








-
The detrimental effects of non-native pathogenic fungi causing disease to agricultural crops or native flora and fauna are well documented[1,2]. Additionally, hundreds of non-native mutualistic fungi have facilitated the establishment and spread of invasive plants in more than 40 countries, and on all continents except Antarctica[3], which can have cascading effects on soil nutrient stocks that facilitate future invasions[4].
Saprotrophic fungi growing outside of their native ranges have been reported for > 150 years[5] but their effects on species and habitats where they have established are poorly understood[6]. Saprotrophic species with the best documented establishments outside of their native ranges are conspicuous. The bright red Clathrus archeri, Devil's Fingers Fungus or Octopus Stinkhorn, spread from its native range in Australia and New Zealand to Europe, North America, Africa, and South America[7] (Fig. 1a). The bright orange Favolaschia claudopus, a member of the F. calocera species complex known as Orange Ping-Pong Bats or Orange Pore Conk, arrived in New Zealand before 1970 and has spread to Australia, western Europe, and Hawai'i[8−10] (Fig. 1b). Both species are saprotrophs on wood, and are thought to have been introduced through wood products, horticultural practices, or in the case of C. archeri in Europe, via wool or soldier deployments[7]. Although F. claudopus and C. archeri have been labeled invasive species, which we define as non-native species likely to cause environmental harm[11], evidence of invasive saprotrophic species is limited to cultivated mushrooms.

Figure 1.
Examples of invasive saprotrophic fungi. (a) Clathrus archeri (Devil's fingers or Octopus Stinkhorn) is native to Australia and New Zealand and has spread to Africa, Europe, and North and South America. Photograph by: Oilys, CC-BY-SA-3.0-migrated (
https://creativecommons.org/licenses/by-sa/3.0 ), plate letter added. (b) Favolaschia claudopus (Orange Ping-Pong Bats or Orange Pore Conk) has spread to Australia, Hawai'i, New Zealand, and western Europe. Its native range is not known. Photograph by: Alan Rockefeller, CC-BY-4.0 (https://creativecommons.org/licenses/by/4.0/deed.en ), plate letter added. (c) Pleurotus citrinopileatus (Golden Oyster Mushroom) is native to east Asia, has established populations outside its native range in Africa, Asia, and Europe, and is a commonly cultivated species that is spreading rapidly throughout eastern North America. Photograph by Kerry Givens, CC-BY-SA-3.0 (https://creativecommons.org/licenses/by-sa/3.0/deed.en ), plate letter added.Cultivated edible and medicinal fungi are a global industry worth up to
${\$} $ -
Actions to stop cultivated fungal invasions will vary depending on the stage of the invasion (Fig. 2). The least ecologically harmful, and often most economically efficient, approach to managing invasive species is to stop an invasion before it happens[17]. To prevent the establishment of non-native fungi and other potential pests, concerned countries or localities have laws limiting the import of certain organisms and plant-growing media (e.g., soil or other material for plant roots)[18]. While fungi can be inconspicuously transported outside their native range on a variety of substrates, cultivated fungi sold commercially are a much less cryptic group as they are intentionally bought, sold, and transported globally.
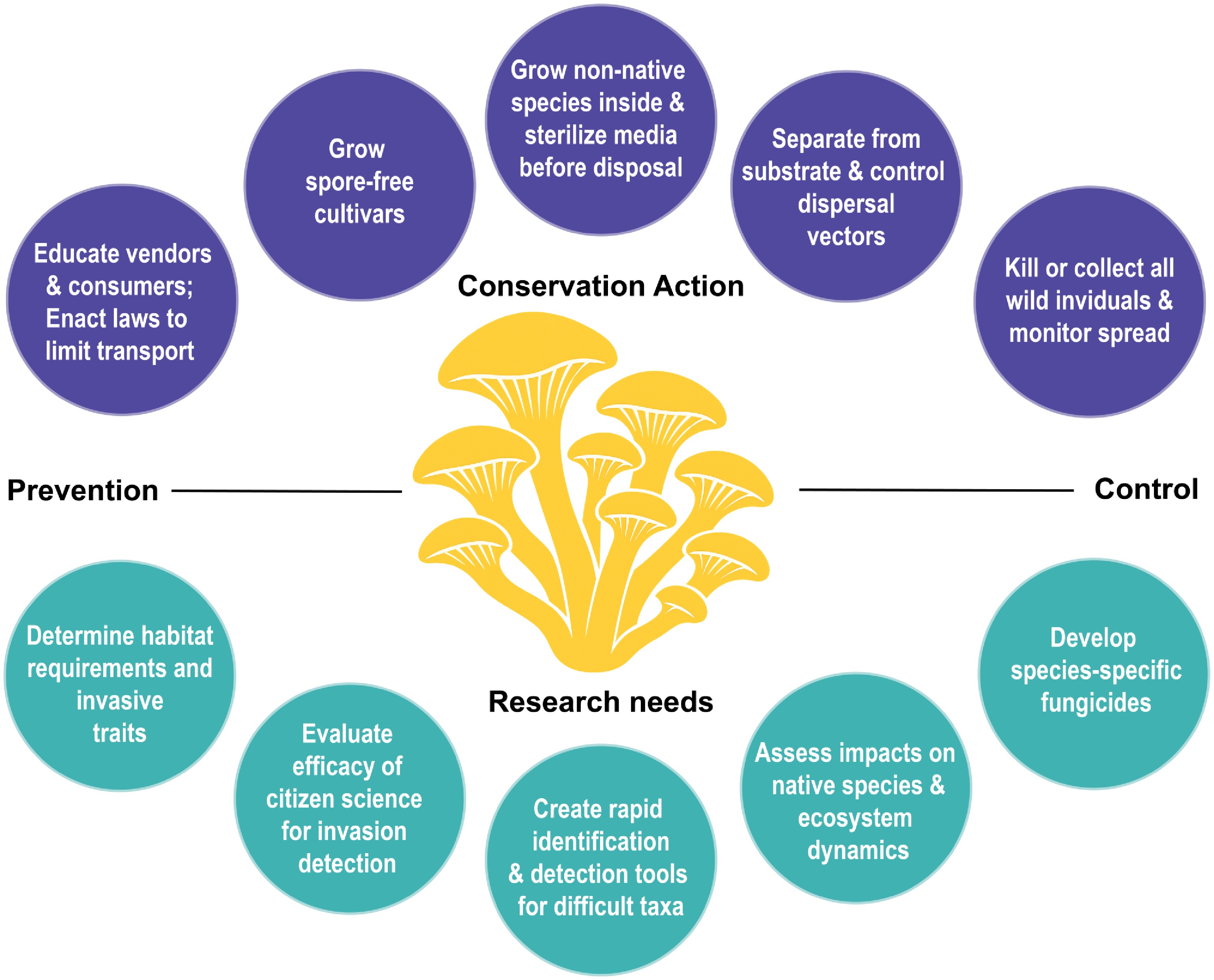
Figure 2.
Conservation actions and research needs to prevent the establishment and control the spread of invasive cultivated fungi.
To prevent establishment in the short-term, we suggest non-native cultivated mushrooms not be grown outdoors, particularly species that do not have a long history of cultivation in a given region. Commercial grow-at-home kits should be disposed of after killing the mycelia with heat > 150 °C for > 1 h in the oven, or via other methods such as pasteurization or pressure cooking. Vendors should include the above suggestions in instructions accompanying their products to help prevent outdoor establishment, and they should voluntarily stop the sale of known invasives, like P. citrinopileatus, to regions where it is not established. Finally, a campaign to educate consumers to not purchase, and pressure vendors to not sell P. citrinopileatus, may prevent its establishment or slow its spread to regions where it is not established, although we note these tools have had only marginal success in the horticultural plant trade[19].
In the long-term, regulation of the transport of fungi and their spores for cultivation should be considered at multiple levels of government to prevent future invasions. For example, the State of Hawai'i, USA, has a list of restricted microorganisms, including all Pleurotus species not previously reported from the state, requiring a permit before legal import (Haw. Code R. § 4-71A-22). Australia maintains a list of mushrooms for propagation with specific import rules for each species or group of species[20]. Additionally, multiple states in the USA have restricted the sales of spores to grow fungi that produce psilocybin or psilocin (e.g., O.C.G.A.§ 16-13-71), which otherwise could be legally sold in the United States because the spores do not contain these substances themselves.
Once established, the elimination or control of invasive species, particularly cryptic taxa with microscopic bodies and propagules, is unlikely to be successful, but higher success may occur in the initial phase when the population is smaller and geographically limited. During an incipient invasion, a fungus may be controlled or potentially eliminated by physically separating it from its required host or substrate, although this is labor intensive[21]. Other methods of control include killing or repelling vectors that disperse the fungus[22] or other organisms that make substrates susceptible to colonization[23]. Finally, manually removing fungi with macroscopic sexual structures before sporulation[21] or using a biocide to kill foliar fungi[24] may also be attempted, although successfully eliminating a pathogen via any of these methodologies has not been documented. The best option will depend on several factors, including the biology of the fungus, host or substrate, environment, and resources available.
For a generalist saprotroph like P. citrinopileatus that immerses itself in its substrate and does not rely on a vector for dispersal, options for control are limited. Removing substrates or creating a barrier between the fungus and its substrate could stop vegetative spread, but is likely not an option. When mushrooms are encountered, they can be collected into an impermeable container, ideally before they mature, to stop spread via spores.
After the incipient phase, when an invasive fungus has achieved a large, widespread population, elimination is unlikely, and control is more challenging. Individually treating susceptible hosts or substrates to prevent or reduce colonization[25], introducing genetic resistance into hosts[26], and using other organisms as a biological control[27] are all potential strategies in addition to those used during incipient invasions. Despite all these strategies, and the ability to limit invasion to individual organisms or small areas of habitat, we are not aware of successful campaigns to control non-agricultural, landscape-scale fungal invasions.
-
While we advocate for preventive practices and science-based control methods, several research gaps remain to better understand the effects of invasions of saprotrophic, and specifically cultivated, fungi. First, repeating the Golden Oyster Mushroom study[15] in different localities, in different species of dead logs, and at different times of the year will help clarify whether the presence of P. citrinopileatus always suppresses native fungal species richness, or if impacts vary. More broadly, the metabarcoding methodology should be used in other localities with non-native saprotrophic fungi to determine if co-occurring native fungi in the same substrate have reduced richness.
Beyond understanding effects and impacts, we need research on how to best predict invasions[28]. Habitat modeling has been used to predict areas of potential expansion of putatively invasive saprotrophic fungi[7], but we are not aware of any modeling for cultivated fungi besides the analysis of Golden Oyster Mushrooms[15] showing that the climate in western North America, where P. citrinopileatus is not widely established, has been suitable for this fungus since 2021. Similarly, it is hypothesized that certain traits, such as fast growth, increased temperature tolerance, mold resistance, and abundant mushroom production, increase the invasion success of Golden Oyster Mushrooms[15]. Asexual reproduction is one trait hypothesized to facilitate invasions in both saprotrophic and mycorrhizal fungi[9,29], while this trait was not shown to have a large effect in a meta-analysis of pathogenic fungi[30].
Habitat modeling and trait examination, which are currently lacking for all cultivated fungi except P. citrinopileatus, would be particularly useful for newer products, or those with limited market share. For example, Chicken of the Woods (Laetiporus spp.) is sold for cultivation, often suggested to be grown outdoors, and several species within North America have non-overlapping distributions[31]. It is unknown if these disjunct distributions are maintained by life history traits, such as preferred climate and host, or natural dispersal barriers. Therefore, it is prudent to generate these and additional data to evaluate establishment and displacement risks.
Breeding cultivars for reduced invasibility is an essential and actionable prevention measure. Sporeless and low-spore varieties of oyster mushrooms have been developed to reduce allergies in mushroom growers[32]. Recently, a key transcription factor essential to spore maturation was identified that is highly conserved in mushroom-forming fungi and has the potential to facilitate the development of sporeless cultivars in a wider range of species[33]. Notably, the identified gene is not involved in mushroom development beyond spore maturation, making it an excellent broad-range target for sporelessness.
While prevention of invasions is preferable, fungi are likely to continue to establish populations outside of their native ranges, and our current methods to detect, track, and control these species are limited. Understanding how effective citizen scientists are in detecting incipient invasions and tracking their spread, and learning how to better incorporate amateur communities into this work, is one promising avenue of research. In the case of P. citrinopileatus, citizen science platforms, including iNaturalist, provided the best retrospective data to date, the first report in the wild, and follow its range expansion over time[15,16]. While citizen science groups have helped detect and track forest pests and pathogens in different situations[34], the only program we are aware of that broadly focuses on non-native fungi began recently in Europe[35].
Detecting and tracking invasions of taxa with native lookalike species is a major challenge. Species differentiation can often be accomplished by DNA barcoding, which is routinely used to identify mushrooms and fungi in environmental substrates[36]. Detecting intraspecific genetic variation in situations where non-native genotype monitoring is needed may require more sensitive genetic markers or tools, like qPCR assays[37]. For taxa where specific markers are not developed, rapid whole-genome sequencing can be deployed, even in the field, for these purposes[38]. The genus Psilocybe is an example where sequence-based detection and monitoring could be applied. There are around 150 species of Psilocybe globally. Differentiating among species morphologically often requires examining microscopic structures[39], and multiple, cultivated strains exist of individual species. Many species produce the hallucinogenic compounds psilocybin, psilocin, and baeocystin, and they are widely cultivated, traded, or intentionally introduced outside their native ranges[40]. Psilocybe species escaping from cultivation have been documented[41], but whether escaped cultivars are restricted to human-altered landscapes remains to be investigated. Sequence-based identification for Psilocybe, and groups with similar challenges, like Favolaschia, offers a reliable detection and monitoring solution.
For species that are identified as major ecological, economic, or health threats, development of species-specific fungicides is warranted, as has been attempted with a few plant pathogens[42]. Pairing fungicide development with identifying invasion mechanisms, specifically determining which attributes lead to exclusion of local taxa, may result in more effective control methods. For instance, is the ability of C. citrinopileatus to exclude native species from substrates due to its rapid growth rate, specific secondary metabolites, other traits, or some combination? Once mechanisms leading to this exclusion are identified, compounds can possibly be designed to specifically target those mechanisms[43]. As with newer, targeted biocides, these techniques are at the edge of scientific discovery, and should be tested for unintentional adverse effects before implementation.
-
Diverse fungal communities are essential for functioning ecosystems and human health[44], and cultivated fungi provide food, medicine, and livelihoods to countless people. Fungal conservation, awareness, and appreciation have improved markedly in the past two decades[44], and new consequences of human-mediated fungal interactions continue to be revealed. While some non-native saprotrophic fungi have been labeled invasive, we now know they can have negative effects on native fungal communities for the first time[15]. Knowledge of this emerging threat to wood-decay fungi, an ecological group with an already diminishing habitat[45], leads to more research needs. However, future research needs do not preclude current conservation actions. We applaud several vendors of P. citrinopileatus in North America and Europe for deciding to phase out sales of this species or ban it from community events until sporeless cultivars are developed[46]. In this study, we give several actions that mushroom growers, foragers, citizen scientists, and research mycologists can take to help address this challenge (Fig. 2). Limiting the spread and preventing the establishment in new locations of P. citrinopileatus and other potentially invasive, cultivated fungi should be a priority to address this emerging threat.
-
Not applicable.
-
Both authors researched and prepared the manuscript, and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study
-
Jessica L. Allen was supported by the BAND Foundation. The authors thank three anonymous reviewers for their suggestions to improve this study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Stallman JK, Allen JL. 2026. What should we do about cultivated mushroom invasions? Studies in Fungi 11: e010 doi: 10.48130/sif-0026-0008 

What should we do about cultivated mushroom invasions?
- Received: 25 November 2025
- Revised: 14 January 2026
- Accepted: 27 January 2026
- Published online: 15 April 2026
Abstract: Fungal invasions can have detrimental impacts on native species. While most invasive fungi are the result of unintentional introductions, cultivated edible and medicinal species used for commercial and at-home production are transported globally, and some have naturalized outside of their native ranges. For instance, the Golden Oyster Mushroom has invaded eastern North America, where it is displacing native species, and has also established outside its native range in Africa, Asia, and Europe. Here, we argue that cultivated fungi are a unique source of invasives that warrant additional research, action, and policy changes. In particular we suggest research to predict invasion potential of cultivated mushrooms and develop sporeless cultivars. We suggest awareness campaigns to producers and consumers focused on reduced transport of known invasives and proper disposal of fungal tissue. Policy changes to limit trade of invasive fungi to certain regions are a longer-term solution to address this emerging threat.
-
Key words:
- Citizen science /
- Invasion biology /
- Invasive species management /
- Policy /
- Trade