








-
The Myxomycetes, also known as slime molds or myxogastrids, represent a diverse group of organisms within the kingdom Protista, exhibiting a distinctive life cycle that alternates between a unicellular amoeboid stage and a multicellular plasmodial stage. During the plasmodial phase, these organisms form large, multinucleate masses capable of movement and phagocytosis. Myxomycetes have been recorded from tropical to temperate regions, found in a variety of habitats, including forests, grasslands, wetlands, and even in agricultural and urban landscapes, where they play critical roles in nutrient cycling and decomposition. They are also valuable model organisms for studying cell biology and development[1−4].
Historically classified as fungi due to the presence of fruiting bodies that produce spores, the Myxomycetes are now recognized as Protozoans. This classification is based on their resemblance to uninucleate, haploid amoeboid cells, which lack a rigid cell wall. Despite being excluded from the fungal kingdom, Myxomycetes remain a subject of mycological study, largely for taxonomic classification, with recent research extending toward ecological investigations[5]. These organisms play a critical role in the decomposition of plant litter and woody substrates, thereby contributing to nutrient cycling and soil conditioning, functions comparable to those of saprotrophic fungi. Owing to their ephemeral and cryptic nature, they appear only transiently within forest ecosystems, yet their contributions to ecological processes are substantial.
Estimating the global diversity of the Myxomycetes remains challenging due to their extensive variability and limited study[6]. The 'Dictionary of the Fungi' by Paul Kirk reports approximately 900 recognized species[7]; however, molecular evidence has suggested that this figure is underestimated, as many ecosystems, particularly tropical forests in Southeast Asia, remain under-surveyed[8].
In India, Myxomycetes have been unevenly documented, with 373 species recorded across six orders and 50 genera. West Bengal alone accounts for 85 species distributed among seven orders, nine families, and 28 genera[9]. This documentation is from largely isolated records from regions with limited forest cover[10]. A more consolidated understanding of national and regional myxomycete diversity has been facilitated by the Indian Myxomycetes Database (IMD), the first centralized repository of verified records, which has substantially strengthened baseline knowledge of myxomycete diversity in India[11].
The Darjeeling Himalayas, in particular, host a rich assemblage of species, including several new records[9]. Neora Valley National Park (NVNP), located in West Bengal, India, represents an important landscape for biodiversity research. Covering approximately 160 km2, NVNP is part of the Eastern Himalayan landscape, a Global Biodiversity Hotspot[12] and a potential UNESCO World Heritage Site candidate[13]. Its altitudinal gradient, ranging from 180 to 3,200 m, supports diverse forest types, including Subtropical Mixed Broadleaf, Lower and Upper Temperate Evergreen, and Rhododendron Forests. The park's spatial contiguity with adjacent protected areas like Pangolakha Wildlife Sanctuary and the Toorsa Strict Reserve further emphasizes its ecological significance. In particular, the temperate forests of NVNP, characterized by high moisture availability and rich biodiversity, provide an ideal setting for investigating the Myxomycetes diversity and ecology[14−17].
A systematic survey of NVNP was conducted between 2019 and 2021 through Annual Biodiversity Assessment Camps (ABACs), generating new insights into the diversity of Myxomycetes present in the region. The survey involved more than 20 experts and aimed to document species diversity, morphological characteristics, and spatial distribution of biodiversity components within NVNP, including the Myxomycetes.
The distribution of Myxomycetes is influenced by various environmental factors, including topographic aspects, soil properties, and vegetation health. Aspect, or the orientation of slopes, affects mesoclimatic conditions such as temperature and moisture, which are crucial for the growth and development of Myxomycetes[18−20]. North-facing slopes, characterized by cooler temperatures and higher moisture availability, often provide ideal conditions for Myxomycetes[1]. Soil properties, particularly bulk density and clay content, further shape habitat suitability by influencing soil porosity, moisture retention, and nutrient availability[21−23]. Soil pH also plays an important role by regulating nutrient availability, thereby affecting Myxomycetes distribution[24].
Vegetation condition indirectly governs Myxomycetes habitats through its influence on organic matter availability and substrate continuity. The Normalized Difference Vegetation Index (NDVI) has been widely applied to assess vegetation health and density, and is an important variable affecting habitats of the Myxomycetes. NDVI values correlate with organic matter availability and vegetation density, which supports Myxomycetes occurrence through vegetation cover and greater availability of decomposing plant substrate[25].
This study integrates field surveys with MaxEnt-based Ensemble of Small Models to document Myxomycete diversity, distribution, and environmental correlates in Neora Valley National Park, providing novel insights into this ecologically significant but understudied group in the Eastern Himalayan biodiversity hotspot.
-
The study was conducted in Neora Valley National Park (NVNP), located in the Kalimpong District of West Bengal, India, within the eastern Himalayan biodiversity hotspot extending from 26.9805°−27.1455° N, 88.6669°−88.8257° E. The park spans approximately 26°52′−27°07′ N and 88°45′–88°55′ E, covering an altitudinal range of 183–3,200 m a.s.l. and representing a gradient from subtropical to temperate evergreen forests.
The study was conducted as part of the third, fourth, and fifth Annual Biodiversity Assessment Camps (ABACs) organized by the Department of Forests, Government of West Bengal, between 2019 and 2021. These consecutive surveys were undertaken at three distinct localities within Neora Valley National Park (NVNP), representing different forest compartments and base camps selected based on accessibility, vegetation heterogeneity, and habitat representativeness (Fig. 1). The three primary forest camps were Bhoteykharka, Thamkharka, and Public Health Engineering (PHE). The third ABAC, held from 20 to 24 October 2019, was based at Bhoteykharka camp (27.0620° N, 88.7748° E; 1,950 m a.s.l.) and covered transects within the Eastnar 21 Forest Compartment of the Lower Neora Range. The fourth ABAC, conducted from 15 to 19 March 2020, was centered at Thamkharka camp (27.0978° N, 88.6906° E; 1,952 m a.s.l.), encompassing transects across Rashet 1–2 and Kolbong 3 of the Upper Neora Range, and Ambiok 1–3 of the Lower Neora Range. The fifth ABAC, organized between 11 and 16 March 2021, was held at the PHE Camp (27.09736° N, 88.72502° E; 2,158 m a.s.l.) and included transects along the PHE (8 No. Veer), Alubari Camp Route, Bhotey Kharka Route (Animal Route), and Zero Route (Fig. 1). Each site comprised multiple accessible forest trails and natural clearings used to establish transect routes for systematic sampling.
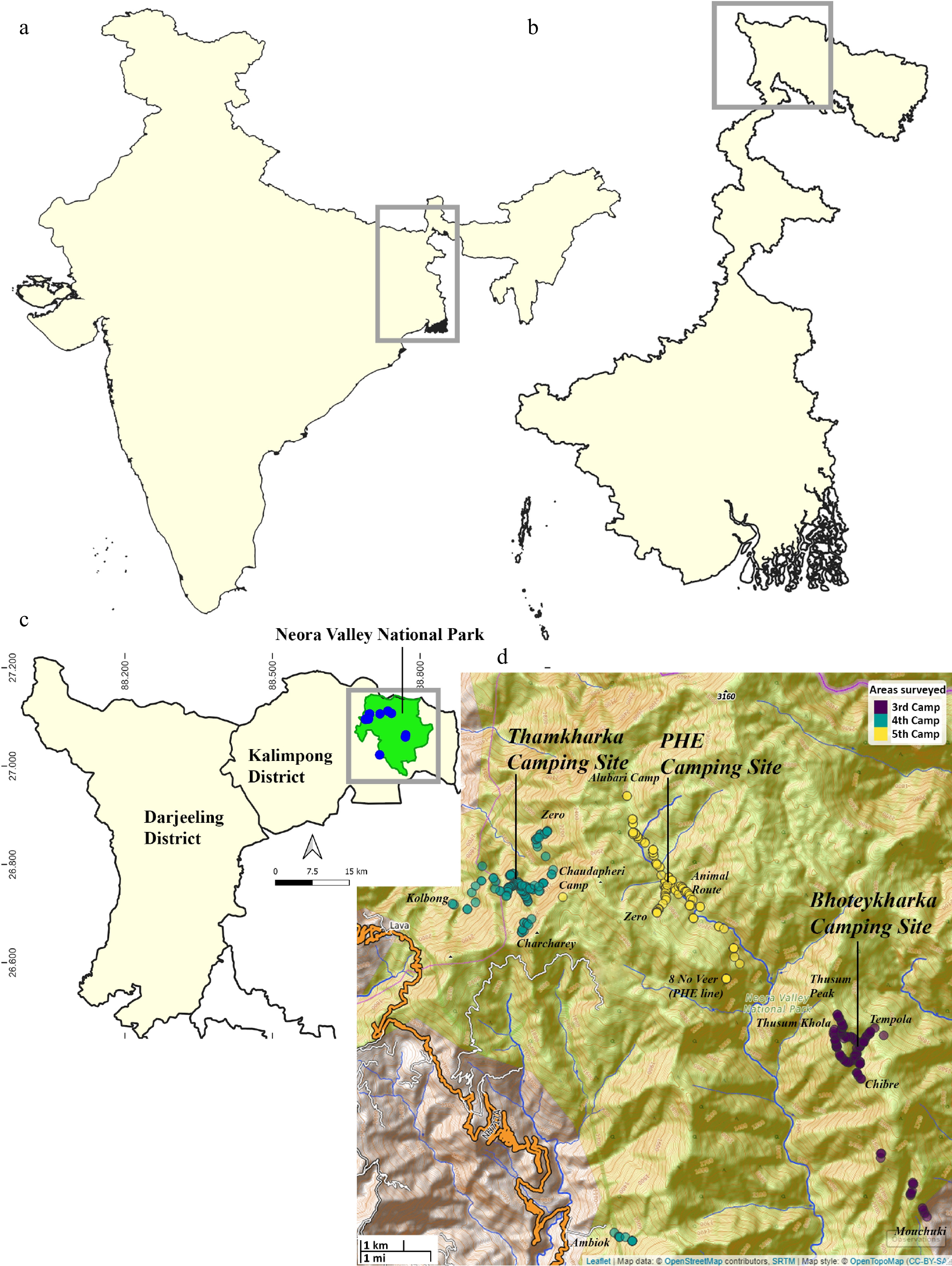
Figure 1.
(a) Map of India showing location of the state of West Bengal (in grey box). (b) Map of West Bengal showing location of Darjeeling and Kalimpong districts (in grey box). (c) Map of Kalimpong district with location of Neora Valley National Park (in green polygon), survey locations (blue dots) in Kalimpong District and area focused for figure d (in grey box). (d) Survey locations covered during 3rd (Purple dots), 4th (Aquamarine dots), and 5th (Yellow dots) ABACs. Maps (a−c) were prepared using QGIS (v. 3.26) and (d) using the mapview package in R (v. 4.2.1).
The spatial context of the study area is illustrated in Fig. 1. At the regional scale (Fig. 1a), the map shows the location of the state of West Bengal within India. At the state level (Fig. 1b), the Darjeeling and Kalimpong districts are highlighted within West Bengal, indicating the broader geographical setting of the study. The Kalimpong District map (Fig. 1c) delineates the Neora Valley National Park (NVNP) as a green polygon, along with survey locations (blue dots) distributed across the NVNP and the specific area focused on in Fig. 1d. At the most detailed scale (Fig. 1d), the map presents the survey locations corresponding to the third (purple dots), fourth (aquamarine dots), and fifth (yellow dots) ABACs, marking the positions of the Bhoteykharka, Thamkharka, and PHE camps, respectively. The NVNP boundary shapefile was created by georeferencing the official park boundary in Google Earth Pro, refining polygon vertex accuracy, and exporting the resultant KML to shapefile format using QGIS (v 3.26). All spatial data were referenced to the WGS 84 coordinate system. Mapping and visualization were carried out using QGIS and the mapview package[26] in R (v 4.2.1)[27] to produce regional and local maps showing site distribution and transect layout.
Sampling design
-
A transect-based survey method was employed at each camp, maintaining an effective observation width of 6 m on both sides of each accessible route. Systematic visual searches were carried out by two trained observers walking slowly between 08:00 and 15:00 h, ensuring consistent survey effort and detection probability.
Field sampling was conducted during the third, fourth, and fifth ABAC surveys, during which a total of 13 transects were traversed: five transects (10.69 km; 0.06414 km2) during the third ABAC, four transects (35.89 km; 0.21534 km2) during the fourth ABAC, and four transects (19.96 km; 0.11976 km2) during the fifth ABAC, totaling a cumulative survey distance of 66.54 km, covering 0.39924 km2.
Each transect was traversed once per camp during the respective survey period. Observations missed during the initial pass were recorded on the return path[28]. All specimens occurring within the 6 m belt on either side of the transect were documented as individual observations. For large fallen logs or tree trunks, each log was treated as an independent sampling unit, with multiple individuals on the same substrate recorded separately. For each record, geographic coordinates were logged using a Garmin eTrex GPS, and associated environmental variables (habitat type, substrate characteristics, decay class, and moisture level) were documented. Habitat descriptors were scored on semi-quantitative ordinal scales (1–5) to maintain consistency among transects. Field photographs were taken using a Nikon D5300 equipped with a 50 mm prime lens and a Tamron 70–300 mm 1:2 macro lens. Specimens that could be reliably identified in the field were recorded without collection, whereas others were collected and identified at camp using standard taxonomic literature and monographs[1–4,9–11,20,28]. Vernacular names of associated trees and substrates were obtained from local forest staff familiar with the vegetation.
Predictor variables
-
Predictor variables for Myxomycete habitat suitability modeling were compiled from multiple open-access sources and represented climatic, topographic, edaphic, and broader environmental conditions relevant to temperate forest ecosystems. Monthly climate layers that included minimum, mean, and maximum temperature (°C), precipitation (mm), solar radiation (kJ·m−2·day−1), wind speed (m·s−1), and water vapour pressure (kPa) were obtained from the WorldClim v 2.1 dataset at 30-arc-second resolution for the period 1970–2000, together with 19 bioclimatic variables describing long-term climatic patterns[29,30]. Soil parameters were retrieved from SoilGrids (
https://soilgrids.org ) following ISRIC (2022) protocols using R[31]. Forest Canopy Height raster (2019) was sourced from the GLAD GEDI dataset(https://glad.umd.edu/dataset/gedi ), and NDVI from 2020 was acquired via EOS Landviewer (https://eos.com/landviewer) . Elevation was derived from the Shuttle Radar Topography Mission (SRTM) Digital Elevation Model raster, while topographic variables, namely slope and aspect, were derived from the elevation raster in R. All raster layers were clipped and masked to the boundary of Neora Valley National Park (NVNP) before analysis.Species occurrence data were recorded using a Garmin eTrex handheld GPS receiver. Given the known spatial uncertainty of consumer-grade handheld GPS units under dense forest canopy, positional error typically ranges from 5–15 m, a consideration that informed the subsequent harmonization of all spatial datasets. To maintain consistency across predictors and avoid mismatches between spatial resolutions, all continuous variables were standardized to the native resolution of the Forest Canopy Height dataset (0.00025° ≈ 27.83 m). Continuous layers, including climate, soil, NDVI, and topography, were resampled in R using bilinear interpolation, which is suitable for continuous surfaces. Categorical variables, such as Landclass, were resampled using nearest-neighbour interpolation to preserve original category integrity.
This scale was ecologically appropriate for Myxomycetes in temperate forests, where the distance decay principle shows that community similarity can decline over short distances, approximately 20 m[32]. This pattern indicates that dispersal limitation and fine-scale environmental heterogeneity operate at spatial scales comparable to, or finer than, standardized resolution. A coarser resolution could obscure these mesoscale ecological processes, while a much finer resolution would be unsuitable because GPS positional uncertainty reduces the benefit of using extremely high-resolution grids. Studies have shown that very fine spatial surfaces provide minimal advantages when location error remains high[33].
The selected 27.83 m grid captured essential meso-scale drivers of Myxomycete distribution, including canopy height variation, topographic complexity, and vegetation productivity, which act as proxies for micro-scale factors such as moisture availability and substrate decomposition. Although individual microhabitats remain below the resolution of the environmental layers, their effects were implicitly integrated through these broader gradients. This modelling approach provided a robust baseline for NVNP, where no previous spatial modeling of Myxomycetes has been conducted. It also offered substantially greater spatial detail than earlier MaxEnt models for the group, viz. 2.5 arc m (4.630 km2 at the equator)[34] and 30 arc s (0.926 km2 at the equator)[35,36]. Consequently, the habitat suitability maps generated here represent potential distributions rather than precise microhabitat occurrences, providing an ecologically coherent interpretation of how micro-scale ecological processes are nested within broader environmental gradients across the NVNP landscape[37].
Variable selection
-
The USDM package was utilized to assess the Variance Inflation Factor (VIF) values for cropped predictor variables, retaining only those with VIF < 10[38,39]. Subsequently, variable selection was performed using the covsel package[40], with partial modification to enhance its functionality. Covsel employs a two-step embedded covariate selection process: collinearity filtering in the first step, followed by algorithm-specific embedding for GAM, RF, and GLM in the second step. Additionally, for this study, MaxEnt outputs from all three subsampling criteria—namely, cross-validation, subsampling, and bootstrapping—were incorporated, with weights assigned to the top-performing methods. While MaxEnt's variable ranking is not inherently included in covsel, its integration into the selection process was essential to refine variable importance and ensure robust species distribution modeling outcomes.
Ecological Niche Model building and model selection
-
Ecological Niche Model (ENM) was performed with flexsdm package in R[41], employing glm (Generalized Linear Model), gbm (Gradient Boosting Machine), svm (Support Vector Machine), MaxEnt (Maximum Entropy), gam (Generalized Additive Models), and net (Neural Network) algorithms along with their respective Ensemble of Small Models (ESM), using nine presence and 10,000 background pseudo-absence data. K-fold random partitioning methods (5 folds, 100 replicates each) were used for partitioning of occurrence and background samples[42]. Thresholds used for individual model runs were maximum sensitivity and specificity 'max_sens_spec', equal sensitivity and specificity 'equal_sens_spec', maximum jaccard 'max_jaccard', maximum sorensen 'max_sorensen', and maximum F-measure on presence-background 'max_fpb'. The model was selected based on the highest values of TSS and AUC[43]. Response curves were generated to show how each environmental variable affects the model prediction by showing the predicted probability of presence changes as each environmental variable is varied. To assess this, 10,000-replicate bootstrap MaxEnt runs were performed.
-
It is important to note that sporocarp production in myxomycetes is inherently episodic and temporally variable, and therefore the observed patterns should be interpreted in light of sampling duration and revisitation frequency, which are known to influence detectability in field-based surveys. The patterns documented in the present study are broadly consistent with reports from tropical and subtropical regions elsewhere, suggesting that the observed ecological responses reflect general myxomycete behavior rather than site-specific anomalies.
Diversity
-
The Eastern Himalayas, particularly the Darjeeling Himalayas, hold numerous exclusive records of Myxomycetes, such as Cribraria aurantiaca Schrad., 1797; Licea erecta K.S. Thind & Dhillon, 1967; Badhamia capsulifera (Bull.) Berk. 1853; Craterium concinnum Rex, 1893; Physarum bethelii T.Macbr. ex G.Lister, 1911; Physarum durjoylingum Lodhi, 1934; Physarum spinulosum K.S. Thind & H.S. Sehgal, 1964; Physarum variegatum K.S. Thind & Dhillon, 1967; Physarum viride var. hinnuleum G. Lister, 1924; Willkommlangea reticulata (Alb. & Schwein.) Kuntze, 1891; Diderma badhamioides K.S. Thind, 1977; Diderma darjeelingense K.S. Thind & H.S. Sehgal, 1964; Diderma simplex (J.Schröt.) G. Lister, 1911; Lamproderma echinulatum (Berk.) Rostaf., 1876; Stemonaria reticulospora Nann.-Bremek., R. Sharma & K.S. Thind, 1984; Arcyria globosa Schwein, 1822; Perichaena quadrata T. Macbr., 1899, some of which are new records to science[9].
The preliminary study demonstrated that the Myxomycetes assemblage of the NVNP includes six species. During the field surveys, eight observations of Myxomycetes were recorded, including two from the third ABAC represented by Physaraceae, five from the fourth ABAC, including representatives from Stemonitidaceae, Reticulariaceae, and Physaraceae, while the fifth ABAC represented members from Stemonitidaceae only. These observations also included multiple sightings of the same specimen in the same ABAC as well as different ABACs.
Members of Myxomycetes were recorded from three families (Table 1, Fig. 2), represented by saprotrophic Stemonitis fusca Roth and Trichia typhoides Bull. from the family Stemonitidaceae; Lycogala epidendrum (L.) Fr. and Tubifera ferruginosa (Batsch) J.F. Gmel. from Reticulariaceae, and moss-associated Fuligo muscorum Alb. & Schwein. and Fuligo septica (L.) F.H. Wigg. from Physaraceae (Table 1). The analysis of species abundance, density, and frequency indicates that Fuligo muscorum, Lycogala epidendrum, Stemonitis fusca, and Tubifera ferruginosa were each observed with an abundance of 2, a density of 2.50, and a frequency of 20. Fuligo septica and Trichia typhoides exhibited an abundance of 1, a density of 1.25, and a frequency of ten each. While S. fusca, T. typhoides, L. epidendrum, and T. ferruginosa have been previously reported from Darjeeling, West Bengal, India, Fuligo septica represents a new record for West Bengal, and Fuligo muscorum is a new record for India.
Table 1. Occurrence records, habitat and brief morphological details of recorded Myxomycetes members from NVNP.
Sl. No. Species Family Habitat Occurrence location Morphological characteristics and remarks 1 Stemonitis fusca Roth Stemonitidaceae Fallen log of Engelhardtia spicata (Mauwa) 27.02289° N, 88.71773° E; 27.10615° N, 88.69744° E; Fruitbodies gregarious, sporangium stiped, elongated, bundle in brown mass. It has been earlier reported from Darjeeling. 2 Trichia typhoides Bull. Stemonitidaceae Upon decaying fern 27.10616° N, 88.71839° E; Fruitbodies gregarious, whitish spathulate jelly like fruitbody with stipe; stipe translucently visible in the upper part. It has been earlier reported from Darjeeling. 3 Lycogala epidendrum (L.) Fr. Reticulariaceae Upon fallen woody debris
including Eurya japonica
(Jhingeni)27.09514° N, 88.68927° E; 27.0957° N, 88.6952° E; Fruitbodies gregarious, sessile, initially semi solid greyish brown, becoming delinquescent with oozing pinkish liquid. It has been earlier reported from Darjeeling. 4 Tubifera ferruginosa (Batsch) J.F. Gmel. Reticulariaceae Upon moss 27.107212° N, 88.741802° E;
27.111986° N, 88.734456° E;Fruitbodies gregarious, salmon coloured, in mass, sporangium in rosette. It has been earlier reported from Darjeeling. 5 Fuligo muscorum Alb. & Schwein. Physaraceae Bark of Bante Katus (Lithocarpus pachyphyllus), upon Moss;
Machilus (Kawlo) bark, upon moss27.063528° N, 88.770917° E; 27.059139° N, 88.770083° E; Fruitbodies gregarious, sub-lunar in appearance, luteous with whitish mucilaginous appendage. It is a new record for India. 6 Fuligo septica (L.) F.H. Wigg. Physaraceae Fallen log of Engelhardtia spicata (Mauwa) tree 27.02289° N, 88.71773° E Fruitbodies gregarious, in bulky mass. It is a new record for West Bengal. 
Figure 2.
(1) Fuligo muscorum. (2) Fuligo septica. (3) Stemonitis fusca. (4) Lycogala epidendrum. (5) Tubifera ferruginosa. (6) Trichia typhoides.
Distribution
Variables driving Myxomycetes distribution in NVNP
-
Initially, the MaxEnt results were tabulated (Table 2), and the top eight variables were selected based on weighted ranking. These MaxEnt rankings were subsequently integrated into the covsel output (Table 3). Variables appearing in at least three out of four models (GLM, GAM, RF, and MaxEnt) were selected. This filtering process identified five key variables: Aspect, NDVI, BD100_200 (Bulk Density at 100−200 cm depth), PH100_200 (pH at 100−200 cm depth), and Clay0_5 (clay content at 0−5 cm depth) as critical for the distribution of Myxomycetes in Neora Valley National Park (NVNP). Notably, NDVI and BD100_200 were represented in all four models, highlighting their significance in predicting Myxomycete distribution.
Table 2. Variable ranks as per MaxEnt runs.
Covariate CA BA SA Rank percentile CPA BPA SPA Rank percentile Rank average WR CA BA SA CPA BPA SPA (1) (2) (3) (4) (5) (6) (7) (8) (9) (10) (11) (12) (13) (14) (15) Aspect 25.8019 17.4301 19.0647 1 1 1 2.3843 19.1714 7.6956 7 1 4 2.5 1 BD100_200 20.5783 14.5925 14.0781 2 2 2 37.6499 4.2306 19.0537 1 6 2 2.5 1 PH100_200 10.0475 11.5682 10.753 5 3 3 0 19.1173 31.1466 14 2 1 4.666667 2 NDVI 12.3987 6.1025 4.3781 3 7 7 12.92 10.563 5.5614 4 3 5 4.833333 3 Forheight 9.6644 8.1107 8.6882 6 5 5 2.7204 4.9563 8.0727 5 5 3 4.833333 3 VegMoistIndex 10.9958 9.1953 9.7247 4 4 4 15.0958 2.2328 1.443 3 12 13 6.666667 4 Clay0_5 0.5626 3.0456 3.1169 8 8 8 2.5117 5.6861 1.5444 6 4 11 7.5 5 Corfrag100_200 0.2428 1.5714 1.6061 9 13 13 1.3705 3.8452 2.1631 10 7 9 10.16667 6 CA = Crossvalidation_Avg, BA = Bootstrap_Avg, SA = Subsample_Avg, CPA = Crossvalidation_Permutation_Avg, BPA = Bootstrap_Permutation_Avg, SPA = Subsample_Permutation_Avg, Rank_Avg = average rank of column no. 5, 6, 7, 11, 12, 13), WR = Weighted Rank. Table 3. Variable selection according top ranks obtained from covsel output fortified by MaxEnt runs.
Covariate Rank Model Covariate Rank Model Covariate Rank Model Covariate Rank Model Aspect 1 glm Aspect 1 gam NDVI 1 rf Aspect 1 MaxEnt NDVI 2 glm NDVI 2 gam BD100_200 2 rf BD100_200 1 MaxEnt BD100_200 3 glm BD100_200 3 gam Clay0_5 3 rf PH100_200 2 MaxEnt Soil_N100_200 4 glm Sand15_30 4 gam OCDen0_5 4 rf NDVI 3 MaxEnt PH100_200 5 glm Forheight 5 gam Slope 5 rf Forheight 3 MaxEnt Slope 6 glm Soil_N100_200 6 gam Sand15_30 6 rf VegMoistIndex 4 MaxEnt Sand15_30 7 glm PH100_200 7 gam − − − Clay0_5 5 MaxEnt Bio3 8 glm Clay0_5 8 gam − − − Corfrag100_200 6 MaxEnt Variables which are present in atleast three models are marked bold. Top five ranked variables were selected for ecological niche modeling. Aspect plays a crucial role in influencing the microclimate of an area by regulating solar radiation exposure, which directly affects soil temperature, vapour pressure, and moisture availability[19]. In NVNP, Myxomycetes strongly favoured northern aspects, specifically between 0−6.5° from north (Fig. 3). This pattern aligns with established mesoclimatic principles: northern slopes in mountainous regions maintain cooler air and soil temperatures with lower vapor pressure deficits, particularly under moderate to dense tree canopy cover[13]. These cooler conditions reduce plant transpiration and soil evaporation, thereby maintaining elevated soil moisture levels essential for Myxomycete survival. On steep northern slopes, cooler temperatures can induce condensation within the litter layer, further enhancing water availability[44]. These combined factors contribute to the optimal conditions for habitat not only for Myxomycetes but also for bryophytes and other moisture-dependent biodiversity elements that characterize such humid forest environments. In contrast, south-eastern and south-western aspects experience higher temperatures and evaporation rates[19], resulting in drier conditions, less conducive to Myxomycetes establishment and sporulation.
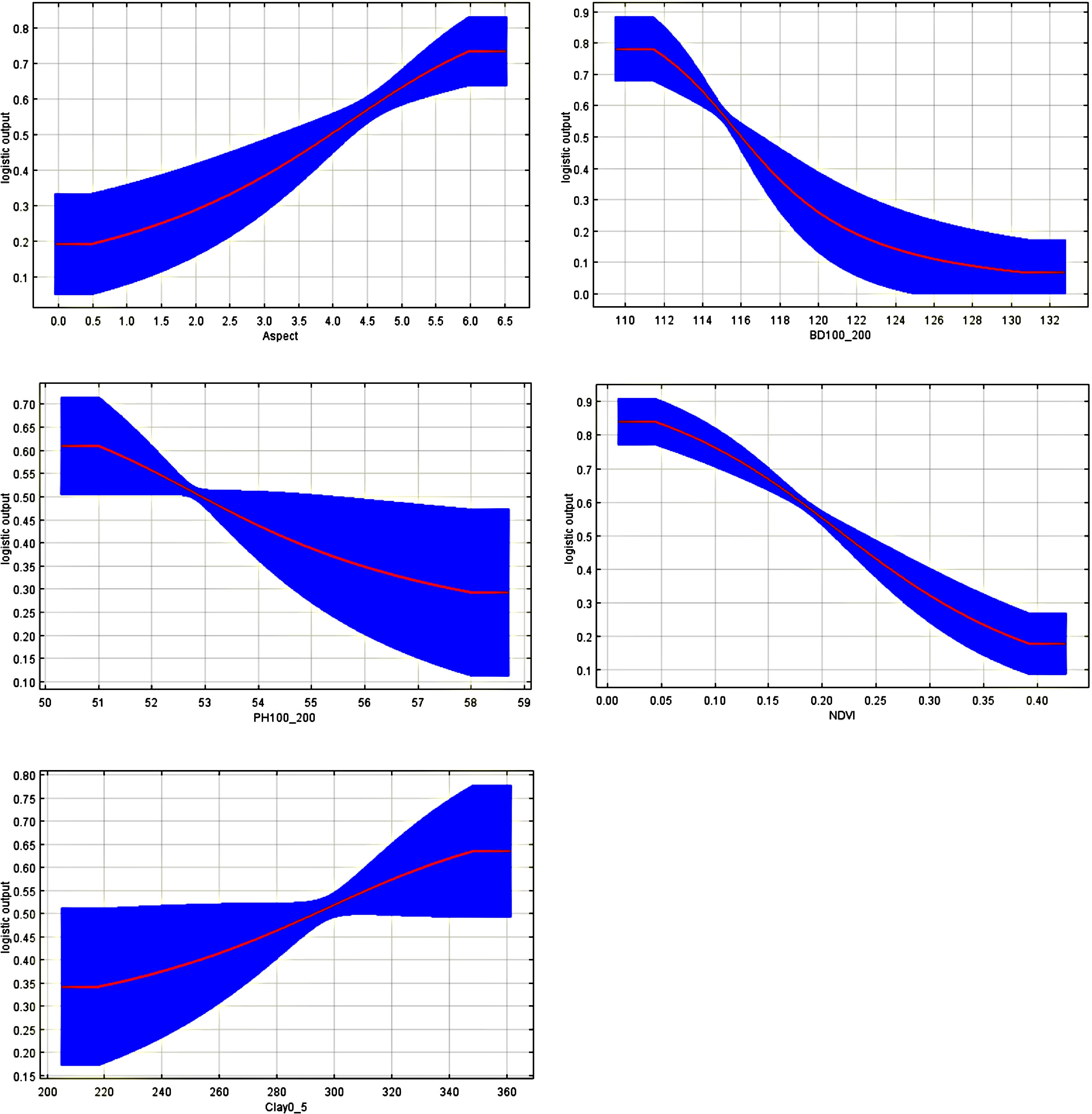
Figure 3.
The curves show the mean response of the 10,000 replicate MaxEnt runs (red), and the mean +/− one standard deviation (blue).
Moreover, aspect also influences the soil organic matter dynamics. Northern slopes typically accumulate higher organic matter content in the top 10 cm of soil compared to southern slopes, primarily because reduced temperatures slow organic matter mineralization rates[19]. This pattern, documented across diverse mountain systems including Mediterranean regions, appears to operate in NVNP as well. Northern and north-western slopes generally exhibit greater soil organic carbon stocks[45], a finding particularly relevant given that organic matter availability emerged as a key driver of Myxomycetes distribution in this system[39]. Furthermore, phosphorus availability, another critical factor for decomposer communities, tends to be higher on northern and western slopes[24], creating nutrients that favour not only Myxomycetes but also their associated microbial communities and substrate-providing vegetation.
The relationship between bulk density and Myxomycetes occurrence, particularly at depth (100−200 cm), proved significant in the models (Fig. 3). Bulk density at deeper soil depths reflects the physical structure available to support tree root systems, with deeper layers typically exhibiting higher compaction due to higher bulk density values[21]. However, the ecological significance of Myxomycetes operates through indirect pathways. Tree root biomass concentrates primarily within the upper 70 cm of soil[22], where nutrient and water availability are highest. When bulk density increases, root elongation, diameter expansion, and mass density all decrease[21], ultimately limiting the root system's overall biomass production. This reduction in root biomass translates to decreased organic material inputs that would otherwise be available for Myxomycetes colonization. Conversely, lower bulk density values indicate reduced soil compaction and improved soil physical structure. These conditions are favorable for Myxomycetes-host interactions, as they allow for better aeration and greater air space, facilitating the decomposition processes that Myxomycetes rely on. Thus, bulk density indirectly influences the organic substrates available for Myxomycetes by regulating tree root systems and soil aeration.
Clay content in surface soils (0−5 cm) plays a multifaceted role in creating a suitable Myxomycete habitat. Higher clay concentrations contribute to stable aggregate formation, which enhances soil porosity and structural integrity, and are vital for microorganism activity, including the organic matter decomposition and nutrient cycling processes (Fig. 3). Clay particles, with their high specific surface area and charged surfaces, retain both water and nutrients[23], resulting in substantially greater water-holding capacity in clay-rich soils compared to sandy substrates. This water retention capacity directly influences moisture availability in Myxomycete habitats while modulating soil pH through effects on the movement and retention of acidic and basic substances within the soil profile. For instance, in the phosphorus-depleted tropical forest systems, increasing clay content from 0% to 67% corresponded with improved tree growth and substantially higher soil organic carbon stocks[23]. Clay content also correlated positively with the arbuscular mycorrhizal fungal abundance, organisms essential for phosphorus mobilization and cycling, although plant-available phosphorus declined with increasing clay content due to strong sorption to clay surfaces. These complex interactions demonstrate how clay-rich surface soils in NVNP contributes to conditions favorable for both mycorrhizal fungi and the organic matter accumulation that supports Myxomycete communities, even in phosphorus-limited environments.
Soil pH has been observed to be one of the determining factors driving the occurrence of species of the Myxomycetes[8], with the Myxomycetes in NVNP showing a preference for acidic conditions, especially at deeper subsurface (Fig. 3). Soil pH is closely intertwined with clay content through buffering capacity. Clay-rich soil resists pH fluctuations when exposed to acidic or basic substances due to the high cation exchange capacity (CEC) of clay minerals such as montmorillonite and kaolinite[24], which minerals adsorb hydrogen ions and other cations, stabilizing pH within ranges that maintain nutrient availability. This stabilization is critical for maintaining the balanced availability of essential macronutrients such as phosphorus, potassium, calcium, and magnesium in acidic soils (pH < 7), while in alkaline soils (pH > 7), it regulates micronutrient availability, particularly iron and zinc. For Myxomycetes and their associated microbial communities, this pH stability likely ensures consistent substrate chemistry and decomposer community composition across temporal and spatial gradients.
Myxomycete occurrence was associated with lower NDVI values rather than with areas of maximum vegetation greenness (Fig. 3). NDVI reflects vegetation greenness and photosynthetic activity and is widely used as a proxy for plant productivity and carbon fixation; however, higher NDVI values in Neora Valley National Park correspond to closed-canopy forests where carbon is largely retained in living biomass and limited amounts enter decomposition pathways. In contrast, lower NDVI values characterize structurally heterogeneous forest patches with canopy gaps, fallen trees, and accumulated coarse woody debris, which provide favorable substrates for Myxomycetes. Although these sites exhibit lower NDVI, they support increased substrate availability and suitable habitats to decomposers. The presence of bryophytes further enhances habitat suitability by retaining moisture, moderating temperature fluctuations, and contributing organic matter through decomposition[46]. Consequently, NDVI functions less as a direct proxy for carbon availability than as an indicator of forest structural complexity and substrate dynamics. In mountainous landscapes, NDVI patterns are also linked to slope stability, as vegetation density influences soil erosion and slope reinforcement through root systems[25].
Predictive biogeography through ensemble of small models
-
This study utilized the Ensemble of Small Models (ESM) approach, a robust methodology applied to various species distribution models (SDMs), including Generalized Linear Models (GLM), Generalized Boosted Models (GBM), Support Vector Machines (SVM), MaxEnt, Generalized Additive Models (GAM), and Neural Networks (NET). By employing both standard and ESM versions of these SDMs, we generated predictive models for the distribution of Myxomycetes within Neora Valley National Park (NVNP). The outcomes of these models are presented in Fig. 4. To identify the most optimal model, we prioritized those exhibiting the highest mean Area Under the Curve (AUC) and True Skill Statistic (TSS) values, following established model performance criteria[42].
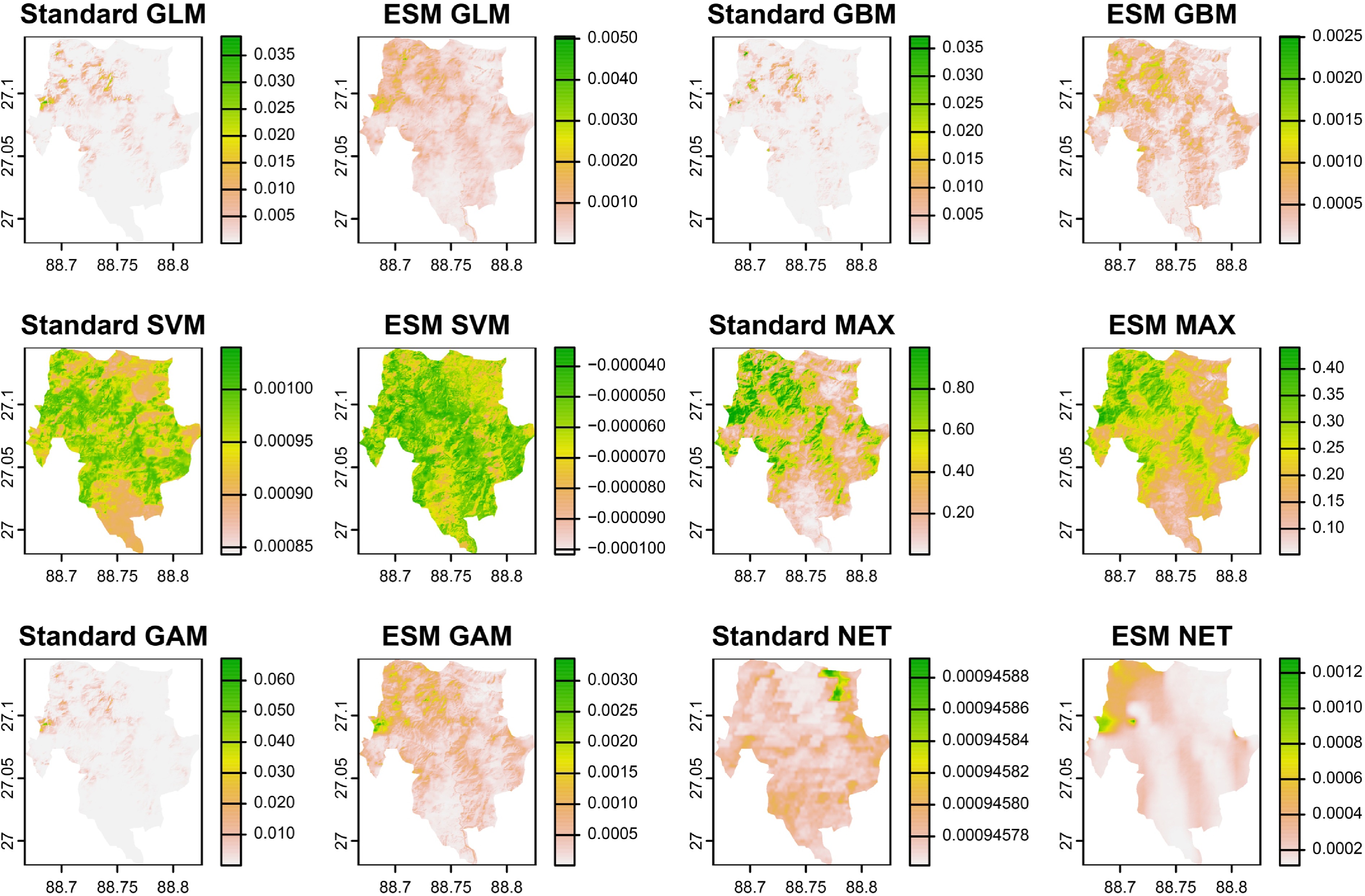
Figure 4.
Environmental suitability of the Myxomycetes from the output of standard and ESM versions of GLM, GBM, SVM, MAX, GAM, and NET performed through flexsdm package in R.
Out of the 65 models constructed, the ESM version of MaxEnt with a threshold of Maximum Sensitivity and Specificity demonstrated the most accurate predictive performance, achieving a mean TSS value of 0.72354 and a mean AUC of 0.816929 (Fig. 5). These high values reflect the model's strong discriminatory power, with maximum sensitivity and specificity thresholds, making it the most reliable predictor for Myxomycetes distribution in NVNP. Notably, the superior performance of the ESM-MaxEnt model is consistent with its efficacy in modeling species with limited occurrence data, as ESM is particularly suitable for rare or poorly collected species[47]. This is achieved by generating bivariate models through pairwise combinations of predictor variables and subsequently averaging suitability values, weighted by Somers' D statistic, to produce a robust ensemble.
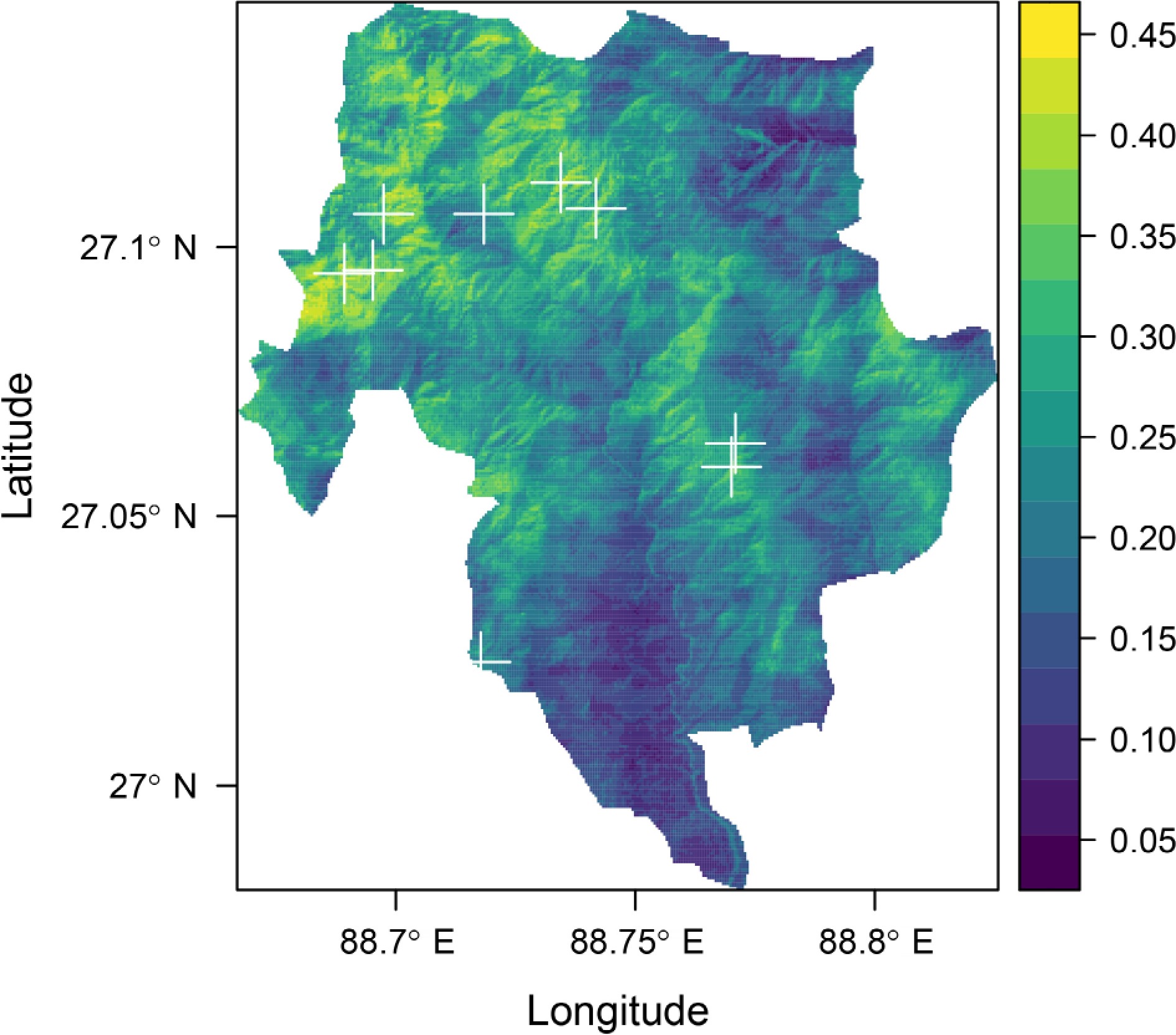
Figure 5.
Environmental suitability of the Myxomycetes from the output of ESM MaxEnt with threshold of maximum sensitivity and specificity in NVNP. (+) symbols indicate occurrence points.
Although the standard MaxEnt model has been successfully applied in previous studies, including to model Myxomycetes distribution in Costa Rica[48], its performance in this study was marginally lower than that of the ESM-MaxEnt model. Specifically, the standard MaxEnt model yielded a mean TSS value 0.0087 lower and a mean AUC value 0.007949 lower than the ESM variant. Despite the small differences, these findings highlight the advantage of employing ESM, particularly in complex, data-limited environments such as NVNP. This advantage can be attributed to ESM's ability to mitigate overfitting and enhance the generalizability of predictions by leveraging smaller, simpler models and integrating them into a more comprehensive framework.
The application of ESM within this study aligns with prior research from Costa Rica and Southeast Asia, where ESM-based approaches demonstrated increased predictive power in regions with insufficient sampling[49]. Findings further validate the suitability of ESM for biogeographical studies in tropical regions, especially in biodiversity hotspots like NVNP, where ecological data collection is often hindered by logistical challenges.
Key environmental variables were identified as significant predictors of Myxomycetes habitat suitability in NVNP. Notably, the ESM-MaxEnt model highlighted northern-facing slopes, lower Normalized Difference Vegetation Index (NDVI) values, and low bulk density soils as crucial factors influencing species distribution (Fig. 5). These variables, which include aspect, NDVI, and soil properties (e.g., bulk density and clay content), are known to significantly impact local habitats by influencing moisture availability, organic matter accumulation, and soil structure—factors critical to the growth and dispersal of Myxomycetes.
The significance of northern-facing slopes as high-suitability areas for Myxomycetes can be attributed to their cooler temperatures and higher moisture retention compared to southern aspects, as documented in several microclimatic studies[18]. These conditions favour Myxomycetes, which are sensitive to desiccation and require moist environments for their plasmodial stage. Similarly, high NDVI values reflect dense vegetation cover, which provides abundant organic substrates for Myxomycetes growth and facilitates active decomposition processes, further contributing to favorable conditions for these organisms[46].
Moreover, the findings related to soil bulk density and clay content align with previous studies that highlight the importance of soil texture and structure in microbial habitat suitability. Soils with lower bulk density have greater porosity and water retention capacity, creating optimal conditions for Myxomycetes colonization by enhancing moisture availability and aeration[21]. In addition, soils rich in clay content offer improved cation exchange capacity, which supports nutrient availability and fosters a more stable, fertile environment for microbial activity[23].
The integration of these environmental factors into the ESM-MaxEnt model not only provides a more comprehensive understanding of the ecological niches occupied by the Myxomycetes in NVNP but also emphasizes the importance of fine-scale environmental heterogeneity in predicting species distributions. This study contributes to the growing body of knowledge on Myxomycetes biogeography and emphasizes the value of employing ESM approaches in regions where species distribution data is sparse or incomplete.
The application of ESM, particularly when combined with MaxEnt, offers a powerful tool for enhancing the accuracy of species distribution models. This methodology is especially pertinent for under-sampled environments like NVNP, where traditional SDMs may be prone to biases due to limited occurrence data. By overcoming these limitations, ESM enables more precise identification of key environmental drivers and enhances the ability to predict species distributions, thereby offering valuable insights for conservation planning and biodiversity management.
Moreover, it is important to highlight the ecological link between Myxomycetes and their host trees. The environmental variables that were found to be significant for Myxomycetes, such as aspect, soil moisture, organic matter, and pH, are also critical for the health and growth of the host trees. The distribution of Myxomycetes is intricately linked with the ecology of their host trees[49]. The presence of healthy, mature trees creates a stable habitat condition with favorable moisture levels, organic matter input, and shade, all of which support the growth of Myxomycetes. Therefore, the environmental factors that promote Myxomycetes distribution are also indicators of the broader ecological health of the forest ecosystem.
-
This study records six Myxomycete species in Neora Valley National Park (NVNP) and elucidates the environmental drivers shaping their distribution in the region. Myxomycetes were found to favour northern-facing slopes with lower NDVI values, higher clay content, lower bulk density, and specific subsurface pH regimes that collectively create cool, moist, organic matter-rich habitats essential for Myxomycete establishment.
The inverse relationship between NDVI and Myxomycete occurrence merits particular attention. Lower NDVI in this context reflects complex forest patches with canopy gaps and accumulated coarse woody debris, which translates to increased availability of carbon sources, which is essential for the establishment of Myxomycetes. Methodologically, the MaxEnt-based Ensemble of Small Models approach proved effective despite limited occurrence data (mean AUC = [0.816], TSS = [0.724]), validating its application, especially in biodiversity-rich but data-limited regions. The 27.83 m raster resolution captured mesohabitat gradients while accommodating GPS uncertainty, providing substantially finer spatial detail than previous global Myxomycete distribution models. However, the temporal snapshot likely underestimates true diversity, and models predict habitat suitability rather than precise occurrence locations, highlighting the need for seasonal surveys and independent validation datasets.
Conservation priorities should focus on protecting northern-aspect forests, maintaining coarse woody debris abundance, and monitoring these climate-sensitive populations as potential indicators of warming-induced shifts. Future research should integrate further biodiversity assessments, seasonal sampling of microhabitats, molecular approaches to detect cryptic diversity, and experimental studies linking substrate chemistry to community assembly, thereby illuminating Myxomycetes' functional roles in nutrient cycling and ecosystem functioning of the Eastern Himalayan ecosystem.
-
The authors confirm contribution to the paper as follows: study conception and design, data collection: Pradhan P; analysis and interpretation of results, draft manuscript preparation: Giri S, Pradhan P. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding authors on reasonable request.
-
The authors would like to thank the Department of Forests, Government of West Bengal, for organizing Annual Biodiversity Assessment Camps in Neora Valley National Park. Furthermore, the corresponding author is thankful to the West Bengal Biodiversity Board for allowing him to participate in the said exercise. This work would not have been possible without support of Shri Ujjal Ghosh, IFS (Retd.); Prof. Silanjan Bhattacharya; Shri Animesh Bose; Shri Rajendra Jakher, IFS; Shri D.B. Basnet, IFS (Retd.); Mrs. Nisha Goswami, IFS; Shri Janmejay Pal, WBFS; Miss Rhea Ganguly, WBFS; Late Sahadev S. Giri, Range Officer, Lower Neora Range; Mrs. Sujata Gurung, Range Officer, Upper Neora Range; Beat Officers of Lower Neora Range and Upper Neora Range; Late Siddhartha Roy, IFS, ex Member Secretary, W.B. Biodiversity Board; Dr. A.K. Sanyal, ex-Chairman, W.B. Biodiversity Board; Dr. Himadri Sekhar Debnath, Chairman, W.B. Biodiversity Board, and Dr. S.N. Ghosh, Senior Research Officer, W.B. Biodiversity Board. Sincere thanks are also due to all the experts participating in ABACs, all the frontline forest staff, especially Shri Buddha Singh Rai, Shri Bobby Bhujel, Shri Yugal Rai, Shri Pritam Mahat, Shri Ajit Rai, Shri Subhash Chettri, Shri Rupen Lepcha, Shri Ram Kumar Rai, other porters, and drivers.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Giri S, Pradhan P. 2026. Beyond the obvious: insights into the diversity and ecological niche of the Myxomycetes in temperate forests of Eastern Himalaya. Studies in Fungi 11: e009 doi: 10.48130/sif-0026-0007 

Beyond the obvious: insights into the diversity and ecological niche of the Myxomycetes in temperate forests of Eastern Himalaya
- Received: 19 September 2025
- Revised: 25 January 2026
- Accepted: 27 January 2026
- Published online: 13 March 2026
Abstract: The ecological distribution of the Myxomycetes, a diverse group of slime molds, is shaped by complex interactions involving environmental and edaphic factors. This study explored the predictive distribution of the Myxomycetes in Neora Valley National Park (NVNP), West Bengal, India, focusing on the influence of aspect, bulk density, clay content, pH, and vegetation health (as measured by NDVI). Field surveys were conducted across varied forest habitats, and environmental niche models were developed using MaxEnt-based Ensemble of Small Models, integrating multivariate environmental data and species occurrence points. Findings suggest that the Myxomycetes are predominantly distributed on northern-facing slopes with high aspect values, favouring cooler and more humid micro-environments. Bulk density and clay content were also critical in influencing the availability of substrates for Myxomycetes, with lower bulk density and higher clay content promoting better soil aeration and moisture retention. Soil pH played an important role in nutrient availability, further modulating habitat suitability. Use of Normalized Difference Vegetation Index (NDVI) as a covariate revealed that Myxomycetes preferred areas with lower vegetation density, where organic matter and dead wood were more abundant, facilitating decomposition processes. MaxEnt-based Ensemble of Small Models showed high predictive performance (mean TSS = 0.724, mean AUC = 0.816) in identifying suitable habitats for Myxomycetes. These findings have important biogeographic insights for the Myxomycetes, as well as an important contribution to the identification of potential areas for protection of their habitats in NVNP. Future studies should focus on further biodiversity assessments, exploring the effects of climate change on mesohabitats, and investigating the interactions between Myxomycetes, their host trees, and other microbial communities. This will enhance the understanding of their role in nutrient cycling and ecosystem functioning.