








-
With global climate change and the impact of human activities on the ecological environment, frequent occurrences of drought have become one of the most severe environmental stressors limiting crop production[1,2]. On the other hand, the increasing population and improved living standards further demand higher levels of safety and stability in agricultural production. It is estimated that 70% of global freshwater consumption is attributed to agricultural production, while the proportion of water absorbed by crop roots from the soil and lost through Tr from leaves can be as high as 99%[3]. Water use efficiency (WUE) refers to the ratio of the amount of water used by a plant or system to the amount of biomass or yield produced. Therefore, enhancing WUE is crucial for achieving water-saving and yield balance in crop production. Although significant progress has been made in drought research over the past decades, translating the findings obtained in laboratory conditions into field crop improvement has remained challenging. The main reasons for this are, firstly, the assessment of drought tolerance in laboratory conditions often relies on plant survival rates, which are difficult to directly correlate with field yield indicators. Secondly, natural environmental factors such as water availability and light intensity exhibit high dynamics, making them more complex than laboratory conditions[4].
Stomata serve as the common pathway for plant water transpiration (Tr) and CO2 absorption, and the ratio of CO2 uptake to water loss at leaf or canopy scales is referred to as WUE[5]. WUE is an important target trait in water-saving crop breeding[6]; however, a long-standing issue is the excessive increase in stomatal sensitivity, which though reduces Tr and prevents plant wilting, hinders photosynthesis due to constrained stomatal conductance. This finally leads to a high WUE but a reduced biomass accumulation and low yield, which is quite common among natural crop germplasm[7,8]. Through combining physiological phenotyping and simulation models, Sun et al.[9] demonstrated that plants exhibiting reduced sensitivity to vapor pressure deficit (VPD) could theoretically gain an improved WUE; however, it is important to take into account the resultant decrease in Tr when aiming for higher yields. In this paper, we assert that the WUE trait is not static but rather a dynamic and plastic attribute, influenced by both environmental conditions and genetics. Plants that can rapidly (within minutes) optimize their WUE to capitalize on favorable environmental conditions exhibit a significant agronomic advantage.
-
It is noteworthy that there is a diurnal asynchrony observed between photosynthesis and Tr, leading to variations in WUE (Fig. 1a)[10]. The primary underlying factor for this difference is that Tr is primarily influenced by the VPD, which typically peaks around or after noon[11]. Conversely, during the morning period following sunrise, there is sufficient light intensity to support high rates of photosynthesis[10]. This suggests the existence of a specific phase characterized by an exceptionally ideal WUE, encompassing a high rate of photosynthesis and a low Tr. Du et al.[12] observed that the photosynthetic rate and stomatal conductance of rice leaves were significantly higher in the morning at 6:00 than in other periods. Fracasso et al.[10] observed that canopy photosynthesis exhibited a rapid increase, reaching a high level shortly after sunrise, while Tr increased slowly in accordance with the VPD trend, ultimately resulting in a high canopy WUE accompanied with high carbon assimilation before 7:00. Similar findings were reported by Yang et al.[13] and Gosa et al.[14] in tomatoes, and Sun et al.[9] in wheat.
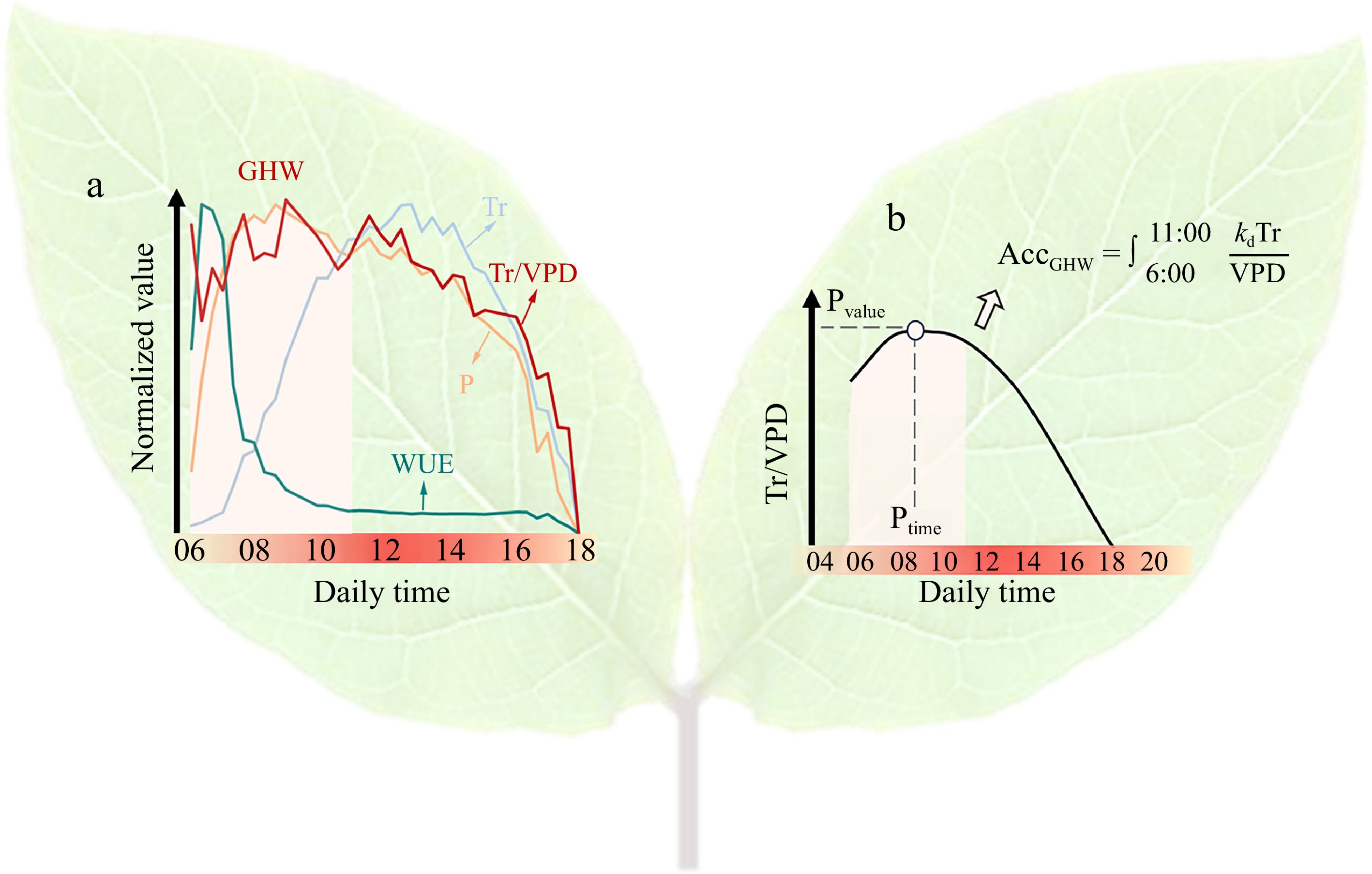
Figure 1.
A diagram illustrating the concept and definition of GHW and its quantitative components. (a) Tr, P, and WUE represent the transpiration rate, photosynthesis rate and water use efficiency measured at the whole-plant scale, respectively. VPD represents the environmental vapor pressure deficit. GHW is defined as the WUE during the 'golden hours', typically ranging from 6:00−11:00, during which WUE is significantly higher compared to other periods. After 11:00, WUE tends to stabilize. (b) Pvalue, Ptime, and AccGHW represent the peak value, peak time, and accumulative value of Tr/VPD during the golden hours, respectively. Kd is a specie-specific parameter. Picture is adapted from Sun et al.[20]. To accurately detect these continuous measurements, the system must precisely differentiate the signals emanating from the plants from environmental noises (see Fig. 2 for explanation).
Hence, there exists a theoretically optimal utilization pattern of WUE, where plants produce more photosynthetic assimilate during periods of high WUE and less during periods of low WUE throughout the day. This pattern can lead to a reduction in water usage while producing the same amount of biomass, which thereby increases the daily and seasonal WUE[15]. Gosa et al.[14] referred to such time period with higher WUE as the 'golden hour'. Building upon these prior studies, we introduce a novel physiological trait termed 'golden-hour' WUE (GHW, Fig. 1a), which is of significant importance for improving water-saving and yield balance in crops. The GHW trait is typically expressed between 6:00 and 11:00 in the morning compared to other periods, according to Sun et al.[9]. During this time window, plants are able to produce more biomass at the cost of consuming less water (Fig. 1a). From a breeding perspective, it is feasible to achieve a balance between water-saving and yield by selecting genotypes with superior GHW trait, i.e., exhibiting higher WUE and higher photosynthesis rate concomitantly for a longer duration of a day. This trait proves particularly advantageous in drought conditions. However, it is notable that the seasonal or climatic variations in sunlight intensity, duration, and temperature, significantly influence GHW traits. Hence, the timeframe from 6:00 to 11:00 is considered a general approximation of the optimal WUE period. In practical applications, it is advisable to monitor the daily variation pattern of 1/VPD, taking into account the reciprocal relationship between WUE and VPD. The period during which noticeable morning fluctuations in 1/VPD occur until stabilization can be identified as the appropriate range for GHW.
-
To enable harnessing GHW in crop improvement, it is prerequisite to establish a quantification method for this newly defined trait. This might involve, for example, comparing photosynthetic capacity during periods of GHW. To achieve this, it first requires dynamically measuring at least two of the following parameters: photosynthesis, Tr and WUE. Measuring plant gas exchange parameters allows for the simultaneous assessment of photosynthesis and Tr, enabling the direct calculation of WUE. However, both leaf-scale gas-exchange measurements (such as the Li-6800 photosynthesis system, LI-COR Inc., USA) and canopy-scale measurements (such as the canopy photosynthesis and transpiration system, CAPTS, Shanghai Millet Hill Biotech Co., Ltd., China) have limitations, such as the need for stable chambers or cuvettes, high sensitivity requirements, slow measurement speed, and high cost. These limitations greatly hinder high-throughput measurements of photosynthetic assimilation and WUE[6], impeding the process of discovering germplasm with excellent WUE utilization patterns and improving crop water relations through large-scale variety screening. With the technical advances in the phenomics era, new high-throughput physiological phenotyping technologies have emerged[16,17]. Among them, the noise-resilient diagnostic lysimeter array is a physiological phenomics platform that can automatically monitor whole-plant-scale water physiological parameters and environmental parameters, continually and simultaneously to all plants in the array, while maintaining cost-effectiveness (Fig. 2)[18] 'Plantarray', an exemplar of the weighing-type lysimeter physiological phenotyping system, enables high-throughput monitoring of whole canopy stomatal conducts, Tr and biomass gain.

Figure 2.
Refining data accuracy: How the Plantarray system enhances signal-to-noise ratio in lysimetry. This figure illustrates the Plantarray system's advancements (Plantarray 3.0, Plant-DiTech, Rehovot, Israel) in addressing challenges commonly associated with traditional lysimeters for measuring plant transpiration, stomatal conductance and biomass gain separately from evaporation. The system's innovative design and technological integration adeptly tackle difficulties such as distinguishing changes in soil weight attributable to soil moisture versus plant biomass, and accurately tracking non-plant related weight alterations due to soil-surface evaporation and pot-dripping. It also skillfully manages the application of diverse treatments and the maintenance of consistent soil water content for numerous plants concurrently, thereby overcoming the pot size effect and preventing salinity build-up in the soil. Key features of the Plantarray system: (1) Advanced sensing technologies: The Plantarray system employs temperature-compensated lysimeters, each connected to an individual controller with dual irrigation valves per plant. This configuration distinctly separates changes in soil weight caused by water dynamics from those due to biomass gain. Being a non-data logger system with short cables and an integrated analog-to-digital solution adjacent to each lysimeter, it ensures swift response times, significantly enhancing measurement accuracy. (2) Controlled drainage and multi-dripper system: Positioned in a controlled drainage green bath and supplied with several drippers, each plant's setup in the Plantarray system precisely monitors every water droplet entering and exiting through dripping or evaporation. This precision enables accurate real transpiration calculations and maintains consistent soil moisture levels, while effectively differentiating plant biomass from other elements. (3) Effective feedback control to handle variations: The Plantarray system's distinctive configuration efficiently addresses challenges such as salinity gradients and soil water content variations. Its carefully considered design also reduces the influence of pot size, promoting more consistent and controlled experimental conditions. (4) Environmental sensors and data analysis: Equipped with a suite of soil and environmental sensors and an advanced data analysis framework, the Plantarray system adjusts to changes in plant biomass and ambient conditions. This comprehensive setup normalizes data for plant size differences and varying environmental conditions, facilitating precise, comparable measurements across different plants and scenarios. For an in-depth understanding of the operational principles and effectiveness of the Plantarray system, see Gosa et al.[14].
However, due to its inability to sense changes in CO2 levels, the lysimeter system does not have the capability to directly monitor real-time photosynthesis. Fortunately, simulation models provide a pathway to estimate WUE by leveraging Tr and environmental parameters[15,19]. Expanding on this theory, Sun et al.[20] used data from Plantarray, which recorded Tr and VPD measurements at 3-min intervals, to estimate dynamic WUE and day matter production (DM):
$ {\mathrm{W}\mathrm{U}\mathrm{E}}_{i,\Delta t}=\dfrac{{\displaystyle\int }_{i}^{i\;+\;\Delta t}\dfrac{{K}_{\mathrm{d}}\mathrm{T}\mathrm{r}}{\mathrm{V}\mathrm{P}\mathrm{D}}}{{\displaystyle\int }_{i}^{i\;+\;\Delta t}\mathrm{T}\mathrm{r}} $ (1) $ {\mathrm{D}\mathrm{M}}_{i,\Delta t}={\int }_{i}^{i\;+\;\Delta t}\dfrac{{K}_{\mathrm{d}}\mathrm{T}\mathrm{r}}{\mathrm{V}\mathrm{P}\mathrm{D}} $ (2) where Kd is a specie-dependent parameter linked to the concentration of CO2 within cells.
$ {\mathrm{W}\mathrm{U}\mathrm{E}}_{i,\Delta t} $ $ {\mathrm{D}\mathrm{M}}_{i,\Delta t} $ When ∆t is shortened, for instance, with a data collection interval of 3 min, the instantaneous
$ {\mathrm{D}\mathrm{M}}_{i,\Delta t} $ https://github.com/Interstingsun/FPP ). Both Accdaily and fGHW exhibited significant inter-variety differences, allowing the selection of genotypes with superior GHW traits. The coefficient of variation (CV) for Accdaily is ~30%. Even with similar Accdaily values, the germplasm lines exhibited considerable variations in fGHW, with some reaching double the value of the lowest. As more germplasm lines undergo similar screening in the future, one can anticipate obtaining even greater disparities in the GHW trait across different genotypes. Therefore, the marriage of high throughput physiological phenotyping and mechanism modeling enables the quantification of the GHW traits. It is important to note that when the phenotyping is conducted in multiple experiments, maintaining consistent environmental conditions, particularly solar radiation, temperature and daylength, is critical to minimize batch effects stemming from environmental factors. -
Since the diurnal variation of WUE is mainly influenced by environmental VPD, it would be highly advantageous for striking a balance between water saving and yield if plants can regulate their physiological processes to achieve higher photosynthetic assimilation capacity during the GHW period. The following are proposed schemes and procedures for leveraging GHW-related traits in breeding practices to achieve this goal (Fig. 3).

Figure 3.
A schematic overview illustrating the practical procedures for leveraging GHW traits in breeding. Measurements of environmental parameters [e.g., air temperature (Tair) and relative humidity (RH)], transpiration rate (Tr), and net photosynthesis rate (Pn) are taken directly or inferred through high-throughput phenotyping platforms, such as spectrum-based, lysimeter-based, or gas exchange-based platforms. GHW traits include parameters related to Tr/VPD or Pn during the gold-hour WUE period (06:00−11:00). KASP: Kompetitive allele-specific PCR. GWAS: genome-wide association mapping. FM: functional mapping, a method to estimate mathematical parameters that describe the developmental mechanisms of trait formation and expression for each quantitative trait locus (QTL). MAS: marker-assisted selection. The superior genotypes possess favorable GHW traits, as reflected by optimized Pvalue, Ptime, and AccGHW. In making this figure, some images were adapted from some references: gas exchange[32], whole-genome resequencing[33], KASP[34], GWAS[35], FM[27], MAS[36].
Step 1, large-scale physiological phenotyping of a population, either natural populations consisting of a number of germplasm, or constructed pedigreed populations derived from crossing and/or backcrossing. Theoretically, any phenomics platform capable of directly or indirectly measuring transpiration or photosynthesis, along with recording environmental variables such as temperature and relative humidity for VPD calculation in a dynamic and continuous manner, can work for this purpose. Subsequently, based on the previous protocol, quantified parameters for GHW traits like AccGHW and fGHW are defined. However, apart from lysimeter systems, specific quantification methods may need to be customized and optimized.
Step 2, following acquisition of the physiological trait data, the genotypes of the measured plants should be resolved. This may typically be through high-throughput genotyping such as whole-genome resequencing, complexity-reduced genotyping, chip hybridization-based genotyping, or high-throughput PCR based genotyping like Kompetitive allele-specific PCR (KASP)[23−26]. Further dissection of the genomic regions and candidate genes controlling GHW traits can be achieved through joint phenotype-genotype analysis. This can be done by conducting genome-wide association analysis for natural populations, or by linkage mapping analysis for pedigreed populations such as F2 and recombinant inbred lines (RILs). More advanced mapping method tailored for dynamic trait mapping, such as joint functional physiological phenotyping-functional mapping, can be employed to more efficiently unravel the genetic network underlying GHW[27].
Step 3, developing molecular markers and utilizing them in molecular breeding to facilitate precise trait improvement. Based on the delimited QTL regions, e.g., AccGHW and fGHW, various types of DNA markers, such as SNPs, Indels and microsatellites that are linked to the GHW traits, will be useful for breeding purpose. SNPs may also be converted into KASP markers prior to use as required. These markers can then be used to screen for elite germplasm line from broader reservoir of germplasm pools or to facilitate selection of favorable offspring plants in breeding populations derived from crossing, backcrossing or selfing. Here, any modernized molecular breeding techniques, such as marker-assisted recurrent selection (MARS) and genomic selection (GS) are theoretically applicable. The ultimate superior genotypes are sought to possess the capacity to maximize assimilation during the golden hour and minimize transpiration during non-golden hours in plants, thereby enhancing the overall water use efficiency of crops while sustaining robust growth throughout the growing season.
-
Building upon the GHW trait, optimizing cultivation management alongside breeding efforts could present a novel strategy for synergistically enhancing crop productivity. Precision irrigation, grounded in integrated water and fertilizer management, currently plays a crucial role in smart agriculture. Current irrigation timing is driven by factors such as reference crop evapotranspiration (ET0), soil moisture, and leaf water potential, whose rationale remains focused on identifying instances of plant water deficit and tailoring irrigation responses accordingly[28−30]. However, future irrigation paradigms could incorporate the dynamics of GHW traits as a guide for feedback irrigation. Specific GHW parameters, such as Ptime, could serve as crucial benchmarks in identifying periods of heightened WUE and priming precise irrigation scheduling. This approach will be particularly beneficial in arid and semi-arid regions, as effective water budgeting relies on plants producing substantial dry matter during the GHW period, compared to conditions with ample water availability. Executing precise irrigation during this specific period will not only improve daily and seasonal WUE but also alleviate ineffective irrigation during alternative periods.
-
Traditional methods for selecting high-WUE crops based on carbon isotope discrimination often fall short of meeting the simultaneous demand for high yield, due to the inability to consider photosynthesis assimilation and transpiration traits concurrently[7,31]. The quantification and utilization of the dynamic GHW provide a promising solution for achieving comprehensive improvements in water saving and crop yield. This approach is distinctive, as it focuses not solely on drought tolerance but also on the plant's water utilization behaviors. This innovative and technologically advanced approach has the potential to reconcile the trade-off between water-saving and high-yield in crop production, facilitating yield increase under adequate water conditions and stable production under water scarcity. The core concept of this strategy is to enhance crop varieties by leveraging their inherent water regulatory traits, without the need for additional chemical inputs like anti-transpiration agents or genetic engineering methods, making it more environmentally friendly. Further elucidating the genetic determinants and responsible genes for the GHW traits in the future will establish a more robust technical foundation for their applications, ultimately contributing to global food security and the conservation of water resources.
-
The authors confirm contribution to the paper as follows: study conception and design: Jiang R, Sun T, Shi Z, Xu P; data collection: Jiang R, Shi Z; analysis and interpretation of results: Sun T, Xu P; draft manuscript preparation: Sun T, Moshelion M, Xu P. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This study is supported by the National Key Research & Development Program of China (China-Israel 2022YFE0198000 and 3013005724), the Key Scientific and Technological Grant of Zhejiang for Breeding New Agricultural Varieties (2021C02066-5, 2021C02067-7), and the Natural Science Foundation of Zhejiang Province (LQ23C150006).
-
The authors declare that they have no conflict of interest. Pei Xu is the Editorial Board member of Vegetable Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
# Authors contributed equally: Rujia Jiang, Ting Sun
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jiang R, Sun T, Shi Z, Moshelion M, Xu P. 2024. Leveraging 'golden-hour' WUE for developing superior vegetable varieties with optimal water-saving and growth traits. Vegetable Research 4: e002 doi: 10.48130/vegres-0024-0001 

Leveraging 'golden-hour' WUE for developing superior vegetable varieties with optimal water-saving and growth traits
- Received: 31 October 2023
- Revised: 26 December 2023
- Accepted: 03 January 2024
- Published online: 17 January 2024
Abstract: Creating high-yielding and water-efficient crop varieties relies on a profound understanding of crop water usage and photosynthetic physiology. Currently, the prevailing strategies for improving drought response in crops center around the regulation of stomata. However, while reducing stomatal conductance can boost water use efficiency (WUE), it results in a decline in photosynthetic assimilation capacity, because stomata function as a shared conduit for both CO2 intake and water evaporation. With the advancement of phenomics, recent research has unveiled disparities in the regulatory patterns of photosynthesis and transpiration (Tr) in plants. Depending on the genotype, the early morning period, when light intensity is sufficient yet vapor pressure deficit (VPD) is low, is referred to as the 'golden hours' for high water use efficiency (WUE). During this window, plants can attain higher photosynthetic intensity with lower Tr levels in a low VPD environment. This is highly advantageous for efficient biomass production under water-saving conditions. Thus, precise and more judicious modulation of WUE through stomatal control becomes pivotal in addressing the delicate balance between water conservation and yield. This perspective paper introduces the concept and significance of the golden-hour WUE (GHW) trait and elucidates the methods for quantitative and high-throughput screening of this trait using modern phenotyping techniques. Building upon this foundation, a systematic approach for screening and leveraging the GHW traits in plant breeding is proposed. This proposed approach holds the potential to offer a solution for achieving a balance between water-saving and plant growth.
-
Key words:
- Golden hour /
- WUE /
- Balance /
- High-throughput screening /
- Breeding