








-
Plant microRNAs (miRNAs) are endogenous small noncoding RNAs ranging from 20 to 24 nucleotides (nt) in length. Despite their tiny size, miRNAs play a crucial role in regulating gene expression, thereby modulating various processes of plant development and plant–environment interactions[1]. MIRNA genes are initially transcribed by RNA polymerase II to generate hairpin-containing primary miRNAs (pri-miRNAs), which are subsequently processed into miRNA/miRNA* duplexes, mainly by the RNase III-type endonuclease DICER-LIKE 1 (DCL1)[2]. The nascent miRNA/miRNA* duplexes undergo 2'-O-methylation on both strands, a reaction catalyzed by the methyltransferase HUA ENHANCER 1 (HEN1)[3]. Following this modification, the miRNA strand is selectively loaded onto ARGONAUTE (AGO) proteins to assemble the miRNA-induced silencing complex (miRISC), but the miRNA* strand is discarded and degraded[1,2]. AGO proteins are highly conserved across almost all three domains of life, with eukaryotic AGO proteins associating with distinct classes of small RNAs and serving as the catalytic core of gene silencing pathways[4]. Typically, miRISC recognizes its target messenger RNAs (mRNAs) and other noncoding RNAs (e.g., the precursors of trans-acting small interfering RNAs (siRNAs)) through sequence complementarity, leading to the target's downregulation via mRNA cleavage or translational repression, or generation of secondary siRNAs[2]. Conversely, miRNAs are also subject to elaborate degradation mechanisms mediated by exoribonucleases, such as SMALL RNA DEGRADING NUCLEASE (SDN)[5] and ATRIMMER 2 (ATRM2)[6]; nucleotidyl transferase proteins (NTPs), including HEN1 SUPPRESSOR 1 (HESO1) and UTP : RNA URIDYLYLTRANSFERASE 1 (URT1)[7−9]; and target mimicry RNAs[10,11]. As master regulators of post-transcriptional gene expression, miRNAs are themselves tightly regulated. Dysregulation of global or even individual miRNA levels, whether through disrupted biogenesis or altered degradation, can have profound physiological consequences, ranging from developmental abnormalities to embryonic lethality[1]. However, in contrast to the well-characterized pathway of miRNA biogenesis, the molecular mechanisms underlying miRNAs' turnover and degradation remain much less well understood. This review focuses on advances in understanding the molecular mechanisms controlling miRNAs' stability, highlighting how the miRISC serves as a central platform where these regulatory events unfold.
-
AGO proteins comprise four conserved domains: the N-terminal, PAZ (Piwi–Argonaute–Zwille), MID (middle), and PIWI (P-element-induced wimpy testis) domains[4,12]. Among these, the MID and PAZ domains are crucial for protecting miRNA: The MID domain binds the miRNA's 5'-end, whereas the PAZ domain shields the 3'-end. This dual protection physically prevents various exoribonucleases and NTPs from accessing the miRNA termini, establishing AGO proteins as central guardians of miRNAs' stability[13].
In eukaryotes, different AGO proteins load distinct small RNA species. The Arabidopsis genome contains 10 AGO proteins, among which AGO1 plays a dominant role in the miRNA pathway[2,12]. The majority of miRNAs are preferentially loaded onto AGO1. Our recent work demonstrated that most miRNAs are highly stable, with half-lives exceeding 48 h, whereas impaired AGO1 activity markedly accelerates their decay[14]. Consistent with this, an early study observed that the malfunction of AGO1 results in reduced accumulation of miRNAs and pleiotropic developmental defects[15]. These findings suggest that AGO1 not only serves as the major effector protein for miRNAs but also acts as a crucial protector of miRNAs' stability. Analyses of different ago1 mutants provide additional insights into the role of different domains in controlling miRNAs' stability. In the putative null mutant ago1-36, which harbors a T-DNA insertion in the PAZ domain, miRNAs undergo significantly increased 3'-end tailing and trimming, hallmark features of 3'-end destabilization of miRNA[16]. By contrast, the weak mutants ago1-11, ago1-25, and ago1-27, which carry point mutations in the PIWI domain and diminish endonuclease activity, show no significant changes in miRNAs' 3'-end tailing and trimming[16]. These observations suggest that AGO1 loading and its subsequent targeting (see below for details), but not the catalytic activity of AGO1, are critical for miRNAs' 3'-end integrity.
Unlike AGO1, AGO10 and AGO7 participate in specialized miRNA regulation. AGO10 sequesters miR165/166 with high affinity and promotes their degradation partially through SDN1/2[5]. Accordingly, miR165/166 levels are elevated in ago10 loss-of-function mutants. AGO7, on the other hand, specifically binds miR390. Intriguingly, miR390's abundance is increased in ago7 mutants, indicating that AGO7 serves as a negative regulator of miR390 abundance in vivo[17].
Animal miRNAs differ fundamentally from plant miRNAs in lacking 2'-O-methylation at the 3' termini, making AGO protein association more crucial for their stability. Human cells express four AGO proteins (HsAGO1–4), with miRNAs predominantly loaded onto HsAGO2. Supporting its protective role, early in vitro studies revealed that the HsAGO2-guided RNA complex exhibits remarkable stability, even in the presence of nucleases[18]. Overexpression of HsAGO2 enhances miRNAs' abundance, whereas loss-of-function mutations reduce miRNA expression[19]. Although HsAGO2 is the only human AGO with intrinsic endonuclease activity, ectopic expression of any human AGO protein, including a cleavage-deficient HsAGO2 variant, leads to increased miRNA levels. These findings demonstrate that AGO slicing activity is not required for miRNAs' stability in human cells[19]. Notably, disruption of the PAZ domain binding pocket of HsAGO2, which results in the exposure of the miRNA's 3'-end, triggers extensive oligouridylation of miRNAs by TUTases and subsequent degradation by the exoribonuclease DIS3L2[20].
On the other hand, the miRNA–AGO interaction also stabilizes AGO proteins. In Arabidopsis, the AGO1 protein levels decline in miRNA biogenesis-deficient mutants such as hyl1[21]. Interestingly, a recent study identified the N-coil of AGO1 as a structural switch that is exposed in empty AGO1 and recognized by the autophagy cargo receptor ATG8-INTERACTING-PROTEIN1 (ATI1) for degradation[22]. In Drosophila, "empty" AGO proteins (those not bound to miRNAs) are degraded via Iruka-directed ubiquitination, followed by valosin-containing protein (VCP)-mediated selective autophagy[23,24]. These findings highlight the mutual dependence between AGO proteins and miRNAs for structural stability. In summary, beyond their canonical gene silencing roles in miRNA-directed pathways, AGO proteins also mediate the critical post-transcriptional regulation of miRNAs' stability. In most cases, AGO binding protects miRNAs from degradation by exoribonucleases and terminal uridylyl transferases. However, certain AGO proteins, such as AGO10, can sequester specific miRNAs and promote their degradation, likely by recruiting exoribonucleolytic machinery.
-
The sorting and selective loading of a miRNA strand from a miRNA/miRNA* duplex is primarily governed by its 5' terminal nucleotide. In plants, AGO1 exhibits a strong preference for binding miRNAs with a 5' terminal uridine (U)[25]. Similarly, nucleotide analyses in humans and Drosophila have revealed a bias for uridine (U) and adenine (A) at the 5'-end of AGO-bound miRNAs[26]. Structural studies further demonstrate that the MID domain of human AGO2 exhibits higher binding affinity for UMP/AMP compared with CMP/GMP[26]. In Drosophila, replacing the 5'-end A with U enhances miRNA's stability and prolongs its half-life, whereas mutating it to guanine (G) does not significantly impact its stability[27]. In mammalian cells, miRNAs with fast turnover rates are significantly enriched in 5'-end G and cytosine (C) but lack 5'-end U[28]. Collectively, these findings suggest that the 5'-end nucleotide serves as a key determinant for miRNA's stability. A plausible explanation is that the first nucleotide at the miRNA's 5'-end serves as a primary signal for AGO protein recognition; as established, AGO's incorporation is essential for stabilizing miRNA.
-
Plants miRNAs typically bind their target mRNAs with perfect or near-perfect complementarity. Inductive pairing rules for miRNA–target interactions have been established: Positions 2–12 allow a maximum of two consecutive mismatches, Positions 9–11 must be perfectly matched, and downstream of Position 13, non-consecutive mismatches are permitted, with a maximum of four in total[29]. Critically, base pairing at Positions 10 and 11 of the miRNA is essential for triggering the endonucleolytic activity of AGO proteins, enabling precise cleavage of the target mRNAs.
In the nascent miRISC complex (i.e., without target RNA), the 3' terminus of miRNA is anchored by the PAZ domain of AGO1. This anchoring mechanism effectively shields the 3' terminus of miRNA from degradation mediated by exoribonucleases and NTPs[13]. Structural analyses revealed that when extensive complementary base pairing is established between miRNA and its target, the AGO protein undergoes a conformational change that exposes the miRNA 3'-end from its PAZ domain[30]. Since plant miRNAs typically recognize target mRNAs through extensive base pairing, the miRNA 3'-end is exposed during target recognition, rendering it vulnerable to enzymatic attack. Following RNA-induced silencing complex (RISC)-mediated mRNA cleavage, HESO1 and URT1 are recruited to AGO1 through protein–protein interaction to uridylate and promote the degradation of RISC-cleaved mRNA fragments[31,32]. In this scenario, the 3'-end 2'-O-methylation of plant miRNAs is crucial to protect them from HESO1- and URT1-mediated uridylation.
In plants, DCL1 sequentially processes a pri-miRNA into a miRNA/miRNA* duplex with characteristic 3'-end 2-nt overhangs. This duplex structure is recognized by the small RNA methyltransferase HEN1, which catalyzes 2'-O-methylation on the ribose of the 3' terminal nucleotide of both strands. This modification is critical for maintaining the stability of plant miRNAs[3]. Besides miRNAs, all kinds of plant siRNAs also bear the HEN1-directed 2'-O-methylation. In hen1 mutants, endogenous miRNA levels decrease significantly, leading to pleiotropic developmental defects. Notably, miRNAs in hen1 mutants undergo extensive 3'-end uridylation[7]. Small RNA uridylation has been identified in various organisms. In Chlamydomonas, the nucleotidyl transferase MUT68 mediates 3'-end uridylation of unmethylated sRNAs and collaborates with the exosome subunit RRP6 for sRNA quality control[33]. HESO1, the homologous nucleotidyl transferase of MUT68 identified in Arabidopsis, catalyzes unmethylated miRNA uridylation in hen1 and promotes miRNA degradation[7]. Additionally, URT1 also catalyzes the uridylation of a subset of miRNAs, particularly in the absence of HESO1[8]. Interestingly, even in a HEN1-competent background, HESO1 and URT1 uridylate a small fraction of miRNAs that undergo a low ratio of methylation (e.g., miR158)[8]. Taken together, HESO1 and URT1 act in a cooperative and redundant manner to mediate the uridylation of AGO-bound unmethylated miRNAs, thereby participating in miRNA turnover.
In contrast, animal miRNAs usually recognize their targets via the "seed region" (Nucleotides 2–7)[34], allowing the 3'-end of the miRNA to remain anchored in the PAZ domain. Accordingly, animal miRNAs lack 3'-end 2'-O-methylation. Nevertheless, the PIWI-interacting RNAs (piRNAs) and certain types of siRNAs, such as the AGO2-associated siRNAs in Drosophila and the ERGO-1-associated 26G RNAs in Caenorhabditis elegans, possess 2'-O-methylation, directed by HEN1-orthologous proteins[35]. In both plants and animals, 3'-end methylation contributes to the stabilization of sRNA. Loss of methylation is consistently associated with 3'-end trimming, uridylation, and the decreased accumulation of sRNAs. Notably, as animal miRNAs lack 2'-O-methylation, nontemplated uridines can be incorporated at the 3'-ends by terminal nucleotidyl transferases, including TUT4 and TUT7. These uridylated miRNAs are then recognized and degraded by the exoribonuclease DIS3L2[20].
In summary, 3'-end 2'-O-methylation is critical for miRNA stability, whereas 3' uridylation promotes its degradation. Importantly, methylation serves as a prerequisite for stability, as uridylation only occurs on unmethylated substrates.
-
Though the canonical mode of target recognition by plant miRNAs relies on extensive base pairing, a notable exception is the Arabidopsis long noncoding RNA INDUCED BY PHOSPHATE STARVATION 1 (IPS1), which contains a sequence that is nearly perfectly complementary to miR399 but features a 3-nt bulge at the central cleavage site. Consequently, IPS1 binds and sequesters endogenous miR399 instead of being cleaved, thereby alleviating miR399-mediated repression of its target, PHOSPHATE 2 (PHO2)[36]. Inspired by this natural mechanism, artificially designed uncleavable "target mimics" (MIMs) containing a central 3-nt bulge have been developed to effectively knock down specific endogenous miRNA families in various plant species[37]. A similar strategy, the short tandem target mimic (STTM), uses two adjacent 3-nt bulged target sites separated by a linker[38,39]. Notably, both the MIM and STTM strategies lead to the destabilization of endogenous miRNAs instead of its initial role as an miRNA "sponge", although miR399 exhibits relative stability when bound to IPS1[36].
The F-box protein HAWAIIAN SKIRT (HWS) was identified through genetic suppressor screens using MIM and STTM transgenic reporter systems[10,11]. Loss-of-function mutations in HWS restored the depletion of endogenous miRNA caused by target mimicry to almost wild-type levels, demonstrating an essential role for HWS in target mimicry-directed miRNA degradation in plants (pTDMD)[10,11]. Interestingly, HWS is crucial for the degradation of short-lived miRNAs under normal growth conditions and for the elimination of specific miRNAs under stress conditions[14]. Notably, in hws mutants, target mimicries and their cognate miRNAs were retained in the miRISC, whereas the normal targets were barely detected[11]. Although HWS probably functions as a key component of the S-phase kinase-associated protein 1 (SKP1)–CUL1–F-box protein (SCF)-type E3 ligase, it remains to be determined whether HWS specifically recognizes and triggers the clearance of the "AGO1–miRNA–target mimicry" ternary complex. Furthermore, genetic studies revealed that STTM-triggered degradation of endogenous miRNAs also depends on the exoribonucleases SDN1 and SDN2, as the sdn1 sdn2 double mutant suppressed both STTM165-induced morphological changes and reductions in miR165[38]. However, whether HWS and SDNs act synergistically or independently in target mimicry-dependent miRNA degradation remains unclear. Additionally, although target mimicry relies on an uncleavable central bulge in the miRNA–target interaction, but the pairing patterns flanking this central region remain to be systematically investigated.
In animals, extensive base pairing between miRNAs and their target mRNAs is not frequently observed. As mentioned above, animal miRNA targeting relies on the seed region (Nucleotides 2–7), which enables AGO proteins to recruit accessory proteins, leading to the target RNA's destabilization and/or translational repression. Notably, introduction of extensively complementary target mRNAs in Drosophila and human cells effectively triggers 3'-end tailing and 3'-to-5' trimming of the cognate miRNAs, a molecular pattern resembling the hen1 mutant in plants[40]. In HeLa cells, copurification analyses suggest that TUT1 and DIS3L2 are involved in miRNA 3'-end tailing and trimming processes[41]. Furthermore, TUT4 and TUT7 are essential for the uridylation of mature miRNAs and turnover of specific miRNA species[20].
In animal cells, introducing competitive inhibitors such as "bulged sponges" and "tough decoys", which harbor base pairing mismatches or a bulge in the central region complementary to cognate miRNAs, also effectively degrade endogenous miRNAs and upregulate the target genes' expression[42]. Viral noncoding RNAs HSUR1/2 can also trigger the host's cellular miRNA degradation during viral pathogenesis[43]. In 2015, the phenomenon whereby highly complementary targets with central mismatches (or bulges) trigger miRNA degradation was termed target RNA-directed miRNA degradation (TDMD)[44]. More importantly, the physiological relevance of endogenous TDMD in animals has been increasingly demonstrated, positioning TDMD as a general mechanism of miRNA turnover[34]. Endogenous TDMD-triggering targets in animals typically exhibit extensive pairing with cognate miRNAs at both the seed region and 3'-end, often forming an internal bulge. Notably, such targets not only induce a specific reduction in endogenous miRNA levels but also introduce 3'-end tailing and 3'-to-5' trimming of cognate miRNAs[42,45], the latter phenomenon now termed target-directed tailing and trimming (TDTT)[46]. Though TDMD and TDTT usually occur concomitantly, their underlying molecular mechanisms are distinct. Notably, the TDTT phenomenon in animal cells parallels the molecular behavior of miRNAs in the hen1 background, which points to the evolutionary conservation of miRNA 3'-end destabilization mechanisms between plants and animals.
The molecular executor of TDMD in animals is the E3 ubiquitin ligase ZSWIM8[46,47]. During TDMD, the extensive miRNA–target pairing induces a conformational change in AGO. ZSWIM8 recognizes this altered state and polyubiquitinates AGO proteins, and thereby triggers proteasome-mediated proteolysis[48]. Following release, the miRNA is rapidly degraded by cytoplasmic exonucleases, whereas the trigger RNA is recycled to initiate further rounds of turnover.
Collectively, target RNAs play a critical role in regulating the stability of their cognate miRNAs, with the degree and pattern of complementarity between miRNAs and their target RNAs being key determinants. In both plants and animals, similar target mimicry strategies are used to destabilize AGO-bound miRNAs. Interestingly, although both HWS and ZSWIM8 function as E3 ubiquitin ligases, they are nonhomologous proteins. Their convergent functions suggest that plants and animals have independently evolved similar mechanistic solutions for target-directed miRNA degradation.
-
In summary, this review consolidates current insights into the regulation of miRNAs' stability from the perspective of AGO1 as a central hub (Fig. 1). First, through recognition of the 5' terminal nucleotide and sensing of the miRNA/miRNA* duplex thermodynamic asymmetry, as well as the pairing structure, AGO1 selectively loads the miRNA, thereby protecting it from exonucleases and nucleotidyl transferases. Second, AGO1 recruits specific factors—such as SDNs, HESO1, and URT1—to promote miRNA's destabilization, and thus prior 2'-O-methylation by HEN1 is crucial for miRNA's 3'-end integrity. Third, centrally bulged target mimicries trigger degradation of the targeted miRNA through an HWS-dependent pathway.
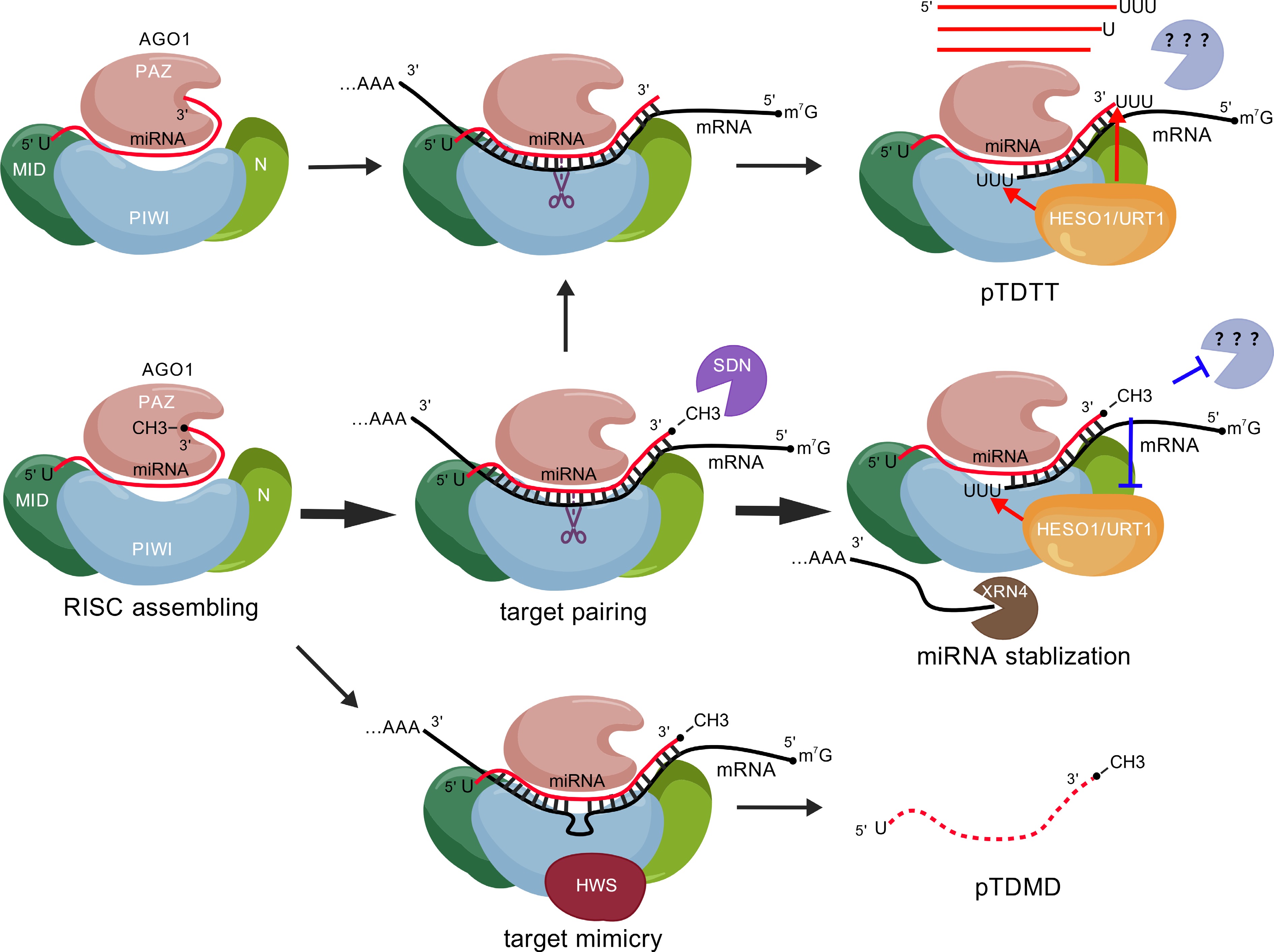
Figure 1.
AGO1-centered regulatory framework governing the dynamics of miRNA's stability. During assembly of the RISC, AGO1 recognizes the 5' terminal nucleotide of miRNAs and sequesters both ends of the miRNA molecule, thereby shielding them from exonucleolytic degradation. Upon target recognition, AGO1 recruits HESO1 and URT1 to uridylate and degrade RISC-cleaved mRNA fragments. The HEN1-catalyzed 3'-end methylation of miRNAs is critical to block mis-uridylation by HESO1/URT1 and 3'-to-5' trimming activities. However, dysfunction of HEN1 or nibbling by SDNs leads to unmethylated miRNAs, which further results subsequent miRNA 3'-end tailing and trimming (pTDTT). Target RNA pairing with a central bulge abrogates the endonuclease activity of AGO1 and induces a conformational change in the protein. This altered state is recognized by HWS, which, in turn, triggers degradation of the AGO1-miRNA-bulged target ternary complex (pTDMD). The figure was created with BioGDPcom[50]. RISC, RNA-induced silencing complex; pTDTT, target-directed miRNA tailing and trimming in plants; pTDMD, target mimicry-directed miRNA degradation in plants.
Even though cumulative research has uncovered multiple layers of mechanisms governing the control of miRNA's stability in plants, several key questions remain to be addressed. First, what is the downstream mechanism of miRNA uridylation? In human cells, the exoribonuclease DIS3L2 degrades uridylated miRNAs; whether the plant ortholog, SUPPRESSOR OF VARICOSE (SOV)[49], plays a comparable role remains to be investigated. Another nonexclusive possibility is that long-tailed miRNAs are more prone to dissociate from miRISC and are subsequently degraded by yet uncharacterized ribonucleases. Second, how does HWS mediate destabilization of the "AGO1–miRNA–target mimicry" ternary complex? It is imperative to investigate the conformational changes and/or post-translational modifications of the AGO1–miRNA–target mimicry complex compared with those of canonical RISC. Moreover, although HWS functions as an E3 ubiquitin ligase, whether the ubiquitin–proteasome system is involved in this process remains unclear. Third, what are the specific roles of SDNs in uridylation and/or HWS-induced miRNA degradation? Addressing these questions through integrated genetic, biochemical, and structural approaches—including mutant analyses, in vitro RISC reconstitution, enzymatic assays, mass spectrometry, and cryo-electron microscopy—will not only elucidate the sophisticated mechanisms of miRNA degradation, but also pave the way for harnessing this knowledge in both fundamental research on regulating gene expression in plants and agricultural applications, such as crop improvement and the development of small RNA-based pesticides.
We apologize to the colleagues whose works were not included due to space limitations. This work was funded by the National Natural Science Foundation of China (32170588, 32470591), the Natural Science Foundation of Guangdong Province (2025A1515011039), the Fundamental Research Funds for the Central Universities (2243200012), and the Shanghai Pilot Program for Basic Research – Fudan University 21TQ1400100 (22TQ014).
-
The authors confirm their contributions to the paper as follows: draft manuscript preparation: Wang X, Guo M, Dong X; literature review: Wang X, Guo M, Dong X, Ren G; manuscript editing: Ren G. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article, as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xiaoyan Wang, Mengwei Guo
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Guo M, Dong X, Ren G. 2026. MicroRNA stability regulation in plants: an ARGONAUTE 1-centric perspective. Epigenetics Insights 19: e007 doi: 10.48130/epi-0026-0004 

MicroRNA stability regulation in plants: an ARGONAUTE 1-centric perspective
- Received: 18 January 2026
- Revised: 17 March 2026
- Accepted: 10 April 2026
- Published online: 21 May 2026
Abstract: MicroRNAs (miRNAs) act as pivotal modulators of endogenous gene expression in eukaryotic organisms. In plants, miRNAs are predominantly loaded onto ARGONAUTE 1 (AGO1). The assembled miRNA-induced silencing complex (miRISC) recognizes the target RNAs through extensive sequence complementarity and mediates target transcript cleavage or translational repression. Although significant advances have been made over the past two decades in elucidating miRNAs' biogenesis and function, the mechanisms governing miRNA turnover and degradation remain relatively poorly understood. This review outlines the current understanding of the molecular mechanisms controlling plant miRNAs' stability from an AGO1-centric point of view. Two aspects of this regulatory process are highlighted: First, AGO1 incorporates and protects miRNA from degradation; second, it recruits various factors to mediate miRNA modification and turnover. We also compare and contrast miRNA catabolism mechanisms between plants and animals. Therefore, future research that unravels how the protective and degradative functions of AGO1 are dynamically balanced will be crucial for a complete understanding of plant miRNAs' homeostasis and active degradation, which may offer novel strategies for crop improvement.
-
Key words:
- miRNA /
- Stability /
- Degradation /
- AGO1 /
- Methylation /
- Uridylation /
- Target mimicry