








-
Melon (Cucumis melo L., 2n = 24) is a kind of dicotyledonous diploid plant of the family Cucurbitaceae, and represents a significant horticultural crop on a global scale[1]. Due to the high commercial and nutritional value, melon is extensively cultivated across temperate, subtropical, and tropical regions worldwide. The species is divided into two subspecies, C. melo ssp. Agrestis and C. melo ssp. Melo, based on the presence or absence of ovary hairs during the early fruit development stage[2]. Melons are favored for their pronounced aroma, elevated sugar content, and superior flesh texture. Data from the Food and Agriculture Organization of the United Nations (FAOSTAT,
www.fao.org/faostat ) shows an upward trend in both the production and yield of melons worldwide from 2018 to 2022 (Fig. 1). Melons are rich in vitamins, carbohydrates, proteins, and essential minerals, along with various antioxidants such as phenolic compounds. Furthermore, melons exhibit significant genetic diversity reflected in variations of fruit size, shape, color, and rind texture, among other characteristics[3]. Consequently, the biological breeding of melons exhibits significant plasticity in the development of hybrids with desirable characteristics. Over the past decade, Spain's average annual melon exports have surpassed 400 kilotons. Furthermore, melons are a heat-resistant crop that exhibits wide adaptability and possesses a short growth cycle, making them highly valuable in the agricultural market. The cultivation of melons can yield significant economic benefits for farmers as they represent one of the key crops for income enhancement.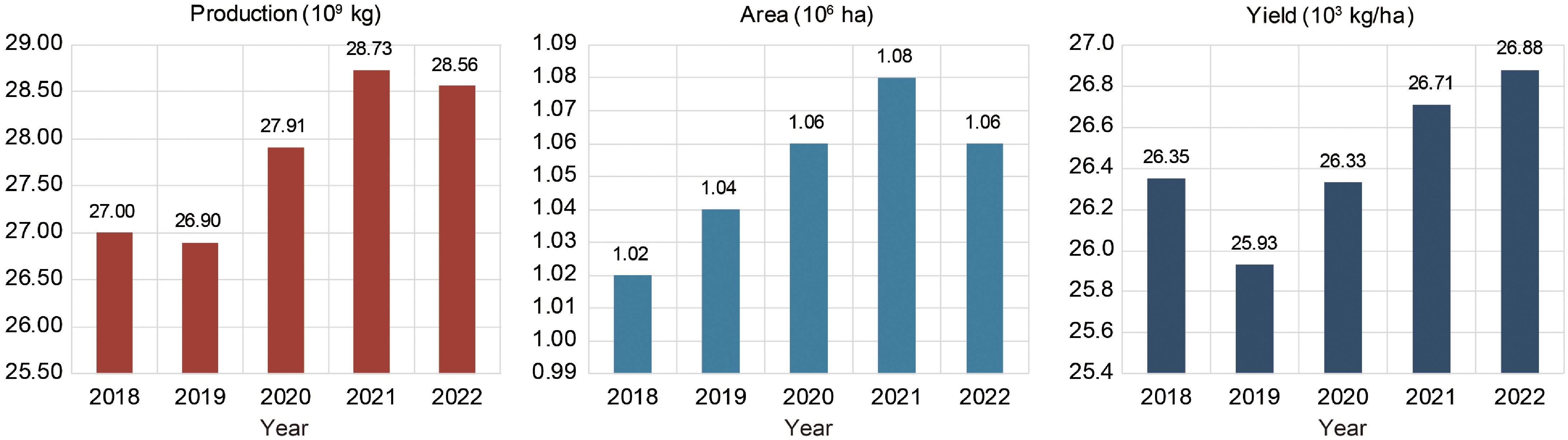
Figure 1.
Global production, cultivated area, and yield of cantaloupes and other melons from 2018 to 2022. Data was collected from the Food and Agriculture Organization of the United Nations (FAO)
(www.fao.org/faostat/zh/#data/QCL/visualize ).The publication of the melon genome has significantly accelerated the research cycle concerning yield and quality traits in melons[1]. Significant advancements have also been achieved in the mapping of QTL and the cloning of genes[4]. Despite significant advancements in genetic enhancement and cultivation methodologies for melons, the challenge of improving both melon quality and yield remains unresolved. In this review, we concentrate on the nutritional and pharmacological properties of melons, while analyzing and summarizing the evolution of melon research from traditional QTL mapping based on linkage maps to contemporary QTL mapping and gene cloning techniques developed through whole-genome resequencing. Furthermore, we emphasize the importance of gene editing and artificial intelligence (AI) technologies, as well as the diversity of germplasms in breeding initiatives. Additionally, we provide recommendations for future strategies and directions in melon breeding, including the integration of QTL associated with desirable traits, identification of disease resistance loci, and optimization of cultivation conditions along with plant architecture. Our insights aim to enhance the understanding of melons and inform future advancements in melon biological breeding.
-
Current melon breeding efforts prioritize yield, disease resistance, aesthetic appeal, and flavor characteristics. However, as a widely consumed food item, melons possess significant nutritional and pharmacological benefits that are frequently overlooked due to their palatable taste. In this context, we present a concise overview of the nutritional and pharmacological properties of melons with the aim of enhancing awareness across various fields.
The nutritional constituents of melon
-
Melons not only possess a delightful sweetness but also offer significant nutritional benefits. With their high-water content, melons provide essential hydration. Additionally, they are abundant in carbohydrates and minerals, making them an ideal choice for consumption. The carbohydrates found in melons primarily consist of starches and sugars, with monosaccharides and disaccharides present in nearly equal proportions. Melons are inherently low in fat content, lacking both monounsaturated and trans fats. The majority of their fatty acids are unsaturated, encompassing beneficial omega-3 and omega-6 fatty acids[5]. Melons are rich in nearly all water-soluble and fat-soluble vitamins, with the exception of vitamins D and E. For instance, honeydew (C. melo ssp. Melo) provides over 18.75 μg/100 g fresh weight of folate, more than 18.0 mg/100 g fresh weight of vitamin C, exceeding 0.42 mg/100 g fresh weight of niacin, and surpassing 2.8 μg/100 g fresh weight of vitamin K[6]. Melon fruits are a valuable source of essential amino acids, providing the body with significant quantities of aspartic acid, glutamic acid, and alanine, while containing lower levels of lysine, arginine, proline, and glycine. Additionally, melons offer modest amounts of protein and dietary fiber that contribute to digestive health[6,7]. These represent numerous advantageous attributes of melons, which, when consumed consistently, can furnish the body with essential nutrition. Melon serves as a significant botanical source of essential minerals for the human body, exhibiting beneficial physiological effects, particularly due to its elevated levels of selenium, calcium, zinc, and potassium, which play a crucial role in supporting immune system functionality[6]. Therefore, melons are highly suitable for consumption, particularly for pregnant women and children. The nutritional value of melons can exhibit considerable variability influenced by factors such as cultivar, ripeness, and cultivation conditions. In light of the current understanding of the nutritional profiles and availability of melons, it is imperative to identify and cultivate varieties with enhanced bioactivity to produce superior products that meet contemporary nutritional and health food standards.
The pharmacological efficacy of melon
-
During metabolic processes in living organisms, a significant quantity of free radicals are generated. These free radicals exhibit high reactivity, and their overproduction can result in damage to macromolecules such as DNA, proteins, and lipids within the organism, ultimately leading to cellular injury and the onset of diseases including cardiovascular and inflammatory disorders[8]. Melon is rich in a variety of natural antioxidants, including flavonoids, phytoene, phenolic acids, 5-methyltetrahydrofolate acid, vitamins, β-carotene, and carotenoids. These compounds function as scavengers of reactive oxygen species within living organisms and are effective in mitigating free radical-induced damage while also reducing the risk of numerous diseases[9,10]. The study also revealed that methanolic extracts from various parts of the melon exhibited distinct phenolic content and antioxidant activity levels, with the pulp extract yielding the highest concentration at 89.6 ± 0.3%. Among these extracts, leaf samples demonstrated the greatest total phenolic content (26.4 ± 0.3 mg GAE/g extract) as well as the highest total flavonoid content (69.7 ± 3.37 μg RE/g extract), thereby showcasing superior antioxidant activity[10]. Melon seeds are abundant in polyphenols and flavonoids. The polyphenolic content of melon seed extract is 37.10 ± 1.11 mg GAE/g of dry extract, while the flavonoid content is measured at 0.43 ± 0.27 mg QE/g of dry extract, and the inhibitory effect of the seed extract on DPPH free radicals can reach up to 69.47%[11]. The investigation into the quantification of phenolic compounds and flavonoids in melon peel revealed that the total content of phenolics and flavonoids was 1.016 mg gallic acid equivalent per 100 g and 262 mg catechin equivalent (CA) per 100 g, respectively. Additionally, it was observed that the aqueous ethanol extract of melon peel exhibited a significant scavenging effect on hydroxyl free radicals, reaching up to 68%[12]. While the antioxidant levels in different melon varieties may vary, their therapeutic potential offers valuable pharmacological insights.
The documented modulatory effects of melon encompass a range of activities, including antioxidant, diuretic, analgesic, hepatoprotective, anti-glycation, anti-hyperlipidemic, and antidiabetic properties, in addition to the inhibition of proliferation in certain cancer cell lines. The diuretic effect of melon is particularly evident in the ether extract derived from melon seeds, which may enhance urine output and urinary chalcogenide levels by reducing renal tubular reabsorption while simultaneously increasing the glomerular filtration rate[13]. The methanolic extract derived from melon seeds exhibits a significant analgesic effect, primarily through the reduction of LTB4 levels in the bloodstream and the attenuation of pro-inflammatory factor release from neutrophils[14]. Dried melon fruit tips have been shown to enhance hepatic gluconeogenesis, exhibit protective effects against CCl4-induced toxicity, and may serve as an adjunct in the management of both acute and chronic hepatitis, as well as hepatic jaundice[15]. Melon exhibits antidiabetic properties, as the abundant ascorbic acid and polyphenols present in melon rind extract can enhance thyroid hormone and insulin levels, thereby potentially ameliorating hyperglycemia and diabetes-related alterations, as well as symptoms associated with hypothyroidism[16,17]. Furthermore, research has demonstrated that melon possesses anticancer properties. Specifically, Cucurbitacin B found in melon can inhibit the Raf/MEK/ERK and STAT3 signaling pathways in the human leukocyte cell line K562, exhibiting anti-leukemic effects[18]. Similar studies have demonstrated that melon peel extract significantly inhibited the proliferation of cancer cells in renal, colorectal, cervical adenocarcinoma, and cervical cancers, with inhibition rates ranging from 20% to 85% at extract concentrations between 0.1 and 1.0 mg/mL[12]. Melon also enhances the human cardiovascular system and mitigates the risk of cardiovascular diseases. Adenosine present in melon extract can inhibit platelet aggregation induced by human adrenaline, thrombin, and collagen, therefore, the consumption of melon may serve as an effective preventive measure against cardiovascular diseases[19,20]. It is crucial to emphasize that the aforementioned medicinal properties are derived from preliminary findings. While melons exhibit certain medicinal potential, further scientific investigation is essential to substantiate their efficacy and safety.
-
Population resequencing based on reference genomes can yield a greater abundance of genetic variation and enable high-throughput genotyping[21]. These variants not only enhance traditional quantitative trait locus (QTL) mapping based on linkage maps, but also facilitate the development of innovative QTL mapping methodologies grounded in resequencing techniques, such as bulked segregant analysis (BSA) and genome-wide association studies (GWAS). These two approaches to QTL mapping are complementary and can be effectively utilized in conjunction with one another, rather than one being unequivocally superior to the other. However, it is evident that advancements in population resequencing technology have significantly accelerated the pace of QTL mapping and gene cloning.
QTL mapping based on linkage map in melon
-
QTL mapping is a sophisticated technique employed to identify genetic loci that correlate genotypes with phenotypes. By constructing a linkage map of melon, researchers can systematically observe and quantify the effects of various genotypes on melon traits, thereby facilitating the identification of QTL associated with specific target traits. Traditional QTL mapping relies on linkage maps derived from recombinant populations[22]. QTL mapping using a linkage map has also yielded excellent results in terms of melon quality and agronomic traits[23]. The recombinant population is established from parental lines exhibiting two extreme phenotypes for a specific trait, encompassing recombinant inbred lines (RIL), near-isogenic lines (NIL), F2, F2:3 generations, and double haploid (DH) lines, all of which exhibit substantial genetic variation among progeny. In addition to the recombinant population, two essential prerequisites for QTL mapping must be met: one involves polymorphic DNA genetic markers such as random amplified polymorphic DNA (RAPD), simple sequence repeats (SSR), insertion-deletion polymorphisms (InDel), and single nucleotide polymorphism (SNP) that delineate the types of variation present in the progeny. The other prerequisite pertains to the phenotypic characterization corresponding to each individual within the recombinant population (Fig. 2a). The initial QTL mapping of melon primarily relied on the characteristics of parental lines. For instance, Díaz et al. identified two QTLs linked to fruit diameter and five associated with fruit length within an F2 population[24]. Amanullah et al. utilized an F2:3 population and 160 CAPS markers distributed across 12 chromosomes to construct a comprehensive genetic linkage map spanning 2,848.68 cM, and identified 24 QTLs associated with fruit morphology and quality, located at various chromosomal positions[25].
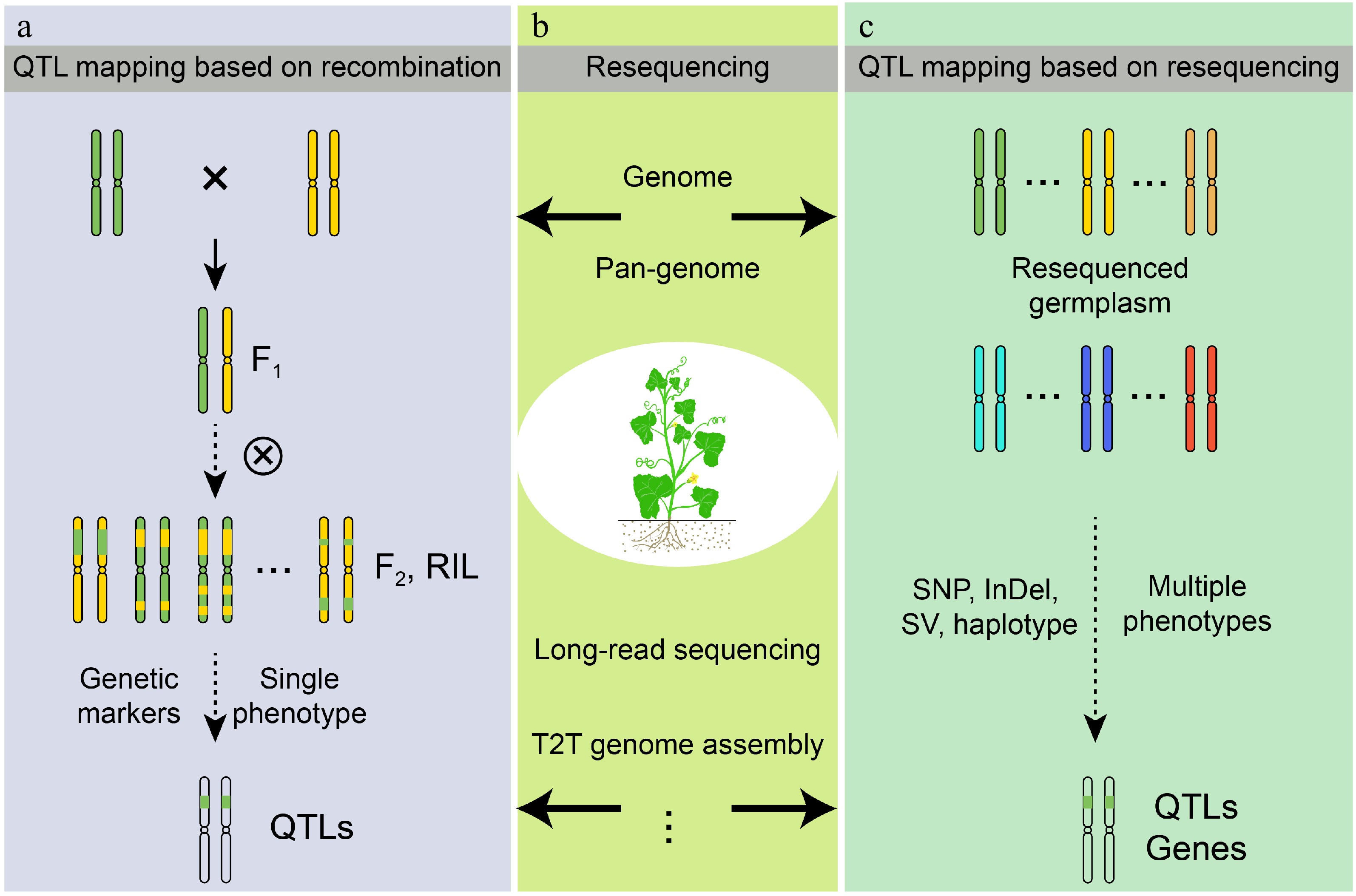
Figure 2.
Development of QTL mapping methods. (a) QTL mapping based on genetic recombination. The progeny population was generated through hybridization and self-fertilization, while QTL mapping was conducted by integrating genetic markers with the phenotypic data of the progeny. (b) Advancements in genome sequencing and assembly technologies serve as formidable instruments for enhancing crop research and development. (c) QTL mapping utilizing resequencing techniques facilitates the swift identification of QTL through the diverse variations and phenotypes present.
The development of melon linkage maps has significantly facilitated the identification of QTL associated with key traits in melon. However, this approach to QTL mapping is subject to certain limitations. Firstly, the low-resolution poses limitations. Substantial chromosomal distances can result in inaccurate QTL localization or the conflation of multiple adjacent QTLs into a single entity. Secondly, constraints arise from genetic markers. The construction of linkage maps is contingent upon genetic markers, yet their quantity and distribution are inherently limited. Furthermore, due to the reliance on genetic linkage maps, the conversion between genetic distance and physical distance obtained through QTL mapping based on these maps is challenging. This difficulty leads to an inability to accurately reflect the true physical positions of QTLs, thereby causing potential location biases in the QTL mapping results. Lastly, constructing recombinant populations, particularly RILs, is labor-intensive and may require seven to eight years for completion. Therefore, there is an urgent necessity to refine the current QTL mapping methodology or to develop novel QTL mapping approaches aimed at enhancing both precision and accuracy in localization.
The genome of melon
-
The utilization of advanced genomic tools and a diverse array of germplasm forms the foundation for employing sophisticated genetic strategies to elucidate complex agronomically significant traits[26,27]. The rapid advancement of next-generation sequencing technology has facilitated the completion of genome assemblies for numerous crops, thereby creating new opportunities to investigate genome evolution and identify genes associated with important traits[1,28]. The cucumber was the inaugural cucurbit crop to have its genome sequenced, significantly facilitating the identification of subsequent cucumber QTL and the investigation of functional genes[28−32]. The sequencing of higher plant genomes is increasingly recognized as a strategic asset in the advancement of accelerated plant breeding techniques, and the accessibility of high-quality representations of the melon genome is crucial. The assembly and annotation of the diploid melon DHL92 genome sequence was published in 2012 (version 3.5.1). The assembled genome spans 375 Mb, which is estimated to represent approximately 83.3% of the total melon genome, and comprises 27,427 genes with an average length of 2,776 base pairs (bp) per gene[1]. The genomic comparison between melon and cucumber has demonstrated a high degree of synteny, with the identification of 19,377 orthologous gene pairs. The utilization of genome sequences will enhance our understanding of cucurbit evolution and refine breeding strategies. However, six years later, the newly assembled melon genome (v3.6.1) along with its annotation has introduced over 8,000 novel genes compared to v3.5.1 and identified a total of 29,980 protein-coding regions, thereby improving the integrity of gene structure[26].
As research advances, the existing single genome data remains inadequate for a thorough genetic analysis and localization of melon, failing to encompass the complete genomic landscape of the species. Various genetic variants are frequently observed in distinct germplasm, exhibiting a random distribution across the genome rather than being confined to a single locus, particularly when substantial segments of SV are not adequately represented by a singular reference[21]. With advancements in sequencing and genome assembly technologies, the accuracy of the melon genome has progressively improved. In recent years, chromosome-scale high-quality assemblies utilizing long-read sequencing, short-read sequencing, Hi-C technology, and single-molecule real-time sequencing have been extensively applied to melons, enhancing the assembly quality of centromeres and telomeres[33,34]. Apparently, significant progress has also been achieved in the research of melon genomics. Yano et al. updated the tissue-wide RNA-seq in the Melonet-DB database[35]. Oren et al. identified potential candidate genes within multi-trait QTLs[36]. Lyu et al. established the melon pan-genome by integrating the chromosome-level genome assembly of 'Mapao' melon (C. melo ssp. agrestis) with sequenced genomic datasets from other melon varieties[37]. Li et al. successfully accomplished the T2T genome assembly of '821' (C. melo ssp.agrestis) at the chromosome level, and identified a significant number of SVs that differentiate the genomes of melo and agrestis[38]. The pan-genome addresses the limitations of a single reference genome, offering additional resources for the identification of significant trait genes in melon and establishing a crucial foundation for its biological breeding. This high-quality genomic assembly data facilitates the identification of increasingly precise variants that are beneficial not only for constructing genetic linkage maps but also for conducting large-scale phenotype-variant correlation analyses (Fig. 2b).
Population resequencing accelerates QTL mapping of melon
-
The advent of genome sequencing has facilitated the emergence of population resequencing, which has proven effective in QTL mapping for melon[27]. Population resequencing can accelerate QTL mapping in melon. Conventional QTL mapping is restricted to constructing populations for a limited number of traits, followed by individual genotyping and phenotyping, which is both time-intensive and laborious. However, QTL mapping facilitated by population resequencing technology can circumvent the laborious process of constructing recombinant populations and instead leverage natural populations derived from wild germplasm for mapping purposes, such as GWAS based on re-sequenced natural populations[39]. Furthermore, the natural population can simultaneously investigate QTL for multiple traits[39]. Population resequencing can also yield more comprehensive information on genomic variants, including SNP, InDel, SV, and copy number variation (CNV). These variant types may significantly influence the phenotypic traits of melon. By examining the associations between these variants and phenotypes, it becomes possible to identify gene regions linked to traits of interest, thereby facilitating the advancement of QTL mapping (Fig. 2c).
In recent years, QTL mapping based on population resequencing has advanced significantly in the study of melon. High-resolution maps generated through genome resequencing significantly enhance the efficiency of QTL localization[23,40]. A research investigation employed GWAS utilizing deletion/presence variants of genes within a pan-genome comprising 297 melon accessions, successfully identifying 13 genes linked to fruit shape in melons[41]. Through the identification of 187 melons via GWAS, a significant QTL was discovered on chromosome 11 that is strongly associated with fruit surface groove[42]. Low temperatures significantly impacted the growth and development of melons, particularly during the seedling stage, potentially resulting in a halt in seedling growth[43]. A total of 51 loci associated with cold resistance were identified through GWAS involving 212 melon accessions. Notably, CMCT505_Chr. 1 was consistently detected across various populations and exhibited a significant phenotypic contribution, suggesting it may serve as a potential key gene regulating cold resistance in melons[43]. Population resequencing facilitates simultaneous QTL mapping for multiple traits. For instance, a GWAS conducted on 1,175 re-sequenced melon accessions identified a total of 208 loci significantly associated with fruit weight, morphology, and quality across 16 agronomic traits[27]. To date, a substantial number of QTLs have been identified, with several genes successfully cloned and their functions validated. Over 1,294 QTLs related to melon have been documented, encompassing seven categories: fruit traits, seed characteristics, disease resistance, floral attributes, vegetative organs, insect resistance, and abiotic stress tolerance. These findings are comprehensively summarized in the context of this recent research study[44].
In comparison to traditional recombinant population-based QTL mapping, QTL mapping through population resequencing offers several advantages. Firstly, it enables genome-wide scanning, thereby facilitating high-resolution QTL identification. Secondly, it is not constrained by a specific genetic background and can be applied across various species and populations. Furthermore, can elucidate the complexity of quantitative traits, including the interactions among multiple genes and environmental factors. Lastly, it can identify candidate genes associated with quantitative traits. In conclusion, genome resequencing holds significant potential for enhancing QTL mapping in melon and presents promising prospects for melon biological breeding.
Population resequencing based on reference genomes not only facilitates QTL mapping but also aids in the cloning of functional genes, applicable to both genotyping of recombinant populations and GWAS of natural populations, thereby expediting the screening and identification of candidate genes. With advancements in biotechnology, the acquisition of new melon germplasm, and the identification of superior traits, it is anticipated that a greater number of melon genes will be cloned alongside comprehensive functional studies being conducted. These investigations will provide a crucial scientific foundation for enhancing our understanding of regulatory mechanisms and improving varietal development in melons.
Application of resequencing in gene cloning of melon
-
Currently, the prevalence of gene cloning and functional studies in melon is significantly lower than that observed in staple crops such as wheat and rice. However, the role of reference genomes and resequencing in facilitating gene cloning should not be underestimated, particularly in their capacity to assist in the screening and identification of candidate genes. In a study concerning the shape of melon fruit, BSA-seq and Bin-Map linkage maps of F2 individuals were employed to precisely locate a region spanning 53.7 kilobase pairs (Kb), ultimately leading to the cloning of the target gene CmFSI8[45]. In a related study, two loci associated with fruit ripening were identified on chromosome 8 through a genome-wide association study (GWAS) involving 211 melon samples. Coupled with fine mapping of the recombinant inbred line (RIL), it was concluded that ETHQV8.1 may serve as a candidate gene for the regulation of fruit ripening[46]. These studies demonstrate that population resequencing can enhance both the speed and accuracy of population genotyping, thereby facilitating the fine mapping of target genes. Furthermore, GWAS can directly pinpoint target genes associated with specific traits. For instance, a GWAS investigating low-temperature germination in cucumber was conducted on 151 accessions, resulting in the identification of seven significant loci across four chromosomes. Through functional annotation and haplotype analysis, the candidate gene CsPPR emerged as a regulatory locus for low-temperature germination, with its localization accuracy further validated by ectopic expression[47]. In a separate investigation, a GWAS involving 318 re-sequenced maize lines was conducted to identify a significant locus associated with sheath blight resistance. The most notable SNP was found within the ZmFBL41, which was subsequently validated through transgenic experiments[48].
-
The improvement of melon varieties is a protracted endeavor that necessitates sustained efforts from breeders and researchers to achieve successful outcomes. Drawing upon existing research, we explore strategies for future melon breeding, which encompasses breeding strategies, biotechnology, and artificial intelligence. For these strategies, we have provided a concise schematic representation and summarized their respective roles in the context of melon breeding (Fig. 3). Figure 3 illustrates an integrated strategy model for melon breeding, encompassing the expansion of melon germplasm resources, the pyramiding of multiple superior QTLs, the application of gene editing technologies, and the development of a high-throughput phenotypic identification system utilizing artificial intelligence.
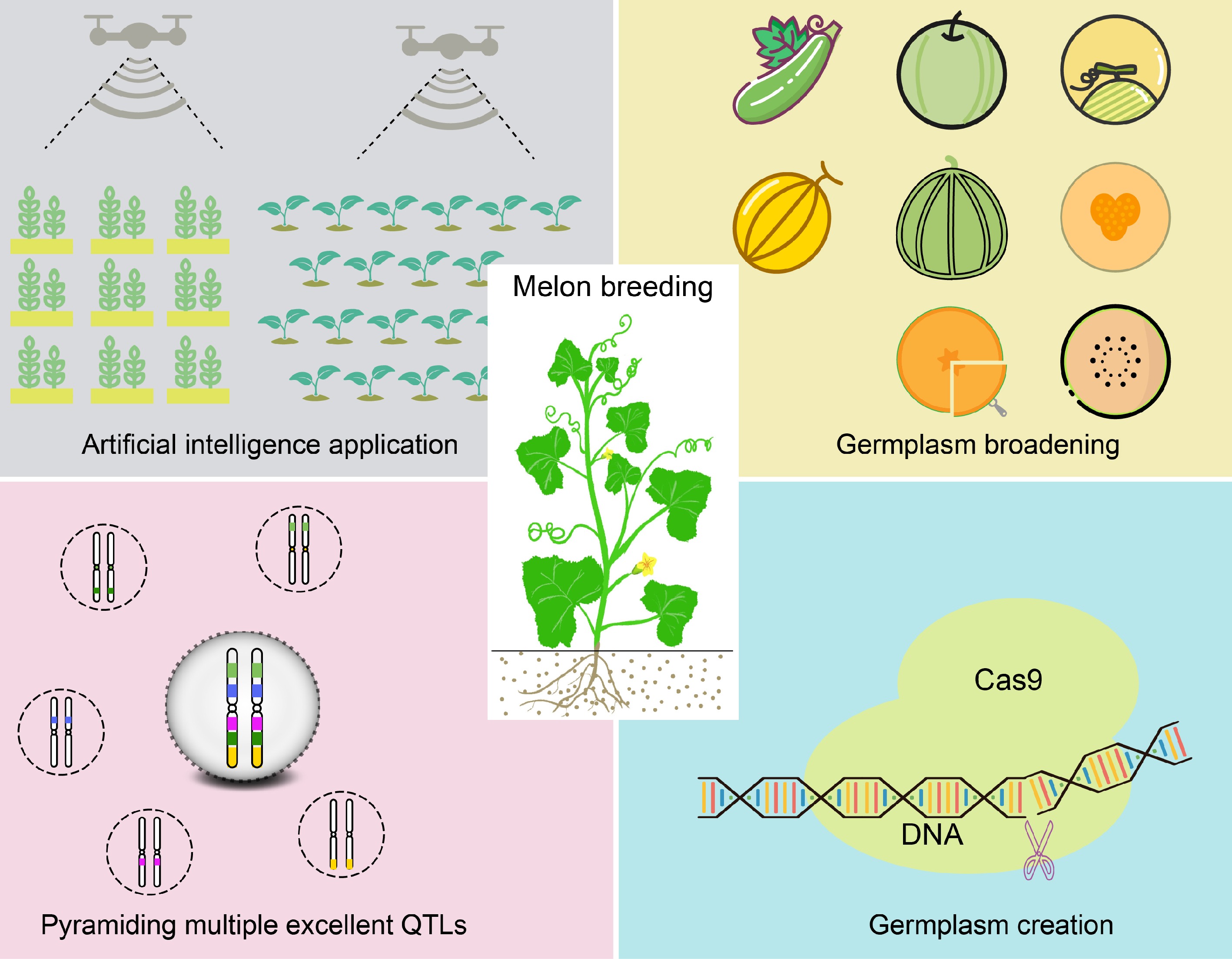
Figure 3.
Integrated strategies for melon breeding. Develop a high-throughput phenotypic identification system utilizing AI technology to expedite melon breeding. Broadening melon germplasm facilitates the identification of novel superior genes. The pyramiding of multiple excellent QTLs aids in enhancing desirable characteristics in melons. Gene editing technologies can be employed to develop new germplasm.
Broadening genetic diversity in melon germplasm
-
Significant genetic and morphological variations exist among different melon germplasm[27]. Melon breeding is essential to achieve high yields and meet diverse consumer demands regarding quality. Melon germplasm resources also display a certain degree of diversity in tolerance levels against pests, diseases, drought, salinity, and other adversities. Currently, research on melons predominantly focuses on QTL mapping, with the majority concentrating on fruit weight, fruit color, and disease resistance[4]. The genetic enhancement of melons extends beyond fruit quality. There is a growing emphasis on the study of melon plant architecture. The architecture of melon plants directly influences land utilization, spatial efficiency, and mechanized management, which in turn can impact overall yield. Research indicates that the number of fruits per plant in melons increases with greater plant spacing[49]. Highly branched melon plants contribute to the development of high-yield varieties that are well-suited for mechanical harvesting[50]. The genetic diversity of germplasm is critically important for the breeding and cultivation of melons. By fully leveraging this diversity, superior varieties can be developed to enhance yield and quality, improve resistance to pests and diseases, and adapt to a broader range of growing environments. Furthermore, the preservation of germplasm diversity plays a vital role in safeguarding the genetic resources of melons, thereby preventing germplasm loss and mitigating reductions in genetic variability.
The findings from current research suggest that both natural germplasm and resynthesized melon lines exhibit substantial DNA sequence variation, including SNPs, InDels, and SVs. Some of these variations are significantly correlated with specific traits in melons, thereby providing direct utility for our applications[21,37]. In instances where germplasm resources are limited or mutations are not sufficiently widespread, chemical mutagenesis or gene editing techniques can be employed to generate new germplasm. Although the alterations induced by these methods may be random, a certain proportion of them is likely to yield beneficial traits. The utilization of such mutants is currently observed in various crops[51]. The generation of new recombination through the construction of introgression lines (ILs) represents a viable strategy for developing novel germplasm. For instance, in tomato breeding, an introgression line was established by incorporating chromosomal segments from wild tomato species into a cultivated variety, ultimately leading to the identification of 326 loci associated with yield-related traits within this high-diversity population[52]. As global climate patterns and growing conditions evolve, plants are compelled by environmental pressures to generate novel mutations for enhanced adaptation, which may also result in the emergence of new superior traits that we can select and utilize.
Pyramiding multiple excellent QTLs
-
Research indicates that the majority of traits, including melon yield, quality, and resistance, are quantitative in nature and governed by multiple genes, and this mode of inheritance is characterized as additive[4,53]. Consequently, efficient molecular breeding techniques can be employed in melon breeding to consolidate favorable alleles across multiple QTLs into elite cultivars. The fundamental concept of QTL pyramiding is to integrate several QTLs with beneficial effects into a single breeding line, aiming for enhanced phenotypic performance; this process necessitates the identification of specific QTL locations and their corresponding molecular markers. QTL pyramiding is accomplished through crosses between two parents exhibiting distinct traits, followed by multiple backcrosses and self-pollinations. Each backcross should incorporate molecular marker-assisted selection to identify progeny possessing the target QTLs, which are then self-pollinated to produce stable and pure germplasm enriched with aggregated QTLs for various traits, ultimately enhancing both the quality and yield of melons. This methodology can also be applied to aggregate different QTLs associated with a particular trait.
QTL aggregation is predominantly employed in rice and wheat cultivation. Research has demonstrated that the integration of alleles from the rice blast resistance QTLs pi21, qBR4-2, Pi34, qBR12-1, and Pi35 into elite varieties possessing major resistance genes can potentially confer broad-spectrum resistance (BSR) in rice[54,55]. The drought yield QTL qDTY2.2, qDTY3.1, and qDTY12.1 were integrated into the variety 'MRQ74'. The resulting improved rice varieties, characterized by different QTL combinations, exhibited varying yields under drought conditions[56]. In wheat, the integration of the QTLs Lr34 and Lr67 with other dominant resistance genes can significantly improve the resistance to rust[57]. In melon, three climacteric QTLs, ETHQB3.5, ETHQV6.3, and ETHQV8.1, were pyramided into the C. melon cultivar 'Piel de Sapo', and resulted in an increased ethylene peak due to ETHQB3.5, enhanced ethylene production promoted by ETHQV6.3, while ETHQV8.1 synergistically augmented the effects of the other two QTLs[46]. In summary, the enhancement of melon quality and yield through QTL pyramiding presents significant potential and holds considerable application value. The aggregation of multiple desirable traits necessitates extensive repetitive crossings and selection processes, which demand a significant investment of time. However, the application of modern biotechnological advancements, such as gene editing and artificial intelligence, can substantially enhance efficiency.
Optimization of melon plant architecture
-
Plants cultivated using vertical farming methods can significantly reduce leaf overlap, thereby enhancing overall photosynthetic efficiency[58]. The planting density of melons exhibited a negative correlation with fruit weight. It was observed that the weight of melon fruits was significantly greater at a planting density of two plants/m² compared to densities of four plants/m² and eight plants/m2[59]. The primary factors contributing to this issue are, firstly, that high-density planting results in an inadequate light interception by the plants, thereby diminishing their photosynthetic efficiency[58,60]. Secondly, such planting practices impair nutrient absorption at the individual plant level[61]. Consequently, it is advisable to minimize inter-plant crossing in melon cultivation. To address these challenges, we propose an ideal plant architecture (IPA) for melons aimed at enhancing production outcomes. The essential characteristics of the melon IPA are manifested in its design concerning plant height, branching structure, stem configuration, and root system.
The optimal height for melon IPA should not exceed 1.5 m, as this facilitates streamlined and mechanized production while minimizing management challenges. At the same planting density, maintaining a low plant height effectively mitigates the sheltering syndrome associated with mutual shading[58]. Similarly, the melon IPA should exhibit fewer branches, which ought to be relatively short and positioned at a more acute angle to the main stem. Ideally, it should have 3−4 branches, each not exceeding 20 cm in length. The branches must possess an adequate number of leaves for effective photosynthesis, thereby supplying sufficient nutrients for optimal fruit development. This approach allows for increased planting density while ensuring that the melon plants do not overshadow one another. At present, the gene responsible for regulating short lateral branches, ClphyB, has been successfully cloned in watermelon, and this architectural design is also appropriate for melons[62]. The melon IPA features a robust root system that effectively absorbs and delivers sufficient water and nutrients to the plant, which can significantly mitigate the nutritional deficiencies resulting from increased planting density. Furthermore, a robust root system enhances the stability of the plant, enabling it to better withstand external forces such as wind and rain, while also improving its resistance to environmental stresses including pests, diseases, droughts, as well as saline-alkaline conditions[63]. The melon IPA should possess a more robust stem, as a thicker stem provides enhanced support for the weight of the fruit, thereby preventing it from toppling or damaging the stalks and facilitating optimal growth and development. The development of such a melon IPA is not solely contingent upon the diversity of available germplasm resources and the integrated application of biotechnological methods; it also necessitates extensive practical experience in screening.
The application of gene editing technology
-
R genes derived from wild species can be introduced into elite cultivars through human intervention, including conventional transgenic techniques, which not only enhance disease resistance in crops but also yield significant economic advantages. However, as pathogens evolve, the efficacy of many R genes may diminish or even be completely lost. Therefore, it is imperative to investigate alternative sources of resistance, such as those conferred by mutations in S genes within the host. Advancements in biotechnology have enabled the application of gene editing techniques to modify the genetic information of crops as needed, including the knockout or deletion of specific genomic segments[64,65], even the modification of individual bases within the genome[66]. Modified genes exhibit changes in their coding products or expression levels, which directly lead to alterations in functional outcomes[67]. This technology offers significant convenience for the modification of the S gene. Furthermore, it is particularly encouraging that this gene editing technique can be applied to a majority of crops with available reference genomes, and that optimizing the DNA sequence at the target site can substantially enhance primer design efficiency.
Numerous reports have documented enhanced resistance achieved through gene editing of susceptibility genes in various crops, with a particularly notable example being the application of mutations in the MILDEW-RESISTANCE LOCUS (MLO) and its homologs to confer resistance against powdery mildew. The simultaneous mutation of all three copies of TaMLO in wheat has successfully resulted in heritable broad-spectrum resistance to powdery mildew in plants[68]. In tomato, the cloning of the recessive broad-spectrum resistance locus ol-2 against powdery mildew revealed that its resistance is attributed to a functional deletion of SlMLO1. Furthermore, the knockout of the direct homolog SlMLO1 resulted in complete resistance to powdery mildew in tomato[69,70]. Similar studies have also been documented in Arabidopsis, pea, rapeseed oil, and pepper, as well as in the monocotyledonous rice species[71,72]. These studies not only suggest that the natural mlo allele may be more effectively adapted to its environment through the loss of MLO function during evolution but also predict that homologous genes could perform analogous functions across different species. This provides a valuable reference for melon breeding. Gene editing technology also contributed significantly to the improvement of plant height, flowering time, yield, and quality[73,74]. The applicability of gene editing technology in melon crops has been validated, and the editing efficiency continues to improve progressively[75]. Wang et al. demonstrated that CRISPR systems incorporating tRNA and Csy4 spacers under the control of Pol II-type promoters can enhance editing efficiency by up to 42.8%[76]. The application of gene editing for enhancing melon phenotypes has been a well-established reality for an extended period. Giordano et al. utilized gene-editing technology to generate ctr1-like and ros1 double mutants in melon, thereby successfully advancing the understanding of melon ripening regulation and improving its ripening time[77]. Another study demonstrated that the knockout of CmACO1 resulted in a significant reduction in ethylene production in melons, a notable increase in fruit firmness, and the successful creation of novel germplasm for melon breeding[78]. Knocking out CmMLO5 in melon confers resistance to powdery mildew[79]. Optimal utilization of gene editing techniques and genomic datasets can profoundly augment the potential for genetic enhancement in melons.
Enhance the support provided by artificial intelligence technology
-
Crop breeding necessitates substantial time and labor for phenotypic identification, which poses a significant constraint on the genetic enhancement of crops. Nevertheless, with advancements in AI and interdisciplinary collaboration, the efficiency of plant phenotypic identification can be significantly enhanced. In recent years, computer-assisted image analysis has significantly contributed to the extraction of morphological features pertinent to plant phenotypes[80]. Building upon this foundational framework, plant phenomics has experienced substantial advancements, facilitating high-throughput phenotypic analysis of complex plant traits[81]. The AI-driven phenotyping system predominantly consists of computer vision sensors, an extensive database, and advanced analytical software[82]. The computer vision component is adept at processing digital images of plant tissues to extract pertinent image features[83]. The database enhances information sharing and resource accessibility[84]. The software employs a diverse array of tools and algorithms to classify and characterize large datasets of crop phenotypic imagery, thereby producing the most insightful analytical outcomes[84]. In addition to AI-driven digital phenotypic data collection, the application of artificial intelligence in crop breeding further includes predictive analytics for elucidating genomic information, the integration of multi-omics big data, facilitating the correlation between genotype and phenotype, advancements in functional genomics and gene discovery, identification of high-quality alleles and causal variants from omics datasets, as well as enhancing practical plant breeding through genomic selection aimed at phenotype prediction[85].
AI technology has also been utilized in melon phenotyping. Research indicates that accurate predictions of fruit weight can be achieved by employing artificial neural networks as a tool for forecasting melon yields[86]. Research has demonstrated that melon production can be accurately estimated through the analysis of overhead color images captured by drones, which facilitate the estimation of both the quantity and individual weight of melons[87]. Beyond that, AI technology plays a significant role in the assessment of melon quality. A predictive model for solid solution concentration and moisture content in melons was developed utilizing hyperspectral imaging technology, demonstrating effective prediction performance[88]. Recently, a software application named CucumberAI has been developed specifically for the analysis of morphological characteristics in cucumber fruits, demonstrating efficacy in identifying up to 51 distinct features of cucumbers[89]. The applications of artificial intelligence in field phenotyping remain nascent; however, we are confident that with ongoing advancements, it will undoubtedly be implemented on a large scale in the future.
AI is increasingly being leveraged for the interpretation of genomic data and the characterization of germplasm resources[85]. By integrating genomic information with germplasm genotype data, genome prediction models can simulate the performance of diverse germplasm resources, thereby facilitating the selection of optimal breeding materials[85,90]. The convergence of AI and genomics has demonstrated significant potential in addressing plant stress biology challenges. Studies have demonstrated that AI can predict genomic hybridization in maize plants, distinguish functional genes from pseudogenes by identifying and characterizing genomic regions[91], predict gene promoters and cis-regulatory elements[92], recognize tissue-specific variations in biosynthetic genes, decode plant metabolic regulatory networks[93], manage bioenergy resources, and optimize biomass production[94].
-
The research in melon genomics offers robust technical support for melon breeding. The release of reference genomes and the utilization of third-generation sequencing technology have markedly enhanced the completeness and precision of genome assembly, thereby establishing a solid foundation for molecular marker development and gene cloning. Genome resequencing technology can produce extensive genomic data, encompassing high-density markers across the entire genome, which facilitates more precise localization and identification of genes associated with target traits. With advancements in molecular marker-assisted selection and whole-genome background selection, breeders are now able to select and improve melon varieties with greater efficiency. Since the initial release of the melon reference genome in 2012, several high-quality iterations have been developed and published. Telomere-to-telomere genome assembly facilitates the construction of a complete genome at the gap-free level[38,44]. The pan-genome integrated 12 distinct ecotypes, encompassing both wild and cultivated species, and uncovered structural variations associated with disease resistance and fruit quality. This provides valuable resources for the identification of superior alleles[37]. Genome resequencing has significantly advanced the genetic analysis of key traits in melon crops. The carotenoid synthesis gene (CmOr) was successfully cloned[95]. The ethylene synthesis pathway, mediated by CmACS7, plays a crucial role in regulating fruit ripening[96]. The localization of powdery mildew resistance genes (Pm-w, Pm-2F) and antiviral genes (Cm-eIF4E) has provided a critical foundation for the development of molecular markers[97]. The regulatory networks of sex-differentiation genes, including CmWIP1 and CmACS7, have been thoroughly elucidated to facilitate the breeding of parthenocarpic flower varieties[98]. In the context of genomics-driven molecular breeding techniques, SNP markers developed through QTL mapping and GWAS are extensively utilized for the precise selection of disease resistance and fruit quality traits[21], CRISPR-Cas9 technology has been successfully utilized to enhance the genetic traits of melon crops[79].
The advancement of genomics has facilitated significant progress in melon breeding; however, further efforts are essential to achieve technological breakthroughs and optimal outcomes. Firstly, enhancing sequencing accuracy and reliability is critical. While high-throughput sequencing rapidly generates vast amounts of data, its precision requires improvement. Issues such as sequencing errors and uneven local sequence coverage can compromise subsequent data analysis. Secondly, accelerating the development of cell engineering breeding technology is imperative. The progress in cell engineering and ploidy breeding for melons has been relatively slow, limiting its widespread application in breeding programs. Improving the seed germination rate of triploid melons could facilitate broader adoption of seedless varieties. Thirdly, establishing an efficient transformation system is necessary. Although gene editing technologies have advanced theoretically, the current melon gene editing system relies heavily on Agrobacterium-mediated genetic transformation, lacking a robust and stable transformation system. Future efforts should focus on developing a mature melon genetic transformation system to maximize the potential of gene editing in breeding practices. Finally, diversifying breeding objectives is crucial. In melon breeding, we must not only prioritize disease resistance and quality enhancement but also integrate emerging technologies like high-throughput phenotyping systems to comprehensively improve plant architecture and growth duration. This will require a substantial number of researchers to collaborate to accomplish the task.
This research was supported by Key Research and Development Plan of Anhui Province (2023z04020019-02), Natural Science Research Project of Colleges and Universities of Anhui Province (2024AH051667), and the Science Fund for Distinguished Young Scholars Higher Education of Anhui Major Science and Technology Project of Anhui Province (2022AH020037). We extend our gratitude to Kuipeng Xu from the College of Horticulture at Qingdao Agricultural University for valuable contributions in revising this paper.
-
The authors confirm contributions to the paper as follows: conceptualization: Liu J, Wang J, Guo Y; methodology and original draft writing: Liu J; investigation: Wang J, Guo Y, Li H, Liu C, Fan Y; data curation: Fan Y, Ma J; resources: Ma J, Zhang H; writing—review and editing: Liu J, Zhang H; project administration: Zhang H. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu J, Wang J, Guo Y, Li H, Liu C, et al. 2025. Novel molecular design breeding strategies enabled by genome resequencing in melon. Vegetable Research 5: e025 doi: 10.48130/vegres-0025-0016 

Novel molecular design breeding strategies enabled by genome resequencing in melon
- Received: 17 November 2024
- Revised: 23 March 2025
- Accepted: 30 April 2025
- Published online: 29 July 2025
Abstract: It has been over a decade since the publication of the melon genome, which has significantly contributed to our understanding of gene function and enhanced interspecific communication. Furthermore, it has facilitated the application of modern breeding techniques such as genomic selection and marker-assisted selection, thereby improving both the efficiency and precision of melon breeding. Nevertheless, in light of the growing global population and unpredictable climate change, melon breeding continues to confront challenges related to increasing production and enhancing environmental resilience. In this review, we provide a comprehensive summary of the nutritional and medicinal properties of melon, while also examining the evolution of quantitative trait locus (QTL) mapping in melon—from traditional QTL mapping utilizing recombinant populations to advanced QTL mapping techniques developed through population resequencing following the publication of its genome. We emphasize the significance of melon genome development and the potential of genetic diversity, along with contemporary biotechnological methods, in enhancing melon breeding. In conclusion, we propose several recommendations for the future direction and strategy of melon breeding, which include the application of gene editing and artificial intelligence technologies, the expansion and exploration of germplasm resources, as well as the optimization of melon plant architecture. Our suggestions will contribute significantly to advancements in melon breeding.
-
Key words:
- Melon /
- Breeding /
- Genome /
- Resequencing /
- Gene editing /
- Artificial intelligence