








-
The persimmon industry has entered a period of rapid development in China owing to the success of large-scale cultivation and advances in scientific research. Persimmons belong to Diospyros, Eb-enaceae, with a chromosome number of 2n = 2x = 30, 2n = 4x = 60, 2n = 6x = 90, or 2n = 9x = 135, and are perennial woody plants[1]. Persimmons not only contain lots of sugar and a variety of vitamins, but also has high medicinal and economic value and is widely used in medicine, health care, cosmetics, and other fields[2−5]. Furukawa et al. found that persimmon tannin could alleviate SARS-CoV-2 virus infection and viral transmission[6]. Itamura et al. have shown that persimmons could dispel the effects of alcohol, and thus reduce the effects of hangovers. The effect of persimmon tannin on the adsorption and recovery of heavy metal ions has also attracted extensive attention[7]. Persimmon tannin can be used as an auxiliary agent of biodegradable materials, high polymer water absorption resin, and other materials in modified polyacrylic acid systems[8].
According to the latest statistics released by the Food and Agriculture Organization of the United Nations (FAOSTAT), Asia has the largest production share of persimmons (Fig. 1a), and China occupies the first position in the world in terms of planted area and annual production of persimmons as of 2020 (Fig. 1b). China is one of the main countries from which persimmons originated, and it is also the country with the longest cultivation history. The production quantities of persimmons in China have been increasing yearly, with an average growth rate of 4.8% from 1994 to 2020 (Fig. 1c). The persimmon production areas in China are mainly in Guangxi, Henan, Hebei, and Shaanxi Provinces (Fig. 1d).
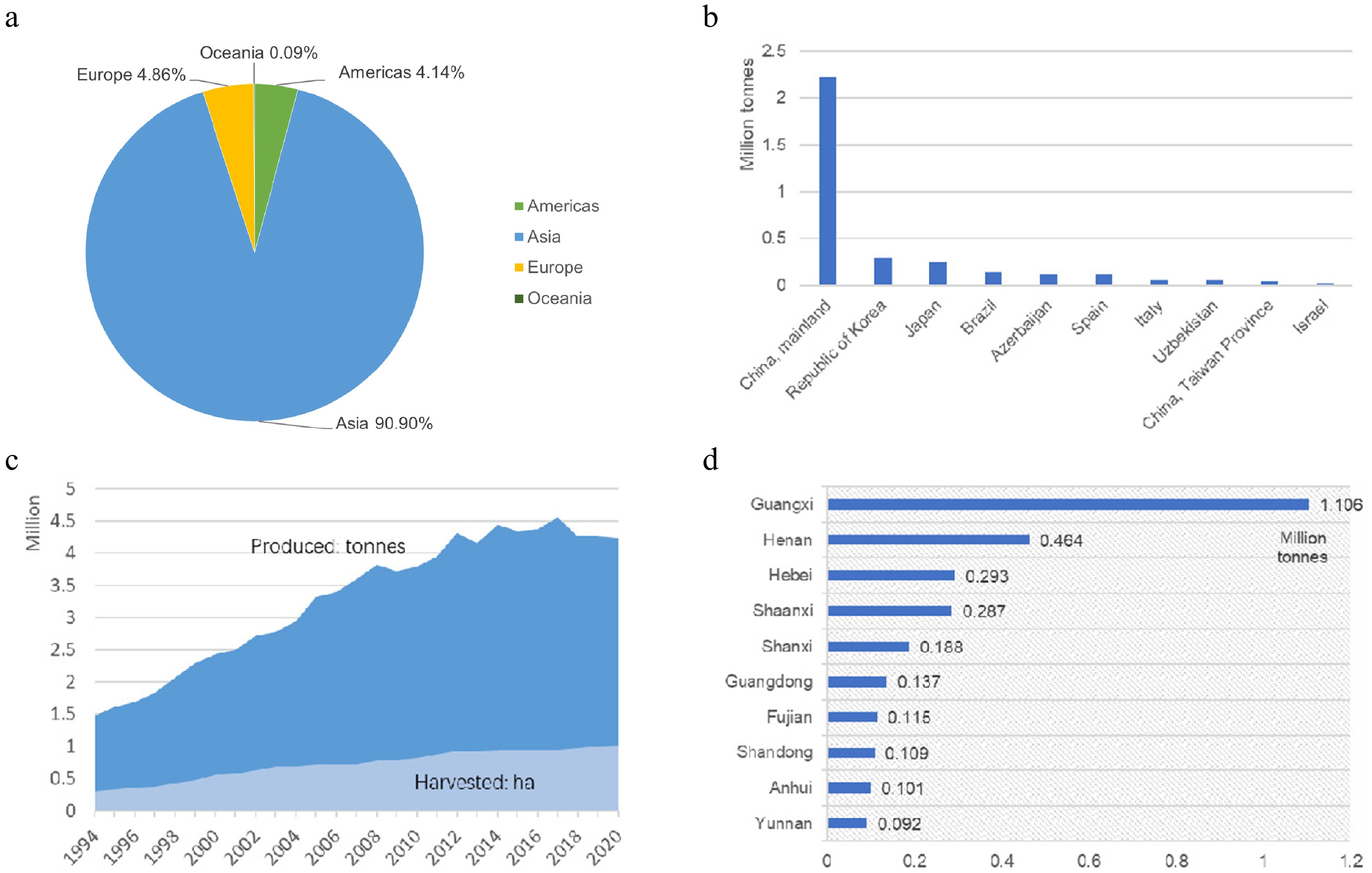
Figure 1.
Current status of the persimmon industry in the world and China. (a) Production share of persimmons by region (average 1994–2020). (b) Top 10 producers of persimmons (average 1994–2020). (c) Production quantities of persimmons in China (1994–2020). (d) Top 10 regions of persimmon production in China.
Although great progress has been made in the research on persimmon cultivation, there are still many significant challenges to the sustainable development of the persimmon industry in China. Pollination-constant nonastringent (PCNA) persimmon is a key target for industry development and variety selection worldwide, because it has the advantages of natural deastringency, great sweetness, and direct edibility[9]. China has the largest cultivated area and annual production of persimmon trees in the world. The main varieties of sweet persimmons currently available in China were introduced from Japan. The natural deastringency of the Japanese sweet persimmon is controlled by recessive genes[10]. In PCNA persimmon breeding, extensive crosses between Japanese PCNA (JPCNA) persimmon cultivars have been performed; however, inbreeding repression has become an issue in JPCNA persimmon breeding owing to high genetic similarity. PCNA persimmon breeding has also been conducted in China using both JPCNA and Chinese PCNA (CPCNA) cultivars, and improved varieties are expected to be produced in the near future. Therefore, the breeding of fine varieties of PCNA persimmon in China is still a great challenge[11]. For astringent persimmons, China is currently promoting CO2 (vacuum packaging) and ethanol treatment and other large-scale techniques for the removal of astringency[12], but artificial deastringency treatment is costly in terms of labor, material, and financial resources. Incomplete removal of astringency in persimmons not only reduces their merchantability but also induces the risk of persimmon bezoars. Another key concern is the screening of rootstocks with wide compatibility[13]. Japan developed the earliest sweet persimmons in the world and promoted their rapid genetic improvement. At present, the main varieties of sweet persimmons all over the world come from Japan, but the soil adaptability of Japanese sweet persimmon is usually poor. For instance, some high-quality varieties, such as Diospyros kaki 'Fuyu' and 'Taishuu', have poor compatibility with the rootstock Diospyros lotus (D. lotus) used in traditional production areas in China. Moreover, the increasingly serious symptoms of delayed incompatibility have become a new problem faced by the persimmon industry[14, 15].
This study summarizes the current status and development of persimmons in China. The current progress in the removal of astringency in persimmons, screening of rootstocks with wide compatibility, and the breeding process of PCNA persimmon is highlighted. This review will facilitate the improvement of persimmon breeding and help promote the sustainable development of the persimmon industry in China. Figure 2 shows the roadmap for persimmon improvement in China.
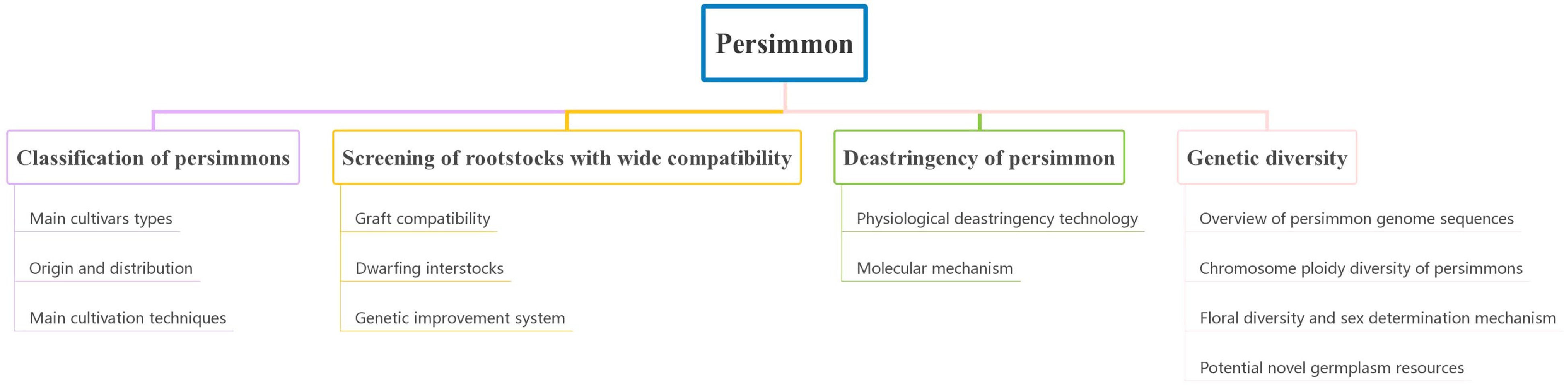
Figure 2.
Roadmap for persimmon improvement in China.
-
Persimmon varieties such as original persimmons (astringent) and sweet persimmons are rich in nutrients. Figure 3 shows the main cultivars of persimmons in China, in which 'Luo-Tian-Tian-Shi' is the earliest and widely distributed PCNA persimmon found in China.

Figure 3.
Main cultivars of persimmons in China.
Chinese persimmons have a cultivation history of more than 2,000 years[16]. As one of the representative cultivars of fruit trees, persimmons can be divided into PCNA persimmons and non-PCNA persimmons, based on the relationship between fruit deastringence and volatile compounds produced by the seeds[17]. Non-PCNA persimmons can be subdivided into pollination-variant nonastringent (PVNA), pollination-variant astringent (PVA), and pollination-constant astringent (PCA) persimmons (Fig. 4).
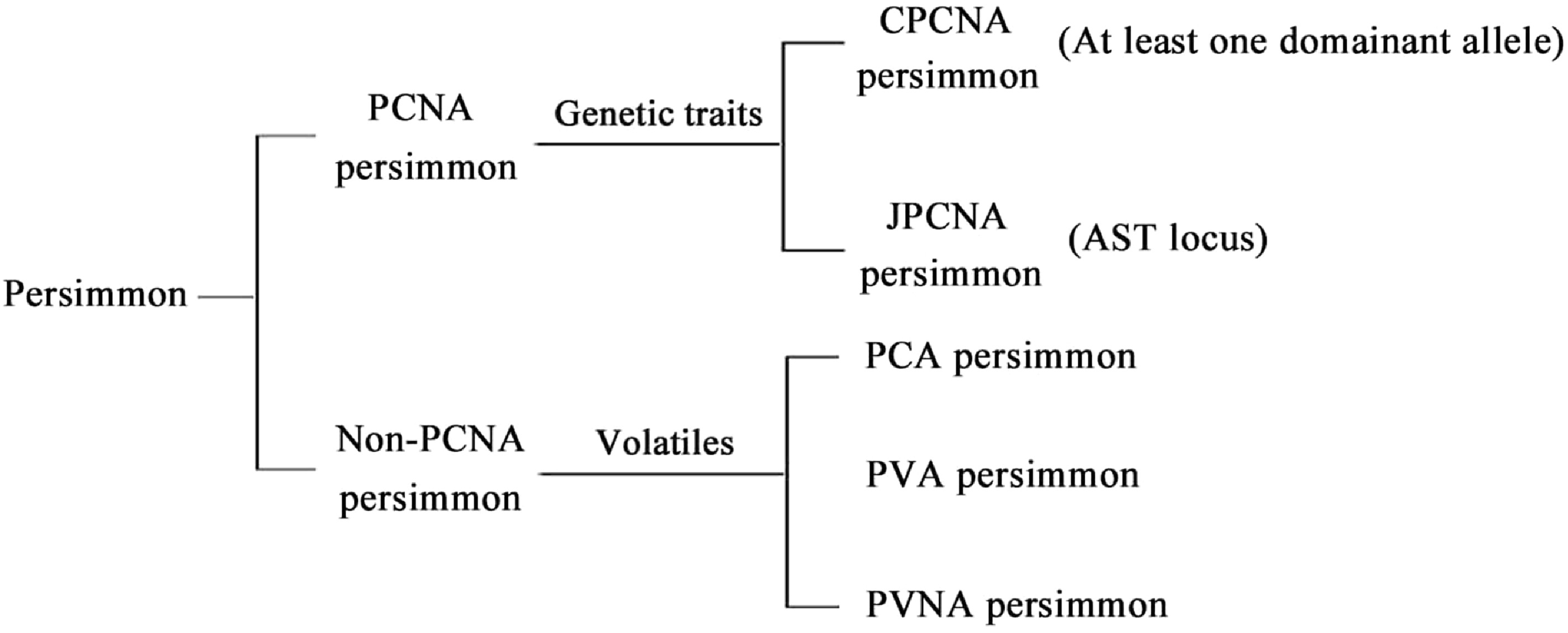
Figure 4.
Classification of persimmons. PCNA: pollination-constant nonastringent. CPCNA: Chinese-PCNA. JPCNA: Japanese-PCNA. PVNA: pollination variant nonastringent. PVA: pollination-variant astringent. PCA: pollination-constant astringent.
PCNA persimmons are believed to be a direct mutation of PCA persimmons, and the Japanese sweet persimmon has specific origin and high genetic consistency. A recent study has shown that the natural astringency reduction characteristic and genotype of the Chinese-origin PCNA persimmon (referred to as the Chinese sweet persimmon), distributed in the Dabie Mountain Area, are different from the Japanese sweet persimmon, whose inbreeding degradation of fruit weight, yield, and tree potential has been evident since a long time[18]. As indicated previously, the natural deastringency of the Japanese sweet persimmon is controlled by recessive genes, while that of the Chinese sweet persimmon is controlled by dominant genes[10]. Therefore, the Chinese sweet persimmon with a relatively independent origin has great application value in the genetic improvement of PCNA persimmons. The vast majority of persimmon varieties in traditional production areas in China are PCA persimmons, and PCNA persimmons are crispy without artificial deastringency and peeling. PCNA persimmons have changed the traditional eating habits and is becoming a popular fruit tree worldwide.
Origin and geographical distribution of sweet persimmons in China
-
Chinese sweet persimmons are distributed in the Dabie Mountain Area at the boundary of Hubei, Henan, and Anhui Province, among which, Luotian county of Hubei Province has the largest area of distribution and the most abundant varieties. In terms of vertical distribution, Chinese sweet persimmons are distributed between 100 and 700 m above sea level, and most of them are in the range of 300−500 m. Few persimmon plants are cultivated above 700 m[19].
According to the "Luotian County Annals", Chinese sweet persimmons were planted in 1032 A.D. (Northern Song Dynasty), more than 180 years earlier than Japan's oldest sweet persimmon variety 'ChanSiWan' (PVNA, discovered in 1214 A.D.). Kanzaki et al.[20] used amplified fragment length polymorphism (AFLP) analysis to obtain similar conclusions. Using sequence-related amplified polymorphism (SRAP), inter-retrotransposon amplified polymorphism (IRAP), and retrotransposon-microsatellite amplified polymorphism (REMAP) techniques to analyze the relationship among Diospyros plants such as CPCNA persimmons and JPCNA persimmons, it was found that the Chinese sweet persimmon and Japanese sweet persimmon are relatively distantly related, but PCNA persimmons and PVNA persimmons could not be separated completely during clustering. Therefore, the original germplasm of Chinese sweet persimmons may be derived from the direct mutation of PVNA persimmon types.
Main cultivation techniques of persimmons
-
The Chinese persimmon usually prefers deep and fertile sandy loam or loam with strong storage capacity, and the slope of the cultivation area should be below 15°. To facilitate soil and water conservation, afforestation land should be thoroughly turned over first. Strong seedlings are used for planting and the soil is loosened 3 to 4 times a year. To expand the canopy, the tree body is trimmed and pruned combined with pruning to control the number of flowers and fruits, and moderate tree vigor is maintained. To improve the serious fruit drop of young persimmon trees, 2 to 3 flower buds are left in the upper and middle parts of each fruiting branch, and all the rest are thinned out, so that the fruits of the whole tree are evenly distributed. The most serious diseases that harm sweet persimmons are persimmon angular spot disease and persimmon anthracnose; therefore, measures should be taken to prevent and comprehensively control them. Firstly, timely removal of diseased leaves and dead branches is required. Secondly, reclamation and fertilization are required during winter, and persimmon tree trunks should be whitened with lime water.
-
The graft compatibility of the rootstock D. lotus used in Chinese traditional production areas is always poor for grafting many super JPCNA cultivars and has become a big problem faced by the persimmon industry. For instance, some high-quality varieties, such as Diospyros kaki 'Fuyu' and 'Taishuu' have poor compatibility with the rootstock D. lotus used in traditional production areas in China. The mechanism of graft incompatibility is still not clear and requires further study. 'Taishuu' is a crossbreed between 'Fuyu' and 'IIiG-16'. However, 'Taishuu' and 'Fuyu' showed no graft compatibility with the commonly used rootstock D. lotus, severely restricting the large-scale production of sweet persimmons. Fortunately, some progress has been made in the research on graft compatibility of rootstocks in China with 'Fuyu' and 'Taishuu' series. Hu et al.[21] showed that the grafting affinity of 'Xiaoguo-Tianshi' and 'Niuyanshi' with 'Fuyu' and 'Jirou' was good through phylogenetic analysis, in vitro culture, and phenotypic observation of field grafting. Therefore, 'Xiaoguo-Tianshi' and 'Niuyanshi' were expected to become the rootstocks for PCNA persimmons. Although sweet persimmons are cultivated and promoted, a rootstock with wide compatibility has become a key issue in the development of sweet persimmons. When the scion is not compatible with the rootstock, the interstock is often used for secondary grafting to improve the graft compatibility of rootstock. The interstock acts as a bridge and regulates the distribution of nutrients and hormones between the scion and the rootstock, thus enabling morphological and physiological effects on the rootstock and the scion[22,23]. Tang et al. found that various interstocks such as the 'Nan-Tong-Xiao-Fang-Shi' and 'Diospyros kaki Thunb. Nishimurawase' could improve graft healing and promote the graft compatibility of 'Diospyros kaki var. sylvestris Mak.,' 'Diospyros galucifolia Metc.,' and 'Diospyros oleifera Cheng' with 'Fuyu'[24]. Wu et al. demonstrated that 'Dabieshan 1' and 'Gaoyou 5' showed good graft compatibility with the scion variety 'Taishuu'[25].
A novel dwarf interstock in China: Nan-Tong-Xiao-Fang-Shi
-
At present, researches are mainly focused on deastringent persimmons and post-harvest softening preservation[26−29], and on obtaining excellent dwarf rootstocks[30]. Persimmon is one of the tallest fruit tree species cultivated in China, and its management is labor intensive. High-density dwarf planting is a labor-saving and efficient cultivation model for woody fruit trees. The vigorous stock, D. lotus, is widely used in persimmon production owing to the lack of suitable dwarfing interstocks. Problems such as long juvenile phase, labor and time consumption, and fruit harvesting difficulties seriously restrict the development of the persimmon industry[13]. 'Nan-Tong-Xiao-Fang-Shi' is a dwarf persimmon variety unique to China and is usually used as an interstock because it does not produce seeds. 'Nan-Tong-Xiao-Fang-Shi' is regarded as a promising dwarfing interstock resource with broad application prospects owing to its excellent dwarfing effect on the scion. Shen et al.[31] explored the dwarfing mechanism of 'Nan-Tong-Xiao-Fang-Shi' as interstocks, and showed that endogenous DkGA2ox1 mRNA which has a high expression level in the interstock of 'Nan-Tong-Xiao-Fang-Shi' was transferred to the scion as a long-distance signal, promoting the accumulation of GA2ox gene in the scion, ultimately reducing the level of active GA, causing scion dwarfing (Fig. 5).

Figure 5.
Schematic diagram of 'Nan-Tong-Xiao-Fang-Shi' as interstocks. (a) Hypothetical model showing DKGA2ox1 transport in grafted plants (Kanshu/ Nan-Tong-Xiao-Fang-Shi/ D. lotus). (b) The diagram of D. kaki 'Youhou'/ Nan-Tong-Xiao-Fang-Shi/ D. lotus grafting.
Genetic improvement system
-
To maintain good agronomic characters, cuttings of root tillers or budding tillers could be selected or propagation methods such as in vitro rapid propagation could be adopted. In addition to the compatibility of rootstock and spike, selection of rootstocks should be based on characteristics such as easy reproduction, high yield, high quality, and strong soil adaptability. The soil condition of persimmon production areas in China is generally inferior to that in Japan; therefore, it is difficult to solve the problem of graft compatibility through the introduction of rootstocks. Li et al.[15] established a protocol for regeneration and Agrobacterium tumefaciens mediated genetic transformation of D. lotus in vitro, and the results showed that the callus induction and regeneration rate reached 100% and 94.28%, respectively, by optimizing the preculture time, bacterial optical density, infection time, co-culture time, and acetyl ethyl ester concentration. Efficient genetic improvement of sweet persimmons was established by integrating traditional hybridization with modern biotechnology and marker-assisted selection (Fig. 6). In addition to increasing heterosis and shortening the breeding cycle, seven out of eight non-target individuals could be eliminated in the early stage by this system[14].
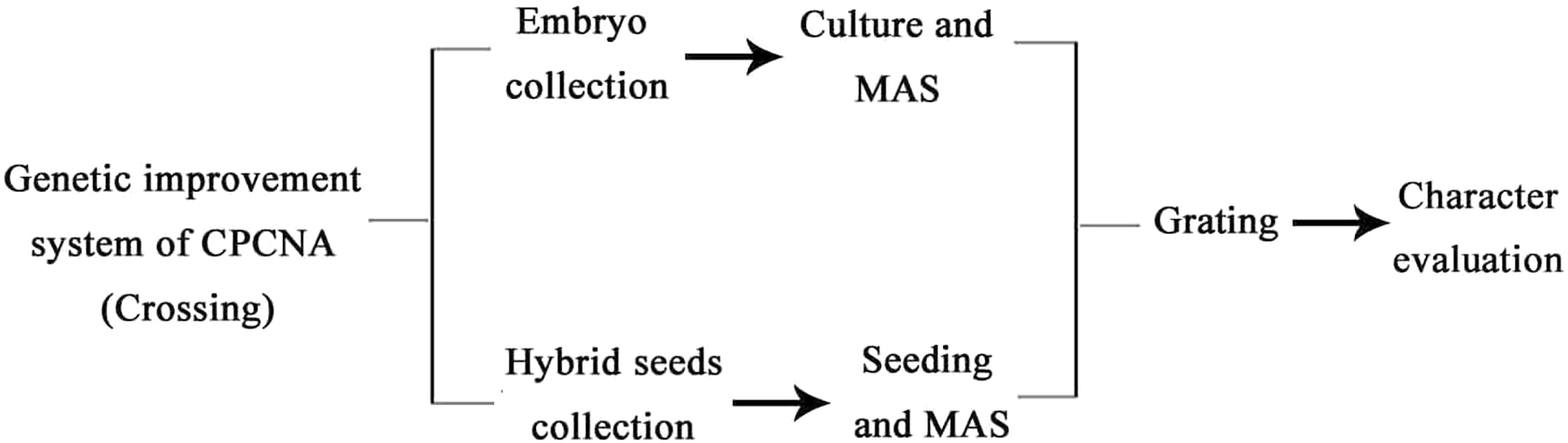
Figure 6.
Diagram of genetic improvement of CPCNA persimmon. MAS: marker-assisted selection.
-
PCA persimmon fruit is still the main product in traditional production areas China. The incomplete removal of astringency in persimmon fruits not only reduces its commercial value, but also has the risk of inducing persimmon bezoars. Therefore, to improve the comprehensive benefits of the industry, it is necessary to carry out research on the technology of deastringency. Astringent persimmons should not be eaten immediately after being picked but only after deastringent treatment. The commonly used deastringent methods for crispy persimmons include warm water treatment, lime water immersion, and CO2 treatment. Most of the soluble tannins are converted to insoluble tannins, and the fruit becomes deastringent[11, 27, 32]. Recently, a new study showed that microwave treatment can significantly inhibit the expression of degradation genes in cells; inhibit the degradation of cellulose, pectin, and the activity of cellulase; and improve the quality of persimmons, indicating it is a promising method for persimmon preservation[12].
The natural deastringency of persimmons is more favorable in the persimmon industry; however, its mechanism is still not fully understood. A study on the natural deastringency characteristics of Chinese sweet persimmon showed that[11]: (1) the tannin biosynthesis of sweet persimmons was reduced five weeks after flowering; soluble tannin content was reduced to a degree that it could not be felt by the taste organs, however, it did not decrease to an edible level until fruit ripening (15 weeks after flowering); (2) Ethanol treatment could induce the deastringency of PVNA persimmons at young fruit stage (early July), and the deastringency of Chinese sweet persimmon could be completed in vivo or in vitro in early July or early September; (3) Majority of 'Luo-Tian-Tian-Shi' contained a high proportion of solidified tannin cells, and the molecular weight of tannins, mainly catechin and gallic acid, was higher. Chinese sweet persimmons and Japanese sweet persimmons have similar characteristics of small tannin cells during early fruit development and natural deastringency in the mature fruit, but the natural deastringency of Chinese sweet persimmon is completed before the fruit is mature. The natural deastringency of Chinese sweet persimmons may be more directly related to the non-dissolution of soluble tannins mediated by acetaldehyde (also known as "solidification effect") before fruit ripening. However, the mechanism of natural deastringency of Chinese sweet persimmons is still not clear.
Recently, Chen et al.[9] found a transcription factor DkMYB14 with dual functions of transcriptional activation and inhibition, which can inhibit the biosynthesis of tannins (procyanidins) as a transcription inhibitor and promote the biosynthesis of acetaldehyde as a transcriptional activator. Thus, the transformation of soluble tannins to insoluble tannins was promoted (Fig. 7). Min et al.[33] indicated that DKNAC 1/3/5/6 might be an important candidate regulator of astringency removal in persimmons. Jamil et al.[34] revealed the regulatory mechanism of zinc finger proteins on the deastringency of persimmons under the action of AHCA. Luo et al.[35] and Yang et al.[36] showed that miR395p-3p, miR858b, miR156j-5p, miR396g, and miR2911a may play key roles in fruit deastringency of persimmons under normal development conditions through miRNA sequencing. The genetic improvement of persimmons is more difficult than that of other fruit trees; therefore, the discovery of new gene resources and improvement of breeding techniques are the key to make a breakthrough in the breeding of sweet persimmons worldwide.
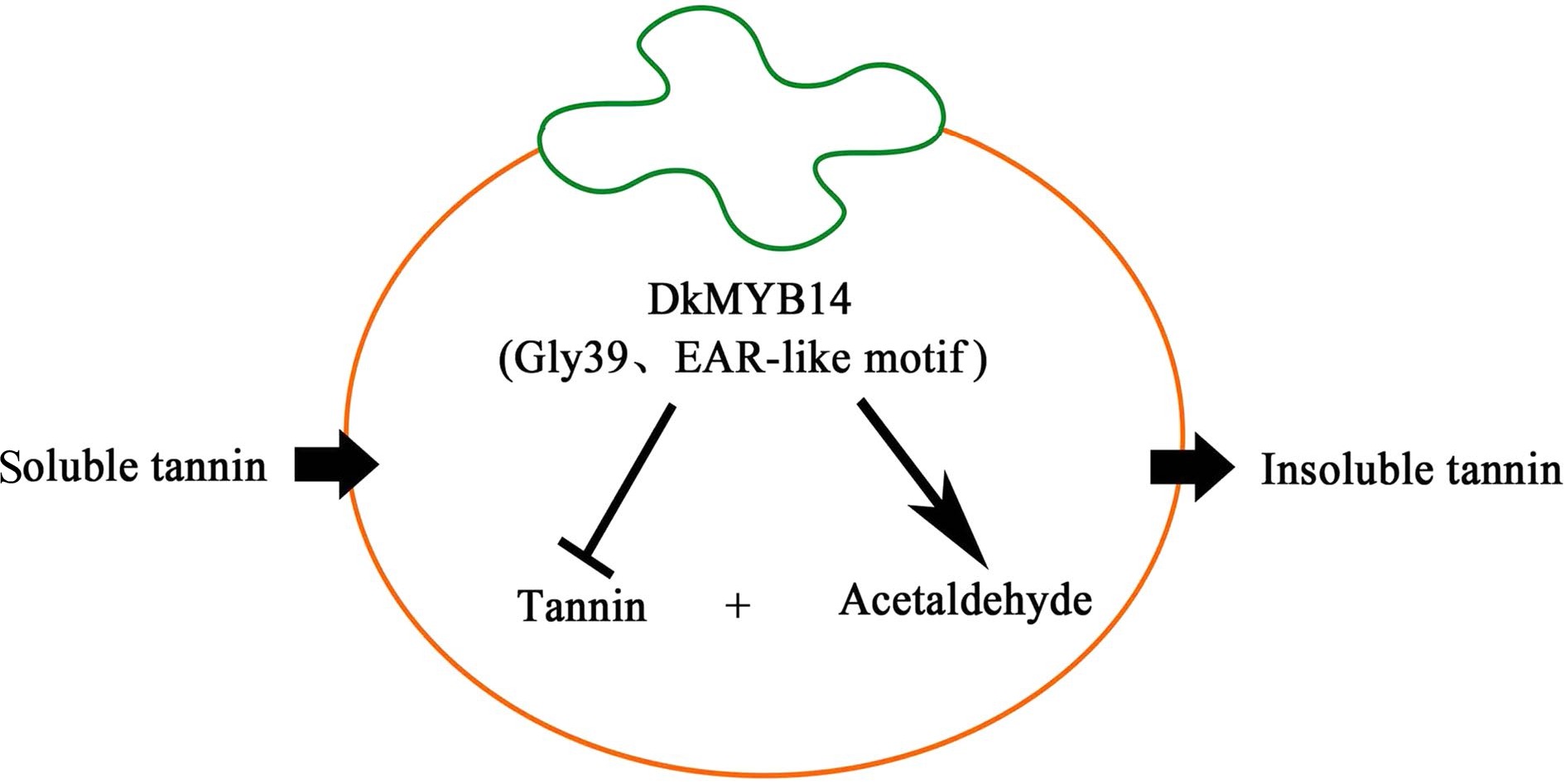
Figure 7.
Hypothetical model showing how DkMYB14 regulates the accumulation of proanthocyanidin (PA) in PCNA persimmon and non-PCNA persimmon.
-
The high ploidy levels and the resulting genetic/genomic complexity often interferes with various genome-wide analyses. Yonemori et al.[37] determined the genomic structure and evolution of D. Lotus (2n = 6x = 90); draft genomes with expected haploid genome size (877.7 Mb from KMER analysis) for long read coverage were collected from CA 65X PacBio using Falcon technology, and it resulted in 3,073 primary contigs and 5,901 "secondary" contigs, which were considered to be alleles of the primary contigs. Thereafter, three genetic profiles from two isolated F1 populations (N = 314 and 119) were created. Genetic maps were created from a total of 5,959 markers from GBS/ddRAD sequencing and allowed contigs to be anchored into a genome sketch containing 15 pseudomolecules that covered approximately 70% of the entire sequence. Transcriptome data were mapped to major genomic contigs, and 40,532 predictive genes were obtained, including 12,058 single-copy number or low-copy number genes (Genome Database of Rosaceae: GDR,
http://persimmon.kazusa.or.jp/ ).Zhu et al.[1] obtained a chromosome scale genome assembly of the diploid persimmon variety 'Yoshi' (Diospyros Oleifera Cheng, 2n = 2x = 30) using Illumina sequencing, single-molecule real-time sequencing, and high-throughput chromosomal conformation capture. The assembled D. oleifera genome consisted of 849.53 Mb, of which 94.14% (799.71 Mb) was allocated to 15 pseudochromosomes and was the first assembled genome in Eb-enaceae. Comparative genome analysis showed that the genome of M. oleracea underwent an ancient γ-genome-wide replication event, which provided a basis for improving persimmon breeding (Genome Database of Rosaceae: GDR,
www.kakiwi.zju.edu.cn ).Chromosome ploidy diversity of persimmons
-
Persimmons have a long history of being cultivated in China, but the genetic diversity among persimmon species and within persimmon species is still unknown. Guan et al.[38] provided the transcriptome resources of related persimmon species for the first time, revealing the genetic differentiation between species. SCoT and IRAP markers were used to investigate the genetic diversity and relationships of 268 persimmon varieties from different regions of China[39]. The results showed that there was gene exchange between persimmons from different regions, which is of great significance for efficient conservation, identification, and utilization of persimmon germplasm resources. Itamura et al.[7] found that the proportions of PCNA persimmon and PVNA persimmon in F1 generation were 50%, respectively, indicating that the natural astringency trait of 'Luo-Tian-Tian-Shi' was a dominant trait. Guo et al.[40] and Du et al.[41] discovered a series of completely male persimmon plants in the Dabie Mountain Area, which were closely related to the Chinese sweet persimmon. Some of the completely male persimmon plants carrying CPCNA persimmon gene, which controls the natural astringency of Chinese sweet persimmon, could be used as the father for PCNA persimmon breeding. It is also a suitable material for the study of flower sex differentiation and is a valuable germplasm resource.
Persimmons are mostly diploid (2n = 2x = 30) and are distributed in tropical or subtropical areas. However, during its migration to northern subtropics and the temperate zone, its chromosomes doubled to produce the cultivated persimmon. Although the details are not fully understood, the evolution process is likely to involve 2n gametes[42]. After the zygote embryos were formed by the 2n pollen pathway, due to the abnormal endosperm balance number, rescue culture of young embryos was necessary to obtain hybrid progenies. 2n oocytes fertilized with normal pollen produce seeds without the need for embryonic culture. Therefore, the evolution from 2-ploid to 6-ploid or 9-ploid in persimmons may be more likely due to 2n egg cells[43, 44]. These results not only provide a new scientific basis for the study of origin and evolution of cultivated persimmons, but also lay a technical foundation for the genetic improvement of persimmons.
Floral diversity and sex determination mechanism of persimmons
-
The floral sex mechanism of persimmons is very complicated. At the single flower level, there are three types: male flower, female flower, and hermaphroditic flower[45]; at the individual plant level, there are seven types: androecious plants (only male flowers), gynoecious plants (only female flowers), dioecious (female flowers and male flowers coexist), gynomonoecious plant (female flowers and hermaphroditic coexist), andromonoecious plants (male flowers and hermaphroditic coexist), androgynomonoecious plants (male flowers, female flowers, and hermaphroditic flowers coexist), and only hermaphroditic plants[46]. Akagi et al.[47] reported that a gene on the Y chromosome regulated a small RNA linked to a non-sex chromosome. The small RNA is localized in the male flower and could inhibit the development of female organs. Monoecious varieties require the Y-encoded small RNA (smRNA) gene OGI. The parthenosexuality of individuals in the dioecious system is often related to genetic determinants on the sex chromosomes. With regard to dioecy, only a few genetic determinants have been identified in some lineages, including persimmons. In dioecious persimmon species, a Y chromosome encoded small RNA (smRNA) gene OGI, which is considered to be the only sex determinant, is responsible for inhibiting the expression of the autosomal counterpart MeGI. In subsequent studies[48, 49], the mechanisms of diploid persimmon and hexaploid persimmon were studied with respect to epigenetic regulation, and another layer of regulation in the form of DNA methylation of the MeGI promoter was found to be associated with the production of male and female flowers in genetically male trees (Fig. 8). The evolutionary history of these genes suggests that they are tied to the origin of separation of sexes in the persimmon family.
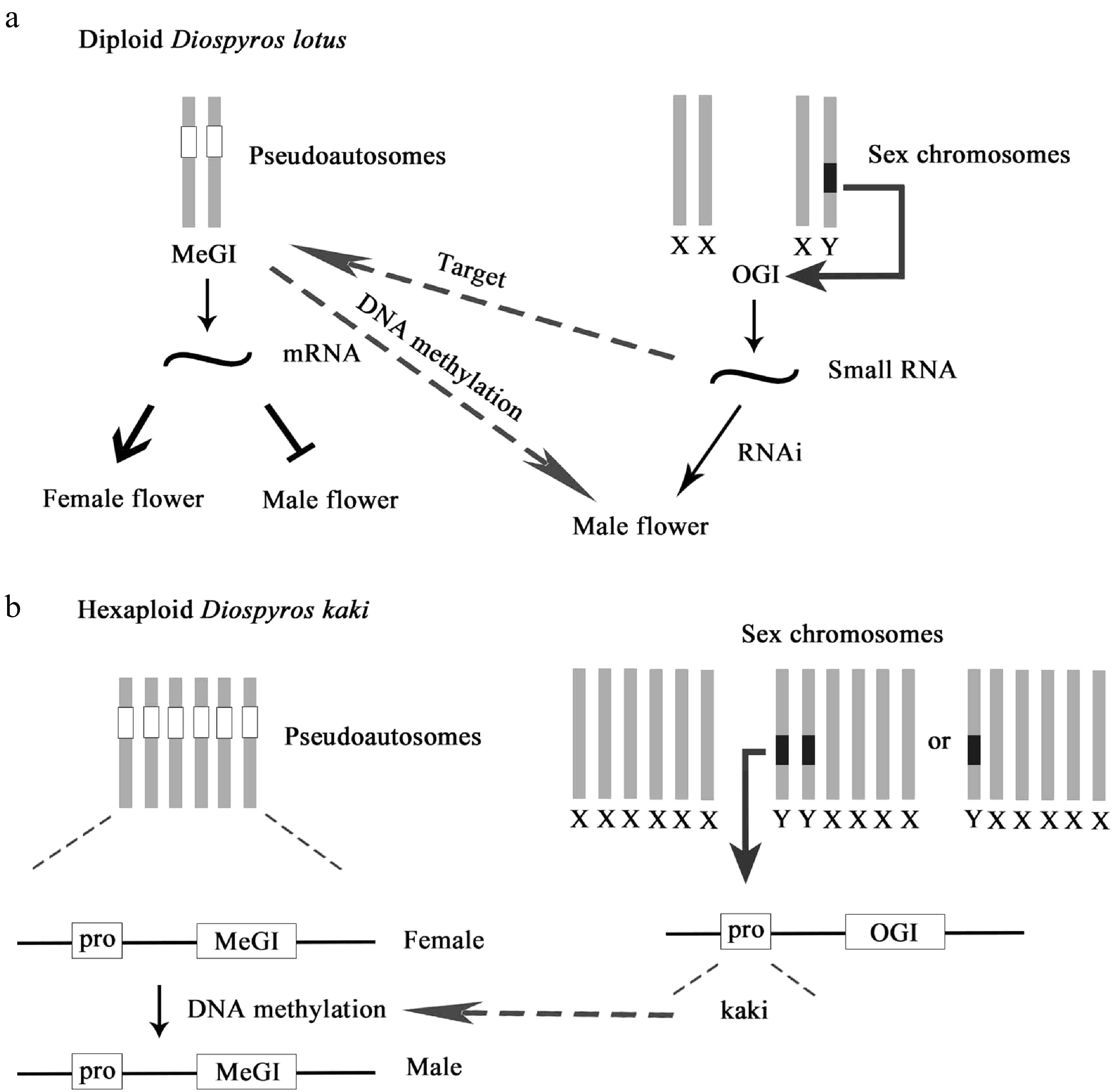
Figure 8.
Diagram of sex evolution model of persimmon. (a) Genetic sex determination in diploid D. lotus. (b) Epigenetic sex determination in genetic males of D. kaki.
The Chinese completely male germplasm has the following characteristics[50−52]: (1) stable male flower character, large pollen quantity, and high pollen germination rate; (2) strong affinity for pollination with the main persimmon variety that enough F1 generations could be obtained through a relatively small number of crosses to provide enough individuals for character screening. Therefore, the completely male persimmon germplasm originating from China has the potential to be developed as a special pollination variety and can be the parent of PCNA persimmon for breeding. In addition, a previous report indicates that hormones are correlated with flower bud differentiation, high GA3 content is positively correlated with male flower bud formation, and high level of ZT and ABA may promote female floral bud differentiation of persimmons[53]. By the utilization of the genotypic pattern of male strains and some F1 generation methods, the sexual orientation of Diospyros kaki was identified in the early stage[46, 54, 55]. The results showed that OGI is an indispensable low-expression gene in male strains. Kali located in the promoter region of OGI showed a high methylation level. This finding will contribute to the improvement of Diospyros kaki's sex theory model and the cultivation of excellent male germplasm resources, which could provide good value for crossbreeding work. PCNA persimmons are believed to be a direct mutation of PCA persimmons, and these mutant individuals have gradually formed the present population through repeated natural crosses with other varieties, and the Chinese and Japanese sweet persimmons are of independent origin. Most persimmon cultivars bear only female flowers, and the androecious plants native to the Dabie Mountain Area are a rare unique germplasm in the world. The natural deastringency traits are controlled by dominant genes, which have the potential to develop special pollinated varieties. It was reported that a series of androecious germplasm in the Dabie Mountain Area were identified, and a kinship analysis based on molecular markers suggested that it may be derived from a variant of the Chinese sweet persimmon. According to a new study[56], in Luojiagang, Jiangxi Province (N 28.53°, E 114.76°), a new persimmon variety was found with only male flowers (PCNA), and was found to carry the sex-linked marker DLSX-AF4S. This above germplasm has potential as a persimmon breeding resource. It is significant to preserve the genetic diversity of persimmon cultivars by protecting the male flower resources of persimmons extensively.
Potential novel germplasm resources in China
-
The main way to create new persimmon germplasm is cross-breeding, and the key is parental selection of cross-breeding, shortening of breeding years, and improvement of breeding efficiency. Genetic improvement is a prerequisite and foundation for the sustainable development of the persimmon industry. In addition, the difficulties in finding excellent rootstock of sweet persimmon varieties in China have not yet been solved. It was reported that a genetic improvement system for PCNA persimmon was established relying on gene editing, molecular marker-assisted breeding, and epigenetic studies, all of which can provide new ideas for persimmon genetic breeding research[55, 56]. 'Heishi No. 1' is a new black, early-maturing and high-quality persimmon variety selected from wild persimmon germplasm resources in Taihang Mountain[57] (Fig. 9a). 'Zhongshi 2' is a new variety with good and stable yield with cold, drought, and disease resistance[58] (Fig. 9b). Zhang et al.[51] crossed the androecious genotype 'Male 8' with non-PCNA persimmon, and the F1 generation obtained was a PCNA persimmon with natural deastringence characteristics, which is of scientific importance for the genetic improvement of Chinese sweet persimmon (Fig. 9c). After years of experimental research, 'Yalin 6' was selected as a widely compatible rootstock and is significant in overcoming the technical difficulties of screening rootstocks with wide compatibility in China, making high-quality sweet persimmon varieties available for production[59] (Fig. 9d). Through the analysis of morphology, chromosome ploidy, pollen morphology, and molecular marker level, it was found that 'De-Yang-Shi' (also named Hong-Hua-Ye-Mao-Shi, tetraploid (2n = 4x = 60)) might be a new germplasm. In particular, the bright color, large number of flowers, and short period of young seedlings are the characteristics that distinguish it from other persimmon materials[60] (Fig. 9e).

Figure 9.
Potential novel germplasm resources in China. (a) A new persimmon cultivar ‘Heishi No. 1’. (b) A new persimmon cultivar ‘Zhongshi 2’. (c) A new persimmon cultivar ‘Male 8’. (d) ‘Yalin 6’ as a potential widely compatible rootstock. (e) A new persimmon cultivar ‘De-Yang-Shi.’ From left to right: ripening fruit with fluff, ripe fruit, male flower, and female flower.
-
The genome of persimmons is highly heterozygous, and the genetic basis of many crucial traits are not yet clear; therefore, targeted and efficient "precision breeding" cannot be fully realized at present. In general, there are many varieties of persimmon that only have female flowers, and it is difficult to choose a suitable male parent. Therefore, conventional hybrid breeding is more difficult than in other fruit trees. This greatly limits the range of selection of cross-breeding parents, and many PCNA persimmon varieties with good traits cannot be fully utilized. Combined with embryo rescue, molecular markers, and high grafting techniques, male flowers could be produced in the second year after grafting and fruit could be borne in the third year, which could shorten the breeding cycle of PCNA persimmon to 7−8 years[61]. At present, the genetic improvement of persimmons is developing rapidly in China, and a relatively complete technical system has been established[15, 62], but further research is still needed.
With respect to persimmon cultivation, China has a superior natural environment, abundant germplasm resources, and large production scale. The market demand for persimmon products, at home and abroad, is growing rapidly. Breeding and cultivation techniques which are suitable for the development of China's persimmon industry continue to improve. However, there are some challenges, such as the occurrence of pests and diseases, single post-harvest processing, and market competition, necessitating China to modify the operation of the persimmon industry. In the future, with the establishment of the persimmon genetic improvement technology system, the persimmon industry in China will develop rapidly.
This study was supported by the Jiangsu Seed Industry Key Research Program (JBGS[2021]019), the Fundamental Research Funds for the Central Universities (KYZZ2022004), the National Key R&D Program of China (2019YFD1000600), the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX21_0606), the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions, and the Jiangsu Belt and Road innovation cooperation project (BZ2019012).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Dong Y, Yu X, Ye X, Gao Z, Wang S, et al. 2022. Current status and perspective of persimmon research in China. Technology in Horticulture 2:4 doi: 10.48130/TIH-2022-0004 

Current status and perspective of persimmon research in China
- Received: 24 February 2022
- Accepted: 30 May 2022
- Published online: 25 July 2022
Abstract: The persimmon, belonging to the genus Diospyros, is one of the most common fruits with high edible and economic value. China has the largest cultivated area and annual production of persimmons in the world. Pollination-constant nonastringent (PCNA) persimmon, which has great advantages of natural deastringency and direct edibility, has been a key target for industry development and variety selection worldwide. However, there are still many challenges in the development of persimmons in China; for example, the ineffective cost of artificial deastringency treatment of persimmons, laborious breeding of fine varieties of PCNA persimmon, and severe symptoms of delayed graft-incompatibility. This review summarizes the current status and development of persimmons in China. The current progress on the removal of astringency in persimmons, screening of rootstocks with wide compatibility, and the breeding process of PCNA persimmon is highlighted. This review will help promote the breeding of excellent persimmon varieties and the sustainable development of the persimmon industry in China.
-
Key words:
- Sweet persimmon /
- Deastringency /
- Graft compatibility /
- Genetic diversity