential for screening and maintaining genetically modified strains. Resistance markers such as hph (hygromycin B), neo (G418), and bar (phosphinothricin) are widely used[92]. Marker recycling systems (e.g., Cre-loxP) enable multiple rounds of genetic modification without marker accumulation[93]. Auxotrophic markers like pyrG, argB, and trp1 require specific mutant backgrounds but allow for clean selection without antibiotics[94]. Fluorescent markers (e.g., GFP, mCherry) facilitate visualization of gene expression and can be used for high-throughput screening via flow cytometry.
-
Filamentous fungi, including G. lucidum, are prolific producers of bioactive natural products. However, their complex genetic backgrounds often hinder functional genomic studies and strain engineering efforts. Gene-silencing and editing technologies provide powerful tools to dissect biosynthetic pathways, identify gene function, and construct high-performance microbial chassis for natural product synthesis. These tools broadly function at two levels: transcriptional-level regulation, such as CRISPR-based systems that disrupt, delete, or precisely edit genes; and post-transcriptional regulation, such as RNA interference (RNAi), which degrades target mRNAs.
RNAi technology
-
RNA interference (RNAi) is an evolutionarily conserved mechanism in which double-stranded RNA (dsRNA) triggers sequence-specific degradation of homologous mRNA transcripts, leading to potent gene silencing[95]. In fungi, RNAi was first demonstrated in Neurospora crassa[96] and has since been successfully applied in over 30 filamentous species[97].
In G. lucidum, RNAi was initially established using the orotidine-5′-monophosphate decarboxylase gene (ura3) as a reporter. Electroporation of protoplasts with dsRNA targeting ura3 resulted in effective gene knockdown, validating the feasibility of RNAi in this species. This approach was subsequently used to silence NADPH oxidase genes (noxA, noxB, noxR), revealing their involvement in reactive oxygen species (ROS) generation and ganoderic acid (GA) biosynthesis. An Agrobacterium tumefaciens-mediated transformation (ATMT) system was later developed to deliver RNAi constructs. Dual-promoter silencing cassettes were found to be more efficient than single-promoter constructs, significantly enhancing gene knockdown efficiency[98]. RNAi has since become a routine tool for functional genomics in G. lucidum, enabling researchers to link specific genes to phenotypic changes and metabolic outputs. Despite its utility, RNAi has limitations. Off-target effects can occur when siRNAs bind to partially complementary sequences, leading to unintended gene silencing. Additionally, the transient nature of siRNA molecules limits the duration of gene suppression, which may hinder long-term studies or industrial applications.
CRISPR/Cas9 technology
-
The CRISPR/Cas9 system has revolutionized genome editing across eukaryotic organisms. Its core components are the Cas9 nuclease, a CRISPR RNA (crRNA), and a trans-activating crRNA (tracrRNA). The crRNA and tracrRNA form a single-guide RNA (sgRNA) that directs Cas9 to a specific genomic locus, where it introduces a double-strand break (DSB). This break is typically repaired by non-homologous end joining (NHEJ), an error-prone process that often results in insertions or deletions (indels) and subsequent gene disruption. Alternatively, homology-directed repair (HDR) can be exploited for precise gene editing when a donor DNA template is provided[99].
In filamentous fungi, CRISPR/Cas9 has been implemented using three main strategies: (1) transformation of pre-transcribed sgRNAs or sgRNA-expressing plasmids into strains constitutively expressing Cas9;
(2) use of a single plasmid encoding both Cas9 and sgRNA[100];
(3) direct delivery of Cas9–sgRNA ribonucleoproteins (RNPs) into protoplasts,
which avoids stable integration and reduces off-target effects. Codon-optimized versions of Cas9, particularly those tailored for fungal expression, are essential for efficient function. Additionally, nuclear localization signals (NLSs) must be fused to Cas9 to ensure proper nuclear import in eukaryotic cells.
CRISPR/Cas9 was first applied in G. lucidum in 2017, with initial editing efficiencies of 0.2–1.78 mutants per 107 protoplasts. Subsequent optimizations, including the use of strong fungal promoters and intron-containing Cas9 expression cassettes, improved editing rates to 14–18 mutants per 107 protoplasts[101]. A CRISPR-based in situ complementation system has also been developed, allowing functional restoration of GA biosynthesis in edited strains.
Recent studies have targeted genes involved in pyrimidine metabolism and GA biosynthesis. For example, disruption of CYP5150L8 significantly reduced the accumulation of several GAs, confirming its role in triterpenoid biosynthesis. Despite these advances, challenges such as low transformation efficiency and off-target effects persist, necessitating further refinement of the system[102].
CRISPR/Cas12a technology
-
CRISPR/Cas12a (formerly Cpf1) is an alternative to Cas9 that offers distinct advantages in certain contexts. Unlike Cas9, Cas12a does not require tracrRNA for crRNA processing and recognizes a TTTN protospacer adjacent motif (PAM), expanding the range of targetable genomic sites. Cas12a also generates cohesive DSBs with 4–5 nucleotide overhangs, which can improve the precision of NHEJ-mediated editing.
Cas12a systems have been successfully established in several filamentous fungi, including Aspergillus species[103] and, more recently, Ganoderma[104]. In G. lucidum, Cas12a has been used to disrupt the lanosterol synthase (LS) gene, leading to altered GA profiles and accumulation of upstream intermediates such as 2,3-oxidosqualene[104]. Although Cas12a generally exhibits lower editing efficiency than Cas9, its higher specificity makes it a valuable tool for precision genome editing.
CRISPR single-base editors
-
Single-base editors are derived from CRISPR/Cas systems but utilize catalytically inactive Cas proteins fused to deaminase enzymes. These tools enable precise C-to-T or A-to-G conversions without inducing double-strand breaks[105]. Cytosine base editors (CBEs) and adenine base editors (ABEs) have been widely used for targeted mutagenesis in microbes[106]. In fungi, base editors can introduce premature stop codons or disrupt start codons to achieve gene inactivation without DNA cleavage. This scarless editing approach is particularly useful for functional studies and strain engineering. However, base editing efficiency can be influenced by sequence context, and degeneracy in the genetic code may limit the number of editable sites.
Gene-silencing technologies such as RNAi and CRISPR/Cas systems have become indispensable for functional genomics and metabolic engineering in G. lucidum. RNAi offers a straightforward approach for transient gene knockdown, while CRISPR-based tools provide precise, heritable genome editing. Continued refinement of these technologies—particularly in improving transformation efficiency, reducing off-target effects, and expanding targetable sites—will be critical for advancing Ganoderma synthetic biology and industrial biotechnology.
-
The rapid development of synthetic biology has significantly transformed traditional metabolic engineering approaches. Direct manipulation of biosynthetic pathway genes enables precise redirection of intracellular metabolic flux toward desired products. Following the complete elucidation of the GA precursor biosynthetic pathway, overexpression of individual key genes, such as HMGR[107], SE[108], FPS[109], SQS[110], and LS[111], has been shown to significantly enhance total GA content as well as specific GA metabolites, including GA-T, GA-S, and GA-Me. Notably, combinatorial overexpression (e.g., HMGR + SE,[108]) consistently outperformed single-gene modifications, indicating synergistic effects in metabolic flux enhancement (Table 2).
Table 2. Advancements in the synthesis and regulation of ganoderic acids.
Strategy Gene/inducer Product Changes in product yield Chassis Ref. RNAi GlSkn7 GAs Increased by 55.9% GM [68] RNAi AreA GAs Increased by 27% in the ammonia source;
Increased by 77% in the nitrate sourceGM [64] RNAi PRMT5 GAs Increased by 1.48-fold GM [77] RNAi Glsirt1 GAs Decreased by 41.8% GM [113] OE HMGR GAs Increased by 1-fold GM [107] OE FPS GAs, GA-T, S, and Me Increased by 1.28-, 1.27-, 1.62-, and 1.80-fold GM [109] OE SQS GA-Mk, T, Me, and S Increased by 1.86-, 1.67-, 0.95-, and 0.25-fold GM [110] OE SE GAs, GA-T, S, Mk, and Me Increased by 0.3-, 2.2-, 1.4-, 0.8-, and 1.9-fold GM [108] OE HMGR, SE GAs, GA-T, S, Mk, and Me Increased by 2.5-, 4.9-, 3.5-, 1.4-, and 4.8-fold GM [108] OE LS GA-O, Mk, T, S, M, and Me Increased by 5.1-, 1.2-, 2.2-, 3.8-, 1.0-, and 0.9-fold GM [111] OE Vgb GA-S, T, Mk, and Me Increased by 0.4-, 1.2-, 0.9-, and 1.0-fold GM [67] OE Vgb GA-O, Mk, T, S, and Me Increased by 1.01-, 0.64-, 1.03-, 1.11-, and 3.05-fold GM [120] OE, ES Vgb, Ca2+ GA-O, Mk, T, S, and Me Increased by 32%, 56%, 24%, 66%, and 48% compared with Vgb overexpression alone GM [120] OE LaeA GA-T, and Me Increased by 25%, and 20% GM [66] OE Glnmnat GAs Increased by 43.1% GM [113] OE WC-2 (combined with blue light) GA-Mk, T, S, and Me Increased by 0.92-, 1.1-, 0.75-, 1.55-, and 0.74-fold GM [121] OE GlbHLH5 GAs Increased by 45% GM [59] OE GlbHLH1 GAs Increased by 38% GM [58] OE SREBP GAs, Ergosterol, Lanosterol and GA-C2 Increased by 1.87-, 1.84-, 1.89-, and 2.75-fold GM [57] ES 100 μM PHB GA-Mk, T, S, and Me Increased by 47%, 28%, 36%,and 64% GM [114] ES 10 mM CaCl2 (Ca2+) GAs, GA-Mk, T, S, and Me Increased by 2.7-, 1.6-, 3.5-, 2.2-, and 2.8-fold GM [116] ES 20 mM ASA GAs Increased by 1.8-fold GM [119] ES 5 mM AcOH GAs, GA-A Increased by 92% GM [122] ES 254 μM MeJA GAs Increased by 45.3% GM [117] ES 100 μM SA GAs Increased by 66% GM [118] ES 4 mM NaAc GAs Increased by 28.63% G. Lucidum fruiting body [115] ES 5 mM NAD+ GAs Increase by 56.2% GM [113] ES 80 μM EGCG GAs Increased by 36.3% GM [113] ES 100 μM GT GAs Decreased by 29.9% GM [113] Comprehensive processing Add Cu2+, carbon and nitrogen sources, tertiary light GAs 4.1 mg/100 mg DW in yield GM [123] HOE CYP5150L8 HLDOA 14.5 mg/L in yield after 120-h fermentation SC [49] HOE, FO, DPAS CYP5150L8, iGLCPR HLDOA 154.45 mg/L in yield SC [112] HOE, DPAS CYP5150L8, CYP5139G1 DHLDOA 2.2 mg/L in yield SC [51] HOE, FACS CYP5150L8, iGLCPR HLDOA 51.36 mg/L in yield SC [52] HOE (Twice) CYP5150L8, iGLCPR, CYP512W2 GA-HLDOA, Y, and Jb 9.66, 51.30, and 56.44 mg/L in yield SC [52] The abbreviations in this table: OE, overexpression; HOE, heterologous overexpression; ES, exogenous stimuli; FO, fermentation optimization; FACS, fluorescence-activated cell sorting; DPAS, dual-plasmid adjustable system; PHB, phenobarbital; ASA, aspirin; SA, salicylic acid; AcOH, acetic acid; NaAc, sodium acetate; MeJA, methyl jasmonate; NAD+, nicotinamide adenine dinucleotide; EGCG, epigallocatechin gallate; GT, gallotannin; GM, Ganoderma mycelium; SC, Saccharomyces cerevisiae. Given the chemical complexity of the native GA profile, the targeted production of individual GA compounds often requires heterologous reconstruction of the biosynthetic pathway. Functional CYPs have been successfully introduced into microbial chassis such as yeast and A. oryzae to achieve targeted synthesis of GA monomers. For instance, co-expression of CYP5150L8[49] and its redox partner iGlCPR[52] in both hosts enabled de novo biosynthesis of GA-HLDOA. A dual-regulon system that modulates the expression of these plasmid-borne genes using different antibiotic concentrations further increased product titers by approximately tenfold[112]. Similarly, fine-tuning the expression levels of CYP5150L8 and CYP5139G1 using the same regulatory system in Saccharomyces cerevisiae led to the production of GA-DHLDOA[51].
As the GA biosynthetic network continues to be deciphered, additional CYPs have been functionally characterized. For example, the introduction of CYP512W2 into yeast facilitated the synthesis of GA-Y and GA-Jb[52], and subsequent engineering of key residues in this enzyme significantly enhanced its catalytic efficiency and product yield.
Influence of regulatory factors
-
Unlike pathway enzymes, regulatory factors do not directly participate in GA biosynthesis but modulate metabolic flux through indirect mechanisms. These regulators typically function via two main routes (Fig. 4). (1) Transcriptional control, by binding to the promoter regions of pathway genes (e.g., HMGR, FPS, SQS, LS) or other genes (e.g., GlPP2C1), thereby modulating their expression levels. (2) Signal transduction, by altering intracellular levels of signaling molecules such as reactive oxygen species (ROS), nitric oxide (NO), and calcium ions (Ca2+), which in turn influence gene expression.
As summarized in Table 2, several positive regulators have been identified, including Glsirt1[77], Glnmnat[113], Vgb[67], LaeA[66], and the transcription factors GlbHLH5[59], GlbHLH7[60], and SREBP[57]. Conversely, negative regulators such as PRMT5[77], AreA[64], and GlSkn7[68] have been shown to suppress GA accumulation. Genetic manipulation—either overexpression or silencing—of these regulatory genes has been demonstrated to enhance GA content by 0.4- to 2-fold.
Effects of exogenous stimuli
-
Beyond genetic modifications, exogenous chemical treatments have been shown to effectively enhance GA biosynthesis. These compounds act by activating key enzymes, inducing the expression of biosynthetic genes, or increasing cellular permeability to facilitate product accumulation. Supplementation of culture media with optimal concentrations of various elicitors, such as NAD+[113], EGCG[113], phenobarbital[114], sodium acetate[115], Ca2+[116], MeJA[117], salicylic acid[118], or aspirin[119], has been shown to reproducibly increase GA titers. The efficacy and specificity of these stimulatory effects vary depending on the compound used and the target metabolite. The types of these exogenous compounds and the resulting stimulatory effects are also summarized in Table 2.
-
Ganoderma breeding techniques have been developed in China for decades, with large-scale artificial cultivation first achieved in the 1950s[124]. Starting with the domestication of wild isolates, a succession of strategies, including mass selection, mutagenesis, hybridization, and transgenic engineering, has been employed to develop strains with improved yield, faster growth, broader adaptability, and enhanced metabolite production. Figure 5 (created with BioGDP.com[125]), shows that this breeding trajectory reflects a progressive shift from phenotype-based selection toward more targeted and mechanism-informed improvement. Using the 'Xianzhi' series as an example, the figure not only illustrates the stepwise breeding pipeline from germplasm development to elite strain generation but also highlights a future transition toward efficacy-driven breeding. In this emerging paradigm, breeding goals are no longer limited to increasing the yield of individual compounds; instead, they are oriented toward defined triterpenoid profiles, stable quality attributes, and target health functions of functional food products. This concept provides the basis for the practical implementation pathway and challenge analysis discussed in the following section.
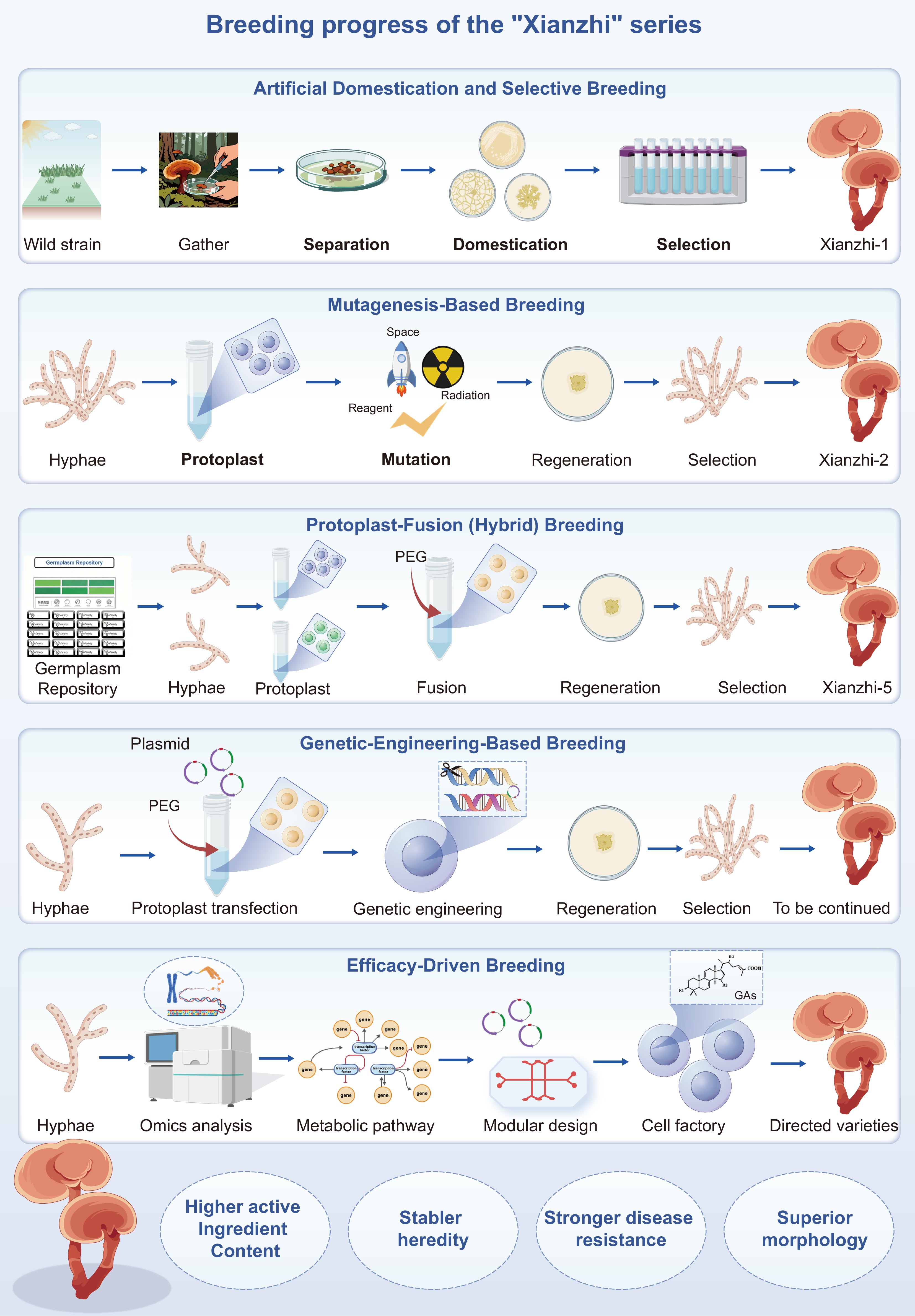
Figure 5.
Breeding pipeline for Ganoderma geared to food-technology outcomes. Using the 'Xianzhi' series as an example, the schematic outlines the germplasm development process, highlighting key breeding techniques aimed at elite edible G. lucidum lines with defined triterpenoid profiles, consistent quality, and targeted health functions for functional-food applications.
Upregulating GA content through breeding technologies
Artificial domestication and selective breeding
-
Wild basidiomes are surface-sterilized and introduced into axenic culture via tissue or single-spore isolation. Repeated sub-culture on sawdust-based formulas adapts the isolate to artificial substrates, preserves genetic diversity and typically doubles GA yield within 5–7 passages[126]. Elite morphotypes are then entered into multi-trait selection programmes. Classic cultivars such as 'Xianzhi-1', 'Kangding Lingzhi', 'Yaochizhi-1' show 20%–40% faster mycelial growth and 15%–25% higher GAs titres than their wild progenitors[127−129]. Integrating germplasm collection with whole-genome sequencing has further accelerated this process. For example, by first identifying genetically diverse wild isolates and then conducting cultivation and small-plot trials, the cultivar 'Xianzhi-3' was developed, which is characterized by elevated active-component content, a shortened growth cycle, superior morphology, and robust disease resistance[130]. Nevertheless, the approach still demands large populations and many generations to fix favourable alleles.
Mutagenesis breeding
-
Due to the thick bilayer sporoderm of Ganoderma spores, isolated protoplasts are preferred for mutagen treatment[131]. Ultraviolet (UV) mutagenesis of protoplasts has yielded strains with elevated polysaccharide[132], or organogermanium production[133], as well as increased basidiospore yield and enhanced resistance to microbial contaminants[134]. Treatment of G. lucidum protoplasts with lithium chloride produced mutants with markedly higher triterpenoid titers[135].
Space-flight mutagenesis (exposure to cosmic radiation and microgravity) led to the development of 'Xianzhi-2', a strain that tolerates temperatures up to 35 °C and contains 30% higher GA and 25% higher polysaccharide levels than its terrestrial counterpart; the polysaccharide fraction also shows enhanced β-1,3-linkages and stronger anti-inflammatory activity[136,137]. Although mutagenesis is technically straightforward, mutations occur randomly, and extensive back-crossing or genome resequencing is often required to separate beneficial from deleterious genetic changes[138].
Protoplast-fusion (hybrid) breeding
-
Hybrid breeding is one of the most widely used and effective strategies in edible-mushroom improvement. Sexual hybridization in Ganoderma is complicated by tetrapolar mating[139]. Monokaryotisation of protoplasts and chemical fusion of polyethylene-glycol-treated protoplasts (monokaryon–monokaryon crosses) bypass this bottleneck and allow whole-genome shuffling. Notable hybrids include 'Zhi-102' (Korean 'Nanhán Lingzhi' × Fujian 'G8-2') yielding 38% more polysaccharides and 20% more GAs than the better parent[140]; 'Xianzhi-5' (wild G77 × 'Xianzhi-1') fruits earlier and contains 1.55% triterpenoids (≈83% increase compared with Xianzhi 1)[141]; and Fusant RS7, generated from two high-ergothioneine parents, shows 30% higher ergothioneine, and a 15% GA bonus after genome-resequencing-guided stabilisation[142]. Protoplast fusion is now routine in China, but its success heavily depends on high-quality parental strains and subsequent marker-assisted selection.
Genetic-engineering-based breeding
-
Stable DNA transfer in Ganoderma is achieved through methods such as Agrobacterium-mediated transformation (ATMT), electroporation, and restriction-enzyme-mediated integration (REMI). Three main genetic engineering strategies have been employed (Table 2):
(1) Heterologous gene expression: the Vitreoscilla hemoglobin gene (Vgb) improves oxygen supply and boosts GA-S, GA-T, GA-Mk, and GA-Me by 20%–60%.
(2) Overexpression of rate-limiting enzymes: HMGR, SQS, SE, and LS single-gene constructs increase total GAs by 1.5- to 2.2-fold; combinatorial overexpression gives additive or synergistic gains.
(3) Silencing negative regulators: RNAi knock-down of PRMT5 or GlSkn7 raises GAs titres by 40%–55%.
To stack multiple desirable traits, monokaryons carrying different transgenes can be crossed. For example, a dikaryon co-expressing Vgb and SQS accumulates squalene and lanosterol at levels 2.4- and 1.8-fold higher, respectively, and the production of GA-P, GA-T, and GA-Me increases by 2.7-, 2.2-, and 1.8-fold, compared to the wild type[143]. It is important to note that most engineered strains have been characterized only in mycelial culture; fruiting-body validation and regulatory safety assessment are still pending.
Efficacy-driven breeding
-
Synthetic biology is transforming mushroom breeding from an empirical process into a precision-engineering discipline. CRISPR base-editors, for instance, can now fine-tune endogenous promoters or introduce premature stop codons without inserting foreign DNA, thereby clearing regulatory hurdles that have historically hindered the adoption of genetically edited fungi. In microbial factories such as yeast or Aspergillus, the modular reconstruction of GA pathways already enables the production of early-stage GAs at tens to hundreds of mg/L in engineered systems[52], and machine-learning-guided iteration may further improve yields in the future.
The next frontier is to reintegrate these synthetic breakthroughs into the mushroom itself. By coupling high-yield fermentation in 'cell-factories' with targeted sexual back-crossing, alleles discovered and optimized in silico can be introgressed into elite fruiting-body-producing strains within months. This approach marries the precision of synthetic biology with the low-cost scalability of traditional mushroom cultivation.
As pharmacological studies increasingly assign specific bioactivities to individual GAs, breeding goals can be redefined in terms of therapeutic efficacy rather than mere yield. We therefore propose an efficacy-driven breeding paradigm: use the synthetic toolkit described above to develop cultivars that hyper-accumulate a single, well-defined GA (e.g., GA-A or GA-T) or, going further, to assemble a tailored blend of GA 'cocktail' whose collective profile is optimized to deliver a desired therapeutic outcome, such as maximal immunomodulation or enhanced anti-tumor activity.
-
Triterpenoids from Ganoderma lucidum, especially GAs, are highly promising as bioactive ingredients for functional-food and nutraceutical applications. Over the past decade, rapid advances in pathway discovery, enzyme characterization, and chassis engineering have created a workable blueprint for GA biomanufacturing. Both native Ganoderma and food-relevant heterologous hosts (e.g., yeasts and filamentous fungi) now serve as platforms for stepwise pathway reconstruction, while the integration of multi-omics and CRISPR-based genetic tools enables targeted flux control. Collectively, these developments are shifting the field from reliance on low-yield extraction toward scalable, specification-driven production of well-defined GA profiles.
Nevertheless, important challenges remain. The GA biosynthetic network is still incompletely resolved, particularly concerning late-stage oxidations, ring rearrangements, and acyl/glycosyl tailoring, which constrains both yields and chemical diversity. Heterologous production is frequently limited by inefficient CYP450 expression, redox balancing, and cofactor supply. At process scale, strain stability, oxygen transfer efficiency, and downstream purification significantly impact production costs. Furthermore, clear and harmonized regulatory pathways for food-grade, fermentation-derived GA ingredients (including safety, quality, and labeling) are still evolving.
Looking forward, the following priorities are critical for advancing the field:
(1) Complete pathway elucidation: Combine targeted genetics with time-resolved metabolomics to assign the remaining biosynthetic steps and unlock late-stage GA diversification.
(2) Optimize CYP450s performance for scale production: Engineer membranes, redox partners, and cofactor regeneration; employ protein design to enhance enzyme turnover, coupling efficiency, and substrate range.
(3) Design for high flux and genetic stability: Use dynamic control circuits, organelle/peroxisome–ER compartmentalization, and stable genome integration to maintain high metabolic flux without compromising strain fitness.
(4) Process intensification: Optimize oxygen transfer and shear using bioreactor engineering; explore in-situ product removal and two-phase systems to mitigate product inhibition and ease downstream recovery.
(5) High-throughput design-build–test–learn cycles: Deploy biosensors, microfermentation arrays, and adaptive laboratory evolution to rapidly identify high-performing strains and novel GA analogues.
(6) Quality by design for food-grade products: Define critical quality attributes (CQAs) for GA fingerprints, impurities, and residual solvents; implement robust, validated quality control systems aligned with GRAS/novel-food regulations.
(7) Safety and efficacy translation: Pair standardized GA compositions with mechanism-anchored bioassays and human-relevant models to substantiate structure–function claims for specific health endpoints.
(8) Sustainability metrics: Use life-cycle assessment and techno-economic analysis early to guide chassis selection, media design (e.g., agri-side-streams), and waste minimization.
(9) Breeding + engineering convergence: Apply genomic selection in Ganoderma breeding to deliver elite, food-grade starting material, while parallel synthetic-biology routes furnish tunable, year-round supply.
Currently, engineering breeding often focuses on individual products, aiming to increase product yield. In the future, efficacy-driven breeding will not be limited to monomeric components with biological activity, but will target a large group of substances with the same biological activity. High-throughput sequencing technology can be used to perform whole-genome scans on breeding populations, constructing high-density genomic marker maps. Through bioinformatics analysis, gene loci or genomic regions significantly associated with target traits can be identified, and gene overexpression or editing can be used to modify the functional gene population in Ganoderma lucidum. In the breeding process, Ganoderma strains with outstanding efficacy, rapid growth and development, and excellent traits can be obtained.
Despite the broad prospects of efficacy breeding, the following risks and challenges still need to be addressed:
(1) Technical complexity and cost: Although the costs of genotyping and sequencing are decreasing, they still represent a significant expense for large-scale breeding populations.
(2) Off-target effects: Gene editing tools may cut or modify non-target sites, leading to unexpected genetic mutations and posing potential biosafety risks.
(3) Health risks: Gene-edited strains may not contain foreign DNA, but off-target effects or unintended disruptions of metabolic pathways may still generate new safety concerns, so their long-term health risks need to be carefully assessed.
(4) Strain degradation risk: During long-term strain cultivation, genes controlling desirable traits may undergo spontaneous negative mutations, resulting in weakened or lost functions, which undoubtedly increases the difficulty of long-term breeding.
By addressing these elements, the field can evolve from artisanal extraction to predictable, industrial-scale biomanufacturing. This will deliver GA ingredients with consistent composition, verified safety, and validated bioactivity. Such an integrated platform will not only broaden the availability of Ganoderma triterpenoids for functional foods and beverages but also provide a generalizable blueprint for the sustainable production of other high-value fungal metabolites.
-
The authors confirm their contributions to the paper as follows: data curation, writing − original draft, writing − review and editing: Yang J; data curation, writing − original draft: Zeng Y; data curation: Zhang G, Li M; writing − review and editing, supervision: Li Z. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
This study was financially supported by the Central Guiding Local Science and Technology Development Fund Project (2024ZY01009), and the Zhejiang Provincial Key R&D Program: Vanguard and Leading Goose Initiative (2025C01133).
-
The authors declared that they have no conflicts of interest to this work.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Jilin Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang J, Zeng Y, Zhang G, Li M, Li Z. 2026. Ganoderma triterpenoids: a review of pharmacological advances, biosynthesis, and synthetic biology strategies. Panfungi 1: e008 doi: 10.48130/panfungi-0026-0002 

Ganoderma triterpenoids: a review of pharmacological advances, biosynthesis, and synthetic biology strategies
- Received: 21 January 2026
- Revised: 30 March 2026
- Accepted: 20 May 2026
- Published online: 24 June 2026
Abstract: Ganoderma lucidum (Lingzhi) owes its broad health-promoting properties primarily to ganoderic acids (GAs), a class of highly oxidized lanostane-type triterpenoids with demonstrated antitumor, hepatoprotective, neuroprotective, antioxidant, and cardioprotective activities. However, natural GAs titres are extremely low and vary across developmental stages, and their structural complexity hampers large-scale extraction or total chemical synthesis, thereby limiting their use as standardized ingredients in functional foods and nutraceuticals. This review synthesizes recent advances in GA pharmacology, maps the biosynthetic network from mevalonate-pathway precursors through cytochrome P450-mediated tailoring, and assesses synthetic-biology strategies—including chassis selection, pathway engineering, and regulatory optimization— to overcome bottlenecks for food-grade production. Although key enzymes and regulators have been identified, the biosynthetic network remains incompletely resolved, and heterologous production yields remain low. The integration of multi-omics analyses, CRISPR/Cas-based genome editing, and metabolic-flux control now enables rational strain design of native Ganoderma or food-relevant filamentous fungal hosts (e.g., Aspergillus spp.) for scalable and consistent GA biomanufacturing. We further outline critical research priorities, including pathway annotation, mechanistic dissection of regulation, and translation of these insights into robust, industry-ready processes tailored to functional-food and nutraceutical applications.
-
Key words:
- Ganoderma lucidum /
- Ganoderic acids /
- Biosynthetic network /
- Synthetic biology /
- Functional foods