









-
Use of digital displays has increased worldwide, and exposure to blue light (380–520 nm) has risen accordingly, with reports linking such exposure to eye strain, sleep disturbance, and diminished concentration[1,2]. Experimental studies indicate that short-wavelength blue light can trigger oxidative stress[3−5], and lipofuscin-related toxicity in retinal cells[6,7], although many of these protocols involve intensities and durations exceeding typical daily exposure. Recent clinical evidence indicates that blue-filtering lenses do not alleviate ocular-surface dryness, tear-film instability, or dry eye symptoms after 120 min of digital reading[8].
Blue light has been investigated in relation to alertness and cognitive performance, although findings remain heterogeneous. Longitudinal observations during continuous wear of blue-light-filtering (BLF) lenses have described declines in sustained attention and visuospatial working memory that reversed after discontinuation[9], whereas nighttime amber filtration improved selected cognitive domains in adults with insomnia[10], and evening filtration in adolescents affected circadian-related measures[11]. Objective studies of accommodation and pupil dynamics have generally not shown symptom relief or accommodative benefit in healthy young adults, despite occasional increases in reading speed[12]. A recent systematic review concluded that short-term BLF use provides little to no advantage for digital eye strain or near-term visual performance relative to non-filtering lenses[13].
Contemporary information use frequently involves brief, time-constrained bouts of reading rather than extended, continuous engagement. Immediate neurophysiological adjustments during such periods are less well characterized, particularly with simultaneous measurements. Oculomotor control and cortical oscillations exhibit task-dependent modulation on short timescales during demanding text processing, supporting designs that probe early adjustments rather than prolonged exposure[14−16].
The present study employs a two-minute timed-reading paradigm to examine acute load under BLF, versus control lenses. Passages were presented under controlled illumination to isolate spectral-input effects on reading. Simultaneous eye-movement and electroencephalogram (EEG) recordings were acquired to provide complementary indices of processing demands and effort allocation[17]. In interpreting lens-related physiological modulation, we distinguish observed differences from functional improvement and evaluate correspondence with reading-comprehension accuracy within the same task window[18].
The following were assessed: (1) whether short-term BLF wear modulates eye-movement and EEG indices during brief, timed reading; (2) whether reading-comprehension accuracy differs between lens conditions within this window; and (3) whether the joint oculomotor-EEG pattern is more consistent with compensatory, load-related adjustment to altered spectral input than with performance enhancement.
-
The required sample size for the study was determined using a G*Power (version 3.1.9.7) power analysis[19]. The calculation was conducted using a paired-sample t-test, reflecting the within-subject crossover design of the experiment. Following Cohen's conventional criteria for behavioral research, and consistent with typical practice in exploratory behavioral and EEG studies, the analysis specified a medium effect size of d = 0.50[20], a two-tailed significance level of α = 0.05, and a statistical power of 0.80. This resulted in a required sample size of 34 participants for detecting medium within-subject differences between lens conditions, rather than for formally establishing non-inferiority or equivalence. The selected sample size aligns in magnitude with that of a previous randomized, placebo-controlled crossover study employing BLF lens interventions in psychological contexts[21].
Accordingly, 34 right-eye dominant participants were recruited from Korea University (24 males, mean age: 23.5 years, SD = 1.88). All participants had normal or corrected-to-normal binocular visual acuity of 0.1 ± 0.1 logMAR or better, and had no history of ophthalmologic or neurological diseases. Normal vision and binocular function were confirmed for all participants through a medical history check and objective testing (Worth-4-dots and Titmus stereo tests), which was administered by an expert from the Korean Optometric Association. All participants reported that they had never previously worn BLF spectacle lenses, and normal spectacles were used for vision correction when needed. Ethical approval was granted by the Internal Review Board (IRB) of Korea University (KU-IRB-15-205-A-1), with written informed consent from all participants.
Lens
-
We used two types of commercially available lenses, both with identical frames and coatings. The experimental lenses featured a BLF coating (see Fig. 1), while the control lenses were clear lenses without this specific filtering property (non-filtering clear lenses). This design allowed for a placebo-controlled comparison. The BLF lenses used in this study were single-vision spherical plastic lenses with a refractive index of 1.56, manufactured by a single commercial vendor (HOYA, Korea). These lenses incorporated a blue-light-filtering treatment combined with a standard multilayer anti-reflective (AR) coating. The control lenses had the same refractive index and AR coating as the BLF lenses, but no blue-light-filtering treatment.
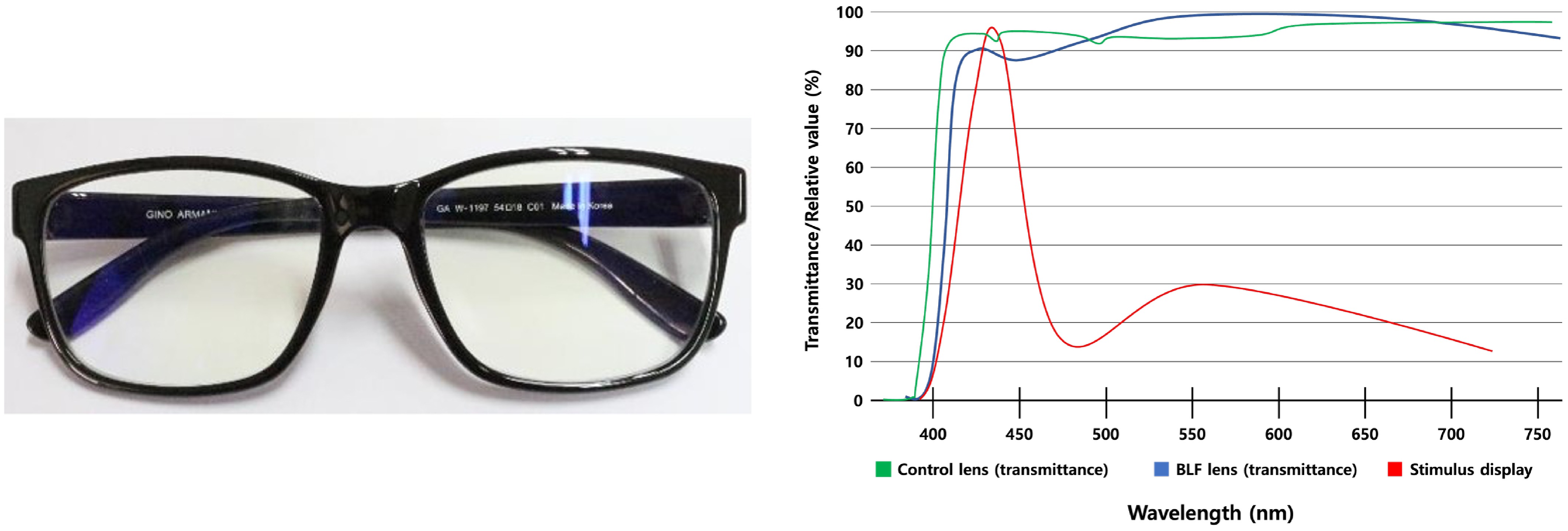
Figure 1.
Blue-light filtering (BLF) spectacles and optical characteristics of the BLF lens and stimulus display. Each participant was fitted with a BLF lens (shown on the left). The spectral transmittance of the BLF lens (blue curve), control lens (green), and the relative spectral output of the stimulus display (red curve), are plotted as percentages of maximum values. Spectral measurements were measured using a spectrophotometer (OPTIZEN 2120UV, Korea,
english.mecasys.co.kr ).Electroencephalogram (EEG)
-
For EEG recordings, we used a Neurofax JE120A, 256-channel, power amplifier (Nihon Kohden, Japan,
www.nihonkohden.com ). The small voltage signals were amplified and sampled at 10 kHz. 20 Ag/AgCl nonmagnetic electrodes (50 μV sensitivity) were positioned on an elastic cap according to the standard 10−20 system with mastoid reference A1↔A2. During setup, gel was applied to the electrodes to ensure resistance below 10 kilo Ohms. The signal was filtered using optimized band-pass filters between 0.53 and 100 Hz. One additional electrode was placed under the left eye for EOG, and the other under the right eye to provide a coarse check for eye-movements.Eye tracker
-
Ocular movements were recorded with a corneal reflection eye tracker (Tobii 120 Eye Tracker, Tobii Technology, Danderyd, Sweden,
tobii.com ). The eye tracker recorded both eyes at 120 Hz with an average accuracy of 0.5° visual angle. Although the eye tracker supports free viewing, the participant's head was fixed using a chinrest to control the distance to the screen at 60 cm (Fig. 2).
Figure 2.
Eye tracking and EEG experimental setup with an observer. Text passages were presented on the integrated display of the eye-tracking system while participants wore either BLF or control lenses. The setup includes a height adjustable chair, table, and chin rest.
The gaze of each participant was calibrated prior to testing. Each participant was calibrated with a standard five-point calibration pattern, in which an expanding contracting circle appeared in every position of a screen-wide grid of calibration points on a white background. Calibration was checked for high error and repeated if necessary.
Display and stimuli
-
The stimuli were generated and controlled using MATLAB (Mathworks, Natick, USA, version 8.4,
mathworks.com ) and Psychtoolbox-3 (Version 2.1,psychtoolbox.org ) for Mac OSX, running on a laptop computer (Apple Mac Pro 5.1, Apple Cupertino, USA,apple.com ). Stimuli were shown on the integrated 17'' TFT stimulus display of the eye tracker (Tobii 120 Eye Tracker, Tobii SE, Sweden,tobii.com ), at 60 Hz frame rate. The illuminance was maintained at ≈ 3 lx (LM-81LX, Lutron, Taiwan,lutron.com.tw ) illumination.Stimuli consisted of short, one-page texts covering six different topics: science, art, astronomy, entertainment, philosophy, and literature. All six passages were excerpted from Korean College Scholastic Aptitude Test (CSAT) reading materials that had been classified as medium difficulty by the official CSAT test committee, and were restricted to a comparable length so that large disparities in reading difficulty were not expected. In a pilot study, six readers (mean age = 24.83 years, SD = 2.23) recruited from the same university as the main experimental participants answered six comprehension questions for each passage; mean accuracy per passage ranged from 4.3 to 4.8 correct responses (out of 6). Subjective difficulty ratings were clustered around 'moderate' (means between 2.3 and 2.7 on a 1−3 scale), suggesting no large between-passage differences in perceived or objective difficulty, although no formal psychometric difficulty indices were computed. Texts were displayed on the screen, one text at a time, at a font size of 10 points (corresponding to a visual angle of ≈ 0.5° per Korean syllable) in Hangeul (Korean script).
Procedure
-
In the present study, we the effect of BLF lenses compared to a baseline, placebo control condition were investigated using three different measurements:
(1) Behavioral performance, in which we asked for people's understanding of the read texts.
(2) Eye tracking metrics that were derived from the eye movements in the different conditions.
(3) Frequency band power estimates that were derived from the EEG data.
The study design compares the two conditions of BLF reading and control lens reading in a within-participant design (see Fig. 3). Six passages were presented in the same fixed order for all participants. Half the texts were read while wearing the BLF lenses, and half with control lenses. To minimize order-related bias, half of the participants read the first three passages with BLF lenses and the remaining three passages with control lenses, whereas the other half read the first three passages with control lenses and the remaining three with BLF lenses. Thus, each passage was read with BLF lenses in half of the sample, and with control lenses in the other half. Participants were randomly assigned to BLF-first or control-first sequences, and lens order was counterbalanced across the sample. Participants were not informed which of the two lenses was the BLF lens or the control lens, and were told only that two different spectacle lenses would be compared, so that testing was effectively single-blind at the participant level.
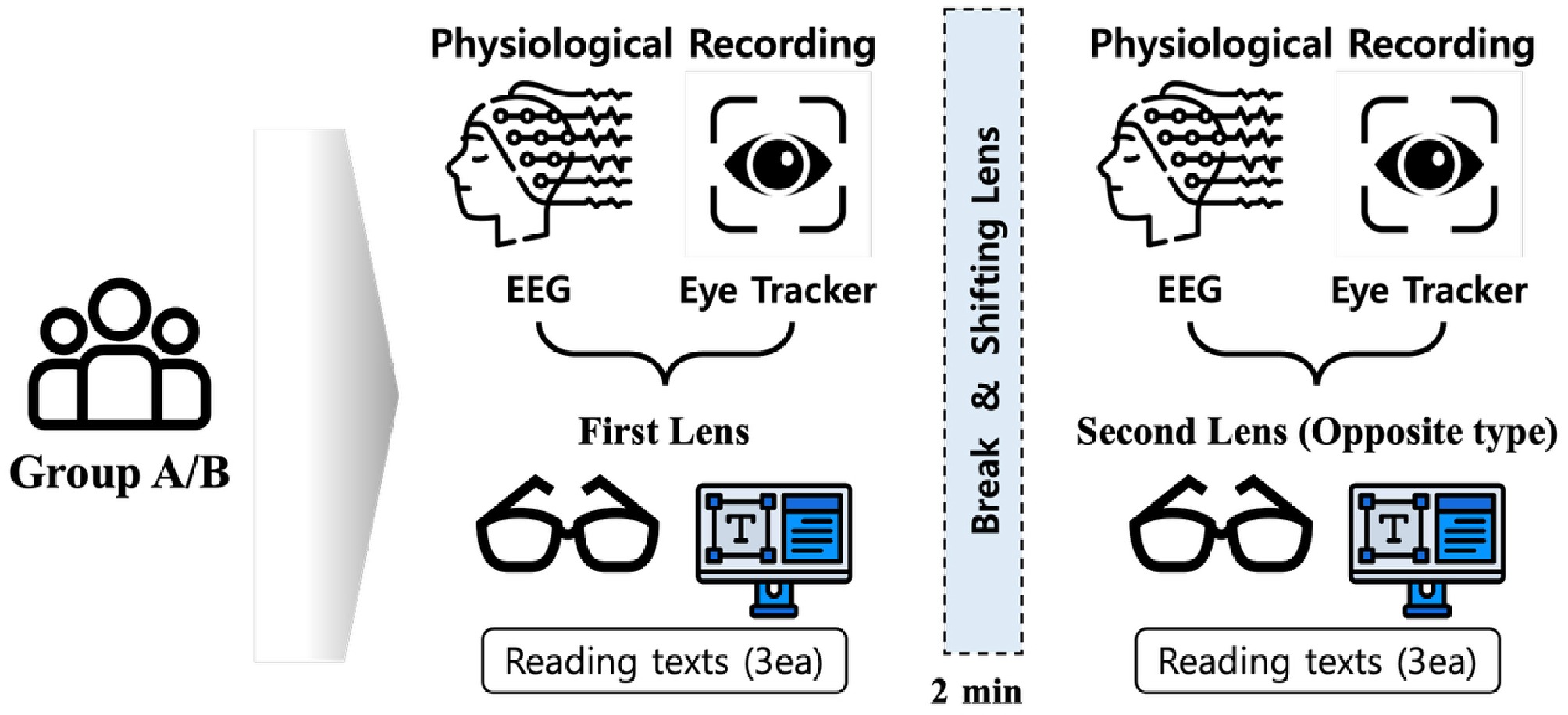
Figure 3.
Schematic representation of the experimental procedure. Each participant underwent reading tasks under two lens conditions (BLF lens and control lens). Two groups initially read three different texts under one lens condition, took a two-minute break and switched to the opposite lens condition, and then read another three different texts. The lens-wearing order was counterbalanced between groups.
Each text was shown for 2 min (120 s), immediately followed by a six-question questionnaire testing the participants' comprehension of the text. Answers were given verbally to the experimenter, who recorded them. A brief 2-min rest and recalibration interval was inserted after the third text, after which the lenses were switched. The duration of this interval was chosen to provide a short recovery from visual and cognitive load, and a clear separation between the two lens blocks, consistent with previous EEG and eye-tracking reading studies that have included short 1–2-min breaks between task blocks, to reduce fatigue and ensure block separation[22−24].
Preprocessing
Electroencephalogram (EEG)
-
We preprocessed the EEG data using the EEGLAB toolbox[25]. Continuous data was band-pass filtered to attenuate slow drifts and high-frequency noise, while preserving the frequency ranges analyzed in the spectral domain. Independent Component Analysis (ICA; runica) was then applied to decompose the data. To handle artifacts, we performed a semi-automated component rejection: IC's clearly associated with ocular (blinks, saccades), or muscle activity were identified for their scalp topographies, time-course characteristics, and power spectra, and were removed. The cleaned continuous signal was subsequently reconstructed for spectral power analysis.
Eye tracking
-
Gaze data was recorded with an integrated remote eye tracker (Tobii 120; 120 Hz). Viewing distance was fixed at 60 cm. Raw gaze coordinates were converted from pixels to degrees of visual angle using the display geometry (17'', 1,280 × 1,024); at 60 cm, 1 px(≈ 0.0252°).
Blinks were defined as consecutive missing/invalid pupil samples with durations of 1,000–1,100 ms, and were excluded from fixation and saccade detection. Fixations were identified using a combined spatial-temporal rule: dispersion radius 35 px (≈ 0.88°) and duration 100–600 ms. These choices are consistent with dispersion-based windows on the order of 0.5–1° and ≥ 100 ms minimum durations described in established identification taxonomies.
Saccades were detected within a velocity-threshold identification (I-VT) framework[26] with additional pragmatic constraints: peak velocity ≥ 60 px/s (≈ 1.51°/s), acceleration ≥ 400 px/s2 (≈ 10.06°/s2), and minimum duration ≥ 5 ms. Given the device sampling (120 Hz; 8.33 ms/sample) events shorter than one sample cannot be registered; operationally the minimum is effectively ≥ 1 sample. We emphasize that the acceleration and minimum-duration constraints are implementation details added in our pipeline while remaining within the I-VT class defined in[26]. All unit conversion, blink masking, and event detection were completed prior to metric computation.
Data analysis
-
Measures were computed per text passage and then averaged within participants for each condition (BLF vs. control). Between-condition differences were tested using Wilcoxon signed-rank tests. Analyses and visualization were performed in Python (v3.9.13) and EEG preprocessing was conducted in EEGLAB.
Behavioral data
-
Accuracy was defined as the number of correct responses to the six comprehension questions per passage. For each condition, passage-level accuracies were averaged within participants across the three passages, yielding one behavioral score per condition and participant. Between-condition differences were assessed using Wilcoxon signed-rank tests.
EEG
-
To explore the topographical distribution of the lens-induced effects on neural oscillations, we analyzed brain activity for each of the 20 electrodes separately. Brain activity overall was assessed by computing average power in each of the five standard EEG frequency bands recovered from the frequency spectrum data (delta: 1–4 Hz, theta: 4–8 Hz, alpha: 8–13 Hz, beta: 13–30 Hz, and gamma: 30–80 Hz) using the ‘spectopo' utility function in EEGLAB. The absolute power values in each of the five bands were normalized to the total power in all frequency bands, and then averaged separately across the three texts for the BLF and the control condition. These values were then tested for differences using paired Wilcoxon non-parametric tests across participants for each channel to find out whether the BLF lenses had an effect on brain activity at that electrode location. Tests were run at an exploratory alpha-level of 0.05, as well as a more stringent Bonferroni-corrected alpha-level of 0.01 (corrected by the number of frequency bands that were tested).
Eye tracking
-
From the preprocessed event stream, nine predefined eye-movement measures were computed per passage: saccade count, saccade amplitude (px), inter-saccadic interval (ISI, ms), mean saccade velocity (px/ms), saccade acceleration (px/s2), fixation count, fixation duration (ms), fixation dispersion (px), and blink count. The metric previously labeled 'saccade duration' reflects the inter-saccadic interval (ISI) returned by the detection pipeline. Mean saccade velocity was defined as amplitude/duration. For each condition, measures were averaged across the three texts within participants. Between-condition differences were assessed using Wilcoxon signed-rank tests. When multiple measures were analyzed, multiplicity was controlled using the Benjamini-Hochberg FDR (q = 0.05).
-
Participant demographics and baseline refractive and binocular characteristics are summarized in Table 1.
Table 1. Demographic characteristics of participants.
Baseline characteristics Participants (n = 34) Demographics Age, years 23.5 ± 1.88 Male sex 24 (70.6%) Spherical equivalent refractive errors (Diopters) Right eye −0.51 ± 0.26 Left eye −0.38 ± 0.89 Best-corrected visual acuity (logMAR) Both eyes 0.1 ± 0.1 Right eye 0.1 ± 0.1 Left eye 0.1 ± 0.1 Binocular function (Titmus) Normal stereoacuity
(< 50 log arcsec)34 (100%) Distance binocular sensory status (Worth 4 Dots) Fusion 34 (100%) Diplopia 0 (0%) Suppression 0 (0%) Behavioral data
-
In responding to the comprehension questions based on the readings, the participants exhibited no statistically significant differences in performance between the BLF and control lens conditions. On average, participants correctly responded to 14.82 questions out of 18 total (82.35% correct). Performance in the BLF condition was slightly higher with 15.03 questions out of 18 total (83.50% correct). The statistical test, however, found no evidence for a statistically significant difference with W = 269, p = 0.68 at our sample size. In summary, the analysis of the comprehension questions did not reveal statistically reliable behavioral differences between lens conditions at this sample size.
Eye tracking
-
Analysis from the eye-tracking data revealed that five out of the nine metrics showed significant differences (Fig. 4). Specifically, wearing the BLF lenses was associated with a pattern of fewer, but longer and faster, saccades. The average number of saccades per text was significantly lower when participants wore the BLF lenses (mean [SD] = 264.87 [114.08]), compared to the control lenses (mean [SD] = 302.76 [113.16]; W = 17, p < 0.001). Correspondingly, the average saccade length increased significantly from 76.86 (32.99) px with control lenses to 88.12 (35.40) px with BLF lenses (W = 56, p < 0.001). The inter-saccadic interval increased from 232.37 (82.07) ms, to 275.80 (130.51) ms (W = 26, p < 0.001). A slight but significant increase was also observed in saccade velocity, from 0.31 (0.06) px/ms in the control condition, to 0.32 (0.06) px/ms in the BLF condition (equivalent to approximately 310 px/s and 320 px/s, respectively; p < 0.05).
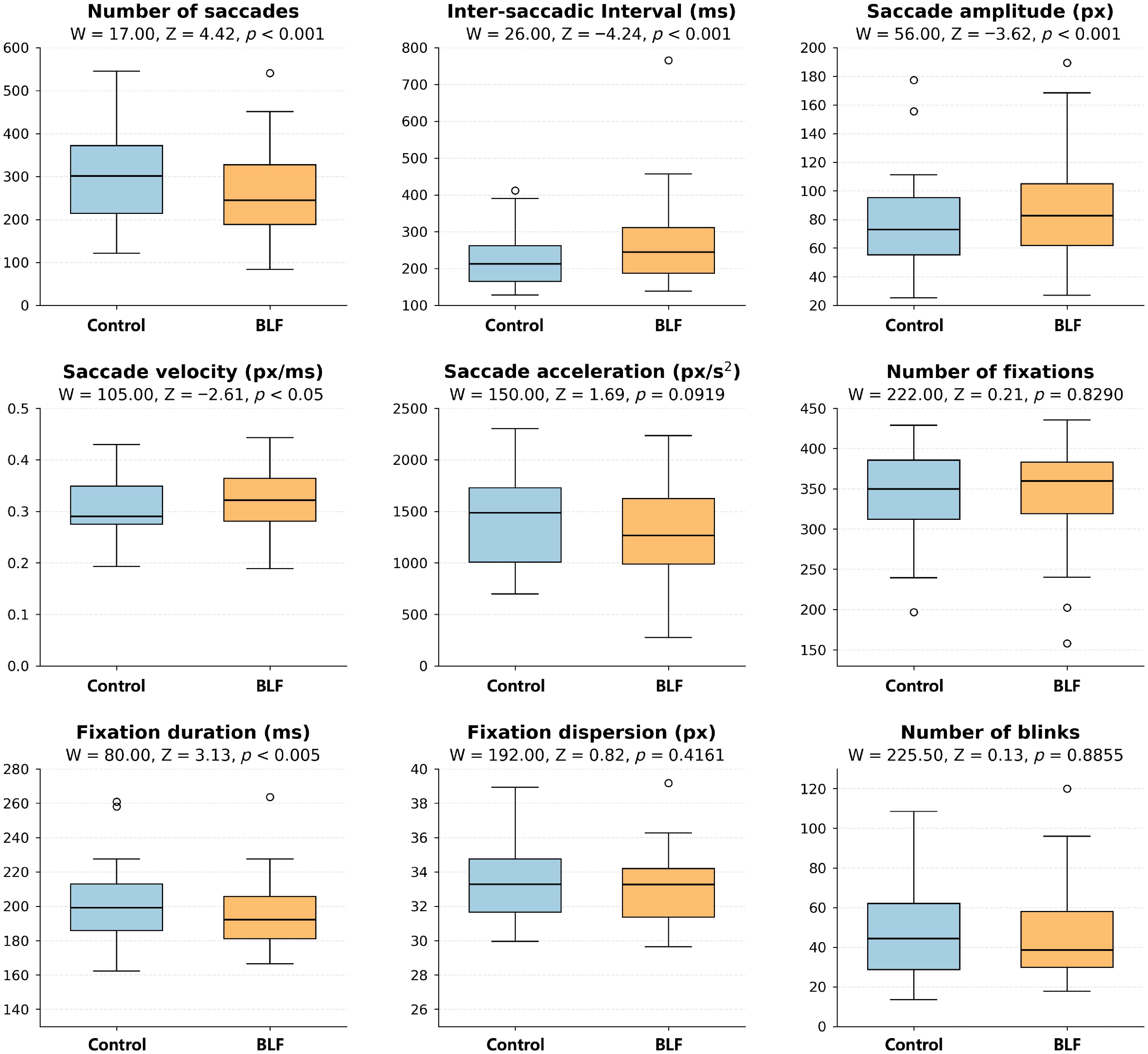
Figure 4.
Box plots of the nine metrics for control and filtered conditions together with associated p-values of non-parametric tests comparing the two conditions. For saccades, the BLF condition yielded fewer saccades, slightly higher mean velocity (px/ms), longer inter-saccadic interval (ms), and larger amplitudes (px); fixation duration was also reduced in the filtered condition. Acceleration is reported in px/s2.
In addition to saccade metrics, fixation duration was also affected. On average, fixation duration was 6.35 ms shorter in the BLF condition (mean [SD] = 194.76 [20.50] ms), compared to the control condition (mean [SD] = 201.11 [22.44] ms; W = 80, p < 0.005). Hence, out of the nine eye-tracking metrics tested, four saccade-based metrics showed potential influences from BLF, as well as one fixation-based metric.
Electroencephalogram (EEG)
-
To investigate the differences in brain oscillation for the BLF and non-filtered control conditions, we recorded the scalp electroencephalogram during the reading task. Power spectral analysis of the EEG during the reading interval was done separately for each electrode, and the standard frequency bands of delta (0 < δ < 4 Hz), theta (4 < θ < 8 Hz), alpha (8 < α < 13 Hz), beta (13 < β < 30 Hz), and gamma (30 < γ < 80 Hz). All electrodes that showed a significant change in power between the two conditions are listed in Table 2.
Table 2. Significant differences in band power for each electrode, in which the BLF condition differed from the control condition (p < 0.05 in Wilcoxon tests).
Frequency band Electrode Change of
BLF conditionStatistics Alpha F3 19% lower W = 154, p = 0.023 Alpha F4 26% lower W = 121, p = 0.004* Alpha C3 30% lower W = 153, p = 0.022 Alpha O2 5% lower W = 164, p = 0.037 Alpha F7 39% lower W = 122, p = 0.004* Alpha F8 23% lower W = 109, p = 0.002* Alpha FCz 22% lower W = 143, p = 0.013 Alpha Cz 17% lower W = 129, p = 0.006* Alpha Pz 2% lower W = 169, p = 0.046 Beta F3 23% lower W = 146, p = 0.015 Beta F4 26% lower W = 141, p = 0.012 Beta C3 33% lower W = 169, p = 0.046 Beta F7 38% lower W = 149, p = 0.018 Beta T8 24% lower W = 146, p = 0.015 Beta FCz 28% lower W = 159, p = 0.029 Gamma F3 21% lower W = 116, p = 0.003* Gamma C3 20% lower W = 148, p = 0.017 Gamma C4 18% lower W = 144, p = 0.014 Gamma F7 15% lower W = 156, p = 0.025 Gamma P8 18% higher W = 160, p = 0.031 Gamma FCz 23% lower W = 145, p = 0.014 Delta F3 5% higher W = 407, p = 0.023 Delta F4 4% higher W = 432, p = 0.006* Delta C3 5% higher W = 417, p = 0.014 Delta F7 5% higher W = 438, p = 0.004* Delta T8 7% higher W = 431, p = 0.006* Delta FCz 4% higher W = 418, p = 0.013 Theta F7 33% lower W = 154, p = 0.023 Theta T8 3% lower W = 156, p = 0.025 The table lists the frequency band, the electrode position in the 10−20 system, the change in power (in percent) compared to the control condition, and the detailed result of the Wilcoxon test. Starts mark the tests that pass a more stringent, Bonferroni corrected test. Bold values indicate statistically significant results (p < 0.05). In detail, we found a large number of electrodes that showed substantial decreases in alpha band power: these included four frontal, two central, one parietal, one temporal, and one occipital electrode. These sites all had lower alpha power in the BLF condition compared to the control condition. For the beta band, we found decreases in frontal and central electrodes, as well as one temporal electrode. Similarly, gamma power was reduced for frontal and central electrodes, with one increase in a parietal electrode. In contrast, we found increases for the delta band in three frontal, two central (C3), and one parietal electrode in the BLF condition. Finally, we found again reduced power for one frontal and one temporal electrode in the theta band.
With a more stringent testing criterion, we still see a significant reduction in alpha power for two frontal, one temporal, and one central electrode, a reduction in gamma power for one frontal electrode, and increases in delta power for two frontal and one temporal electrode.
Despite small performance differences, we therefore found several significant differences in sites in the EEG data, which suggest effects of the BLF lenses on brain activity. The majority of these sites include frontal areas, but for both alpha and delta bands, we also found other sites to be affected by the lenses compared to the control condition.
-
The present study utilized a multimodal approach, recording simultaneous EEG and eye-movement measures to analyze neurophysiological changes during a display-based reading task. The experiment compared the effects of wearing BLF lenses against non-filtering placebo (control) lenses. A clear dissociation emerged: short-term wearing of BLF lenses induced statistically significant alterations in both eye-movement patterns and brainwave activity, yet these physiological changes were not associated with any meaningful improvement in reading-comprehension performance. The paradigm targeted acute load during brief, timed reading, and the effects were detectable within a two-minute window; generalization to prolonged wear or day-long reading was not addressed.
Notably, the changes in eye-movement patterns often labeled as 'efficient' did not lead to an actual improvement in reading-comprehension. We interpret this constellation as compensatory rather than efficient. The results demonstrate that such metrics are not universally indicative of enhanced performance. These changes are more appropriately interpreted as a compensatory visual scanning strategy in response to the altered visual environment[27]. The brain may be adapting to the altered color and luminance profile of the filtered lenses by adopting a more deliberate scanning pattern to ensure equivalent information intake, rather than processing the text more efficiently, a finding consistent with studies on compensatory eye movements during difficult reading[28,29].
The decrease in alpha power and increase in delta power, while often associated with increased attention, are also established markers of acute cognitive load during learning tasks[30−33]. Therefore, the EEG changes suggest adjustments in effort allocation to process the unfamiliar visual input. Taken together, fewer but longer, and slightly faster saccades alongside spectral modulation align with a reallocation of effort to maintain performance under altered spectral input, indicating compensatory adjustment rather than enhancement[34−38]. Intrinsically photosensitive retinal ganglion cells (ipRGCs) are known to primarily mediate non-image-forming functions such as signaling environmental light, controlling pupil size, and entraining circadian rhythm, and they also contribute to visual detection, temporal processing, and color processing[39]. ipRGC's are maximally sensitive to short-wavelength light and light, and project both to non-image-forming centres (such as the suprachiasmatic nucleus and brainstem arousal systems), and to image-forming thalamocortical pathways, allowing short-wavelength-weighted input to modulated cortical excitability and background arousal during visual tasks[40−42]. Experimental manipulations of blue-enriched, or short-wavelength light have been associated with changes in occipital alpha and low-frequency power in parallel with altered alertness and cognitive performance[43,44]. In this context, the BLF-related EEG pattern observed here—modest reductions in alpha power together with localized increases in delta power at frontal and central sites—can be interpreted as a compensatory adjustment of the thalamocortical state to maintain performance under altered spectral input, rather than a marker of improved processing efficiency. Given that the present study did not include direct indices of ipRGC-driven input (e.g., melanopic weighted irradiance or post-illumination pupil response), the EEG alterations observed here should be considered compatible with ipRGC-mediated modulation, but cannot be regarded as ipRGC-specific.
Short-term wear of BLF lenses induces measurable changes in brain activity and eye-movement patterns, but these physiological adaptations do not translate into a tangible improvement in task performance (reading-comprehension accuracy) in the present sample[45−47]. Our findings align with this trend. Several systematic reviews and placebo-controlled trials have found insufficient evidence to support claims that these lenses alleviate visual fatigue or improve cognitive performance during short-term computer use[13,48]. The dissociation between physiological adaptation and unchanged reading comprehension should be interpreted in light of the participants profile. Participants were healthy university students in their twenties with normal or corrected-to-normal visual acuity and binocular function. Most participants were accustomed to intensive near work and digital reading in an academic context. In such readers, oculomotor and EEG responses are likely tuned for fast, goal-directed text processing. BLF-related changes may therefore reflect a reallocation of effort within a robust visual system, rather than removal of a limiting bottleneck. In routine practice, an important target group for BLF spectacles is working-age adults who perform prolonged screen-based tasks in office environments. Occupational visual demands, cumulative digital eye strain, and sex-related differences in susceptibility to visual symptoms may alter the balance between physiological adaptation and any perceived benefit in working-age office workers. Future studies with more diverse samples, particularly office workers with high screen exposure, are needed to test the hypothesis that occupational visual demands and sex-related vulnerability modulate the relationship between physiological adaptation and perceived benefit.
Multimodal assays serve as sensitive probes of effort allocation in time-constrained reading, revealing adjustments that coarse behavioral scores miss[18,49]. The present pattern cautions against equating physiological difference with functional benefit in lens evaluation. Within acute-load contexts, interpretive weight rests on converging multimodal evidence rather than isolated metrics[17].
Limitations
-
The paradigm targeted acute load during brief, timed reading and does not address prolonged wear, or day-long reading. Passages were standardized but limited in number, and testing occurred under a single, controlled level of ambient illumination using one display configuration. Behavioral outcomes were restricted to reading-comprehension accuracy; endpoints such as reading speed and fluency were not assessed.
EEG inferences were based on relative band-power summaries without explicit aperiodic-periodic separation. Electrode-wise comparisons were exploratory; a Bonferroni threshold was reported across the five frequency bands within each electrode, and no correction was applied across electrodes. These choices constrain the scope of topographical claims.
Eye-movement event detection was expressed in degrees of visual angle and constrained by the 120 Hz sampling rate, which sets the effective minimum event duration. Finally, individual spectral transmittance of the BLF lenses (including melanopic weighting) was not quantified, limiting inferences about person-specific spectral drive under BLF.
-
Short-term wear of BLF lenses altered oculomotor and EEG indices during brief, timed reading, without accompanying gains in reading-comprehension accuracy. The data indicate compensatory, load-related adjustments under altered spectral input rather than performance enhancement. This dissociation constrains claims about cognitive benefits during reading and highlights the value of acute-load paradigms for revealing adjustments that behavioral scores alone may miss.
Generalization to prolonged wear, varied illumination, and broader reading outcomes remain to be established.
-
The study protocol was conducted in accordance with the tenets of the Declaration of Helsinki, and all participants provided written informed consent after receiving a full explanation of the experimental procedures. The study was approved by the Internal Review Board (IRB) of Korea University (KU-IRB-15-205-A-1) on February 13, 2016. This study did not require clinical trial registration, as it did not involve a therapeutic intervention or outcomes classified as a clinical trial under applicable regulations.
-
The authors confirm their contributions to the paper as follows: conceptualization, visualization: Ryu H, Wallraven C; data curation, formal analysis, investigation, methodology, writing—original draft: Ryu H; project administration, validation: Wallraven C; writing—review and editing, funding acquisition: Wallraven C, Kim J; supervision: Wallraven C, Kim J. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are not publicly available because they contain information that could compromise the privacy of research participants, but are available from the corresponding author upon reasonable request.
-
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (RS-2025-22902971).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ryu H, Wallraven C, Kim J. 2026. A multimodal analysis of eye-movement and EEG responses during short-term blue-light filtering lens wear in a reading task. Visual Neuroscience 43: e008 doi: 10.48130/vns-0026-0006 

A multimodal analysis of eye-movement and EEG responses during short-term blue-light filtering lens wear in a reading task
- Received: 09 September 2025
- Revised: 12 December 2025
- Accepted: 23 December 2025
- Published online: 12 March 2026
Abstract: Blue light filtering (BLF) lenses are widely marketed to enhance cognitive function during digital device use, yet objective evidence for their immediate effects remains limited. We investigated the immediate neurophysiological impact of short-term wear by comparing BLF lenses to non-filtering placebo lenses in a reading task. For a within-subjects experiment, 34 participants read texts for a total of 12 min while electroencephalogram and eye-tracking data were recorded for both lens conditions in a counter-balanced order. No significant difference in reading comprehension performance was found between the two conditions within this sample. Wearing the BLF lenses, however, induced significant physiological changes: eye-tracking revealed fewer, longer saccades and decreased fixation duration, while EEG showed decreased alpha and increased delta power. The present findings suggest that the physiological changes induced by BLF lenses did not result in any immediate behavioral or cognitive improvements.
-
Key words:
- Blue light filtering lenses /
- Electroencephalogram /
- Eye movements /
- Reading performance