










-
Rice (Oryza sativa L.) is a staple food for more than half of the world's population, providing essential calories and nutrition[1]. The quality of rice, particularly its eating and cooking properties, is largely determined by starch composition, primarily amylose and amylopectin. One of the most important genes influencing rice starch properties is the Wx gene, which regulates amylose synthesis in the endosperm. Consequently, understanding the genetic and molecular regulation of the Wx gene is critical for breeding rice cultivars with desirable cooking and eating qualities[2].
As an essential food and key agricultural product, rice holds significant economic and nutritional importance in China[3]. Rice quality is primarily determined by the synthesis, composition, distribution, and accumulation of nutrients and storage compounds, particularly starch and protein. Among the various quality traits, cooking and eating quality are considered key indicators influencing consumer preference. In current breeding practices, improving the eating quality of rice largely depends on allelic variation in the Waxy (Wx) gene, which encodes granule-bound starch synthase I (GBSSI)—a key enzyme responsible for amylose biosynthesis. Variations in Wx allow for the development of rice varieties with different amylose contents, thereby enabling breeders to tailor cooking and textural properties to meet the diverse demands of global consumer markets[4].
The expression of Wx gene is finely controlled at the post-transcriptional level, with a critical GT/TT polymorphism at the 5′ splice site of its first intron affecting splicing efficiency. This polymorphism gives rise to two predominant alleles: Wxa, associated with intermediate to high amylose levels, and Wxb, linked to low to intermediate amylose content[5]. Additionally, the null wx allele completely disrupts amylose synthesis, resulting in starch composed solely of amylopectin, commonly known as waxy or glutinous rice[6]. This waxy phenotype, highly valued for its sticky texture, is integral to many traditional Asian dishes and has been cultivated in China for over 2,000 years[7]. Today, glutinous rice remains highly valued for its unique chewiness and versatility, with waxy starch offering exceptional stickiness and freeze-thaw stability, making it a key resource in food and industrial applications[8]. Glutinous rice, also known as sticky or waxy rice, has a rich history, particularly in East and Southeast Asia[7].
The sticky texture of waxy rice made it ideal for traditional dishes like zongzi (rice dumplings), mochi, and fermented products like rice wine. In Southeast Asia, varieties like Thai sticky rice (Khao Niao) became staples in regions where steaming, rather than boiling, was a preferred cooking method. The null wx allele likely emerged through natural mutations, which farmers then selected for specific purposes. Genetic studies suggest that waxy rice spread across Asia via trade and migration, with distinct mutations in the Indica and Japonica subspecies indicating independent domestication events[9].
Rice germplasm collections play a critical role in advancing genetic research by offering a wide range of variation for traits associated with eating quality[10]. Ancestral rice species of the genus Oryza provide rich genetic diversity and adaptability, offering valuable traits such as stress resistance. The reduced genetic variation caused by intensive breeding can be mitigated through systematic use of ancestral germplasm, supported by ecological data and advanced identification methods[11]. In this context, Malaysia's tropical climate and its diverse rice cultivars make it a valuable hotspot for investigating eating quality genes. Of particular interest is the identification of novel Wx alleles in Malaysian rice, which will not only highlight the genetic uniqueness of the local germplasms but also raise important questions about their evolutionary origin. Therefore, systematic screening of Malaysian rice accessions for Wx gene variation is essential for uncovering novel alleles that can be harnessed in breeding programs aimed at enhancing rice eating quality.
This study aims to investigate the genetic and phenotypic basis of eating quality in Malaysian rice varieties through allele characterization and trait analysis. By focusing on this underexplored germplasm, the research seeks to uncover functional variations of key eating quality-related genes and clarify their contributions to consumer-relevant traits. The findings will support future breeding programs aiming to enhance rice quality and will add valuable information to the global understanding of rice eating quality diversity. Ultimately, this work emphasizes the importance of Malaysian rice varieties as a genetic resource for improving the culinary value of rice in regional and international markets.
-
Fifty nine Malaysian rice varieties were collected to investigate their genetic diversity through genotyping analysis using marker development at the Waxy gene locus. The seeds of the Malaysian varieties were planted at the Lingshui Experiment Station in Hainan, China (18.25° N, 109.50° E) for trait analysis. All the varieties were assigned to three major subgroups, including 18 Tropical Japonica, 36 Indica, and five admix type varieties, respectively, based on the International Rice Research Institute (IRRI) classification system (Supplementary Table S1).
Determination of AAC, (gel consistency) GC, GPC, RVA, and DSC
-
Rice grains of each sample were harvested at the mature stage and dried in a 40 °C oven for seven days, then stored at room temperature for measuring grain quality traits. For each variety, more than 100 g of brown rice was prepared for measuring grain quality, including AAC, GC, GPC, DSC, and RVA. AAC was measured using the iodine colorimetric method with some modifications described by Juliano[12]. It is noted that the iodine colorimetric method may slightly overestimate amylose levels in some varieties, as long-chain amylopectin fractions can also bind iodine in addition to amylose[13]. GC was measured by placing 100 mg of rice flour in a 10 mL test tube with 0.2 mL of 0.025% thymol blue ethanol solution, and 2.0 mL of 0.2 mol/L KOH. After mixing, the tubes were covered with glass beads and heated in a boiling water bath for 8 min, then cooled at room temperature for 5 min and in ice water for 20 min. Tubes were then laid horizontally at 25 ± 2 °C, and after 1 h, the gel length was recorded in millimeters as the GC value[14].
The GPC was determined by near-infrared spectroscopy using an Infratec 1,241 Grain Analyzer (Foss Tecator, Sweden). Briefly, approximately 100 g of dry seeds per sample were dehulled and scanned three times per sample; results are presented on a 0% moisture basis.
RVA, was analyzed with 3 g rice flour (12% moisture) in 25 ml distilled water. Heating and cooling cycle followed standard protocols: 50 °C for 1 min; ramp to 95 °C at 12.2 °C/min, hold 2.5 min, then cool to 50 °C at 11.8 °C/min, with stirring at 160 rpm after an initial 10 s at 960 rpm[15].
DSC was conducted using a Hart Scientific model 7701 thermal analyzer. Samples (~500 mg, 70% moisture) were heated from 20 to 110 °C at 0.17–1.33 °C/min. Water was used as a reference[16].
Development and genotyping of allele-specific markers
-
To quickly discriminate different alleles of Waxy gene, molecular markers for InDel, dCAPS and ARMS-PCR were developed. For InDel markers, the primers were designed by the software Primer Premier 5 directly. The dCAPS Finder 2.0 tool (
http://helix.wustl.edu/dcaps/dcaps.html ) was used to design dCAPS markers and forward primers with restriction enzyme sites were selected. To match the forward primers, Primer Premier 5 was used to select reverse primers. The program developed by Ye et al.[17] was used to design the tetra-primers ARMS–PCR.Genomic DNA was extracted using the TPS method[18]. PCR products were separated on 3% agarose gels and visualized under UV light. For the dCAPS marker, PCR products were digested with the EcoRI at 37°C for one hour, and then run on 3% agarose gel for discrimination.
Extraction of total protein, GBSSI, and SDS-polyacrylamide gel electrophoresis
-
Total protein was extracted from 70 mg rice flour with the extraction buffer (125 mM Tris-HCl, pH 6.8, 4 M Urea, 4% SDS, 5% 2-Mercaptoethanol) and shaken at 37 °C for 3 h. Proteins were centrifuged, and the supernatant was stored at 4 °C for analysis.
For GBSSI protein extraction, 50 mg of rice flour mixed with SDS buffer containing 0.05 M Tris-HCl (pH 6.8), 2.8% SDS, 10% glycerol, and 5% 2-Mercaptoethanol, was centrifuged, washed with acetone, vacuum-dried, resuspended in SDS buffer, and proteins were denatured by boiling[19]. SDS-PAGE was performed following the procedure of Sambrook & Russell[20].
Statistical analysis
-
All statistical analyses were performed using Minitab 21. Descriptive statistics and one-way analysis of variance (ANOVA) were conducted to assess variability and detect significant differences among genotypes. Post-hoc comparisons were performed using Tukey's HSD test at a significance level of p < 0.05. All graphs and visualizations were generated using GraphPad Prism 10 and Microsoft Excel.
-
To investigate the genetic basis of grain quality in Malaysian rice varieties, functional molecular markers were developed for allele variants of the Wx gene, which could associate with this trait (Fig. 1 and Supplementary Table S1). Specifically, InDel markers were designed for the wx23 bp allele. Tetra-primer ARMS-PCR (TETRA) markers were developed to target Wxin and Wxlv alleles.
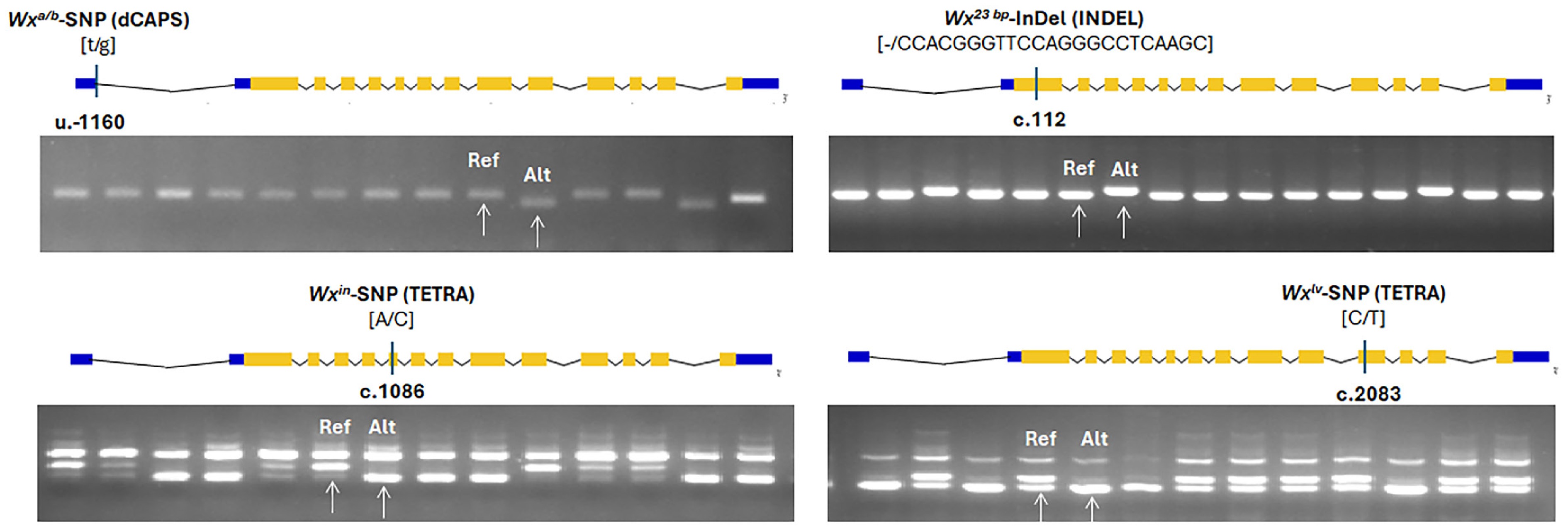
Figure 1.
Allele type and position of candidate FNPs of Waxy (Wxa/b, Wx23 bp, Wxin, Wxlv) alleles, and corresponding PCR results. The UTRs, exons, and introns of different genes are represented by blue boxes, orange boxes, and fold lines, respectively. The position of each variation is labeled as the relative distance to the ATG in the UTRs (u) and CDS (c). Allele variation of each gene is shown in the square bracket, with the front nucleotide variation representing Nipponbare (Reference) Allele and the back nucleotide variation representing non-NIP-type. The white arrows denote the band pattern of the reference (Ref) allele and alternative (Alt) allele, respectively.
A derived cleaved amplified polymorphic sequence (dCAPS) marker was constructed for the Wxa/Wxb allele variant. PCR products were subjected to gel electrophoresis, allowing allele discrimination based on banding patterns between reference alleles (Ref) and alternative alleles (Alt) identified in the Malaysian varieties (Fig. 1 and Supplementary Table S1). Based on the published Wx allele variations (Supplementary Fig. S1), five Wx alleles were identified among the Malaysian varieties, namely Wxlv (five varieties), Wxa (23 varieties), Wxin (23 varieties), Wxb (four varieties), and wx (four varieties).
Association of Wx alleles with AAC, and physicochemical properties in Malaysian rice varieties
-
We focused on the trait comparison of AAC, GPC, RVA, and DSC analysis among allele groups of the Wx gene (Fig. 2, Supplementary Table S1, and Supplementary Figs. S2 & S3). The analysis revealed significant differences in apparent amylose content (AAC) among the five Wx alleles (p < 0.05, Tukey's test). The Wxlv and Wxa alleles exhibited the highest AAC values, with means of 22.84% and 21.13%, respectively, and were grouped under statistical group A, indicating their classification as high-amylose types. The Wxin allele also clustered within group A, though it presented a moderately lower AAC value of 18.41%. In contrast, the Wxb allele showed a significantly lower AAC of 12.79% and was categorized as group B. The wx allele, consistent with a waxy phenotype, exhibited the lowest AAC at 3.62% and was assigned to group C. Interestingly, one genotype within the Wxin group displayed low amylose content, making it a promising candidate for in-depth functional and genetic analysis.
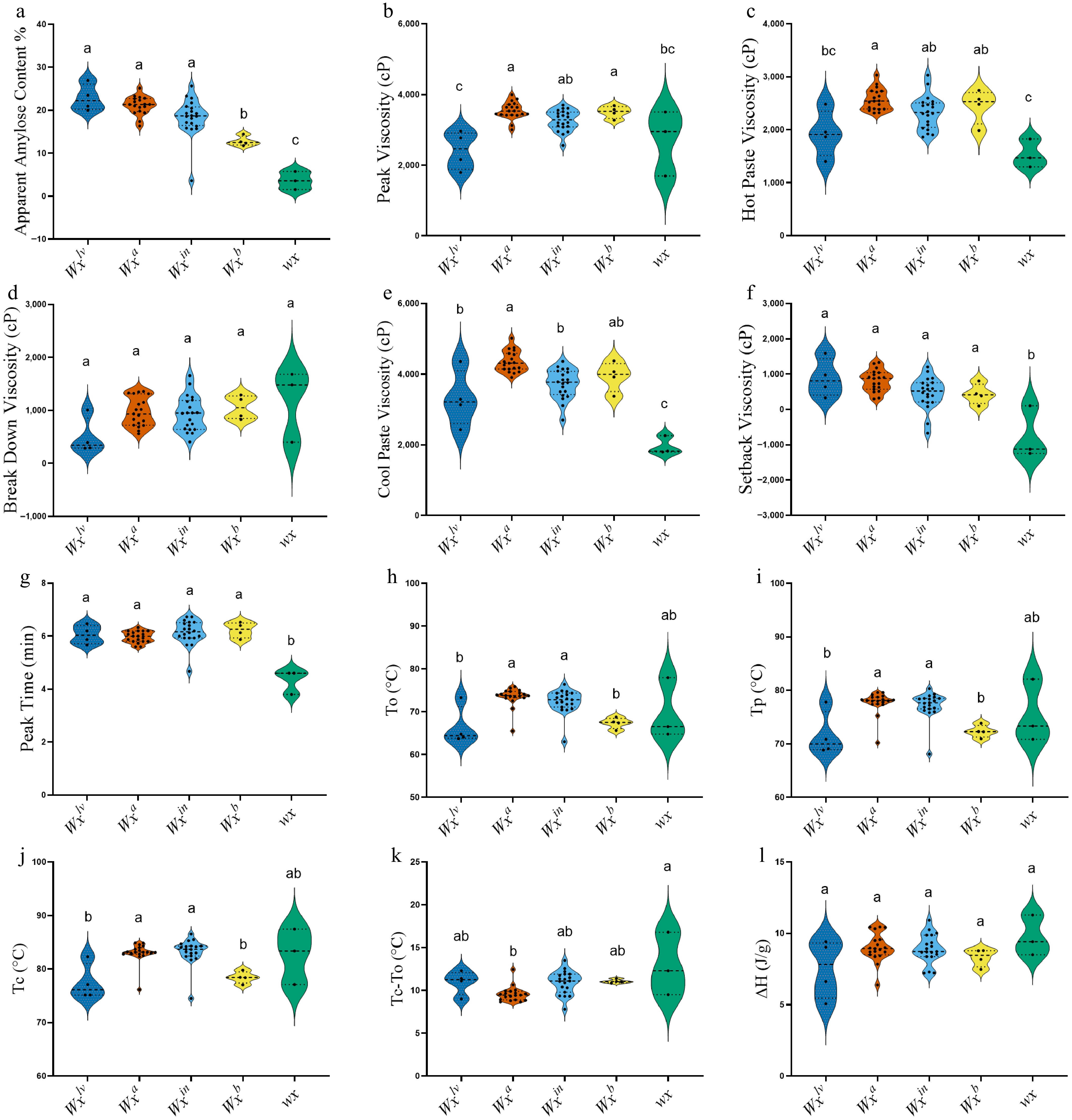
Figure 2.
Violin plot of Malaysian rice varieties grouped by different Wx alleles (Wxlv, Wxa, Wxin, Wxb, and wx), showing variation in (a) apparent amylose content, and (b) physicochemical parameters of RVA (Rapid Visco Analyzer), Peak Viscosity (PKV), (c) Hot Paste Viscosity (HPV), (d) Break Down Viscosity (BDV), (e) Cool Paste Viscosity (CPV), (f) Setback Viscosity (SBV), (g) Peak Time (PT), and DSC (differential scanning calorimetry), (h) To, (i) Tp, (j) Tc, (k) Tc–To, (l) ΔH (J/g). Significant statistical differences exist between groups with no shared character above the columns.
RVA analysis revealed substantial differences in starch pasting behavior among the Wx alleles. PKV was highest in Wxa (3,541.4 cP) and Wxb (3,503.3 cP), both of which were categorized in group A. The lowest PKV was recorded in Wxlv (2,422 cP), classified under group C. HPV followed a similar trend, with Wxa exhibiting the highest value (2,567.8 cP) and wx showing the lowest (1,531 cP), which was grouped as C. BDV, representing the difference between peak and trough values was greatest in wx (1,187 cP), indicating lower paste stability and higher susceptibility to shear thinning. CPV, representing the starch paste's ability to retrograde upon cooling, was highest in Wxa (4,352.9 cP), while wx displayed the lowest CPV (1,962 cP), consistent with a weak tendency toward retrogradation. SBV values further confirmed this behavior, with positive values in Wxlv (884 cP) and Wxa (811.5 cP), and a distinctly negative value in wx (–756 cP), reflecting minimal reassociation of amylose chains. Peak time (PT) varied among alleles, with wx showing the shortest time to reach peak viscosity (4.33 min), and Wxin the longest (6.14 min).
DSC analysis provided further insight into the thermal transitions of starch associated with different Wx alleles. The onset (To), peak (Tp), and conclusion (Tc) temperatures were notably higher in Wxa (To = 73.48 °C, Tc = 83.05 °C), and wx (To = 69.77°C, Tc = 82.63 °C), indicating more delayed gelatinization. In contrast, Wxlv exhibited the lowest gelatinization temperatures (To = 66.47 °C, Tc = 77.43 °C), suggesting earlier granule disruption during heating. The gelatinization temperature range (Tc–To) was widest in wx (12.87 °C), placing it in group A, and indicating a more heterogeneous starch crystalline structure. Narrower ranges were observed in Wxa (9.56 °C) and Wxin (10.91 °C), suggesting more uniform starch properties. The enthalpy change (ΔH), which reflects the energy required for starch gelatinization, did not differ significantly among alleles, with all variants statistically grouped as category A. However, the wx allele exhibited the highest ΔH value (9.73 J/g), suggesting a more ordered or compact crystalline structure within the endosperm starch.
In-depth comparison of waxy rice varieties
-
Based on the AAC content, four waxy rice varieties from Malaysian varieties were identified, and three harbor the known wx allele (PPM430, PB411, PS434), while the remaining one belongs to the Wxin group (PT438). These varieties provide an opportunity to further understand the differences in starch composition and its physicochemical properties between waxy and non-waxy rice. The four waxy Malaysian rice varieties were compared with the Wxin non-waxy reference cultivar KATUNG (KT405) using Apparent Amylose Content (AAC), Gel Consistency (GC), Rapid Visco Analyzer (RVA), and Differential Scanning Calorimetry (DSC) as primary indicators. Also, grain protein content (GPC) was also analyzed to learn the impact of low AAC on protein accumulation (Fig. 3).
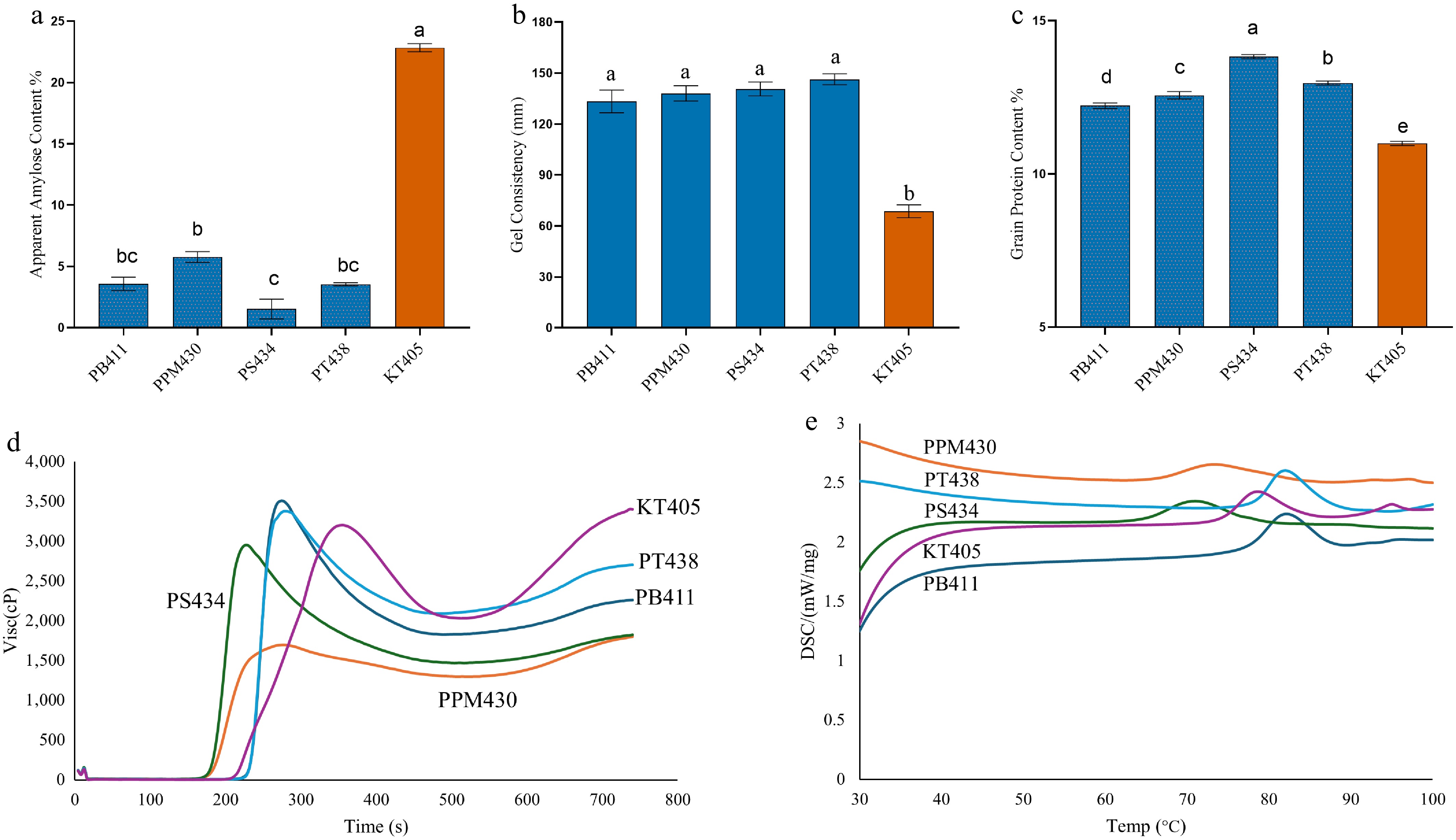
Figure 3.
(a) Apparent amylose content, (b) gel consistency, (c) grain protein content, (d) RVA (Rapid Visco Analyzer), and (e) DSC (Differential Scanning Calorimetry) graphs of Malaysian waxy rice varieties compared with KT405 (Wxin). PB411 (PULUT BURUNG), PPM430 (PADI PULOT MELAYANG), PS434 (PULUT SELEMYAM), PT438 (PULUTAN), KT405 (KATUNG). Significant statistical differences exist between groups with no sharing character above the columns (a and b).
Significant variation in AAC was observed among the tested varieties (Fig. 3a). The non-waxy reference cultivar KATUNG (KT405) exhibited the highest AAC value (22.83%), statistically grouped as A, consistent with its classification as a typical non-waxy variety. Among the waxy genotypes, PPM430 recorded the highest AAC (5.73%), categorized in group B. PT438 (3.57%) and PB411 (3.55%) were grouped as BC, indicating substantially reduced amylose levels, while PS434 displayed the lowest AAC (1.54%), assigned to group C, reflective of a strong glutinous phenotype.
Significant differences in gel consistency were observed among the studied varieties (Fig. 3b). The non-waxy reference variety (KT405) exhibited the lowest GC value (~70 mm), statistically grouped as B, consistent with its classification as a typical non-waxy variety with firmer gel texture. In contrast, all waxy alleles displayed markedly higher GC values (> 120 mm), reflecting the softer gel characteristic of waxy starch. Among them, PS434 and PT438 recorded the highest GC, both exceeding 140 mm, followed closely by PB411 and PPM430, which showed similar values (~130 mm). These waxy genotypes were statistically assigned to the same group A, indicating no significant differences in gel consistency among them.
In contrast to AAC, GPC values exhibited an inverse trend (Fig. 3c). The highest GPC was observed in PS434 (13.83%), followed by PT438 (12.97%), PPM430 (12.57%), and PB411 (12.23%). KT405, the non-waxy variety, recorded the lowest protein content (11.00%), statistically grouped in group E, whereas PS434 was uniquely categorized in group A. This apparent inverse relationship between AAC and GPC suggests that the suppression of starch biosynthesis in waxy genotypes may alter carbon–nitrogen partitioning, thereby promoting increased protein accumulation in the grain.
The Rapid Visco Analyzer (RVA) was performed to investigate the viscosity of Malaysian varieties in four waxy rice varieties and a non-waxy variety (KT405). The RVA graph and corresponding RVA parameters, including PKV, HPV, BDV, CPV, SBV, and PT, are shown in Fig. 3d and Supplementary Table S2, respectively. The waxy rice varieties show a wide range of PKV, with PB411 having the highest PKV (3,506 cP), and PPM430 the lowest (1,697 cP). The non-waxy variety KT405 has a PKV of 3,199 cP, which is within the range of the waxy varieties. The HPV, which indicates the stability of the starch paste under heat and shear, varies significantly between waxy varieties. PT438 has the highest HPV (2,092 cP), while PPM430 has the lowest (1,297 cP). Although the non-waxy variety had the highest HPV (2,198 cP) compared to other waxy varieties, it was lower than PT438. The BDV parameter reflects the stability of the starch paste during cooking. PB411 shows the highest BDV (1,680 cP), indicating less stability, whereas PPM430 has the lowest (400 cP), suggesting greater stability. The CPV, which indicates the starch's ability to form a gel upon cooling, is highest for non-waxy, KT405 (3,569 cP), and lowest for PPM430 (1,800 cP). Variety PT438 among waxy varieties had the highest CPV (2,704 cP). The SBV, which measures the retrogradation tendency, is negative for most waxy varieties, indicating less retrogradation. The non-waxy variety KT405 has a positive setback (200 cP), indicating a higher tendency for retrogradation. The peak time varies among the varieties, with KT405 for non-waxy varieties having the longest peak time (5.93 min), but waxy varieties had lower values with ranging between 3.8–4.67 min.
Differential Scanning Calorimetry (DSC) analysis was performed to investigate the thermal properties of four Malaysian waxy rice varieties. The onset temperature indicates the beginning of the gelatinization process, as shown in Fig. 3e and Supplementary Table S3. The waxy varieties showed a wide range of To values. The highest To was observed in PB411 (77.95 °C), indicating a delayed gelatinization onset. The lowest To was recorded for PS434 (64.8 °C), indicating a quicker onset of gelatinization. The non-waxy variety (KT405) exhibited a To of 74.35 °C, slightly higher than most of the waxy rice varieties. The peak temperature represents the point of maximum gelatinization. The highest Tp was recorded for PB411 (82.1 °C), while the lowest was for PS434 (70.85 °C). The non-waxy rice (KT405) showed a Tp of 78.35 °C, relatively higher than the average of the waxy varieties. Conclusion Temperature (Tc) marks the end of the gelatinization process. The highest Tc was observed for PB411 (87.45 °C), and the lowest was for PS434 (77.1 °C). The non-waxy variety (KT405) had a Tc of 83.7°C. This range (Tc–To) represents the duration of the gelatinization process. The widest range was noted in PPM430 (16.8 °C), indicating prolonged gelatinization. The narrowest range was in non-waxy KT405 (9.35 °C), suggesting a more rapid transition. Enthalpy Change (ΔH) reflects the energy required for gelatinization. The highest ΔH was observed in PB411 (11.28 J/g), indicating higher starch crystallinity and stability. The lowest ΔH was noted in PT438 (7.22 J/g), implying lower crystalline stability. The non-waxy variety (KT405) displayed a ΔH of 9.88 J/g.
Waxy rice varieties generally show lower onset and peak temperatures, indicating faster gelatinization compared to non-waxy varieties. The enthalpy change (ΔH) was notably higher in waxy varieties, suggesting higher crystalline amylopectin content. In contrast, the non-waxy variety exhibited a comparatively lower ΔH, reflecting higher amylose content, which contributes to its more gradual gelatinization profile.
Discovery of a new Waxy allele
-
The functional marker sets well dissected the allele variation of the Wx gene among the Malaysian varieties. When comparing the genotype of four waxy rice varieties, it was found that PT438 shared the allele type of Wxin, which is not consistent with the known allele contribution (Supplementary Fig. S1, and Supplementary Table S4). This inconsistency suggests that PT438 might have undergone significant genetic alterations within the Wx gene, making it different from the other waxy rice varieties. Further sequencing was conducted to explore the genetic differences between PT438 and the other waxy genotypes. Six primers were initially used to amplify the entire Wx gene for sequencing analysis (Supplementary Fig. S4 and Supplementary Table S5).
A simple nucleotide polymorphism (SNP) variation was identified in exon 6 at position 62 from the sequencing result. This variation involves a nucleotide substitution, where adenine (A) is replaced by cytosine (C) at this specific location (Supplementary Fig. S4a). Notably, this variation is identical to the Wxin mutation, which has been revealed by the functional marker. The Wxin mutation is known to influence AAC moderately, and our identified SNP variation could potentially have a comparable effect on starch synthesis pathways. Upon sequencing, primers 1 to 4 successfully amplified regions of the Wx gene in PT438, while primers 5 and 6 did not produce any bands. This result suggested that a segment at the end of the Wx gene might be deleted in PT438. To confirm this hypothesis, additional primers were designed to cover the entire gene, including the suspected deletion region. Three specific primers, P1-FR, P2-FR, and P3-FR, were utilized for this purpose (Fig. 4a and Supplementary Table S5). Primers P1-FR and P2-FR did not produce any bands in PT438, but P3-FR produced a sharp band, confirming the presence of a large deletion towards the end of the Wx gene (Fig. 4b).
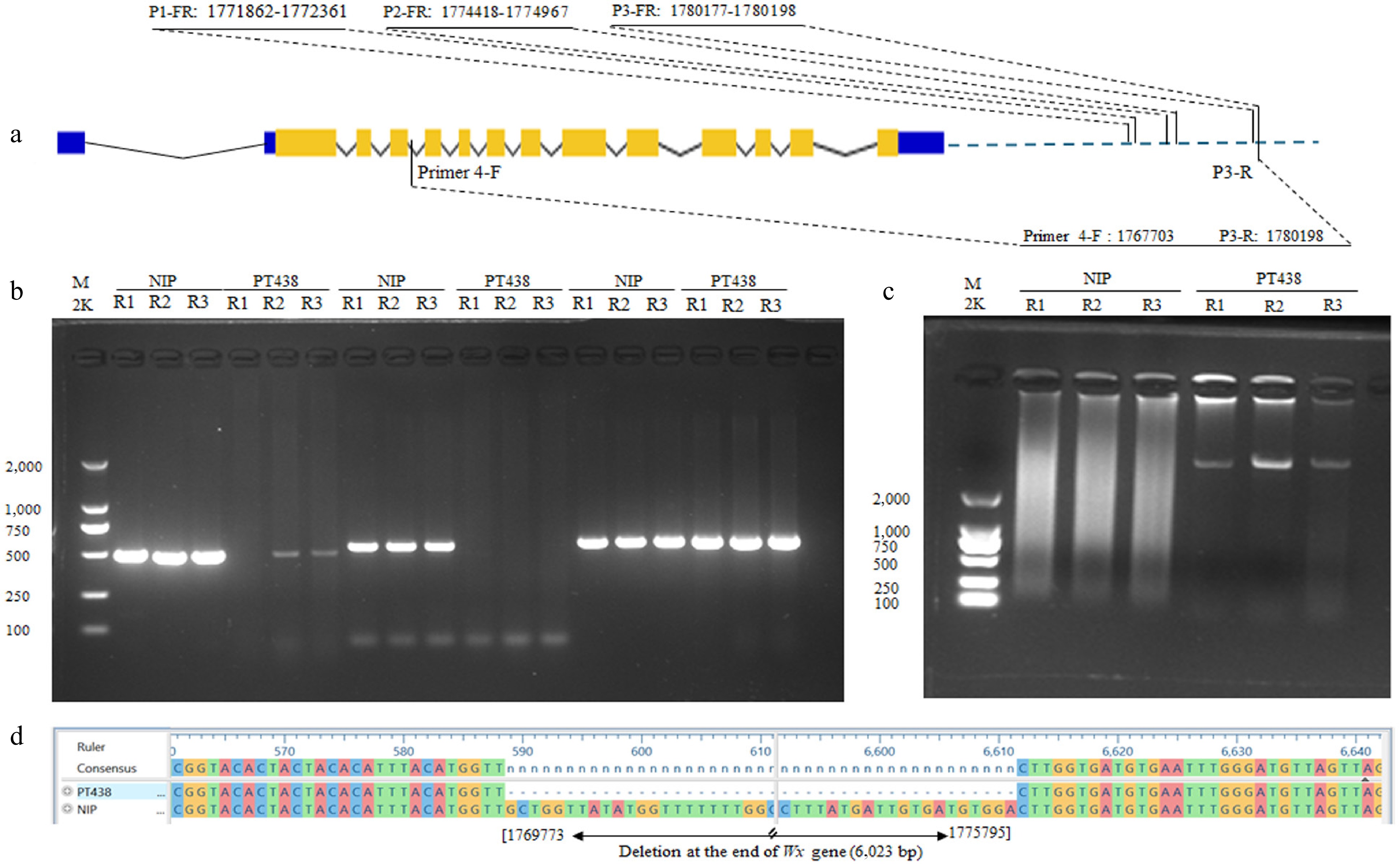
Figure 4.
(a) Deletion mapping, and (b) PCR amplification profile of the Malaysian rice variety PT438 (PULUTAN) using three primer pairs (P1-FR, P2-FR, and P3-FR) at the downstream of waxy gene. The position of each primer is shown on the gene structure, and NIP was amplified as a control. (c) and (d) are PCR amplification profiles and sequencing results showing a 6,023 bp deletion at positions 1769773−1775795 of the Malaysian rice variety PT438 (PULUTAN) compared with NIP. The promoters, UTRs, exons, and introns of the Waxy gene are represented by blue lines, blue boxes, orange boxes, and hat lines, respectively. R1, R2, and R3 are three repeats for each PCR amplification profile.
Primers 4-F, toward P3-R, targeting the region at positions 1767703-1780198, successfully produced a band in PT438 but not in Nipponbare, indicating the primer pairs cover the whole deletion region (Fig. 4c). Based on the sequencing result of available band, a 6,023 base-pair deletion was identified at the end of the Wx gene in PT438 (Fig. 4d). This discovery suggests that the Wx gene in this variety has undergone a significant structural modification, resulting in the deletion of a large segment. This large deletion potentially affects the amylose synthesis pathway, leading to altered starch properties and unique cooking qualities in PT438. Additionally, this deletion has led to the identification of new alleles of the Waxy gene in this variety, further differentiating PT438 from other waxy rice varieties, and the new allele was designated as wx6,023.Del hereafter.
To assess the function of proteins in different waxy rice varieties, including PT438, and to compare them with known Waxy alleles such as Wxlv, Wxa, Wxb, Wxin, SDS–PAGE was performed for total protein and starch granule-bound GBSSI (Fig. 5). The varieties analyzed include NIP (I1 with Wxb), PS434 (I2 with wx), PB411 (I3 with wx), PPM430 (I4 with wx), PT438 (I5&I6 with wx6,023.Del), MR167 (I7 with Wxlv), MR84 (I8 with Wxlv), PL425 (I9 with Wxlv), KT405 (I10 with Wxin), MR69 (I11 with Wxa), MR185 (I12 with Wxb), ZJ116 (I13 with Wxa), 9311 (I14 with Wxb).
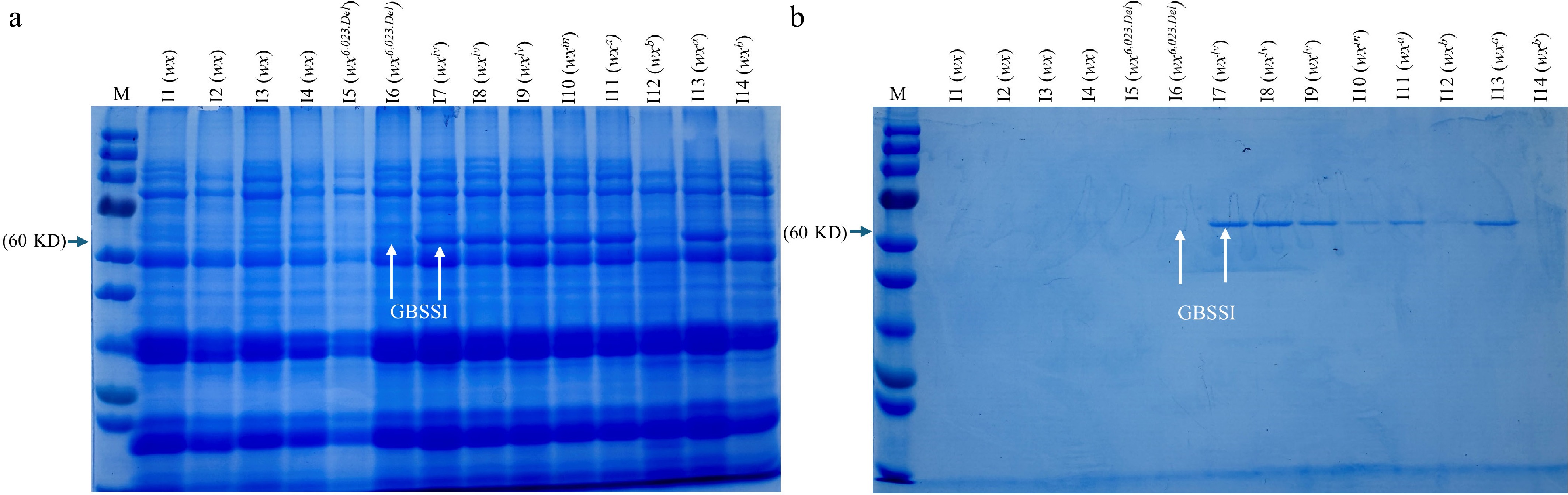
Figure 5.
SDS–PAGE assay of (a) total grain proteins and (b) starch granule-bound GBSSI in mature grains of NIP (I1 with Wxb), PS434 (I2 with wx), PB411 (I3 with wx), PPM430 (I4 with wx), PT438 (I5&I6 with wx6,023.Del), MR167 (I7 with Wxlv), MR 84 (I8 with Wxlv), PL425 (I9 with Wxlv), KT405 (I10 with Wxin), MR69 (I11 with Wxa), MR185 (I12 with Wxb), ZJ116 (I13 with Wxa), 9311 (I14 with Wxb). White arrows denote the protein level of wx6,023.Del and Wxlv genotype.
The SDS–PAGE results provide insights into the protein profiles of the tested varieties (Fig. 5). The variety PT438 showed no detectable bands for both total protein and GBSSI, the same as other waxy varieties (PS434, PB411, and PPM430), indicating that it did not produce this protein under the assay conditions. In contrast, other non-waxy varieties exhibited distinct bands corresponding to their respective protein profiles. This lack of bands in PT438 suggests that the 6,023 bp deletion affects both the expression of total protein and GBSSI, particularly in the context of starch biosynthesis pathways. The absence of GBSSI in PT438, a key enzyme in amylose synthesis, further supports the hypothesis that this variety may be deficient in amylose production, consistent with its classification as a new waxy rice variety.
-
The analysis of Wx allelic variation in Malaysian rice varieties revealed clear and consistent associations with key starch quality traits, particularly apparent amylose content (AAC) and pasting properties, reaffirming the central role of the Wx gene in determining rice eating and cooking quality.
As expected, apparent amylose content (AAC) varied significantly across the five Wx alleles. These findings align with previous studies demonstrating the functional hierarchy of Wx alleles in controlling GBSSI expression and amylose biosynthesis in the endosperm. The Wxlv and Wxa alleles, known to be more transcriptionally active, supported higher AAC levels and were grouped as high-amylose types. In contrast, Wxb, common in many Japonica varieties, consistently led to reduced AAC, while the wx null allele, characterized by loss-of-function mutations, produced the expected waxy phenotype. This pattern confirms that the Wx allele type is a strong genetic determinant of starch composition and can be used as a reliable marker in breeding for specific amylose content targets[21].
The influence of Wx alleles on RVA pasting parameters was substantial, highlighting their role in modulating functional starch behavior. The Wxa and Wxb alleles exhibited the highest peak viscosity (PKV) and cool paste viscosity (CPV), indicative of good paste formation and a strong tendency toward retrogradation. The high breakdown value observed in wx further underscores its instability under heat and shear, aligning with the soft, sticky texture preferred in certain culinary applications such as sticky rice and desserts[22].
DSC data added another dimension to these insights. While enthalpy changes (ΔH) did not vary significantly among alleles, gelatinization temperatures (To, Tp, Tc) showed distinct differences. Moreover, the wx allele was associated with the widest gelatinization temperature range (Tc–To) and the highest ΔH value, implying a more heterogeneous starch structure and a potentially more complex gelatinization process, despite its waxy nature[23].
Taken together, these results reinforce the pivotal role of Wx allelic diversity in shaping rice starch quality. The strong and consistent associations with AAC and RVA traits suggest that Wx genotyping can serve as a practical selection tool in rice breeding programs aimed at tailoring cooking properties to consumer preferences. For example, Wxb and Wxa may be desirable for general table rice types with firmer texture and better retrogradation, whereas wx suits specialty applications requiring sticky or soft textures. Meanwhile, Wxlv, although linked with high AAC, exhibits unique pasting and thermal characteristics that warrant further investigation, possibly involving detailed molecular analysis of its transcript levels or GBSSI enzymatic activity[21].
Insight from the comparison of waxy rice varieties
-
The comparative analysis between Malaysian waxy rice varieties and the non-waxy reference KATUNG (KT405) offers critical insights into how the waxy phenotype affects starch composition, pasting behavior, and thermal properties. The results align well with the established biochemical framework of Wx gene function, while also highlighting intra-group variability among waxy genotypes, suggesting the influence of secondary modifiers beyond the core Wx locus[24].
As expected, apparent amylose content (AAC) showed a marked distinction between waxy and non-waxy types. However, variation within the waxy group (e.g., higher AAC in PPM430 compared to PS434) suggests residual GBSSI expression or the influence of modifier genes such as OsAGPL2 or OsSSIIIa affecting amylose biosynthesis in a Wx background[25,26]. This phenotypic gradient among waxy types is consistent with the findings of Muto et al.[27], who reported that glutinous landraces in northern Laos exhibit substantial heterogeneity due to indica–japonica recombination and introgression in the genomic background surrounding the Wx locus.
In contrast to AAC, significant differences in GC and GPC were observed between non-waxy and waxy genotypes. The non-waxy reference variety (KT405), with detectable amylose content, exhibited the lowest GC value (~70 mm), indicating a firmer and less cohesive gel texture. Conversely, all waxy alleles exhibited markedly higher GC values (> 120 mm), corresponding to softer gel textures and enhanced stickiness, in agreement with previous reports that waxy starch, composed almost entirely of amylopectin, produces a tender and cohesive gel structure[28]. Among the waxy varieties, the lack of significant differences in GC (all clustered in group A) suggests a uniform starch behavior, though slight variation was noted, with PS434 and PT438 showing the highest GC (> 140 mm). Such variation may reflect subtle differences in amylopectin fine structure[29].
Grain protein content (GPC), on the other hand, was significantly higher in waxy varieties, particularly PS434 (13.83%), while the non-waxy variety (KT405) recorded the lowest GPC (11.00%). This inverse relationship between AAC and GPC supports the hypothesis that starch biosynthesis suppression may redirect assimilates toward protein accumulation, as previously reported in the glutinous rice study[22].
The RVA analysis further distinguished waxy and non-waxy profiles. While the non-waxy variety (KT405) had a high peak viscosity (PKV) (3,199 cP), some waxy varieties, such as PB411 exceeded it (3,506 cP), highlighting the structural diversity among waxy types. The negative setback values observed in most waxy varieties further confirm their reduced retrogradation capacity, consistent with their low amylose content. These results align with previous findings by Thomas et al. and Fei et al., who reported weak gel formation and poor textural stability in glutinous rice due to minimal amylose re-association during cooling[30,31].
DSC analysis confirmed that thermal behavior is tightly linked to starch composition. PB411 displayed the highest To, Tp, Tc, ΔH, and heat flow, suggesting greater starch crystalline stability, despite its waxy phenotype. This unique profile sets PB411 apart from other waxy varieties and may reflect a higher ratio of short-chain amylopectin (DP6–12), known to form tightly packed crystallites that resist gelatinization, as described by Tester & Morrison[32]. Interestingly, the non-waxy variety (KT405) had a moderate ΔH and broader gelatinization range, suggesting a more heterogeneous starch granule structure composed of both amylose and amylopectin, leading to a gradual gelatinization process. These properties are highly relevant to textural quality, as broader gelatinization ranges, often resulting from amylose–amylopectin interactions and heterogeneous granule structures, can influence cooking uniformity and textural quality[33].
Structural deletion in the Wx gene became a new choice for manipulating eating quality
-
The functional marker analysis revealed that PT438 exhibits a unique profile compared to other waxy rice genotypes, and further sequencing confirmed the presence of a SNP at exon 6 (A→C) and, more notably, a large terminal deletion in the distal regions of the gene. This deletion is of particular interest, as it removes a substantial portion of the Wx gene, which likely includes critical exonic and regulatory sequences. Such large deletions have been rarely documented, and the wx6,023.Del allele could represent a novel class of Wx mutations. Previous studies have described various null/weak alleles of Wx, including small indels, point mutations, or splicing variants (e.g., wx, Wxb, Wxin) that reduce or eliminate GBSSI expression and result in a low amylose phenotype[21]. However, the extent of structural disruption observed in wx6,023.Del suggests a more profound effect on gene function and downstream starch metabolism.
Supporting the structural findings, SDS–PAGE analysis further validated the functional consequences of the wx6,023.Del allele. PT438 showed no detectable bands for granule-bound starch synthase I (GBSSI), consistent with the complete loss-of-function of the Wx gene. This was comparable to other waxy varieties (PS434, PB411, and PPM430), which also lacked GBSSI bands, confirming the link between waxy phenotype and GBSSI protein deficiency. In contrast, non-waxy varieties harboring functional Wxlv, Wxa, Wxin, or Wxb alleles showed distinct GBSSI bands, reflecting active protein synthesis.
The genetic and protein-level data collectively explain the low amylose content (AAC) observed in PT438, reinforcing the role of GBSSI as the core enzyme responsible for amylose synthesis in rice endosperm[34]. The novel wx6,023.Del allele, by eliminating GBSSI production, effectively classifies PT438 as a waxy genotype. Interestingly, although functionally similar to the classic wx allele, the underlying mechanism—a large deletion—represents a unique molecular basis.
The phenotypic implications of this deletion are reflected in the RVA and DSC profiles of PT438, including low final and setback viscosities and late gelatinization, all typical of waxy starch behavior. However, further functional validation through transcriptomic analysis or GBSSI activity assays would help elucidate whether the deletion affects additional elements of the starch biosynthetic pathway, such as interactions with branching enzymes or starch synthases.
The discovery of the wx6,023.Del allele adds to the growing list of Wx gene variants contributing to rice starch quality diversity. From a breeding perspective, PT438 offers a valuable germplasm resource for developing glutinous rice with specific cooking or industrial properties, such as rapid gelatinization, low retrogradation, and high stickiness. Moreover, the distinct deletion pattern provides a robust molecular marker for precise introgression and identification in breeding populations.
-
A novel null wx allele in Malaysian rice could reflect local adaptation or historical introgression from neighboring regions like Thailand or Indonesia, both known for sticky rice traditions[35]. In conclusion, the identification of the novel wx6,023.Del allele in the Malaysian rice variety PT438 (PULUTAN) represents a significant advancement in understanding the genetic regulation of low amylose content (AAC) in rice. The discovery of a unique nucleotide change at Ex6-62 (A/C) and a large 6,023 bp deletion spanning the terminal exons of the Wx gene highlights the complexity of genetic variation affecting cooking quality. The non-functional granule-bound starch synthase (GBSSI), encoded by wx6,023.Del, contributes to the Null AAC phenotype, providing valuable insights into the molecular mechanisms underlying soft rice varieties. These findings not only deepen our knowledge of Wx gene expression but also open new avenues for breeding rice cultivars with improved eating and cooking qualities. Further research to explore and manipulate this novel allele may prove instrumental in developing high-quality rice varieties tailored to consumer preferences.
This work was supported by grants from Jiangsu Province Government (BE2022336, BZ2021017, and 20KJA210002) and the grant from the Ministry of Science and Technology of the People's Republic of China (DL2022014009L).
-
The authors confirm contributions to the paper as follows: study conception and design: Yeilaghi H, Zhang L, Liu Q; data collection: Yeilaghi H, Fatima I, Qazi M, Qin B, Bian Z, Zhang L; draft manuscript preparation: Yeilaghi H, Wong M, Yu L, Zhang C, Arzani A, Zhang L, Liu Q. All authors reviewed the results and approved the final version of the manuscript.
-
All data supporting the findings of this study are available within the paper and its supplementary data published online.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Variety name, marker variation, AAC, Rapid Visco Analyzer (RVA), Differential Scanning Calorimetry (DSC) Profiles of Malaysian Rice Varieties. For the allele types of Waxy, orange indicates the NIP (Ref) allele, and blue indicates the alternative (Alt) allele.
- Supplementary Table S2 Apparent Amylose Content (AAC), Gel Consistency (GC), and Grain Protein Content (GPC) with Rapid Visco Analyzer (RVA) data in four waxy varieties (PB411, PPM430, PS434, PT438) and KT405 (Wxin).
- Supplementary Table S3 Differential Scanning Calorimetry (DSC) data in four waxy rice varieties (PB411, PPM430, PS434, PT438) compared with KT405 (Wxin).
- Supplementary Table S4 Marker variation for key waxy allele types in waxy rice varieties (PB411, PPM430, PS434, PT438), compared with KT405 and NIP.
- Supplementary Table S5 Primer names and sequences for the Wx target gene.
- Supplementary Fig. S1 Genotypes and phenotypes of different Wx alleles.
- Supplementary Fig. S2 Rapid Visco Analyzer (RVA) Profiles of Malaysian Rice Varieties planted in China.
- Supplementary Fig. S3 Differential Scanning Calorimetry (DSC) Profiles of Malaysian Rice Varieties planted in China.
- Supplementary Fig. S4 Locations of six primer pairs (a) and PCR amplification profile (b) of the Waxy gene in Malaysian rice variety PT438 (PULUTAN).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yeilaghi H, Fatima I, Qazi M, Qin B, Bian Z, et al. 2025. Amylose variation reveals a novel null wx allele in Malaysian rice. Seed Biology 4: e025 doi: 10.48130/seedbio-0025-0023 

Amylose variation reveals a novel null wx allele in Malaysian rice
- Received: 05 August 2025
- Revised: 27 September 2025
- Accepted: 24 October 2025
- Published online: 22 December 2025
Abstract: Low amylose content is a key trait associated with the cooking and eating quality of rice (Oryza sativa L.), and is a major breeding target in the development of soft-textured rice varieties. In this study, 59 rice varieties from Malaysia were evaluated, a broad range of variation in apparent amylose content was observed. To investigate the genetic basis underlying this variation, a set of functional markers were employed to determine allelic types at the Waxy (Wx) gene. The results confirmed the central role of Wx in regulating apparent amylose content. Among the tested varieties, PT438 (PULUTAN) was identified as having a very low apparent amylose content phenotype. Genetic analysis revealed that this trait is governed by a novel Wx allele, designated wx6,023.Del, which encodes a non-functional granule-bound starch synthase I (GBSSI). Sequence comparison with the reference Wxb allele from Nipponbare showed a nucleotide substitution at exon 6 (Ex6-62, A/C; position 1,086 from ATG), along with a 6,023 bp deletion beginning at position 2,853 from ATG and extending to the end of the Wx gene, removing the terminal exons and generating a completely novel wx allele. PT438 exhibited soft texture traits, characterized by high gel consistency, low viscosity, and enthalpy, indicating a low-amylose starch structure. These findings provide new insights into the genetic and physicochemical mechanisms underlying low amylose content in Malaysian rice, including the discovery of wx6,023.Del enriches the Wx allele pool, and presents a valuable genetic resource for breeding rice varieties with improved eating and cooking quality.
-
Key words:
- Apparent amylose content /
- Null Waxy Allele /
- Rice /
- Malaysia /
- Eating and cooking quality