








-
Low-temperature (LT) stress stands as a pivotal environmental limiting factor that dictates the geographical distribution of plants and agricultural productivity. From 1975 to 2023, cumulative global economic losses from extreme climate reached 69 billion USD[1]; horticultural crops—representing 35% of global agricultural output (7 billion tons) in 2017—are especially vulnerable to such yield-limiting stress[1]. To counter this challenge, plants have evolved an intricate set of adaptive strategies aimed at mitigating LT stress while, to the greatest possible extent, sustaining the processes of growth and development. This delicate balance is not a simple 'on-off' switch, but rather an actively adaptive process dynamically regulated by a multi-tiered signaling network, commonly referred to as the 'growth and tolerance trade-off'. When plants perceive LT through alterations in plasma membrane fluidity and calcium signaling, they promptly initiate a comprehensive cold-response cascade. This encompasses the accumulation of compatible solutes (such as proline), the expression of cold-regulated proteins, and the remodeling of membrane lipid composition to maintain cellular homeostasis[1−3]. The activation of this defensive program consumes substantial amounts of energy, inevitably inhibiting growth processes such as cell division and elongation. Plants therefore face a core dilemma in resource allocation: how to optimally partition limited resources between defense and growth? Evidence indicates that this trade-off is not passive but is centrally coordinated through an intricate endogenous hormone network.
Early research tended to categorize hormone functions simply as promoting growth (e.g., auxin, gibberellin [GA]), or mediating stress responses (e.g., ABA, jasmonic acid [JA]). However, this 'one hormone, one function' linear model has been revised by modern evidence. For instance, LT promotes the accumulation of abscisic acid (ABA), which orchestrates a suite of adaptive responses, including the activation of LT defense mechanisms and the induction of seed dormancy[4]. Concurrently, elevated ABA levels inhibit seed germination and cause pollen abortion in plants[4,5]. Similarly, LT affects the synthesis and transport of auxin, thereby inhibiting root growth and morphogenesis. However, a moderate reduction in auxin levels can, conversely, contribute to enhancing the overall cold tolerance of plants[6−8]. A growing body of research indicates that hormones can simultaneously participate in stress responses and growth balance, with a complex network of hormonal crosstalk, serving as the core for achieving a regulatable balance. Despite a profound understanding of the interactions among hormones, a core question remains incompletely elucidated: how do plants, upon perceiving LT signals, integrate multiple hormonal signals into a holistic resource allocation decision-making system, thereby achieving the optimal solution for survival and reproduction in dynamic environments? This signal integration mechanism is pivotal for understanding plant environmental adaptability, and also provides potential targets for crop cold tolerance breeding.
This review systematically elucidates the molecular pathways of LT signal perception and transduction, analyzing LT impacts from cellular to tissue levels to provide an integrated understanding of plant adaptation. It particularly highlights the hormonal network's role in balancing growth and cold-tolerance defense. Focusing on key hormones—ABA, auxin, GA, ethylene (ET), cytokinin (CK), brassinosteroid (BR), salicylic acid (SA), strigolactone (SL), and JA—the review examines how they integrate LT-responsive, growth-related, and stress-resistance factors, and interact with one another to form a coordinated regulatory network. This network orchestrates resource allocation, managing the growth-cold tolerance trade-off to ensure optimal adaptation. Finally, based on the dynamic equilibrium of LT-triggered hormone signaling, we outline prospective applications and key research directions for genetically improving crop cold tolerance.
-
Plants respond to LT stress through a highly coordinated molecular network, the core of which lies in the precise perception and transduction of cold signals. Perception occurs primarily at the plasma membrane: LT induces a phase transition of membrane lipids from the liquid-crystalline state to the gel state, reducing membrane fluidity, which in turn activates plasma membrane-localized calcium channels, causing a transient increase in cytosolic calcium ion concentration ([Ca2+]cyt) that constitutes one of the earliest cold signaling events[2,9]. Recent research indicates that specific plasma membrane ion channels and transport proteins play key roles in cold-induced calcium signaling: Arabidopsis mechanosensitive channel of small conductance Mid1-COMPLEMENTING ACTIVITY (MCA1/2), ANNEXIN 1 (ANN1), as well as rice CYCLIC NUCLEOTIDE-GATED CHANNEL 9 (OsCNGC9) have been demonstrated to mediate the cold shock-induced rise in cytosolic Ca2+ concentration[1−3]. The rice CHILLING TOLERANCE DIVERGENCE 1 (COLD1) protein, which is dual-localized to both the plasma membrane and the endoplasmic reticulum, functions as a G-protein regulator. By interacting with the Gα sub-unit RGA1, it enhances the GTPase activity of RGA1 and collaboratively activates calcium channels to initiate LT signaling[10]. In addition, the plasma membrane-localized protein complex COLD6-OSM1 perceives cold signals, triggers the accumulation of the novel intracellular second messenger 2',3'-cAMP, and thereby activates cold defense responses[11]. The plasma membrane-localized calcium-dependent protein kinase CALCIUM-DEPENDENT PROTEIN KINASE 28 (CPK28) is rapidly activated by cold shock in a Ca2+-dependent manner, subsequently phosphorylating the cytosolic NIN-LIKE PROTEIN 7 (NLP7) and inducing its nuclear translocation. Within the nucleus, NLP7 directly binds to, and activates the promoters of cold-regulated genes such as C-REPEAT BINDING FACTOR (CBF), driving the expression of downstream COLD-REGULATED (COR) genes, thus forming a crucial Ca2+ signal perception-decoding module[2,9].
In Arabidopsis, the calcium/calmodulin (Ca2+/CaM) signaling pathway plays a central role in LT signal transduction and the establishment of cold tolerance by regulating two key MITOGEN-ACTIVATED PROTEIN KINASE (MAPK) cascades. Research shows that Ca2+/CaM-MEDIATED RECEPTOR-LIKE CYTOPLASMIC KINASES (CRLK1) and CRLK2 negatively regulate cold-induced MPK3/MPK6 activity through a dual mechanism: they can directly inhibit the phosphorylation and activation of MPK3/MPK6, and indirectly suppress MPK3/MPK6 by activating the upstream MEKK1-MKK2-MPK4 cascade[2,12,13]. Notably, MAPK cascades exhibit antagonistic functions in the cold response: the MKK4/MKK5-MPK3/MPK6 module negatively regulates plant cold tolerance, whereas the MEKK1/MEKK2-MKK2-MPK4 module positively regulates the cold response by inhibiting MPK3/MPK6 activation. Acting as an upstream regulatory node, CRLK1/2 integrates calcium signals and coordinates the activity of these two downstream MAPK cascades, precisely regulating the phosphorylation signaling network and gene expression reprogramming in cells under LT stress, ultimately enhancing plant cold tolerance[2,12,13]. These findings reveal that plants employ a multi-tiered signaling network—spanning from membrane receptor perception and calcium signal decoding, to kinase cascade regulation and transcriptional reprogramming—to precisely perceive and transduce early cold signals and initiate efficient adaptive physiological responses.
Following perception at the plasma membrane, the LT signal is relayed via calcium signaling and MAPK cascades to transcriptional regulatory hubs. Among these, the ICE-CBF-CORs transcriptional cascade constitutes the core pathway of the cold response network. In Arabidopsis, the MYC-type bHLH transcription factor INDUCER OF CBF EXPRESSION 1 (ICE1) acts as a specific activator of CBF gene, and its activity is regulated by multiple post-translational modifications: HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE 1 (HOS1), PLANT U-BOX 25 (PUB25), and PUB26 mediate its ubiquitination and degradation; SAP AND MIZ1 DOMAIN-CONTAINING LIGASE 1 (SIZ1) stabilizes ICE1 via SUMOylation, while the kinases OPEN STOMATA 1 (OST1), MPK3/6, and BRASSINOSTEROID INSENSITIVE 2 (BIN2) modulate its transcriptional activity through phosphorylation[1−3,14] (Fig. 1). The active form of ICE1 directly binds to the MYC recognition elements in the CBF promoters, activating the expression of CBF1, CBF2, and CBF3[1−3].
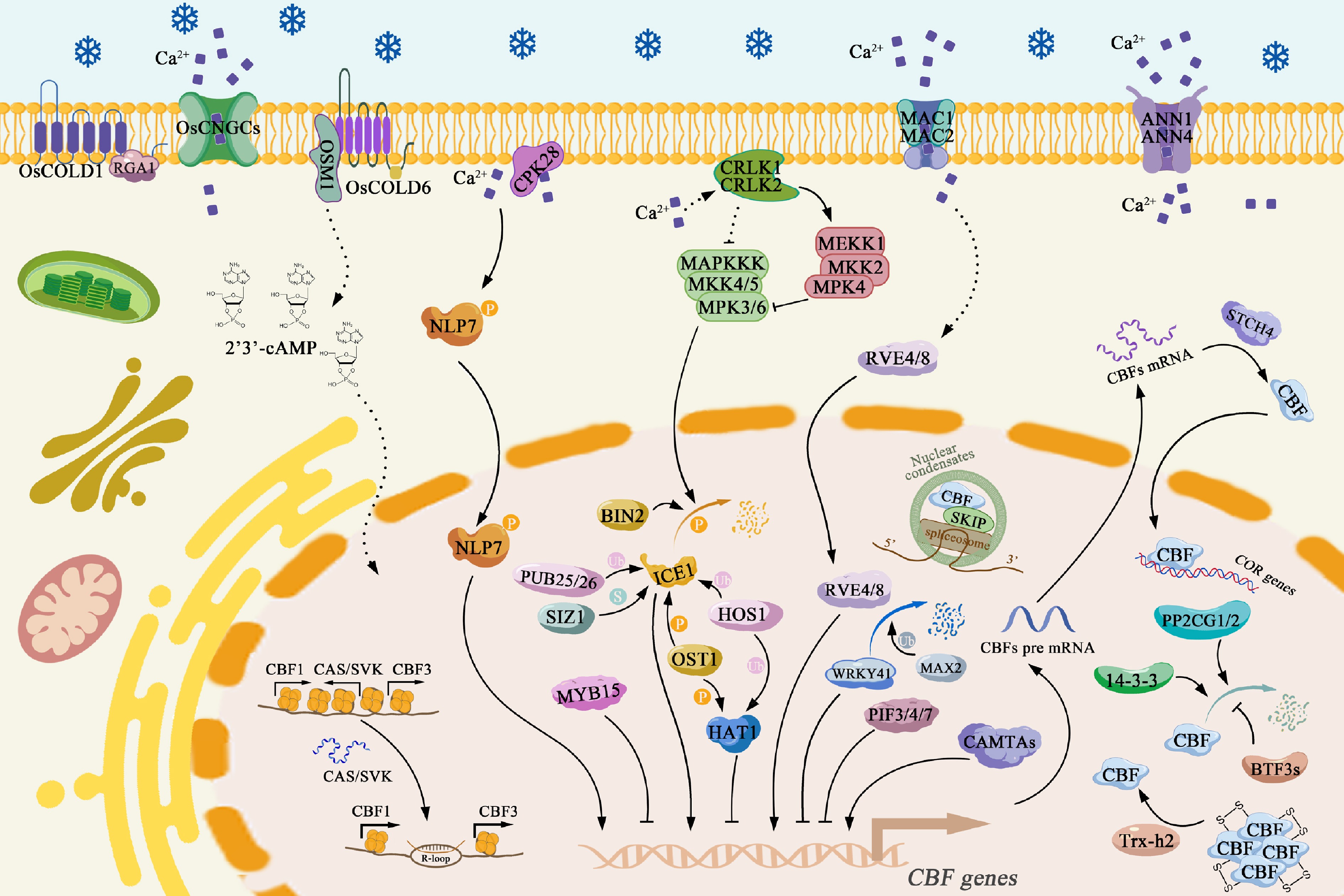
Figure 1.
Model of LT perception and signaling transduction in plants. Proteins such as MCA1/2, ANN1/4, CPK28, CRLK1/2, MKK4/MKK5-MPK3/MPK6, and MEKK1-MKK2–MPK4 in Arabidopsis, as well as OsCNGC9, COLD1, and COLD6-OSM1 in rice, perceive LT stress and regulate Ca2+ influx, thereby transmitting the cold signal into the nucleus. ICE1 protein activity is post-translationally modulated by BIN2, MPK3/6, HOS1, SIZ1, OST1, and PUB25/26, which in turn affects its regulatory function in cold signaling. Transcription of CBF genes is controlled by NLP7, HAT1, ICE1, MYB15, WRKY41, RVE4/8, PIF3/4/7, CAMTAs, and CAS/SVK. CBF proteins' stability, translation, and functional activity are influenced by STCH4, PP2CG1/2, 14-3-3 proteins, BTF3s, and Trx-h2, which affect their DNA binding to COR genes or splicing regulators, such as SKIP. Arrows indicate positive regulation, bar-headed lines represent negative regulation; solid lines denote direct interactions, and dashed lines represent indirect regulation. Abbreviations: MCA, Mid1-COMPLEMENTING ACTIVITY; ANN1/4, ANNEXIN1/4; CPK28, CALCIUM-DEPENDENT PROTEIN KINASE 28; CRLK1/2, Ca2+/CaM-REGULATED RECEPTOR-LIKE KINASE 1/2; MEKK1, MPK KINASE KINASE 1; MKK, MPK KINASE; MPK, MITOGEN-ACTIVATED PROTEIN KINASE; OsCNGC9, CYCLIC NUCLEOTIDE-GATED CHANNEL 9; COLD1/6, CHILLING TOLERANCE DIVERGENCE 1/6; OSM1, OSMOTIN-LIKE 1; ICE1, INDUCER OF CBF EXPRESSION 1; BIN2, BRASSINOSTEROID INSENSITIVE 2; HOS1, HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE 1; SIZ1, SAP AND MIZ1 DOMAIN-CONTAINING LIGASE 1; OST1, OPEN STOMATA 1; PUB25/26, PLANT U-BOX 25/26; CBF, C-REPEAT BINDING FACTOR; NLP7, NIN-LIKE PROTEIN 7; HAT1, HOMEODOMAIN-ASSOCIATED TRANSCRIPTION FACTOR 1; MYB15, MYB DOMAIN PROTEIN 15; WRKY41, WRKY DNA-BINDING PROTEIN 41; RVE4/8, REVEILLE 4/8; PIF3/4/7, PHYTOCHROME-INTERACTING FACTOR 3/4/7; CAMTAs, CALMODULIN-BINDING TRANSCRIPTION ACTIVATORs; CAS, CBF ANTISENSE TRANSCRIPT; SVK, SVALKA; STCH4, SENSITIVE TO CHILLING 4; PP2CG1/2, PROTEIN PHOSPHATASE 2C G 1/2; BTF3s, BASIC TRANSCRIPTION FACTOR 3s; trx-h2, thioredoxin h2; COR, COLD-REGULATED; SKIP, SnRNA KINASE INTERACTING PROTEIN; P, phosphorylation; Ub, ubiquitination; S, SUMOylation. This figure was created with templates from
https://BioGDP.com .CBF proteins, belonging to the AP2/ERF transcription factor family (also known as DEHYDRATION-RESPONSIVE ELEMENT-BINDING PROTEIN 1s/DREB1s), are master regulators of cold acclimation in higher plants. They specifically recognize the CRT/DRE cis-element (core sequence: CCGAC) in the promoters of COR genes and activate their expression by recruiting co-activators or chromatin remodeling complexes[1−3]. The expression and function of the CBF gene is governed by a multi-layered and intricate regulatory network during LT stress. At the transcriptional level, besides ICE1, they are regulated by various other transcription factors, such as NLP7, MYB DOMAIN PROTEIN 15 (MYB15), CALMODULIN-BINDING TRANSCRIPTION ACTIVATOR 3 (CAMTA3), and so on[2,9,15]. Within the photoperiod signaling pathway, PHYTOCHROME-INTERACTING FACTOR 3 (PIF3), PIF4, and PIF7 suppress CBF expression under dark or LT conditions by directly binding to G-box or E-box motifs in their promoters[16]. Conversely, under LT stress, CBFs interact with PIF3, disrupting its binding to the photoreceptor Phytochrome B (phyB). The stabilized phyB then acts downstream of CBFs to positively enhance cold tolerance by regulating stress-responsive and growth-related genes[17]. Furthermore, the circadian clock imposes temporal regulation on CBF expression through clock-associated transcription factors: the PSEUDO-RESPONSE REGULATORS5/7/9 (PRR5/7/9) negatively regulate CBF expression and cold tolerance[18], whereas the clock-related MYB transcription factors RVE4 and RVE8 translocate from the cytoplasm to the nucleus upon cold exposure, directly binding the CBF3 promoter to induce its expression[15]. The roles of other clock components like CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) in CBF regulation are also noted, though their specific mechanisms remain somewhat controversial[15,19]. Recent studies have revealed additional regulatory layers. Cold triggers OST1-mediated phosphorylation of HOMEODOMAIN-ASSOCIATED TRANSCRIPTION FACTOR 1 (HAT1), promoting its interaction with HOS1, leading to HAT1 ubiquitination and degradation. This process relieves HAT1's repression on CBF and COR genes, thereby activating the cold response[1]. At the post-transcriptional level, cold-induced antisense non-coding RNAs (CAS) derived from CBF1 and CBF3 loci facilitate CBF expression by forming R-loop structures that reduce nucleosome density[20]. Post-translational modifications and protein stability are also critical: factors like 14-3-3 PROTEINS, BASIC TRANSCRIPTION FACTOR 3/BTF3-LIKE PROTEIN (BTF3/BTF3L), PROTEIN PHOSPHATASE 2C G GROUP 1 (PP2CG1), and PP2CG2 modulate CBFs' stability under LT stress[1−3,15]. The nuclear-localized desulfuration Thioredoxin h2 (Trx-h2) reduces oxidized, inactive CBF1 polymers into active monomers, enabling the activation of CORs[21]. The ribosome biogenesis factor STCH4 (encoded by the cold-induced TCH4/REIL2) enhances CBF translation by maintaining rRNA processing efficiency[22]. The F-box E3 ubiquitin ligase MAX2 mediates the degradation of the transcriptional repressor WRKY DNA-BINDING PROTEIN 41 (WRKY41), thereby de-repressing CBF expression[14]. Furthermore, CBFs interact with the spliceosome core component SnRNA KINASE INTERACTING PROTEIN (SKIP), promoting the formation of SKIP nuclear condensates and enhancing the splicing efficiency of cold-responsive transcripts[23]. Multitiered regulation of CBF gene, from transcription to post-translational control of protein stability enables their precise induction and function in cold acclimation. This finely tuned regulatory network is evolutionarily conserved; phylogenetic analyses show that the CBF transcriptional module is shared between monocots and dicots and has been identified in key crops such as rice, maize, wheat, barley, tomato, and rapeseed[2]. Functional studies confirm that heterologous or endogenous overexpression of CBF gene consistently activates downstream CORs expression and enhances freezing tolerance across these species, underscoring the central role of the CBF pathway in plant adaptation to LT stress (Fig. 1).
In summary, plants employ a multi-tiered perception and regulatory system to respond precisely to LT stress. This system spans from primary perception (e.g., membrane fluidity changes) to rapid signal transduction (e.g., Ca2+, ROS bursts), and extends to fine-tuning at the levels of transcription factors (ICE1, CBFs), epigenetic modifications, non-coding RNAs, and post-translational modifications. This ensures the timely activation of cold tolerance genes while maintaining growth balance, and enhances stress resilience through cross-pathway integration and inter-organellar coordination.
-
LT stress acts as a potent environmental signal that triggers a profound reprogramming of plant growth, prioritizing a survival-over-growth strategy. Under the ensuing energy and resource constraints, plants actively suppress growth to reallocate photoassimilates toward the establishment of cryoprotection. This chapter will systematically elaborate on the multi-level inhibitory effects of LT stress on plants, from microscopic to macroscopic scales, and from vegetative to reproductive growth, along with their molecular and physiological bases.
Cellular-level inhibition
-
The inhibitory effect of LT on plant growth initiates at the cellular level, primarily targeting the core processes of life maintenance and structural construction—cell division and expansion. The plasma membrane is the primary site of cold injury. Under LT stress, ice crystals formed in the apoplast can cause cellular dehydration and irreversible damage to membrane structure, subsequently triggering a series of cellular dysfunctions[24−27]. Membrane fluidity and cytoskeletal reorganization play roles in early cold signal transduction. LT induces a phase transition of membrane lipids from the liquid-crystalline to the gel state, reducing membrane fluidity, which leads to the malfunction of membrane proteins (e.g., H+-ATPase, ion channels, and transporters). This results in ion leakage, disruption of the transmembrane electrochemical gradient, and efflux of cellular contents, ultimately causing metabolic disorders and cell death[25−27].
Studies show that compared to plants grown under normal temperatures, seedlings subjected to 4 °C treatment exhibit no significant change in average cell length, but a marked rearrangement of the cell cycle progression: the proportion of cells in the G1 and G2 phases increase, while the proportion in the S phase decreases. Concurrently, expression of the G2-M phase-specific cell cycle marker CYCLIN B1;1 is significantly reduced, confirming the suppression of mitotic activity under LT stress. Furthermore, beyond directly causing cell cycle arrest, LT may also induce nucleic acid damage. Significant upregulation of DNA repair genes during cold acclimation in winter wheat indicates that while LT does not inhibit cell elongation, it can indirectly suppress cell proliferation by affecting the cell division process and genomic stability[7,28,29].
LT stress inhibits cell expansion through dual mechanisms involving water balance regulation and cell wall remodeling. Regarding water regulation, LT reduces the activity and expression of plasma membrane aquaporins, concurrently diminishing root water uptake capacity and xylem hydraulic conductivity. This leads to cellular water deficit and an inability to maintain the turgor pressure required for driving expansion[26,27,30]. In terms of the cell wall, significant metabolic and structural remodeling occurs: Expansins such as AnEXPA1 are upregulated, loosening the cellulose-hemicellulose network via non-hydrolytic mechanisms, whereas the activity of XYLOGLUCAN ENDOTRANSGLUCOSYLASES/HYDROLASES (XTHs) is inhibited, affecting the cross-linking and restructuring of cell wall polysaccharides[26,27,31]. Additionally, adaptive changes in cell wall composition include increased pectin content, enhanced wall strength, and reduced porosity[32]. Notably, WALL-ASSOCIATED KINASES (WAKs) are upregulated under LT stress and perceive cell wall integrity through interaction with oligogalacturonides or pectin, suggesting the cell wall itself may act as a sensing element for LT signals involved in the regulatory process[27,33]. In summary, LT coordinately suppresses normal cell division and expansion processes through direct impairment of membrane integrity, disruption of water balance, alteration of cell wall structure and plasticity, and effects on nuclear genetic material stability. These multifaceted impacts constitute the physiological basis for growth inhibition at the cellular level (Fig. 2).
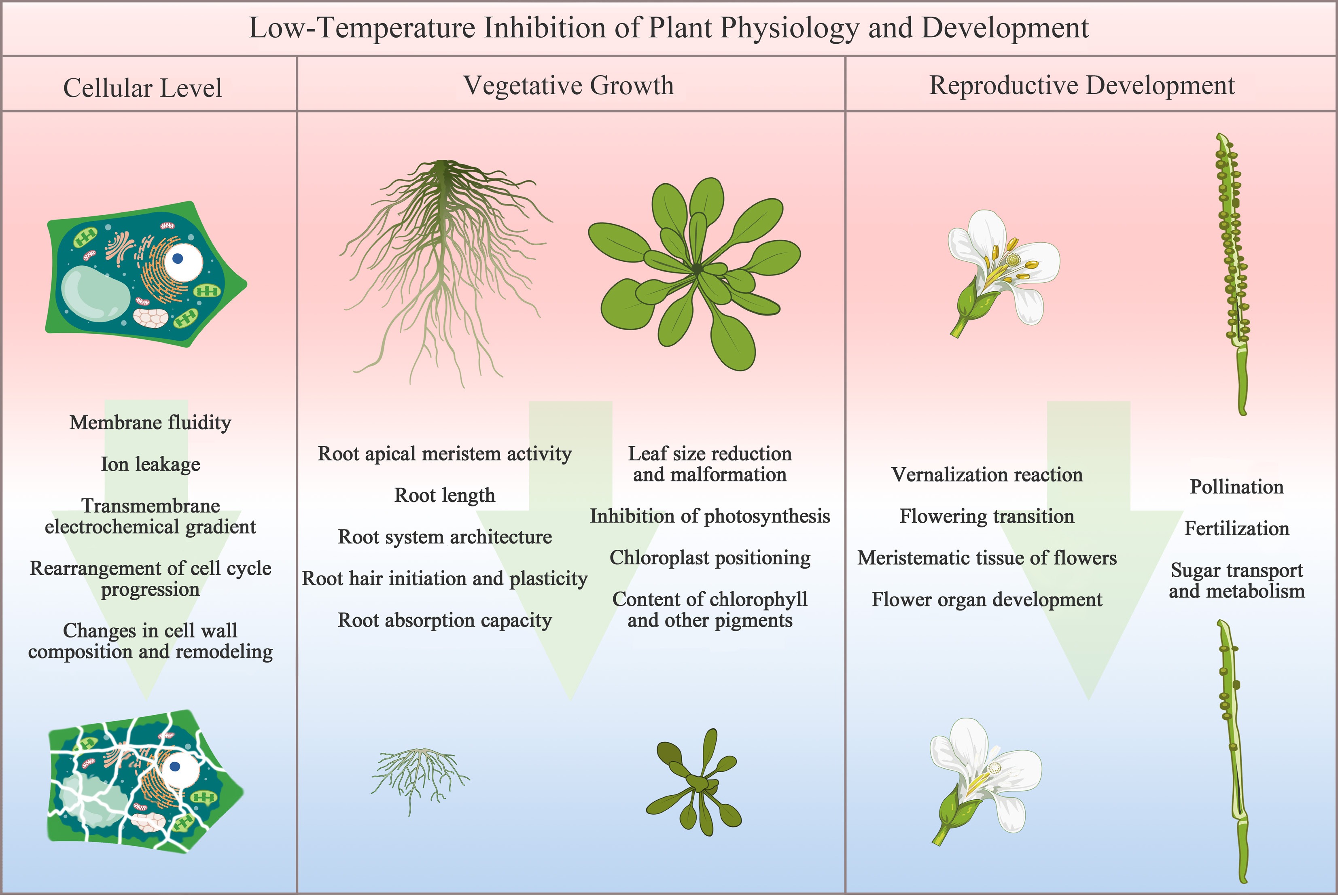
Figure 2.
Schematic representation of LT-induced inhibition at the cellular, vegetative, and reproductive levels in plants. At the cellular level, LT causes structural damage, including membrane dysfunction, cytoskeletal disorganization, and disruption of ion and water transport. Vegetative growth is suppressed through impaired leaf morphogenesis and photosynthetic performance, as well as restricted root system development and nutrient uptake capacity. Reproductive development is affected by altered flowering time, aberrant floral organ formation, compromised pollination and fertilization, and disrupted carbohydrate translocation, leading to a reduced fruit set. This figure was created with templates from
https://BioGDP.com .The overall slowdown in the growth of vegetative organs
-
Under LT stress, root growth in plants is significantly inhibited, with adaptive remodeling occurring in both morphological construction and physiological function. As the primary organ directly sensing the soil environment, roots are highly sensitive to LT: it initially suppresses meristematic activity in the root tip and disrupts the polar auxin transport and spatial distribution mediated by PIN-FORMED family proteins[34] (Fig. 2). This leads to abnormal gravitropic responses, inhibition of lateral root initiation and elongation, manifested as significant reductions in total root length, root volume, and root biomass, resulting in a simplified root architecture[35,36]. LT also directly affects the cell cycle progression, inhibiting cell division in the root meristem, which reduces the size of the meristematic zone and cell number, whereas cell elongation remains largely unaffected within a certain temperature range[6,7]. Furthermore, LT disrupts root hair initiation and plastic adjustment, reducing the root absorptive surface area and the efficiency of water and nutrient acquisition[37]. These morphological and cellular alterations collectively limit the plant's resource uptake capacity and, through source-sink relationships, exacerbate the inhibition of shoot growth, ultimately leading to reduced whole-plant biomass accumulation.
The functionality of leaves as photosynthetic organs is critically dependent on their proper development, a process severely compromised by LT. LT stress impedes leaf morphogenesis at its cellular basis by slowing both mitotic activity and cell expansion (Fig. 2). Consequently, plants exhibit typical stress-induced phenotypes, most notably a reduction in leaf area and lamina thickness, alongside morphological deformations like curling and wrinkling. Concurrently, LT suppresses photosynthesis through multiple mechanisms involving diffusional, metabolic, and photochemical limitations. Firstly, it induces stomatal closure, reducing CO2 diffusion into the mesophyll cells, and thereby directly limiting the substrate supply for photosynthetic carbon assimilation[3,38]. Secondly, it decreases the activity of key photosynthetic enzymes, such as Rubisco, and those involved in RuBP regeneration (e.g., SBPase, FBPase), which weakens the biochemical driving capacity of the photosynthetic reactions[39]. Thirdly, when combined with high light, LT causes damage to the Photosystem II (PSII) reaction centers. This occurs primarily because the degradation rate of the D1 protein surpasses its repair rate, leading to reduced photochemical efficiency (Fv/Fm) and increased photoinhibition[40]. Additionally, LT disrupts normal chloroplast positioning and function, inhibits chlorophyll synthesis and accelerates its degradation, leading to decreased total leaf chlorophyll content, impaired photosynthetic processes, and reduced accumulation of photosynthetic products, ultimately inhibiting overall plant growth[38,41]. The transcription factor CsCBF2 plays a key regulatory role in this process by repressing the expression of genes related to chlorophyll and carotenoid biosynthesis, thereby further aggravating the damage to leaf photosynthetic function under LT[42]. These cascading reactions ultimately lead to a reduction in photosynthate accumulation, which becomes a critical physiological bottleneck constraining overall plant growth.
Important influence on the reproductive growth process
-
To optimize the balance between survival and reproduction under LT stress, plants delay flowering via molecular mechanisms that integrate LT signals into the photoperiodic pathway. In rice, the thermosensor OsphyB stabilizes the flowering repressor OsGhd7 under LT, which suppresses the florigen genes OsHd3a and OsRFT1[43]. In soybean, LT upregulates GmCOL2b to enhance the repressor GmE1, forming a GmphyA-GmE1 module that coordinates LT and photoperiod signals to inhibit flowering. In Arabidopsis, the circadian clock component GIGANTEA (GI) acts as a key integrator; its stability is downregulated under LT through COP1/ELF3-dependent and HOS15-mediated pathways, leading to reduced expression of the florigen FT and thus delaying flowering[43].
LT precisely regulates flowering time and disrupts floral organ development through multi-layered molecular networks, with underlying mechanisms being progressively elucidated across multiple crop species. In chrysanthemum, LT activates the repressor CmMAF2 to suppress GA biosynthetic gene CmGA20ox1, downregulating CmLFY, and impairing floral primordium initiation[44]. Concurrently, LT-upregulated CmFLC-like represses CmFTL3 and CmAFL1, further delaying the floral transition[45]. In tomato, LT destabilizes the shoot apical meristem modules SlWUS-SlCLV3 and SlWUS-TAG1 and alters endogenous hormone balance by promoting ABA accumulation and reducing GA content, resulting in abnormal floral organ development[46]. In wheat, LT differentially regulates key TFs (e.g., TaMADS6, TaERF4, TaWOX6, TaNAC48), disrupting floret, ovule, and stamen morphogenesis[47]. Extreme LT can also trigger carbon-nitrogen metabolic imbalance, leading to floret degeneration[48] (Fig. 2). Epigenetically, H3K27me3 modification positively regulates LT-induced flowering in litchi[49], while BARENTSZ 1 (OsBTZ1)/OsBTZ2 maintains floral organ stability in rice, with defects exacerbated by LT stress[50].
LT stress critically affects pollen fertility and fruit set (Fig. 2). It disrupts anther structure and sugar transport, causing pollen sterility[51]. In rice, osmorn1/osmorn2 alter lipid metabolism and induce ROS accumulation under LT stress, reducing fertility[52]. Loss of PWIN1 disrupts sucrose metabolism in pollen and the SNF1-RELATED KINASE 1 (SnRK1)/TARGET OF RAPAMYCIN (TOR) signaling pathway, while its overexpression enhances cold tolerance[53]. The lipid transfer protein COLD TOLERANCE AT THE BOOTING STAGE 6 (OsCTB6) scavenges ROS to maintain tapetum development[54], and OsCTB5 interacts with OsHox12 to modulate OsEUI1, promoting the accumulation of active GA in the anthers[55,56]. During fruit set, LT-induced pollination failure and assimilate shortage cause fruit drop. The cold tolerance gene OsCTB3 in rice regulates sugar content via the OsTCP19 and OsTPP1 trehalose pathway, a key adaptation in japonica rice[57]. In tomato, LT inhibits assimilate transport; loss of sugar transporter SlSTP2 disrupts the arabinosylation of SlCLV3/SlWUS, causing multi-ovary formation[58].
-
As established, LT is a stringent environmental stressor that challenges plant cellular structure, physiological stability, and developmental continuity, leading to characteristic negative phenotypes such as altered membrane fluidity, reduced photosynthetic efficiency, and decreased growth rates[26,27]. Plants have evolved sophisticated adaptive strategies to cope with such stresses, with LT response being a critical mechanism. At the phenotypic level, cold-induced aberrations reflect internal physiological and developmental dysregulation, centrally governed by hormonal regulatory networks. These intrinsic chemical messengers perceive LT signals and activate downstream transduction cascades. By orchestrating gene expression, they systematically modulate cellular physiology and growth trajectories, mitigating LT stress and restoring homeostasis. This clear causal chain—from environmental signal to hormonal network to adaptive phenotype—highlights how hormone-mediated instructions determine plant acclimation strategies. Building on this linkage, the following sections will elucidate the dynamic balancing mechanisms within hormone signaling networks under LT. Through intricate cross-regulation, these networks establish an equilibrium between cold acclimation and normal growth, forming the core molecular basis for plant adaptation to LT environments.
Abscisic acid
-
Plants integrate LT and ABA signals through a sophisticated transcriptional network to balance cold resistance and growth. Under LT stress, ABA biosynthesis genes (e.g., NCED4) are upregulated, while ABA metabolic genes (e.g., CYP707A2) are suppressed, leading to ABA accumulation. This subsequently activates LT defense mechanisms, induces seed dormancy and inhibits germination[4]. During germination, ICE1 fine-tunes ABA signals by repressing ABA-responsive genes EM1, EM6, and ABI5[59]. During reproductive development, LT-elevated ABA in anthers disrupts pollen wall formation and sugar transport[51], causes abnormal m6A modification of genes like SlABCG31[5], and leads to pollen abortion. ABA also enhances cold tolerance by facilitating the interaction between its response factor MdABI4 and the core regulatory protein MdICE1, thereby boosting the expression of MdCBF1[60]. The C2H2-type zinc finger transcriptional repressor OsZOS2-19 responds to LT and ABA signals. Its overexpression impairs plant LT adaptability, osmotic balance, and ROS scavenging capacity, and negatively regulates stress pathways such as sugar and glutathione metabolism[61]. Conversely, Oryza sativa UBIQUITIN-CONJUGATING ENZYME 12 (OsUBC12) enhances the germination ability of japonica rice under LT conditions by promoting OsSnRK1.1 degradation, thereby weakening its activation of OsABI5[62]. LT also activates OsTPP1 via OsMYB30 and the OsbZIP23-OsTPP3 module through ABA accumulation mediated by OsNCED3, synergistically promoting trehalose to regulate germination sensitivity[63,64]. In tomato, SlERF15—a downstream ET component—also acts as a positive ABA regulator, binding and activating the expression of SlCBF1 and SlWRKY6, while modulating ABA-induced cold resistance, revealing a molecular bridge for ET and ABA signaling in regulating cold adaptation[65]. In summary, ABA acts as a dual-regulatory node in plant LT adaptation: it serves as both an early stress signal, and a central integrator that balances cold acclimation with growth. Following rapid accumulation, ABA drives broad transcriptional reprogramming, redirecting resources toward defense while concurrently restraining high-cost developmental processes like seed germination and pollen development. This strategic trade-off—short-term growth restraint for long-term viability—forms the physiological and molecular basis of the classic 'tolerate now, recover later' strategy under LT stress.
Auxin
-
Under normal development, auxin maintains the stem cell niche and organogenesis through genes such as WUS, PLT1, and PLT2, while precisely regulating the morphogenesis of leaves via components like IAA CARBOXYL METHYLTRANSFERASE 1 (IAMT1) and YUCCA[26,34,66]. However, LT stress reprograms auxin signaling. Cold-induced miR169 suppresses NF-YA2 binding to the YUC2 promoter, inhibiting auxin biosynthesis[67], while impaired GNOM-mediated PIN2 recycling disrupts basipetal transport, reducing root-apex auxin levels. This perturbs the cell cycle regulatory network (e.g., suppressed CYCDs and E2Fb, enhanced E2Fc and RBR), thereby inhibiting root growth and morphological construction. Disrupted polar auxin transport also weakens root gravitropic responses[6,7,34]. During reproduction, LT-reduced auxin in pollen leads to ROS and starch accumulation, ATP deficiency, and decreased germination[68]. For cold tolerance, INDOLE-3-ACETIC ACID-AMIDO SYNTHETASE (OsGH3-2) enhances rice cold-tolerance by converting active IAA into its inactive conjugated form[26,34,69]. Natural variations in the auxin efflux transporter OsHAN2 (OsABCB5) are linked to cold tolerance. The temperate japonica allele significantly enhances cold tolerance at the seedling and heading stages due to suppressed expression[70]. In apples, MdHY5 coordinates IAA and ABA signaling: LT weakens MdHY5-mediated repression of MdGH3-2/12 to reduce IAA content, while enhancing its activation of MdNCED2 to promote ABA accumulation. The lowered IAA/ABA ratio induces anthocyanin synthesis and improves adaptation[8]. Notably, moderate LT activates an adaptive pathway dependent on TAA1/YUCCA-mediated synthesis, PIN2/PIN4/AUX1-mediated polar transport, and TIR1/AFB2-ARF6/7/8 signaling, promoting root hair elongation[71]. This demonstrates auxin's ability to drive specific morphogenesis under LT conditions. In summary, under LT stress, auxin acts not merely as a growth promoter, but as a dynamic regulator of developmental programming. Through precise reprogramming of its biosynthesis, polar transport, and signal transduction, auxin undergoes spatiotemporal redistribution in response to LT. This reconfiguration restrains energy-consuming processes such as root elongation and pollen development, while promoting adaptive morphogenesis like root hair formation. Often coordinating with hormones such as ABA, auxin helps optimize resource allocation. Thus, its central role lies in integrating environmental cues to establish a dynamic balance between basal survival and adaptive development under LT stress.
Cytokinin
-
CK plays an essential role in plant growth and development, precisely controlling cell division via MYB3R4-dependent activation of mitotic genes and coordination with auxin to regulate leaf morphogenesis and apical dominance[34,66]. Under LT stress, CK signaling is reprogrammed: biosynthetic genes are downregulated, including ISOPENTENYLTRANSFERASE (IPT), tRNA ISOPENTENYLTRANSFERASE (TRIT1), CIS-ZEATIN O-GLUCOSYLTRANSFERASE (CISZOG), and D11, while the degradation gene CYTOKININ OXIDASE/DEHYDROGENASE (CKX) is upregulated[34,66,72]. Reduced CK levels in roots may help sustain basic growth, whereas elevated CK concentrates cold-induced root inhibition[34,73,74]. CK is transmitted through a phosphorelay involving ARABIDOPSIS HISTIDINE KINASE 2 (AHK2), AHK3, ARABIDOPSIS HISTIDINE PHOSPHOTRANSFER PROTEINS (AHPs), and RESPONSE REGULATORS (RRs)[73,74]. In Arabidopsis, AHK2 and AHK3 function as negative regulators of cold signaling, with the ahk2-ahk3 double mutant exhibiting enhanced cold tolerance[34,75]. Type-A RRs (e.g., ARR5, ARR6, ARR7, ARR15) are rapidly induced under LT. ARR7 negatively regulates cold tolerance in a CBF-independent manner[75], while overexpression of ARR5/7/15 can enhance cold tolerance, indicating context-dependent functionality[76]. Type-B RRs such as ARR1/12 primarily suppress root growth under LT stress by reducing auxin accumulation in the root tip[6]. Under LT stress, AHK2/AHK3 and type-A ARRs negatively regulate both cold tolerance and ABA responses[75]. In drought, ABA stabilizes ARR5 to amplify its signaling and inhibit growth, whereas CK antagonizes ABA by suppressing B-type ARR-SnRK2 interactions[77]. While these findings reveal a general CK-ABA antagonism, their specific interplay under LT remains to be fully elucidated. Notably, although endogenous CK signaling often impairs cold tolerance, exogenous CK application can enhance cold tolerance in both Arabidopsis and sugarcane[72,75]. Moreover, ZmRR1 in maize has been reported to improve cold adaptation[78], further highlighting the functional diversity of CK across species and conditions. In summary, under LT stress, CK finely balances growth and cold adaptation. LT generally downregulates CK biosynthesis and promotes its degradation, while suppressing the AHK2/AHK3 signaling pathway to shift resources from growth to defense. Notably, AHK2/AHK3 serve as negative regulators of cold tolerance, their loss-of-function mutants show enhanced cold tolerance—a suppression mechanism that may prevent over-investment in defense under mild or transient cold, thus preserving growth potential. CK function, however, is context-dependent and multifaceted: specific downstream regulators (e.g., certain type-A ARRs) can enhance cold tolerance, and exogenous CK application improves tolerance in several species. Thus, CK acts not merely as a growth promoter or cold-response repressor, but as a dynamic signaling hub that integrates environmental and developmental cues to optimize the growth-stress survival trade-off under LT conditions.
Gibberellin
-
GA serves as a central phytohormone regulating plant growth and development, including germination, elongation, and flowering[26,66]. Under LT stress, plants precisely balance growth and cold adaptation by modulating GA metabolism and signaling. Cold-induced CBFs upregulate Gibberellin 2-Oxidase (GA2ox) activity, reducing bioactive GA and subsequent accumulation of DELLA proteins. This growth restraint enhances cold tolerance[79,80]. DELLA proteins further interact with growth-regulating factors such as GROWTH-REGULATING FACTORS (GRFs) to coordinately regulate developmental processes under LT stress[81]. During anther development, LT suppresses GA biosynthetic genes such as OsGA20ox1, OsGA20ox3, and OsGA3ox1 through OsWRKY53, decreasing active GA and reducing pollen fertility[80,82]. In contrast, in rice, OsCTB5 interacts with OsHox12 to suppress OsGA2ox6 and activate OsGA3ox1 in anthers, promoting GA accumulation and supporting tapetum development under LT stress[56]. Notably, OsCTB5 is also induced by ABA and enhances seedling cold tolerance, regulating ABA signaling components such as SERINE/THREONINE-PROTEIN KINASE (OsSAPK6) and OsbZIP23, and by directly activating the PYR1-LIKE PROTEIN 9 (OsPYL9) receptor[56]. In summary, the GA pathway mediates strategic resource reallocation under LT stress. Cold-induced accumulation of growth repressors (e.g., DELLA proteins) restrain growth, thereby redirecting energy and metabolic resources toward cold acclimation. This trade-off—between reproductive development and stress survival, notably in processes like pollen fertility—underscores GA signaling as a central hub that integrates developmental progression with cold adaptation.
Brassinosteroids
-
BRs regulate key architectural traits such as leaf angle and tiller number[66,83]. Under LT stress, BRs participate in resource reallocation strategies. For instance, in chrysanthemum, cold-induced CmbZIP19 represses the BR biosynthetic gene CmDWF1, reducing branching—a phenotype reversible by exogenous BR[84]. Physiologically, BRs enhance membrane stability and cold tolerance by promoting membrane lipid ordering[85]. Molecularly, LT upregulates the biosynthetic gene DE-ETIOLATED2 (DET2) and represses catabolic genes CYP734A7/8, resulting in BR accumulation and activation of the core transcription factor BRASSINAZOLE RESISTANT 1 (BZR1), which positively regulates cold tolerance[86,87]. The kinase BIN2, a negative regulator of BR signaling, phosphorylates and destabilizes BZR1/BES1 under normal conditions[87]. However, under LT stress, BIN2 exhibits a dual temporal function: early attenuation allows BZR1 accumulation and direct activation of CBF1/2 and COR genes; later reactivation negatively regulates cold tolerance by degrading ICE1[3]. Additionally, BR signaling components such as CESTA directly promote CBF expression, synergistically enhancing the cold response[83]. BR signaling also integrates with other hormonal pathways. BR enhances auxin transport capacity by promoting PIN2 accumulation from the root tip to the elongation zone, thereby improving tropic responses[88]. BZR1 upregulates NCED1 to elevate ABA levels, whereas BIN2 suppresses NCED1 expression[87]. In apple (Malus × domestic), BR INSENSITIVE1-EMS-SUPPRESSOR1-INTERACTING MYC-LIKE PROTEIN1 (MdBIM1) activates MdCBF1 to enhance cold tolerance, while JASMONATE ZIM-DOMAIN Proteins (MdJAZ1/2) disrupt the MdBIM1-MdCBF2 complex, revealing an antagonistic interaction between BR and JA signaling under LT[89]. Furthermore, the BR-BZR1 module also promotes H2O2 production by activating RBOH1, induces photoprotective proteins, and activates autophagy-related genes ATGs and the receptor NEIGHBOR OF BRCA1 GENE 1 (NBR1) to maintain protein homeostasis[1]. In summary, BR signaling integrates multiple responses to LT. Its core transcription factor BZR1 directly enhances cold tolerance by activating key pathways like CBF and coordinating processes including ROS homeostasis and autophagy. BR signaling also extensively interacts with other hormonal pathways (e.g., auxin and ABA) to dynamically balance growth suppression and defense activation. A key temporal regulator is the kinase BIN2; its early attenuation promotes cold acclimation, while its later reactivation facilitates growth recovery. This time-dependent switch enables BR to precisely balance the growth-defense trade-off under LT stress.
Ethylene
-
ET plays a complex role in modulating cold tolerance and developmental plasticity under LT, acting as either a positive or negative regulator depending on species and conditions[3,76]. Notably, although SlNAC3 can activate CBF and CORs under LT, its primary function is to negatively regulate cold tolerance by directly upregulating key ET biosynthesis genes—ACC OXIDASE 1 (SlACO1), ACC SYNTHASE 2 (SlACS2), and SlACS4—thereby promoting ET release[90]. In contrast, MaDREB1F can promote ET accumulation by activating the promoter of MaACO20[91]. At the signal transduction level, components are differentially regulated under LT and perform opposing functions. Receptors ETHYLENE RESPONSE 1 (ETR1) and ETHYLENE INSENSITIVE 4 (EIN4) are rapidly upregulated under LT conditions, enhancing cold tolerance[76]. However, the expression of the EIN2-EIL1 module is suppressed, together with stabilized EIN3, which inhibit CBF and ARR5/7/15, negatively regulating cold tolerance[76]. Downstream ET RESPONSE FACTORS (ERFs) play as central integrators. In various species, ERFs such as PtrERF9, VaERF092 and MfERF1 enhance cold tolerance by modulating antioxidant defenses, glutathione metabolism, polyamine/proline accumulation, and interacting with WRKY or GST pathways[3,92−95]. In summary, ET orchestrates a multilayered regulatory system that balances growth and stress responses under LT through the dynamic control of biosynthesis, differential regulation of signaling components, and activation of a diverse ERF transcriptional network. ET functions as a context-dependent switch that fine-tunes the balance between cold adaptation and growth maintenance. Its role varies across species and even within components of the same signaling cascade, reflecting a sophisticated plasticity that tailors responses to stress intensity, duration, and developmental stage. This regulatory duality allows ET to enhance cold tolerance under certain conditions while limiting excessive growth inhibition in others, positioning it as a dynamic modulator of resource allocation under LT stress.
Jasmonic acid
-
JA enables plants to dynamically balance freezing tolerance with growth under LT stress through a sophisticated molecular network. Biosynthetic genes—such as LIPOXYGENASES (LOXs) and ALLENE OXIDE CYCLASES (AOCs) in Arabidopsis, and LIPOXYGENASE D (SlLOXD) and ALLENE OXIDE SYNTHASE (SlAOS) in tomato—promote JA accumulation[96−98]. In wheat, TaPAP6L enhances JA synthesis by improving the precursor α-linolenic acid and simultaneously inhibiting Jasmonate Resistant 1 (TaJAR1)-mediated JA degradation[99]. Accumulated JA activates the core COI1-JAZ-MYC2 module: JAZ repressors are degraded, releasing ICE1 and MYC2 to activate CBF and CORs[96,100,101]. JA signaling integrates with other pathways through multi-layered mechanisms. JA signaling extensively integrates with other pathways. MdJAZ1/2 suppress MdBBX37-mediated activation of MdCBF1/4, and disrupt MdBBX37-MdICE1 interaction, while MdMIEL1 degrades MdBBX37, attenuating cold responses[102]. JAZ proteins interact with EIN3/EIL1 and ICE1 to regulate the CBF cascade[96,103]. Far-red light-activated phyA sequentially activates ABA and JA to enhance cold tolerance via the CBF[104]. Positive feedback occurs through SlWRKY50-mediated JA biosynthesis and SlMYC2-activated SlWRKY50 transcription[97]. Homeostasis is maintained via negative feedback, e.g., OsHAN1 hydroxylates JA-Ile to its inactive form[105]. Interestingly, JA also participates in a temporally regulated network in response to LT: SlMYB15 activates the SlLOXD promoter during early LT stress to promote JA synthesis, while simultaneously repressing SlMYC2[99]. At this stage, rising JA and ABA jointly mediate antioxidant defense[98]. In later stages, SlMYB15 is suppressed, leading to decreased ABA levels and the derepression of SlMYC2. SlMYC2 then directly binds to the promoters of SlLOXD and SlCBF1/2, ensuring the temporally precise contribution of JA signaling during LT stress[98]. JA acts as a temporally tuned modulator of the growth-defense equilibrium under LT stress. Its signaling follows a phased program: early in LT, JA cooperates with ABA to initiate antioxidant defenses; under prolonged stress, core components like MYC2 are derepressed to amplify CBF-mediated cold adaptation, while selectively restraining growth. This time-resolved rewiring prioritizes resource allocation, exemplifying how JA optimizes survival during extended LT conditions.
Salicylic acid
-
SA serves as a key signaling molecule in plant responses to LT stress, contributing to cold tolerance and balancing immunity and cold adaptation[3,66]. Under prolonged LT conditions (e.g., 4 °C for more than 1 week), repression of SALICYLIC ACID-DEFICIENT 1 (SARD1), CBP60-LIKE g (CBP60g), ISOCHORISMATE SYNTHASE 1 (ICS1), and ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) by CAMTA1/2/3 complexes is released, activating the isochorismate pathway and driving SA accumulation[106−108]. SA signaling is transduced through multiple mechanisms. The SA receptor NON-EXPRESSER OF PR GENES 3 (NPR3) relieves TGA2 repression, activating the proline biosynthesis gene Δ1-PYRROLINE-5-CARBOXYLATE SYNTHASE 1 (P5CS1) to enhance cold tolerance, and forms a positive feedback loop by promoting ICS1[1]. Meanwhile, SA activates immune responses via the NPR1-dependent upregulation of PATHOGENESIS-RELATED PROTEIN 2 (PR2) and PR5[107]. Independently, NPR1 interacts with HSFA1 to activate cold-tolerance genes[109], while LT induces an ICE1-NPR1-TGA3 complex that directly activates immune-related genes like PR1, thereby synergistically enhancing responses to both biotic and abiotic stresses[110]. SA action is dose-dependent: excess SA suppresses COR genes, impairing cold tolerance and delaying growth. This reflects a dynamic interaction between SA and cold-response pathways—while LT can co-activate both, persistently high SA levels eventually inhibit CORs expression, mediating a resource trade-off between growth and tolerance[111]. At the hormone cross-talk level, ET signaling via EIN3 suppresses ICS1/2 and EDS5, inhibiting both SA biosynthesis and the CBF pathway[108]. ABA potentially attenuates SA-mediated immunity by promoting CUL3-NPR3/NPR4-mediated ubiquitination and degradation of NPR1[112,113]. Epigenetically, TaBZR1 recruits TaHAG1 to enhance histone acetylation and activate TaSAMT1, promoting SA methylation and enabling coordinated regulation between BR and SA signaling[86]. In summary, the specific accumulation of SA under prolonged LT exposure demonstrates the recruitment of immune components during abiotic stress. Through modules like NPR, SA enhances basal cold tolerance and integrates immune responses. This synergy, however, is dose-dependent: excessive SA can compete with classical cold-response pathways (e.g., the CBF regulon) and suppress growth. Thus, SA acts as a sensitive regulatory valve, whose dynamic concentration determines the prioritization of resources among cold resistance, immune vigilance, and growth, making it a key integrative node in adaptation to complex environmental pressures.
Strigolactone
-
SL plays a significant regulatory role in plant adaptation to LT stress. LT induces the expression of key SL biosynthetic genes such as MORE AXILLARY GROWTH 1 (MAX1), MAX3, and MAX4, promoting SL accumulation[14,114]. Signal transduction through the MAX2-dependent ubiquitination pathway enhances cold tolerance via two parallel mechanisms: degradation of the transcriptional repressor WRKY41 to inhibit CBF, and degradation of SUPPRESSOR OF MAX2 1-LIKE PROTEINS (SMXLs) to activate anthocyanin biosynthetic genes[14]. Simultaneously, SL signaling induces the accumulation of HY5, which triggers the autophagy-related gene ATG18a, promoting the clearance of ubiquitinated proteins and further enhancing cold tolerance[115]. SL signaling integrates with multiple hormonal networks to optimize cold adaptation. It upregulates ABA biosynthetic genes, such as NCED6 and ABA signaling components, including ABF4, thereby reinforcing ABA-dependent stress responses[2]. In apple, the MdGLK1-MdBZR1 module enhances cold tolerance by activating MdCBF1/2, while ET negatively regulates this process through MdEBF1-mediated degradation of MdGLK1. Additionally, the SL signaling repressor MdSMXL8 fine-tunes this pathway by interfering with the MdGLK1-MdBZR1 interaction[116]. Furthermore, kairavins (KARs) perceive LT through the receptor KAI2, regulating MAX1-dependent SL biosynthesis and SnRK2.5-mediated ABA signaling. This coordination activates the CBF pathway and enhances antioxidant capacity, simultaneously maintaining both growth and stress tolerance in tomato[117]. In summary, SL functions as a systemic integrator in cold adaptation. Through MAX2-dependent signaling, SL enhances cold tolerance via multiple direct mechanisms: degrading WRKY41 to derepress CBF expression, promoting anthocyanin accumulation via SMXL protein degradation, and inducing HY5 to activate autophagy for proteostasis. Furthermore, SL signaling reinforces ABA biosynthesis and interacts extensively with light, BR, ET, and KAI2-mediated karrikin networks, thereby fine-tuning resource allocation and the growth-defense balance under LT stress. This multi-layered integration establishes SL as a key regulatory hub linking developmental programs, environmental signals, and the cold-tolerant phenotype.
Hormonal crosstalk
-
Under LT stress, plants balance cold tolerance and growth through a dynamic, multi-layered phytohormone network. This system operates via progressive, interwoven interactions that precisely control physiological outputs, optimizing the survival-growth equilibrium (Fig. 3). Multiple hormones converge on core LT pathways (e.g., CBF transcription factors) to set initial strategic priorities. For instance, SlERF15 (ET-responsive) and MdABI4 (ABA-responsive) directly activate CBF gene, reinforcing basal cold acclimation[60,65]. Concurrently, competition and counterbalance occur: CK signaling via AHK2/AHK3 type-A ARRs negatively regulate both CBF expression and ABA responses, preventing over-investment in defense under mild or transient LT[75]. This synergy-antagonism dualism toward shared targets defines the network's foundational logic, determining whether the plant hardens, or maintains growth based on signal strength and stress severity.
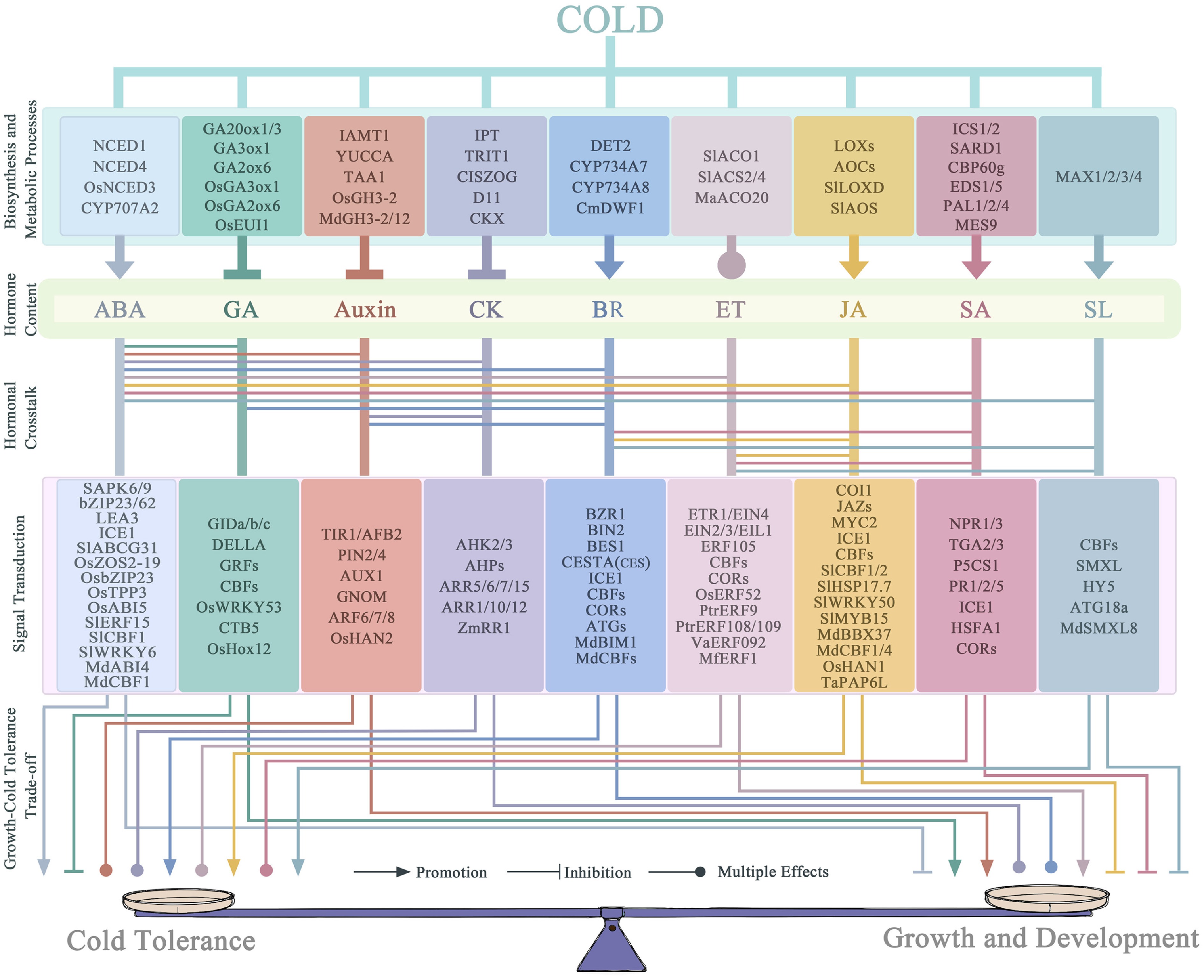
Figure 3.
A model of hormone-mediated trade-offs between growth and cold tolerance in plants under LT stress. LT stress triggers the reconstruction of hormonal homeostasis in plants and activates a multi-layered signaling interaction network. This regulatory process initiates with the perception and transduction of cold signals through plasma membrane-associated receptors and downstream signaling components, thereby modulating the expression and activity of key enzymes involved in the biosynthesis and metabolism of several essential hormones, such as ABA, GA, Auxin, CK, BR, ET, JA, SA, and SL, leading to dynamic shifts in endogenous hormone levels. Subsequently, these hormones engage in cross-talk between pathways, involving both synergistic and antagonistic interactions, to relay signals downstream. The hormonal signals are further integrated with transcriptional regulators and post-translational modification systems, collectively coordinating resource allocation and physiological balance between cold acclimation responses and normal growth and development. Ultimately, this hormone-regulated network precisely reprograms developmental progression and carbon-nitrogen metabolic partitioning, enabling the maintenance of physiological homeostasis and plant adaptation under LT stress.
The network further enables dynamic buffering and feedback through bidirectional regulation of hormone biosynthesis and signaling (Fig. 3). For example, BR promotes ABA synthesis via BZR1, while BIN2 kinase inhibits it, forming an intrinsic balancing switch[87]. ET suppresses SA biosynthesis through EIN3, limiting resource diversion to immune responses[108]; ABA can further modulate this by promoting degradation of NPR1, a key SA-signaling regulator[112,113]. Such mutual regulation provides flexibility: defense-related hormones are enhanced during sustained stress, while growth-promoting hormones are suppressed, with rapid reversal upon recovery.
At the most sophisticated level, hormone signals are integrated by specific transcription factors or protein complexes into functional modules that execute localized, precise adjustments. MdHY5, for instance, coordinately downregulates auxin and upregulates ABA metabolism, inducing anthocyanin synthesis for photoprotection without stimulating overall growth[8]. The MdGLK1-MdBZR1 module (BR-related) enhances cold tolerance but is constrained by ET and fine-tuned by SL repressors, ensuring a measured defense[103]. JA signaling interacts via JAZ proteins with ET components and ICE1, fine-tuning the CBF cascade[96,103]. In apple, MdJAZ1/2 interacts with MdABI4 to disrupt the MdABI4-MdICE1 complex, thereby integrating JA and ABA signals into the CBF pathway[60]. These multi-hormone modules act as 'specialized task forces' executing localized adaptive tasks—enhancing resistance, modifying organ morphology, or redirecting metabolism—to optimize the growth and defense balance at a fine scale.
In summary, the hormonal network governing plant LT response forms a complete regulatory system: it sets global direction through core-pathway convergence, dynamically reallocates resources via inter-hormone crosstalk, and achieves localized adaptation through integrated modules (Fig. 3). This hierarchical, networked design provides remarkable plasticity and resilience, maximizing survival under stress while preserving growth potential for recovery. a deeper understanding of this network is central to elucidating plant environmental adaptation and guiding the genetic improvement of crop stress tolerance.
-
In exploring how plants balance growth and development with cold tolerance under LT stress, this review synthesizes the sophisticated molecular mechanisms that enable this equilibrium. As a key environmental factor influencing plant geographical distribution and agricultural productivity, LT stress compels plants to optimize the trade-off between survival and reproductive success. This balance is achieved through a complex and finely tuned molecular network encompassing LT perception and signal transduction, expression of cold-tolerance genes, and regulation of hormonal signaling pathways. Plant perception of LT signals represents the initial step in the entire response cascade and serves as the foundation for subsequent complex regulatory events. Plants perceive LT signals through plasma membrane-localized receptors and ion channels, rapidly triggering changes in intracellular calcium ion concentrations and subsequently activating a series of signal transduction pathways, such as the MAPK cascade system. Ultimately, these intracellular signals converge transcriptionally, centered on the ICE1-CBF module. This module specifically activates cold-tolerance genes (e.g., CORs), whose products enhance membrane stability[2,9]. Together, they establish a rapid, coordinated molecular defense, enabling plants to mount effective protection against LT stress within a short timeframe.
Plant hormones play a central role in balancing cold tolerance and growth and development. Hormones such as ABA, auxins, CK, GA, BR, ET, JA, and SA collectively shape plant stress-resistant phenotypes and developmental trajectories through intricate and finely tuned synergistic and antagonistic interactions. For instance, ABA accumulates under LT stress and functions as a central 'cold response commander', enhancing defense by promoting stomatal closure and antioxidant synthesis, while concurrently inhibiting cell elongation and division[4,63,64]. Within this context, phytohormones collectively enable plants to dynamically prioritize between defense and growth. ABA accumulates The signaling pathways of growth-promoting hormones, such as auxin and CK, are reprogrammed; their inherent functions are temporarily suppressed to reallocate resources toward adaptive responses like deeper root growth for improved resilience[34,73,74]. GA and BR fine-tune the balance through distinct mechanisms: GA-mediated growth is slowed via suppression of its biosynthesis and stabilization of DELLA proteins, while BR transcription factors like BZR1 coordinately regulate genes for both cold tolerance and growth. ET, JA, and SA further contribute in a context-dependent manner, integrating defense signals and modulating immune responses.
These hormones form a resilient, interconnected network by converging on common targets (e.g., the CBF pathway), engaging in mutual regulation of biosynthesis and signaling, and assembling into multi-hormone modules (e.g., MdHY5 coordinating IAA and ABA metabolism). This architecture allows flexible resource allocation between growth and defense according to stress intensity and duration.
Consequently, plants adopt a 'survival-first' strategy under LT. Growth processes—including cell division, expansion, and reproductive development—are temporarily suppressed or delayed. This redirects energy and materials toward acclimation mechanisms, enhancing short-term survival at the cost of delayed growth and yield—an evolutionary trade-off that prioritizes viability under stress.
In summary, the plant LT response is a systematic, multi-layered process. Signal perception provides accurate environmental input. The hormonal network integrates these signals through synergy and antagonism to shape phenotypic outcomes, and resource allocation executes these decisions, optimally distributing limited resources between survival and development. Future research should further dissect the dynamic architecture and key regulatory nodes of this network. Such efforts will deepen our understanding of how plants balance growth and development with cold tolerance under LT stress and to provide strategies for sustainable agricultural development: (1) Elucidate the mechanisms of hormone signal cross-talk: Conduct in-depth investigations into the interaction networks among multiple hormones at the levels of transcriptional regulation, epigenetics, and non-coding RNAs to reveal the precise mechanisms underlying their coordinated regulation of stress resistance and growth. (2) Explore and utilize cold-tolerance gene resources: Cold-tolerance genes can be mined from crop wild relatives via genome-wide association studies and functionally validated. Using gene editing, these elite alleles can then be precisely integrated into modern cultivars. This molecular design-based breeding approach effectively enhances both crop cold tolerance, and yield stability. (3) Develop green cold tolerance strategies: Research and develop environmentally friendly cold-tolerance agents based on plant hormones or natural products to reduce the negative impacts of chemical agents on ecosystems. (4) Promote multidisciplinary collaborative research: Integrate approaches from botany, molecular biology, ecology, and climate science to systematically elucidate plant cold adaptation mechanisms, providing theoretical foundations and technical support for agricultural production in the context of global climate change. In conclusion, by thoroughly analyzing the molecular mechanisms of plants under LT stress, the cross-talk within hormone signaling networks, and the development and application of environmentally friendly cold tolerance agents, we will be better equipped to address the challenges posed by global climate change and ensure the sustainability of agricultural production. This knowledge also advances our comprehension of plant adaptation strategies and ecological functions within natural ecosystems, thereby providing valuable insights for ecological conservation and sustainable development.
-
The authors confirm contribution to the paper as follows: study conception and design: Ye J, Yu C, Hu Y; data collection: Ye J; analysis and interpretation of results: Ye J, Hu Y; draft manuscript preparation: Ye J, Yu C. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
Received 3 December 2025; Accepted 2 March 2026; Published online 26 March 2026
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ye J, Yu C, Hu Y. 2026. Decoding the cold signal: hormonal networks orchestrating the trade-off between growth and tolerance. Tropical Plants 5: e008 doi: 10.48130/tp-0026-0006 

Decoding the cold signal: hormonal networks orchestrating the trade-off between growth and tolerance
- Received: 03 December 2025
- Revised: 06 February 2026
- Accepted: 02 March 2026
- Published online: 26 March 2026
Abstract: Low-temperature (LT) stress represents a pivotal environmental factor that significantly impacts plant growth, geographical distribution, and agricultural productivity. As sessile organisms, plants have evolved a suite of sophisticated mechanisms to adapt to LT environments, achieving an optimal balance between growth and cold tolerance, a phenomenon termed the growth and tolerance trade-off. This balance is not a passive outcome but rather an active, dynamic, and adaptive decision-making process regulated by a complex molecular network. This review systematically summarizes the molecular mechanisms underlying plant responses to LT and the equilibrium between growth and cold tolerance, with a particular emphasis on LT perception and signal transduction, the suppression of growth and developmental processes, and the regulatory role of hormonal networks. It underscores the central role of hormonal signaling networks in balancing cold tolerance and growth, and also explores potential future applications in this field.
-
Key words:
- Low-temperature (LT) /
- Growth /
- Cold tolerance /
- Hormonal signaling /
- Trade-off