









-
Cereal crops, including rice (Oryza sativa), wheat (Triticum aestivum), barley (Hordeum vulgare), and maize (Zea mays), form the foundation of global food systems and are indispensable to human nutrition, agricultural economies, and food security worldwide[1,2]. These crops are cultivated across diverse agroecological regions, where their productivity increasingly faces pressure from climate variability and extreme weather events. Among the challenges associated with unstable harvest conditions, pre-harvest sprouting (PHS), defined as the premature germination of physiologically mature grains on the mother plant, has emerged as a major constraint in cereal production. PHS reduces grain yield, impairs end-use quality, disrupts market value, and threatens production stability in regions exposed to rainfall, prolonged humidity, and fluctuating temperatures during grain maturation and harvest[1,3,4].
A deeper consideration of PHS necessarily leads to the broader question of how cereals, which underpins both global nutrition and national food security[1,2], can preserve resilience amid accelerating climate variability. Building on this urgency, it has been thought that PHS in these food crops has revealed that it is not a simple trait, but a multifaceted phenomenon shaped by the interplay of genetics, physiology, and environment[1,5]. At its core lies the delicate balance between seed dormancy and germination potential[3,6], an evolutionary safeguard in wild relatives that has been gradually weakened through domestication and breeding for rapid germination and uniform growth[1,3]. While these improvements favored agricultural efficiency, the cumulative effects of domestication and modern breeding also rendered many contemporary varieties more susceptible to sprouting under unseasonal rains and humid conditions[7]. Recent advances in molecular biology, quantitative genetics, and genomics have uncovered key regulatory networks involving hormone signaling, transcriptional control, and seed coat characteristics that shape dormancy and sprouting responses. With climate models projecting more erratic rainfall in key growing regions, the relevance of PHS has expanded from being a sporadic production issue to a systemic threat to global food security. This shifting context underscores the need for an updated understanding of PHS that not only synthesizes decades of research but also integrates emerging molecular and physiological insights to guide breeding and management strategies tailored to the realities of a changing climate.
The aim of this review is to provide a comprehensive understanding of PHS in cereal crops, with rice serving as the principal reference system for detailed discussion. This review integrates current understanding on environmental triggers, physiological mechanisms, hormonal regulation, and genetic determinants that collectively govern seed dormancy and sprouting behavior across cereals. By drawing comparative evidence from wheat, barley, and maize, while emphasizing the substantial advances achieved in rice, the review highlights both conserved and crop-specific mechanisms underlying PHS resistance. It also addresses the domestication-related trade-offs that have weakened dormancy in modern cultivars and discusses how these insights can support the development of climate-resilient cereal breeding strategies, with particular relevance to rice improvement under increasingly unstable harvest conditions.
-
Environmental conditions are decisive drivers of PHS, while genetic factors define the baseline resistance of a cultivar. The actual incidence of sprouting at harvest is strongly shaped by rainfall, humidity, temperature dynamics, and the timing of these conditions relative to crop maturity[3,8]. Structural features of the ear or panicle further modulate how environmental stimuli translate into sprouting risk. Table 1 summarizes the principal environmental drivers of PHS and illustrates their manifestation in diverse cereal crops.
Table 1. Species-specific environmental parameters influencing pre-harvest sprouting in cereal crops.
# Environmental factor Rice Wheat Barley Maize 1 Rainfall and humidity ≥ 10–15 mm rainfall over several days with relative humidity
> 85% during late maturity strongly increases vivipary.Moderate rainfall with relative humidity > 80% near harvest promotes sprouting, especially in white-grained cultivars. Short rainfall events with high humidity rapidly induce sprouting due to shallow dormancy. Sustained rainfall that maintains kernel moisture for several days increases vivipary when husks are loose. 2 Temperature × moisture Warm temperatures > 25 °C combined with high moisture accelerate dormancy loss. Cool temperatures (10–20 °C) with prolonged moisture extend grain hydration and favor sprouting. Moderate temperatures (15–25 °C) with moisture rapidly activate germination. Warm and humid conditions slow kernel drying and favor vivipary. 3 Rainfall timing Rainfall after physiological maturity or during early after-ripening presents the highest risk. Rainfall during late grain filling and immediately before harvest increases PHS incidence. Rainfall close to harvest readily induces sprouting. Rainfall after black layer formation increases vivipary under exposed kernels. 4 Wetting-drying cycles Multiple wet-dry cycles destabilize dormancy and increase sensitivity to later rainfall. Alternating wet and dry conditions weaken dormancy within spikes. Repeated moisture fluctuations accelerate dormancy breakdown. Wet-dry cycles increase kernel moisture when husk protection is incomplete. 5 Duration of grain wetness Continuous spikelet wetness > 48 h strongly correlates with vivipary. Prolonged spike wetness enhances water uptake through glumes. Short wetness periods are sufficient to initiate germination. Extended kernel wetness occurs when husks retain moisture. 6 Morphology and architecture Hull tightness, hull thickness, and panicle openness regulate water entry. Spike compactness, glume closure, and pericarp permeability control hydration. Spike architecture and hull adherence influence moisture retention. Husk tightness, husk length, and ear orientation determine kernel exposure. Rainfall, particularly when combined with prolonged high relative humidity, constitutes the primary environmental trigger of PHS across cereal crops by sustaining kernel hydration long enough to reactivate embryo metabolism[3,8]. Previous studies show that 10–15 mm of rainfall is typically needed to induce sprouting in susceptible cultivars of wheat, although smaller amounts may still be sufficient under conditions of prolonged wetness and high relative humidity[1,8]. Barley, with its open spike structure and long awns, is even more vulnerable, as light but persistent rain often results in more water retention than short, heavy downpours that quickly run off[1,9]. Once water enters the grain, kernels swell, and metabolic processes resume, including enzyme production and radicle emergence, which signal the onset of germination. Relative humidity amplifies the effect of rainfall by prolonging moisture retention on the crop surface[8,10]. In the absence of substantial precipitation, relative humidity levels above 85%–90% can prolong moisture retention on the panicle, which maintains kernel hydration for a sufficient duration to initiate embryo reactivation and metabolic activity[11].
This is a major concern in growing regions affected by monsoon rains, where high humidity lingers after showers and panicles remain wet for extended periods[12]. In maize, ears with loose husk coverage are particularly at risk under humid tropical conditions, as kernels stay moist long after rainfall ends[13,14]. Prolonged humidity after rain is particularly damaging in wheat, barley, and rice because it prevents the rapid drying of spikes or panicles and maintains a moist microenvironment that promotes sprouting. This extended wetness is especially problematic in regions where post-rain humidity remains high, as grains in these cereals retain surface moisture for long periods. In wheat and barley, the architecture of the spike slows moisture loss, while in rice, the compact arrangement of spikelets within the panicle further delays drying. Even limited rainfall followed by persistent humidity can keep kernels hydrated long enough to initiate embryo reactivation and trigger sprouting. This explains why regions with scattered showers and extended periods of damp air, such as northern Europe for barley or East Asia for wheat and rice, experience higher PHS incidence than regions where a single heavy rain is followed by rapid drying[9]. Crop architecture also modifies the effects of rainfall and humidity. Wheat spikes that remain upright tend to shed water more effectively than spikes that droop, which trap moisture around the grains[8,11,15]. In barley, the presence of long awns increases water interception[16], while in rice, the compact arrangement of spikelets within the panicle allows droplets to accumulate and persist[12,17]. The tightness of glumes likewise plays a role across cereals, as looser glumes expose the kernel to more moisture. The interaction between structural traits and environmental conditions dictates the extent of water penetration, the permeability of seed coats, and the hormonal balance within kernels, which together determine how long grains remain hydrated and the likelihood of sprouting before harvest[1].
Temperature and moisture interact in ways that significantly influence the risk of PHS. In wheat, cool and moist weather after physiological maturity creates ideal conditions for sprouting[18]. Low temperatures retain moisture around the kernel for longer periods and also accelerate the decline of seed dormancy. Once dormancy weakens, even a short rain shower can cause germination. This pattern is often seen in northern Europe and Canada, where harvest overlaps with frequent cool rains or heavy dew that leaves the spikes wet for many hours[19,20]. High temperatures during grain filling produce the opposite outcome. Grains that mature under hot conditions often reach harvest with weak dormancy. When rain arrives, they sprout quickly and in a uniform manner[3,21]. In barley, where dormancy is already low, hot and dry grain filling reduces resistance even further. Malting cultivars show high levels of sprouting when rain falls late in the season. Moisture stress can complicate the picture[22]. Under drought, high temperatures sometimes strengthen resistance. Drought hardens the seed coat and dries the kernel more quickly, which makes it harder for water to re-enter[23]. In rice, drought before harvest can temporarily reduce vivipary in panicles[24]. However, once monsoon rains become persistent, this protection fades. In maize, heat and drought may delay water uptake when husks are tight, but extended rainfall soon overcomes this barrier, and sprouting begins inside the ear[25]. Temperature also plays a central role in after-ripening. Warm and dry conditions between maturity and harvest shorten the dormancy period, which leaves kernels more likely to sprout when rain arrives (Fig. 1). This outcome has been reported in Mediterranean wheat and barley, where dormancy loss occurs early, and unexpected showers cause severe damage. Cooler conditions slow after-ripening and extend resistance, although in humid areas such as tropical Asia, high atmospheric moisture can cancel this advantage in rice[1,26].
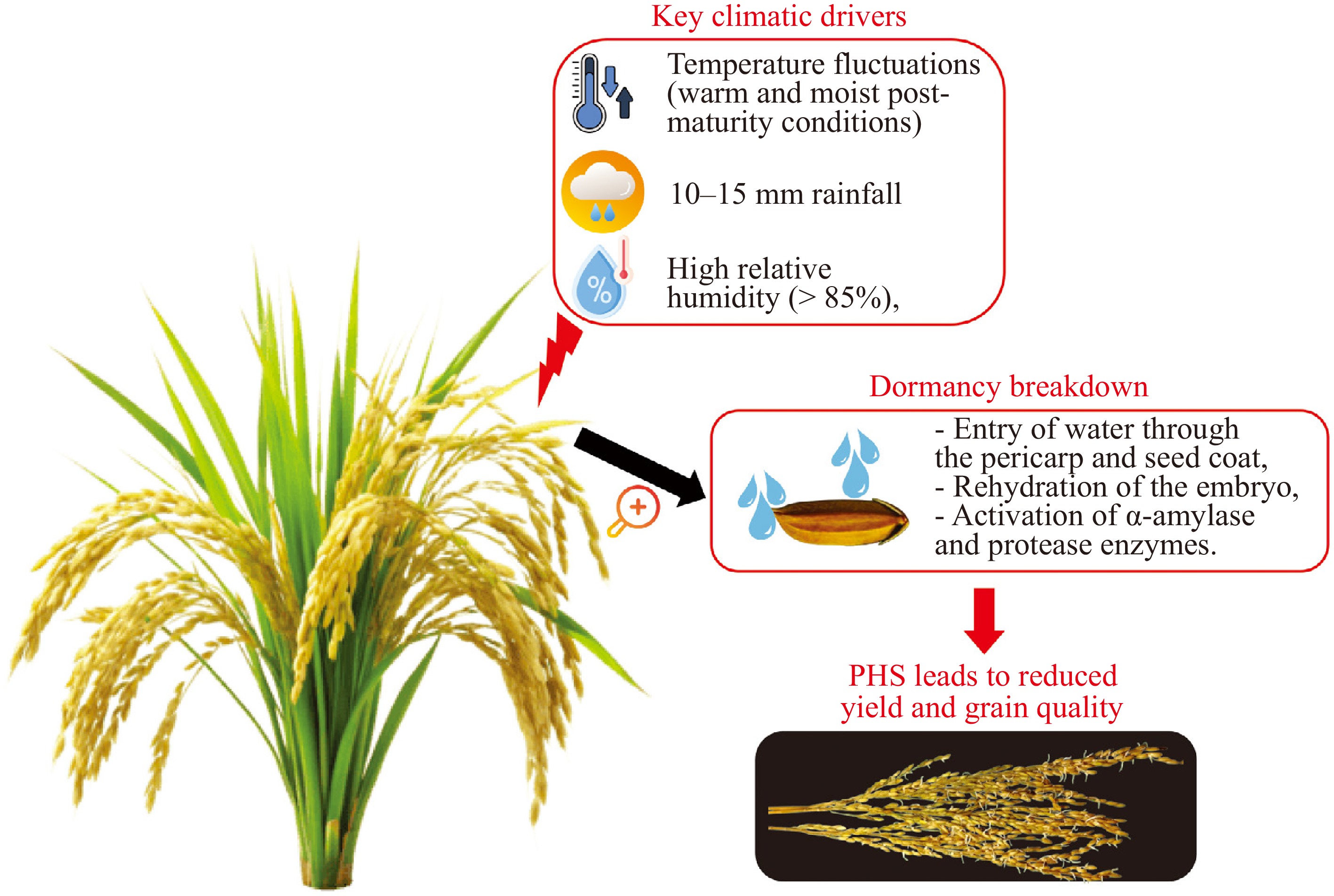
Figure 1.
Conceptual overview of pre-harvest sprouting in rice. Environmental triggers such as rainfall, humidity, and temperature affect moisture uptake by panicles and spikelets. Water infiltration through glumes and hulls reactivates metabolic enzymes and causes dormancy breakdown and premature germination. The physiological consequence of pre-harvest sprouting results in lower grain yield, reduced processing quality, and decreased storage stability.
The impact of temperature and moisture continues through the timing of rainfall during the final stages of grain development[1]. Rain that falls 10 to 20 d before harvest rarely triggers sprouting on its own, since kernels still hold some level of dormancy. Yet these early showers often shorten the dormancy period. When a later rainfall event takes place near harvest, seeds that were exposed earlier sprout within 1 or 2 d of continuous wetness[27]. In wheat, this sequence of early and late rain is a major cause of falling number reductions in Europe and North America[18]. In barley, the consequences are more severe, since low dormancy means that rainfall before maturity leaves the crop highly vulnerable if harvest is delayed[28]. In rice, the outcome depends on the variety. Susceptible lines lose dormancy quickly, so even light showers before harvest make them prone to vivipary once monsoon rains intensify[12,29]. In maize, rainfall timing is most critical in open-husked types. Showers that arrive close to harvest cause the highest risk, but earlier rainfall can still weaken dormancy and increase the chance of later sprouting[30]. These examples show that PHS risk depends not only on how much rain falls, but also on when it occurs and how temperature shapes dormancy. The combination of these factors explains why different cereals and different regions experience such variable levels of sprouting damage[1,9,31].
-
The physiological basis of PHS arises from the interaction of multiple processes that determine whether a mature grain remains dormant or initiates germination when exposed to moisture before harvest. PHS does not result from a single factor; instead, it emerges from the combined action of several mechanisms that regulate the transition between dormancy and germination. These mechanisms include the strength and duration of seed dormancy, the balance between abscisic acid and gibberellin signaling, the reactivation of metabolic enzymes, the protective contributions of hulls and other structural barriers, and the timing of dormancy release during after-ripening[4]. Together, these processes determine whether grains resist or succumb to sprouting, and their expression differs across cereal crops[1,5,6,31,32].
Dormancy serves as the most important physiological barrier preventing seeds from germinating on the mother plant[33,34]. It provides a period of developmental restraint that allows grains to withstand temporary wetting events before harvest. The strength, duration, and genetic basis of dormancy vary widely among cereals, and these differences largely determine the extent of PHS observed in the field[35]. In wheat, dormancy is strongly influenced by the seed coat and glume tissues, which limit both oxygen diffusion and water entry into the kernel[36]. Red-grained wheat cultivars show lower susceptibility to PHS than white-grained cultivars, a pattern that is most often attributed to tight genetic linkage between pericarp color loci and genes controlling seed dormancy[37]. In addition to linkage effects, pigment-related metabolic processes may also contribute indirectly to dormancy maintenance. Abscisic acid (ABA) biosynthesis originates from carotenoid precursors, and key intermediates in carotenoid metabolism, such as zeaxanthin epoxide, participate in the ABA biosynthetic pathway[5,38−40]. Enhanced activity of pigment biosynthesis pathways could coincide with higher ABA accumulation during seed maturation. Although the direct causal relationship between pigment content and ABA levels remains incompletely resolved, this metabolic convergence is consistent with the central role of ABA in reinforcing dormancy and suppressing premature germination[39−42]. Consequently, genetic linkage and shared metabolic pathways provide a coherent framework for understanding the association between red pericarp coloration and increased resistance to PHS in wheat. This explains why white-grained wheats in regions such as North America and Australia often suffer higher levels of PHS[43]. Furthermore, dormancy tends to be stronger in winter wheats grown under cooler environments, while spring wheats in warmer regions lose dormancy earlier, predisposing them to sprouting when rainfall coincides with harvest[44]. In barley, dormancy is also a key protective mechanism, but its role has been compromised by breeding priorities. Because the malting process requires rapid and uniform germination, barley breeding has historically favored reduced dormancy[45]. While this trait improves malting efficiency, it inadvertently increases the crop's vulnerability to PHS. Modern malting barley cultivars typically exhibit minimal or absent seed dormancy, which results in high susceptibility to PHS under cool and humid harvest conditions, characteristic of Northern Europe[46]. In contrast, landraces and wild barley accessions contain genetic variants that give the seeds stronger dormancy through greater sensitivity to ABA and slower activation of germination signals[42,47]. These traits serve as valuable sources for developing barley lines that resist sprouting before harvest. In rice, dormancy expression is highly variable and strongly dependent on genetic background. Indica varieties grown in tropical Asia often lose dormancy soon after maturity[12]. Thus, they become vulnerable to sprouting when the air turns humid during the monsoon season. Japonica varieties, particularly those grown in temperate regions, generally retain dormancy for longer, which provides some protection against sprouting in the panicle. However, the short duration of dormancy in many widely cultivated lines remains a major challenge, as rainfall near harvest readily induces germination on the panicle[6]. In maize, dormancy is inherently weak compared with wheat, barley, or rice[1,3,14]. Instead, the main safeguard against PHS is physical rather than physiological; particularly, the husk that surrounds the cob provides a barrier that limits water entry and protects kernels from prolonged wetness. Tight husk coverage can delay sprouting, while loose husks, which are common in tropical and subtropical varieties, allow rain and humidity to reach kernels more easily. Thus, in maize, husk morphology often determines whether ears remain sound or sprout during wet harvest periods[1,13]. These comparisons demonstrated that dormancy functions as the primary line of defense against PHS across cereals, but its strength and expression differ widely[1]. Wheat relies on genetic associations between seed coat pigmentation and dormancy loci, barley has traded dormancy for malting efficiency, rice shows sharp varietal differences linked to ecogeographic adaptation, and maize depends mainly on husk architecture. Understanding these crop-specific dormancy mechanisms is critical for designing breeding strategies that balance agricultural use with resistance to PHS.
The regulation of PHS is tightly linked to the interplay between two key phytohormones, especially ABA, which enforces seed dormancy[34,42,47], and gibberellins (GA), which promote germination[48,49]. The balance between these two hormones determines whether mature seeds remain quiescent or shift toward sprouting when exposed to favorable conditions. In wheat, ABA accumulates during seed maturation and plays a central role in establishing primary dormancy[50]. Dormant genotypes often show higher ABA levels in developing embryos and stronger sensitivity to the hormone compared with sprouting-susceptible lines[51]. Genes, such as TaVp-1 (an ortholog of maize Viviparous-1 and Arabidopsis [Arabidopsis thaliana] abscisic acid-insensitive-3), regulate ABA response pathways[52]; defective alleles in modern hexaploid wheat reduce ABA sensitivity and increase PHS risk[52]. Meanwhile, GA stimulates the synthesis of α-amylase in the aleurone layer, which then causes starch in the endosperm to break down. Wheat varieties with lower ABA responsiveness or higher GA sensitivity tend to lose dormancy rapidly and sprout under rainfall near harvest[53]. In barley, ABA similarly promotes dormancy, while GA drives germination. During the malting process, the GA-induced production of hydrolytic enzymes is deliberately exploited to mobilize starch reserves for brewing[54]. However, in the field, this same GA response under wet conditions accelerates PHS. In rice, the hormonal regulation of dormancy and sprouting is also mediated by ABA and GA, but with distinct genetic underpinnings[12]. Indica varieties grown in monsoon climates often display weaker ABA accumulation or reduced ABA sensitivity, which contributes to their susceptibility to vivipary[55]. Mutations in ABA biosynthetic genes, such as OsNCED3 (encoding 9-cis-epoxycarotenoid dioxygenase), increase the likelihood of sprouting on the panicle[56]. Japonica varieties typically retain higher ABA levels and longer dormancy, which provides stronger resistance under temperate growing conditions[57]. Studies have shown that rice vivipary mutants often have defects in ABA synthesis or ABA signal transduction. These defects highlight the essential role of this pathway in maintaining seed dormancy during the harvest period[58,59]. In maize, ABA is critical for maintaining dormancy in developing kernels, and mutants with impaired ABA biosynthesis or signaling exhibit precocious germination (the classic viviparous or vp mutants)[60]. For instance, vp14 mutants lack a key enzyme required for ABA production. As a result, they show weaker dormancy and tend to sprout directly on the cob[61]. Similarly, Vp1 (Viviparous1), the maize ortholog of wheat TaVp-1[62], encodes a transcription factor required for ABA-mediated gene expression[63]. Mutations in Vp1 produce kernels that fail to enter dormancy and germinate prematurely[14]. Although maize generally exhibits weaker physiological dormancy than wheat or rice[1,3,14], these hormonal pathways[64], combined with husk coverage, still play an important role in determining whether kernels resist sprouting under wet and humid harvest conditions. Across cereals, the physiological state of the seed can therefore be described as the outcome of an ABA-GA balance. High ABA accumulation and strong ABA sensitivity favor dormancy, whereas elevated GA activity and weaker ABA signaling favor germination[3,39]. This balance differs among crops and cultivars. Some maintain strong resistance to PHS, while others are far more vulnerable. Understanding this hormonal antagonism provides a foundation for breeding programs that aim to enhance sprouting resistance without undermining end-use quality.
Once dormancy is lost and moisture enters the kernel, PHS proceeds through the reactivation of metabolic pathways that are normally quiescent in mature seeds. The most notable physiological change is the induction of hydrolytic enzymes, particularly α-amylase, which breaks down starch in the endosperm, and various proteases that mobilize storage proteins[2]. These processes, while essential for germination and seedling establishment, are highly detrimental when they occur prematurely in the field. In wheat, the premature induction of α-amylase activity has direct consequences for grain processing quality[65,66]. Even a small percentage of sprouted kernels in a bulk lot can lower the falling number value, which is a key measure of α-amylase activity. Reduced falling numbers correspond to lower dough viscosity, decreased loaf volume, and poor bread-making performance[65]. This explains why millers and bakers place strong emphasis on sprouting resistance in wheat breeding. Hard red wheats with strong dormancy typically show lower α-amylase activity under wet conditions, whereas soft white wheats, which often lack strong dormancy, are more susceptible to enzyme activation and subsequent quality losses[67,68]. In barley, enzyme activity induced during PHS is essentially the same as that harnessed during the malting process[54]. GA secreted by the embryo stimulate the aleurone layer to produce α-amylase and β-glucanases, which degrade starch and cell wall polysaccharides[69]. While these changes are beneficial in brewing, they reduce grain soundness and feed quality when triggered in the field[54]. Malting barley, already selected for rapid enzyme induction, is particularly vulnerable. This creates a direct conflict between malting quality and sprouting resistance in barley breeding programs[16]. In rice, PHS leads to vivipary within the panicle, where kernels germinate before harvest[6,12]. Enzyme reactivation in rice reduces grain weight and nutritional quality, while also altering texture and eating quality after cooking. Studies have shown that sprouted rice accumulates high levels of α-amylase and proteases, which degrade starch and proteins required for stable grain storage[2]. For producers in monsoon climates, this enzymatic breakdown translates into significant post-harvest losses and reduced market value. In maize, sprouting on the cob activates α-amylase and proteases within individual kernels. Because maize is often harvested and stored at relatively high moisture levels, enzyme activation during PHS accelerates deterioration in storage and reduces seed viability for planting[13,64]. Hybrids with tight husk coverage delay water infiltration and slow enzyme induction, while open-husked genotypes show rapid kernel sprouting and enzyme activation when exposed to humid harvest conditions. The impact of enzymatic reactivation underscores why PHS is not simply a problem of yield loss, but also a major issue of grain quality. Elevated α-amylase levels reduce the processing value of wheat and barley, while protein and starch degradation lower the eating and storage quality of rice and maize[1]. Thus, understanding and mitigating enzyme activation during PHS remains a central priority in cereal improvement programs.
-
PHS is a quantitative trait regulated by multiple genes and hormone-responsive pathways that coordinate seed dormancy and germination. Comparative studies across cereal crops have revealed several conserved regulatory pathways controlling dormancy stability and sprouting resistance, although the specific genes and their functional contributions often differ among species[1,50,70]. Rice has emerged as one of the most extensively characterized systems for dissecting the molecular regulation of PHS[7,71]; however, key regulatory components have also been identified in wheat, barley, and maize.
First, ABA signaling represents a central regulatory axis controlling seed dormancy in cereals[5]. In rice, the transcription factor OsVP1, an ortholog of maize Vp1[63,72], maintains embryo quiescence during late seed development and is required for proper ABA responsiveness[73,74]. Loss-of-function mutations in OsVP1 result in frequent vivipary under humid conditions[73], which confirms its essential role in dormancy maintenance in rice[41,75,76]. Orthologs of Vp1 in wheat and barley also contribute to dormancy regulation, although their effects are influenced by breeding history and selection for end-use traits, such as malting quality in barley[62,77,78].
ABA biosynthesis represents another core layer of PHS regulation[5]. In rice, OsNCED3, which catalyzes a key step in ABA biosynthesis, plays a major role in promoting ABA accumulation in developing seeds and strengthening dormancy[56]. Allelic variation at this locus contributes to the divergence in dormancy strength between the indica and japonica subspecies[71]. Direct experimental evidence for the importance of ABA biosynthesis in rice PHS was provided by Fang et al., who identified viviparous mutants defective in carotenoid biosynthesis genes, including phytoene desaturase (OsPDS), ξ-carotene desaturase (OsZDS), carotenoid isomerase (OsCRTISO), and lycopene β-cyclase (β-OsLCY)[79]. Disruption of these genes reduced carotenoid-derived ABA production and caused severe PHS under humid field conditions, while ABA signaling capacity remained intact[79]. These findings established a causal link between ABA biosynthesis and the maintenance of dormancy in rice. Comparable ABA-related regulatory mechanisms have been reported in maize, where mutations affecting ABA biosynthesis genes, such as Vp14, lead to viviparous kernels that germinate prematurely on the cob[80].
Next, downstream of ABA accumulation, ABA-responsive transcription factors reinforce dormancy. For example, key regulators, such as ABSCISIC ACID INSENSITIVE 3 (ABI3), ABI4, and ABI5, mediate ABA-responsive gene expression and mutations in these genes reduce ABA sensitivity and promote vivipary[81]. In maize, Vp1 is functionally equivalent to ABI3 in Arabidopsis[72], while homologs of these ABA signaling genes in wheat contribute to natural variation in dormancy, though their effects differ depending on genetic background[8,82,83].
In contrast, GA promotes germination and acts antagonistically to ABA[41,48]. Overexpression of genes involved in GA biosynthesis (GA3 and GA20) accelerates germination[84], while loss-of-function alleles delay sprouting[85,86]. GA-dependent induction of hydrolytic enzymes, such as α-amylases (namely Amy1 and Amy2) in the aleurone layer, initiates reserve mobilization after dormancy release[87]. In other cereals, variation in α-amylase regulation in barley contributes to differences in sprouting behavior, but this layer primarily provides a comparative context for rice[88].
The locus qSD7-1 corresponds to the Rc gene, which encodes a basic helix-loop-helix (bHLH) transcription factor originally characterized for its role in pericarp pigmentation[75]. Functional analyses have demonstrated that Rc exerts pleiotropic effects on seed dormancy through modulation of ABA biosynthesis, partly via coordinated regulation of flavonoid metabolic pathways. Natural variation at Rc explains the strong association between red pericarp color and enhanced seed dormancy in weedy and wild rice, as well as reduced susceptibility to PHS[75]. Loss-of-function alleles selected during domestication confer white pericarp and weakened dormancy, which increases vulnerability to sprouting under humid harvest conditions[75].
Next, fine-scale modulation of dormancy persistence in rice involves DELAY OF GERMINATION1 (DOG1)-like regulators. It has been noted that the contribution of DOG1-like genes to seed dormancy is less prominent in rice than in Arabidopsis, which can be attributed to fundamental differences in dormancy regulatory architecture between the two species. In Arabidopsis, DOG1 functions as a central regulator of primary seed dormancy and exerts a strong genetic effect[89]. In contrast, seed dormancy in rice is governed by a polygenic regulatory system that integrates hormonal metabolism, transcriptional control, and grain structural characteristics[15,90]. Within this framework, DOG1-like homologs, including OsDOG1L, act as modulatory components rather than as primary determinants of dormancy[90]. Functional diversification among DOG1-like family members and redundancy across dormancy-related pathways attenuate the phenotypic impact of individual DOG1-like genes in rice[59]. Moreover, ABA biosynthesis and catabolic processes constitute dominant regulatory layers in rice dormancy control[6,56,90,91], which further constrains the relative contribution of DOG1-like factors. These observations indicate that the regulatory hierarchy governing seed dormancy differs between dicot and monocot lineages.
Additionally, recent studies have reported several additional regulators that expand the dormancy network. Among them, SEED DORMANCY4 (Sdr4) functions downstream of OsVP1 and coordinates late seed maturation processes that stabilize dormancy at physiological maturity[73,92]. Natural variation at Sdr4 correlates with regional rainfall patterns and exhibits adaptation during rice domestication[92,93]. The qSD1-2 locus encodes OsGA20ox2 and links GA biosynthesis to endosperm-imposed dormancy[94]. Other regulators, including PRE-HARVEST SPROUTING8 (PHS8) and PHS9, connect carbohydrate metabolism and redox homeostasis with ABA signaling[95]. Transcription factors such as SD6 and MOTHER OF FT AND TFL2 (MFT2) integrate temperature cues and ABA signaling into dormancy regulation[96−98], while the NF-YB1-SLRL2-bHLH144 module coordinates ABA-mediated control of both seed dormancy and grain quality[99].
Quantitative trait loci underlying pre-harvest sprouting resistance in rice
-
Considering the high collinearity of genetic loci among gramineous crops, we summarize current PHS-related quantitative trait loci using rice as a primary example. Genetic analyses confirm that PHS resistance in rice arises from a distributed QTL architecture (Fig. 2, Table 2). Numerous loci of small to moderate effect are distributed across almost all chromosomes, and each contributes partially to dormancy strength, hormone sensitivity, or grain protection. Resistance emerges from the combined action of these loci rather than from a single dominant determinant.
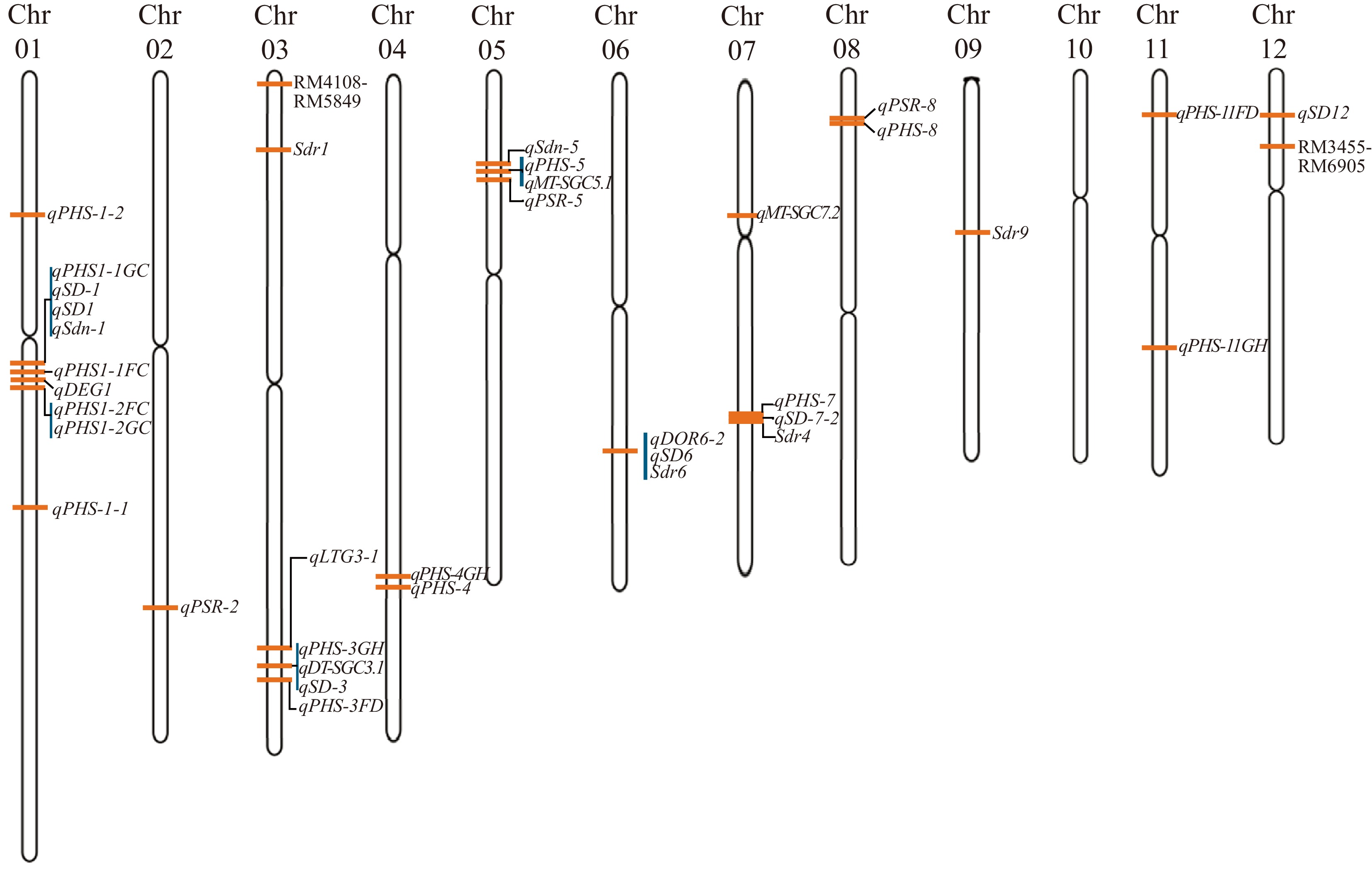
Figure 2.
Physical distribution of reported pre-harvest sprouting-associated quantitative trait loci on the rice genome. Rice chromosomes (Chr01–Chr12) are shown as vertical bars. Orange horizontal bars indicate the approximate physical positions of pre-harvest sprouting-related QTLs.
Table 2. Quantitative trait loci associated with pre-harvest sprouting in rice.
QTL Chr Mapping population Parents Ref. ♀ ♂ qPHS-1-1 1 RIL IR64 (Indica) Asominori (Japonica) [100] qPHS-1-2 1 RIL IR64 (Indica) Asominori (Japonica) [100] qPHS1-1FC 1 RIL Jinsang (Japonica) Gopum (Japonica) [101] qPHS1-2FC 1 RIL Jinsang (Japonica) Gopum (Japonica) [101] qPHS1-1GC 1 RIL Jinsang (Japonica) Gopum (Japonica) [101] qPHS1-2GC 1 RIL Jinsang (Japonica) Gopum (Japonica) [101] qSD-1 1 − 14 Japonica rice varieties 7 Indica rice varieties [102] qSD1 1 − 14 Japonica rice varieties 7 Indica rice varieties [102] qDEG1 1 − 14 Japonica rice varieties 7 Indica rice varieties [102] qSdn-1 1 − 14 Japonica rice varieties 7 Indica rice varieties [102] qPSR-2 2 F2 K81 G46B [103] qPHS-3GH 3 RIL Odae (Japonica) Unbong40 (Japonica) [104] qPHS-3FD 3 RIL Odae (Japonica) Unbong40 (Japonica) [104] Sdr1 3 − 14 Japonica rice varieties 7 Indica rice varieties [102] qDT-SGC3.1 3 − 14 Japonica rice varieties 7 Indica rice varieties [102] qSD-3 3 − 14 Japonica rice varieties 7 Indica rice varieties [102] RM4108 - RM5849 3 BIL Nipponbare (Japonica) Koshihikari (Japonica) [105] qLTG3-1 3 − 14 Japonica rice varieties 7 Indica rice varieties [102] qPHS-4GH 4 RIL Odae (Japonica) Unbong40 (Japonica) [104] qPHS-4 4 RIL IR64 (Indica) Asominori (Japonica) [100] qPHS-5 5 RIL IR64 (Indica) Asominori (Japonica) [100] qPSR-5 5 F2 K81 G46B [103] qSdn-5 5 − 14 Japonica rice varieties 7 Indica rice varieties [102] qMT-SGC5.1 5 − 14 Japonica rice varieties 7 Indica rice varieties [102] qDOR6-2 6 − 14 Japonica rice varieties 7 Indica rice varieties [102] qSD6 6 − 14 Japonica rice varieties 7 Indica rice varieties [102] Sdr6 6 − 14 Japonica rice varieties 7 Indica rice varieties [102] qPHS-7 7 RIL IR64 (Indica) Asominori (Japonica) [100] Sdr4 4 − 14 Japonica rice varieties 7 Indica rice varieties [102] qMT-SGC7.2 7 RIL Milyang 23 Tong 88-7 [102] qSD-7-2 7 − 14 Japonica rice varieties 7 Indica rice varieties [102] qPHS-8 8 RIL IR64 (Indica) Asominori (Japonica) [100] qPSR-8 8 F2 K81 G46B [103] Sdr9 9 − 14 Japonica rice varieties 7 Indica rice varieties [102] qPHS-11FD 11 RIL Odae (Japonica) Unbong40 (Japonica) [104] qPHS-11GH 11 RIL Odae (Japonica) Unbong40 (Japonica) [104] qSD12 12 − 14 Japonica rice varieties 7 Indica rice varieties [102] RM3455 - RM6905 12 BIL Nipponbare (Japonica) Koshihikari (Japonica) [105] Particularly, chromosome 1 contains several of the most consistently detected QTLs for PHS resistance. Early work identified qPHS-1-1 and qPHS-1-2 in an indica × japonica population. Later studies revealed additional loci such as qPHS1-1FC, qPHS1-2FC, qPHS1-1GC, and qPHS1-2GC in japonica backgrounds. Interestingly, DELAY OF GERMINATION1 (DOG1)-like regulation of dormancy in rice is mediated by loci on chromosome 1. OsDOG1L-3, which corresponds to the dormancy QTL qSD1-1 regulates seed dormancy through the ABA pathway[90]. Functional characterization indicates that OsDOG1L-3 influences dormancy depth without affecting pericarp pigmentation, which distinguishes its regulatory role from that of Rc[75]. Furthermore, it has been noted that the contribution of DOG1-like genes to seed dormancy is less prominent in rice than in Arabidopsis, which can be attributed to fundamental differences in dormancy regulatory architecture between the two species. In Arabidopsis, DOG1 functions as a central regulator of primary seed dormancy and exerts a strong genetic effect[89]. In contrast, seed dormancy in rice is governed by a polygenic regulatory system that integrates hormonal metabolism, transcriptional control, and grain structural characteristics[15,90]. Within this framework, DOG1-like homologs, including OsDOG1L, act as modulatory components rather than as primary determinants of dormancy[90]. These findings point to a major dormancy region where allelic differences influence ABA accumulation, ABA sensitivity, and the timing of after-ripening. Diversity analyses also uncovered qSD-1, qDEG1, and qSdn-1, which strengthen the evidence that chromosome 1 shapes fundamental dormancy contrasts between indica and japonica rice. The recurrence of these loci across unrelated studies indicates that chromosome 1 functions as a genomic center for maintaining dormancy depth[100−102]. Chromosomes 2 and 3 introduce additional layers of PHS resistance. For example, qPSR-2 on chromosome 2 affects how the embryo responds to moisture exposure near harvest and influences the stability of ABA-dependent germination control[103]. Chromosome 3 contains a broad cluster of loci that include qPHS-3GH, qPHS-3FD, Sdr1, qSD-3, and qLTG3-1. These loci influence the transition from primary to secondary dormancy and shape the strength of the ABA-GA balance during late seed development. Their repeated detection across distinct populations suggests that chromosome 3 coordinates environmental signals with internal hormonal adjustments that maintain embryo quiescence when wet conditions occur close to harvest[102,104]. QTLs on chromosomes 4 and 5 reinforce the idea that PHS resistance requires both biochemical and structural defenses. Particularly, qPHS-4, qPHS-4GH, and the well-characterized dormancy gene SDR4 help extend primary dormancy into the late maturation phase and prevent premature activation of α-amylase and other hydrolytic enzymes[100,102,104]. Chromosome 5 includes qPHS-5, qPSR-5, qSdn-5, and qMT-SGC5.1, each contributing to seed desiccation stability, endosperm integrity, and resistance to GA-driven germination[100,102,103]. These loci also influence hull tightness and glume adherence, which slow the entry of rainwater into the caryopsis and reduce the likelihood of vivipary under monsoon-like conditions. Chromosomes 6 and 7 add further regulatory depth to the genetic landscape of PHS resistance. Particularly, qSD6, qDOR6-2, and Sdr6 modulate the rate at which dormancy weakens and adjust the embryo's responsiveness to ABA during late seed maturation[102]. Chromosome 7 contains qPHS-7 and qSD-7-2, which contribute to moisture-related transitions in dormancy strength[100,102]. The presence of multiple PHS-associated loci near this region indicates that chromosome 7 acts as a regulatory zone where dormancy pathways and environmental sensing converge. Chromosomes 8, 9, 11, and 12 complete the distributed network of dormancy control. Chromosome 8 includes qPHS-8 and qPSR-8[100,103], and regions defined by RM4108–RM5849 and RM3455–RM6905 are shown on chromosomes 3 and 12, respectively[105]. These loci influence hull firmness, husk-grain adhesion, and moisture retention properties of the spikelet[100,103,105]. The Sdr9 locus on chromosome 9 affects dormancy depth and extends the protective window during which grains resist sprouting[102]. Chromosomes 11 and 12 contain qPHS-11FD, qPHS-11GH, and qSD12, all of which help suppress metabolic activation at the onset of wet conditions and delay the biochemical events that initiate germination[102,104]. Taken together, these QTLs reveal that PHS resistance in rice results from the combined action of diverse genomic mechanisms. Some loci enhance ABA biosynthesis or strengthen ABA signaling. Others reduce GA responsiveness, delay after-ripening, or stabilize protective seed structures such as hulls and glumes. Several loci also influence enzymatic thresholds that determine how quickly α-amylase and proteases activate once the grain encounters moisture. The geographic pattern of allelic variation mirrors the divergent domestication histories of indica and japonica rice: japonica cultivars retain stronger dormancy alleles, while many indica cultivars lost them during selection for rapid and uniform germination. This distribution highlights the need to reintroduce favorable dormancy alleles into PHS-susceptible germplasm as climate change intensifies the frequency of wet harvest conditions.
While rice provides the most detailed model, similar quantitative architectures underlie PHS resistance in other cereals. In wheat, major loci such as Phs1 on chromosome 4A and dormancy QTLs linked to TaVp1 and red pericarp genes shape sprouting tolerance[62,106]. In barley, strong dormancy loci on chromosomes 5H and 7H contribute to resistance, but interact with malting quality traits[107]. In maize, PHS resistance depends on loci associated with Vp1, ABA biosynthesis genes such as Vp14, and husk morphology, which jointly regulate kernel desiccation and exposure to moisture[61].
-
Susceptibility to PHS in cereal crops originates from a fundamental domestication trade-off between rapid and uniform germination after sowing and the maintenance of seed dormancy at maturity. Wild progenitors of cereals evolved strong dormancy as an adaptive trait under fluctuating rainfall and unstable soil moisture conditions, which delayed germination until conditions supported seedling establishment[108]. Agricultural domestication favored early and synchronous germination because these traits facilitated predictable crop establishment under managed systems. Selection for rapid emergence reduced dormancy strength and associated protective mechanisms across cereals, which increased vulnerability to sprouting before harvest.
Evidence from wheat, barley, and maize illustrates this general pattern. Wild and early domesticated wheats exhibit strong dormancy and red pericarp pigmentation, both associated with resistance to sprouting[18,109]. Selection for white-grained wheat improved processing quality but coincided with the loss of dormancy-linked alleles, which increased PHS susceptibility in cool and wet harvest environments[43,110]. In barley, breeding for malting quality emphasized rapid and uniform germination, which reduced dormancy and elevated PHS risk under humid conditions[54,111]. In maize, physiological dormancy is inherently weak, and resistance to PHS relies mainly on husk-mediated physical protection. Domestication favored looser husk architecture to improve drying and harvest efficiency, which increased kernel exposure to moisture and susceptibility to vivipary under wet harvest conditions[14,108,112].
In contrast to these brief comparative examples, rice provides a particularly clear illustration of domestication-driven trade-offs between dormancy and early germination. In wild rice species, the development of strong seed dormancy and delayed germination was not accidental, but a product of adaptation under natural selection. These traits acted as safeguards in monsoon ecosystems, where rainfall was plentiful, yet irregular and often short-lived. Dormancy kept the seeds from reacting to short rain showers that were not enough to sustain young seedlings[113]. Instead, seeds remained quiescent until soil moisture was sufficient to sustain growth. This protective delay reduced the risk of crop failure under unstable weather patterns. Another layer of protection came from staggered germination. Not all seeds germinated at the same time, which spread the risk across several growth cycles. Even if one cohort failed due to drought, flooding, or disease, other seeds remained in the soil seed bank and preserved the population. This staggered strategy was vital in flood-prone lowlands and unpredictable upland fields, where conditions could shift rapidly[26]. These mechanisms together gave wild rice resilience against vivipary, the premature germination of seeds on the mother plant. In humid environments, especially in regions subject to prolonged monsoon rains, the danger of vivipary was ever present. Strong dormancy and slow germination served as natural defenses. They allowed seeds to survive wet conditions without sprouting before the right time. By holding back germination until the right season arrived, wild rice maintained its ecological foothold across diverse and challenging habitats[6]. Domestication altered these natural safeguards in profound ways. As rice transitioned from wild stands to cultivated fields, farmers favored traits that suited agricultural management rather than ecological resilience. Strong dormancy, which once helped rice survive irregular rainfall, later became an obstacle because it slowed and disrupted uniform crop growth. Early farmers selected seeds that germinated quickly and consistently after sowing, especially in irrigated paddies where water availability could be controlled. Over many generations, this selection pressure reduced dormancy, and the resulting decline in dormancy strength weakened the natural barrier against pre-harvest sprouting[3]. Wild grains were tightly enclosed by firm hulls that slowed water entry and reinforced dormancy (Fig. 3). Cultivated rice was bred for easier threshing and milling, which required hulls that were less rigid and more loosely attached. This change made processing easier, but also exposed the kernels more to rain and moisture. As a result, they became more likely to sprout while still on the panicle[114]. The legacy of domestication is visible in the contrasting ecotypes of rice today. Indica rice varieties, widely grown in tropical Asia, often show very weak dormancy and are highly prone to vivipary under monsoon harvest conditions. Japonica rice varieties, domesticated in temperate East Asia, generally retain stronger dormancy and show better resistance to sprouting[115,116]. This difference came from both nature and human choice. Indica varieties fit well in irrigated, long-season fields where farmers prefer quick germination. Japonica varieties developed in cooler, shorter growing seasons where dormancy still gave an advantage.
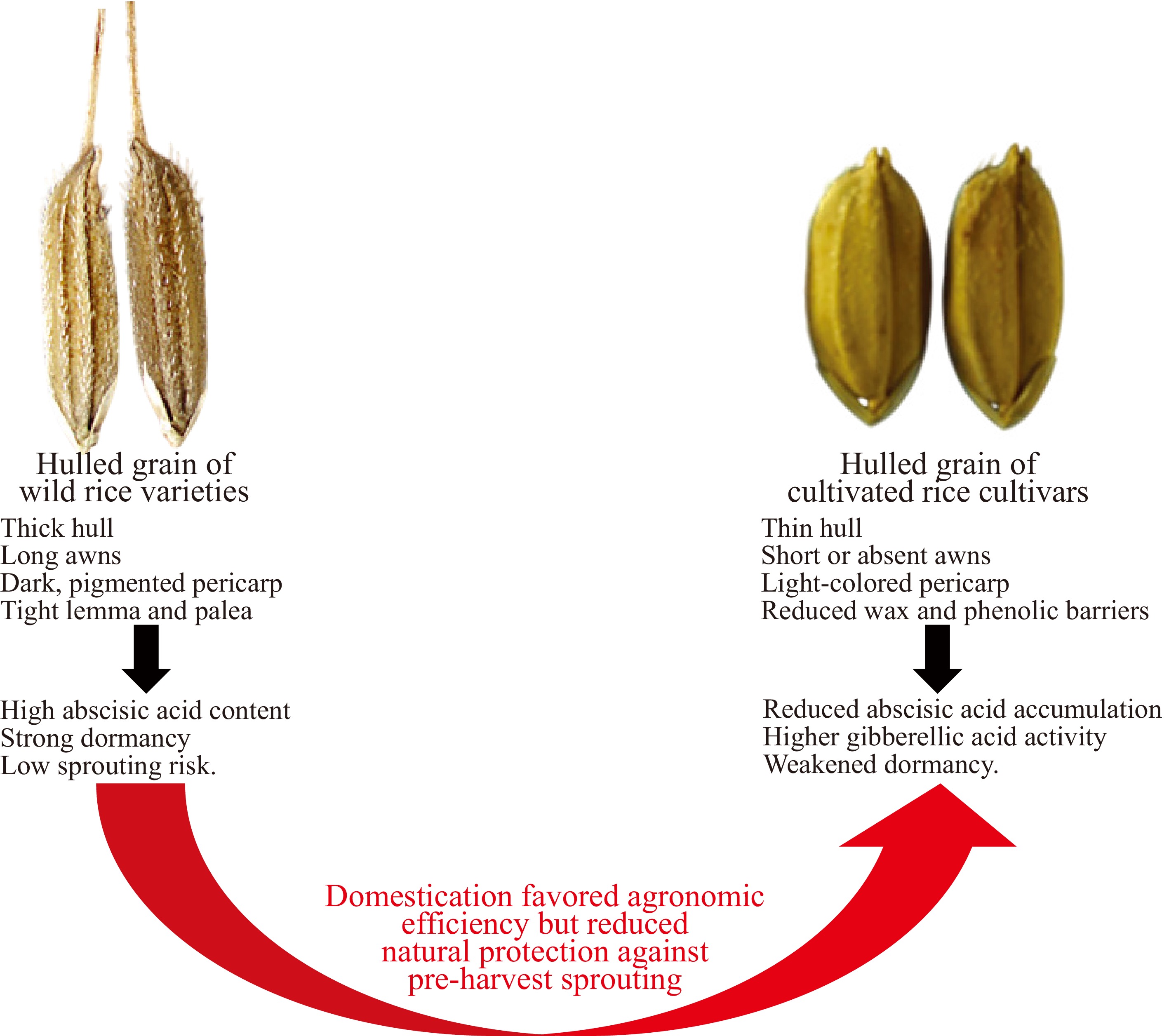
Figure 3.
Evolutionary and domestication-related changes affecting pre-harvest sprouting in rice. Wild rice species possess firm hulls, pigmented pericarps, and high abscisic acid levels that maintain strong seed dormancy and resist moisture penetration. Domestication selected for traits that favor rapid germination and easy threshing, including thinner hulls and reduced dormancy, which increased pre-harvest sprouting susceptibility in cultivated rice varieties. These evolutionary trade-offs highlight the need to reintroduce dormancy alleles and structural defenses into modern breeding programs.
The story of rice and PHS demonstrates how natural and artificial selection has pulled in opposite directions. Wild rice relied on strong dormancy and protective hulls to survive erratic rainfall, while domestication gradually dismantled these defenses in favor of traits that suited agriculture[7]. Indica varieties, shaped by cultivation in tropical regions with controlled irrigation, embody this trade-off most clearly: they germinate quickly and uniformly, but are highly vulnerable to sprouting when harvest coincides with monsoon rains. Japonica varieties, by contrast, retained part of the natural dormancy and structural protection, which allows them to resist sprouting more effectively in temperate climates. These contrasting outcomes underline the challenge faced by modern breeders. On one hand, agriculture demands rapid and uniform germination; on the other, food security requires resilience against unpredictable rainfall. Current research seeks to reintroduce dormancy alleles from japonica or wild rice into indica backgrounds, aiming to rebuild natural safeguards without undermining the agricultural efficiency that domestication has achieved. The domestication history of rice illustrates the contrasting pressures imposed by natural and artificial selection. Wild rice depended on strong seed dormancy and structural protection to persist under unpredictable rainfall, while domestication favored traits that enhanced agricultural efficiency but reduced resilience to moisture exposure at harvest. This trade-off accounts for current patterns of PHS susceptibility in rice and highlights a key challenge for modern breeding. Current efforts aim to reintroduce favorable dormancy alleles from wild rice or japonica backgrounds into indica germplasm to improve resistance to PHS while retaining desirable agronomic traits.
-
Future research on PHS should continue to advance from crop-specific descriptions toward a more integrated understanding of seed-level regulatory mechanisms that operate across cereal species. Although rice, wheat, barley, and maize differ in their domestication histories, inflorescence structures, and ecological adaptations, comparative evidence indicates that PHS susceptibility emerges from common biological processes that govern seed dormancy maintenance, hormonal balance, and responses to moisture exposure at maturity. Expanding research frameworks that emphasize these shared mechanisms will strengthen the conceptual foundation of PHS biology.
A central priority lies in refining the understanding of dormancy regulation as a dynamic trait that responds to both developmental cues and environmental signals. While the roles of ABA and GA in controlling dormancy and germination are well established[48,49,70,117], further work is needed to clarify how hormonal sensitivity, biosynthesis, and turnover are coordinated during late seed maturation across cereals. Comparative analyses of ABA-related pathways, including NCED-mediated biosynthesis and downstream transcriptional regulation, may reveal conserved modules that stabilize dormancy under transient wet conditions[56]. Meanwhile, species-specific differences in hormone responsiveness should be examined to explain contrasting PHS phenotypes among crops.
Genetic studies will remain essential for dissecting the polygenic nature of PHS resistance. Future efforts should emphasize the integration of QTLs, genome-wide association analyses, and functional genomics to define how multiple loci interact to shape dormancy depth, after-ripening behavior, and germination thresholds. Cross-cereal comparisons of key dormancy regulators will help distinguish universal regulatory principles from lineage-specific adaptations.
Structural traits of seeds and reproductive organs represent another important research frontier. Hull adherence in rice, pericarp properties in wheat, husk coverage in maize, and spike architecture in barley all influence the extent and duration of water exposure at maturity[1,110]. Greater attention to the developmental and genetic control of these traits will improve understanding of how physical barriers interact with physiological dormancy to suppress sprouting. Integrating structural phenotyping with physiological and molecular analyses will provide a more complete picture of PHS resistance at the whole-seed level.
Finally, future studies should strengthen the linkage between seed biology and environmental variability. The timing, duration, and intensity of rainfall, humidity, and temperature fluctuations near harvest interact with intrinsic dormancy states to determine sprouting outcomes. Coordinated experiments that combine controlled environmental treatments with field observations across cereal species will improve the ability to interpret PHS as a context-dependent trait. Such integrative approaches will support a biologically grounded understanding of PHS and contribute to more resilient cereal production systems under increasingly unpredictable climatic conditions.
-
PHS represents a major constraint on cereal grain quality and yield stability when harvest coincides with humid or rainy conditions. Evidence synthesized in this review shows that resistance to PHS depends on the maintenance of seed dormancy at maturity and on the coordinated action of hormonal regulation, genetic control, and grain structural traits. Across cereals, ABA biosynthesis and signaling form a central regulatory system that suppresses premature germination, whereas GA pathways promote dormancy release when environmental conditions become favorable. Genetic analyses also indicate that PHS resistance has a polygenic basis in which numerous loci contribute partial effects to dormancy strength, hormone sensitivity, and grain permeability to water. Domestication and breeding have reshaped this system because selection favored rapid and uniform germination. This shift weakened natural dormancy barriers and increased susceptibility to sprouting in many modern cereal cultivars. Comparative evidence from different cereal crops, with rice as a key reference, also shows that distinct domestication histories and environmental adaptation have influenced present-day variation in PHS susceptibility. By integrating physiological, genetic, and domestication-related evidence, this review provides a consolidated framework for understanding the biological basis of PHS in cereals and for interpreting variation in sprouting resistance across production environments.
-
The authors confirm their contributions to the paper as follows: conceived the manuscript: Chu HD, Duong CN, Tran LSP; drafted the manuscript preparation: Nguyen TQ, Chu HD; literature review and data collection: Dong GH, Nguyen LV, Chuong NN, Le TD, Li W. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and analyzed in the current study are available from the corresponding author upon reasonable request.
-
This research has been done under the research project QG.25.61 of Vietnam National University, Hanoi.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Nguyen TQ, Dong GH, Nguyen LV, Le TD, Chuong NN, et al. 2026. Integrated physiological, genetic, and environmental insights into pre-harvest sprouting in cereal for climate-resilient breeding. Seed Biology 5: e017 doi: 10.48130/seedbio-0026-0014 

Integrated physiological, genetic, and environmental insights into pre-harvest sprouting in cereal for climate-resilient breeding
- Received: 05 December 2025
- Revised: 12 March 2026
- Accepted: 16 March 2026
- Published online: 02 June 2026
Abstract: Pre-harvest sprouting (PHS) poses a major threat to cereal production during grain maturation under humid and unstable climatic conditions. PHS reduces yield and grain quality and disrupts production stability in many growing regions. This review integrates physiological, hormonal, genetic, and environmental evidence to clarify the regulatory basis of PHS resistance in cereal. Seed dormancy represents the primary barrier that prevents premature germination, and its maintenance depends on abscisic acid and gibberellin balance, dormancy-related transcriptional regulators, and grain structural traits that restrict water entry. To our interest, genetic analyses reveal that PHS resistance in rice (Oryza sativa) follows a polygenic architecture, with numerous quantitative trait loci distributed across most chromosomes that contribute partial effects to dormancy strength and vivipary suppression. Recent molecular and multi-omics studies have identified key regulatory genes and pathways that stabilize dormancy at maturity and modulate germination thresholds under moisture exposure. Domestication and breeding have weakened these protective mechanisms in many modern cultivars, particularly in indica rice, which explains current vulnerability under monsoon harvest conditions. By synthesizing advances from rice and comparative evidence from other cereals, this review outlines genetic and physiological principles that support the development of climate-resilient cereal cultivars with improved resistance to PHS.