








-
Oligodendrocytes (OLs) and Schwann cells (SCs) are myelin-forming cells in the CNS and the PNS, respectively. Their main function is to form insulating myelin sheaths around axons, thereby increasing the speed and efficiency of action potential propagation. Despite the similarity in axon wrapping, OLs and SCs employ entirely different mechanisms to initiate myelination. SCs require biochemical signals provided by axons, specifically Neuregulin-1 (NRG1), to initiate myelination[1,2]. By contrast, the process of myelin formation in CNS OLs does not appear to involve the same biochemical signals[3]. The myelin formation by OLs can be activated solely by geometrical cues from axons. In vitro studies have shown that oligodendrocyte precursor cells (OPCs) can differentiate and form myelin sheaths on pseudo-axons such as plastic nanofibers[4] and silica micropillars[5]. While Schwann cells can sense environmental stiffness during peripheral myelin formation through the mechanosensitive channels Piezo1/2, they cannot initiate myelination on pseudo-axons with proper signals omitted[6]. In vivo evidence indicates that OLs prefer to wrap axons larger than 0.5 μm, demonstrating that geometrical cues from axons bias the selection of OLs[4,7]. However, the machinery by which OLs sense the geometric shape of axons and transduce this information into intracellular pro-differentiation signals remains largely unknown. Previous studies have shown that OPCs can sense substrate stiffness via the mechanosensitive ion channel PIEZO1, thereby regulating OPC differentiation and myelination[8]. Recently, mechanosensitive channels (MCs), molecular sensors that read the physical environment, have attracted significant attention for their role in regulating glial cell development.
In the study by Dereddi et al., it was first demonstrated that mechanical tension is an important signal in the process of myelin formation in the CNS, and the transmembrane protein 63A (TMEM63A) of the mechanically gated ion channel[9] is the most important mechanically-sensitive channel for sensing mechanical tension in OLs[10]. This channel can convert the membrane's stretching force into calcium (Ca2+) signals, thereby regulating myelin's geometric shape (thickness and length). The study not only demonstrated the crucial role of TMEM63A in developmental myelin formation but also linked its functional defect to human transient infantile hypomyelinating leukodystrophy-19 (HLD19)[11], providing a new molecular basis for understanding and treating myelin-related diseases (Fig. 1).
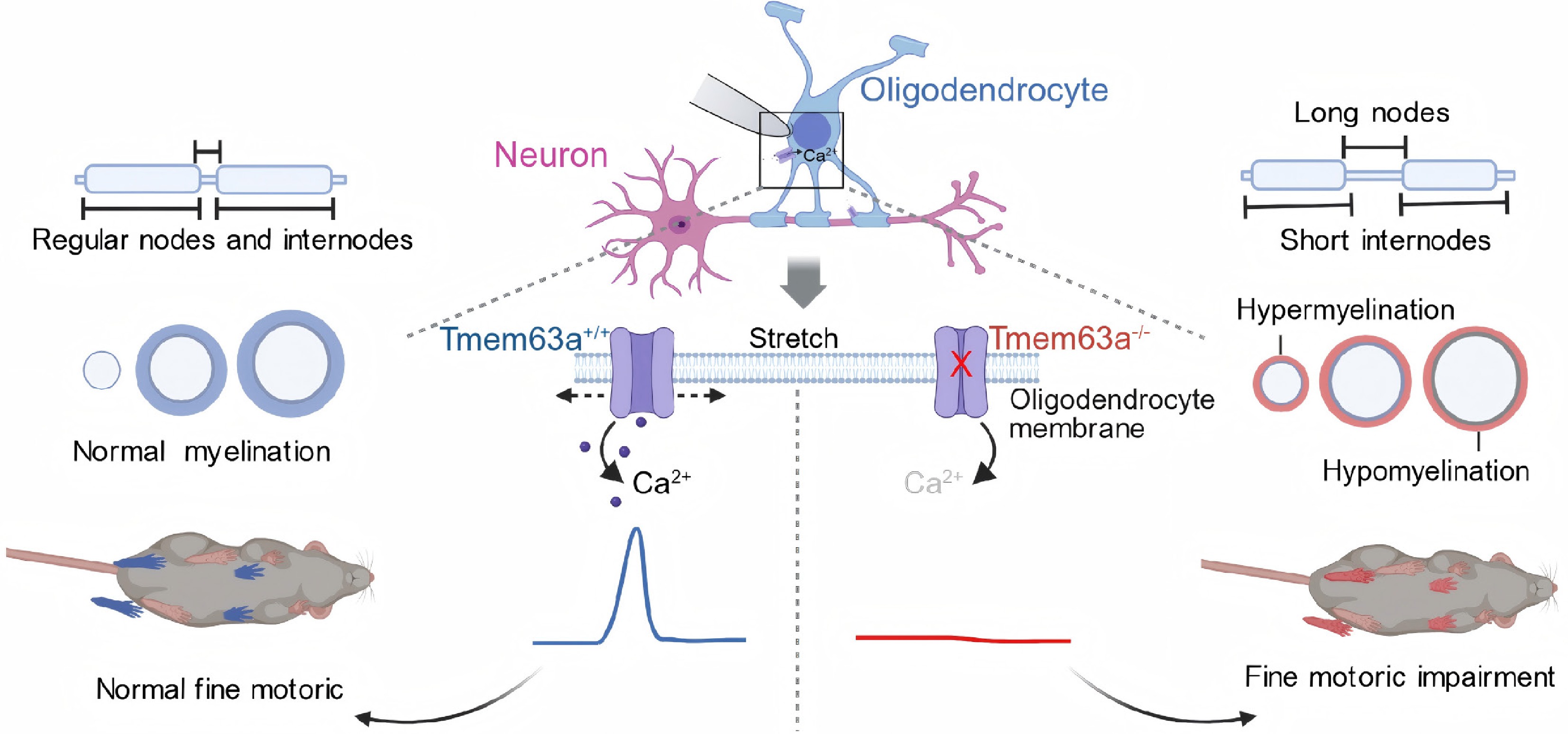
Figure 1.
TMEM63A-mediated mechanosensation regulates myelin geometry in the central nervous system. Oligodendrocytes sense axonal membrane stretch via the mechanosensitive ion channel TMEM63A, leading to Ca2+ influx and activation of intracellular Ca2+ signaling pathways. Through this mechanism, oligodendrocytes dynamically adjust myelin thickness and internode length according to axonal diameter. Loss of TMEM63A causes dysmyelination, characterized by thinner myelin on large-diameter axons, aberrantly thick ectopic myelination on small-diameter axons, reduced and more uniform internode lengths and increased node lengths.
Dereddi et al. first addressed an acute mechanical stimulus to one side of the OLs, confirming that OLs would experience a significant, rapid increase in intracellular Ca2+ when stimulated by mechanical force[12]. Moreover, the Ca2+ signal response could be transmitted throughout the cell, including its distal terminal, indicating that the mechanical signal perceived at a certain location can trigger a coordinated response across the entire cell. Further results proved that this mechanically sensitive Ca2+ response is not mediated by PIEZO1. Previous results have confirmed that PIEZO1 enables OPCs to sense the hardness of the basal environment, thereby regulating their proliferation and differentiation. However, PIEZO1 is not involved in the activation process of the Ca2+ in the mechanical response, indicating that this is a developmental stage-specific mechanical signal mechanism: OPCs can rely on the PIEZO1 signal pathway to sense the physical environment where they are located, while mature OLs regulate myelin formation through another mechanical pathway. This difference also makes it clear that mechanical perception in the oligodendrocyte lineage is not a single, fixed process but changes with cell differentiation and myelin formation.
To confirm that OLs possess mechanical sensitivity, the authors used single-cell RNA sequencing (scRNA-seq) to find that TMEM63A is the most abundantly expressed mechanosensitive channel in OLs, especially highly expressed in both newly formed and mature OLs, while other mechanical ion channels, such as PIEZO1, are expressed less. By using TMEM63A-eYFP reporter mice, it was further confirmed that TMEM63A is specifically expressed on the membrane of OLs. Further electrophysiological patch clamp recordings of OLs revealed that mechanical stimulation could induce inward currents in OLs, while OLs lacking TMEM63A showed almost no current response to mechanical tension stimulation. This finding indicates that TMEM63A is the main mediator of mechanical gating currents in OLs. More importantly, the loss of TMEM63A does not affect OL survival, suggesting that mechanical sensitivity is not a necessary condition for cell survival but is used to fine-tune myelin formation.
Next, the research was conducted in vivo in mice, revealing that in TMEM63A-knockout mice, there was a transient delay in myelin formation during development. In the early stage of development (P11), severe hypomyelination occurred in the cortical and corpus callosum regions, with a decrease in oligodendrocyte numbers; however, by P35, myelin levels had largely recovered, showing characteristics of 'temporary hypomyelination' similar to those of HLD19 patients. However, the ultrastructural analysis by electron microscopy revealed that in the TMEM63A-knockout mice, the abnormal thickness and length of the myelin sheath persisted into adulthood. It was found that the myelin sheath on large-diameter axons became thinner (with an increased g-ratio). While on small-diameter axons (which normally do not form myelin), there was an abnormally thickened ectopic myelin sheath. At the same time, the internode length of the formed myelin shortened and became more uniform, while the node length increased significantly. These results indicate that the loss of TMEM63A allows the myelin sheath to form, but the geometric shape of the myelin sheath cannot perfectly match the diameter of the wrapped axons. This abnormal change can directly affect the speed and efficiency of neural signal transmission (Fig. 1).
The long-term intravital imaging of zebrafish also further proved that the lack of TMEM63A would lead to a slower growth rate of myelin sheath, increased myelin sheath regression, and a decrease in the frequency of Ca2+ signal responses. These results also indicated that TMEM63A can regulate myelin sheath stability by modulating local Ca2+ signaling responses. In order to explore how mechanical tension guides the differential formation of myelin sheaths. Dereddi et al. conducted transcriptomic analysis. The results showed that the loss of TMEM63A led to downregulation of multiple vesicle transport-related gene expression in OLs, among which the most significant was myosin 5A (MYO5A)[12]. MYO5A is a motor protein regulated by Ca2+ signals and participates in the transport of mRNA particles within the cell. Immunofluorescence and single-molecule fluorescence in situ hybridization (smFISH) results indicated that MYO5A was co-localized with Mbp mRNA at the myelin interstitial regions, while the knockout of TMEM63A led to a decrease in Mbp mRNA expression. In summary, these results show that the mechanical tension of the membrane can trigger a Ca2+ signal response through TMEM63A and further mediate transport through the MYO5A-mediated pathway, transporting Mbp mRNA to sites where myelin formation is required, thereby achieving precise matching between myelin geometry and axons. Murthy et al. first reported that the absence of Tmem63A leads to a delay in the process of myelin formation[13]. However, Dereddi et al. further advanced this study, demonstrating that TMEM63A is not merely a switch for myelin formation but rather a precise regulatory factor governing myelin geometry. Alternatively, this result suggests that compensatory regulatory pathways may exist that do not involve mechanical signals, ultimately restoring the total amount of myelin but not correcting its ultrastructural defects. This finding suggests the existence of compensatory regulatory pathways, potentially independent of mechanical signals, that can ultimately restore the total amount of myelin without correcting the underlying ultrastructural defects (Fig. 1).
Although Dereddi et al. have elaborated in detail on how OLs regulate myelin formation through the mechanosensitive ion channel TMEM63A, some key issues remain unresolved. The exact mechanism by which the diameter of the axon is converted into the membrane mechanical tension that OLs can sense remains unclear. Is it possible that the curvature radius of the axon determines the membrane tension applied to the anterior edge of the inner tongue during the wrapping process? Specifically, compared to axons with smaller diameters, axons with larger diameters may increase the membrane tension during each successive wrapping, thereby activating TMEM63A. If the function of TMEM63A is rescued during the developmental period in TMEM63A knockout mice, will it be able to correct the defect in the geometric shape of myelin? Nevertheless, the work was the first to establish a direct link between TMEM63A-dependent regulation of mechanical sensing and myelin-related diseases. It demonstrates the direct regulatory role of mechanical tension in myelin formation. In clinical studies, mutations in TMEM63A can cause myelin dysplasia in HLD19, but the symptoms of this disease gradually improve with age. Similar phenomena were also observed in the TMEM63A-knockout mice, including abnormal myelin structure, slowed neural signal transmission, and fine motor defects. These findings not only explain the pathological mechanism of HLD19 but also indicate that TMEM63A may be a potential therapeutic target.
HTML
This work was supported by the National Natural Science Foundation of China (Grant No. W2511025).
-
Not applicable.
-
The authors confirm their contributions to the work as follows: manuscript conception, revision, and supervision: Mei F; manuscript writing and figure creation: Si T. All authors reviewed and approved the final version of the manuscript.
-
Data sharing is not applicable to this commentary as no datasets were generated or analyzed.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of China Pharmaceutical University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Si T, Mei F. 2026. Oligodendrocyte mechanosensitivity: translates geometric cues into myelin patterns? Targetome 2(3): e022 doi: 10.48130/targetome-0026-0021


|