









-
The Rosa L. genus, belonging to the Rosaceae family, is renowned for its ornamental, medicinal, nutritional, and economic value. China serves as one of the global centers of Rosa distribution, hosting 95 species[1], of which 65 are endemic, accounting for roughly half of the world’s total. Rosa resources in China are particularly rich in traits such as flower color, fragrance, continuous blooming, and resistance to disease, cold, and drought, making them vital germplasm resources for modern rose breeding[2,3].
However, the widespread distribution of Rosa resources and frequent interspecific exchanges complicate their classification. Factors such as natural hybridization, gene introgression, polyploidy, and human influence have led to extensive phenotypic, genotypic, and ecological variation[4,5]. These interconnected factors pose significant challenges to classification, hindering the investigation, assessment, and utilization of Rosa resources. As such, clarifying the systematics, interspecific differentiation, and phylogenetic relationships within the genus is crucial. This effort aids in accurately understanding Rosa’s germplasm resources and constructing core germplasm collections, providing a rich genetic foundation for rose breeding and development of new cultivars, and further promoting the conservation and utilization of Rosa resources.
We have been involved for years in the investigation, classification, and breeding of Rosa resources, culminating in the publication of Genus Rosa L. in China. Over time, the classification of the genus has been refined, updated, and supplemented. This paper offers a summary of the latest developments in the distribution, research, and classification of Rosa in China. It discusses current challenges in Rosa taxonomy and presents new insights into species delimitation, classification methods, and key taxonomic groups, with the goal of exploring the phylogenetic relationships of Rosa taxa and offering references for the conservation and utilization of these resources.
-
The survey of Rosa germplasm resources in China began in earnest in the latter half of the 20th century. Taxonomists undertook efforts to identify, revise, and compile Rosa resources in China, resulting in key publications such as Flora Reipublicae Popularis Sinicae[6] and Flora of China[1]. Since the 1990s, surveys of Rosa resources have been conducted across various regions of China, with evaluations completed in areas including Xinjiang, Qinghai, Gansu, Ningxia, Xizang, Yunnan, Anhui, and Shandong. In parallel, efforts in introduction and hybrid breeding have been initiated[7−16]. The most comprehensive germplasm resource garden for Rosa in China has also been established. Meanwhile, investigations into key distribution areas and important species of the genus have been conducted concurrently (Supplementary Table S1). These studies primarily focused on aspects such as species resource distribution, morphological traits, reproductive strategies, population diversity, and resistance. The findings provide valuable baseline information for understanding Rosa resources in China and form an essential foundation for their conservation and sustainable use.
Update of distribution
-
Rosa resources are widely distributed across the cold temperate and subtropical zones of the Northern Hemisphere, with Central Asia and Southwest Asia identified as the primary diversity centers. To date, no wild Rosa species have been found in the Southern Hemisphere[17−19]. Due to the extensive variation and natural hybridization within the genus, subtle interspecific differences and transitional forms often lead to confusion in classification. As a result, the global number of Rosa species is not fixed, with estimates ranging from 150 to 200 species[20,21].
China is an important global center for Rosa distribution. Although Rosa species are found across various provinces and regions (spanning 75.13° E~133.56° E, 18.89° N~53.04° N, and elevations ranging from 3~5,455 m), the distribution is notably uneven geographically. Species richness is highest within the latitudinal range of 26.19° N~34.29° N, and at mid to high elevations (956~3,518 m). Wild Rosa resources primarily increase from Southeastern China towards the southwest and northwest. Notably, the Southwestern Hengduan Mountains are recognized as the diversity hot spot for the genus, while the Tianshan, Altai, and Changbai Mountains are local concentration areas. The Qinling Mountains, acting as a transition zone between the north and south, also house rich Rosa resources and considerable variation[22−25]. Combining molecular evidence[26], researchers identified two major diversity centers of Rosa in China: (1) The northwest center, which includes many ancestral traits and more early-diverging lineages; (2) The southwest center, where favorable hydrothermal conditions and minimal seasonal climate variation offer optimal conditions for the propagation and differentiation of Rosa populations (Fig. 1). The resource distribution of different sections within Rosa varies (Supplementary Table S2).
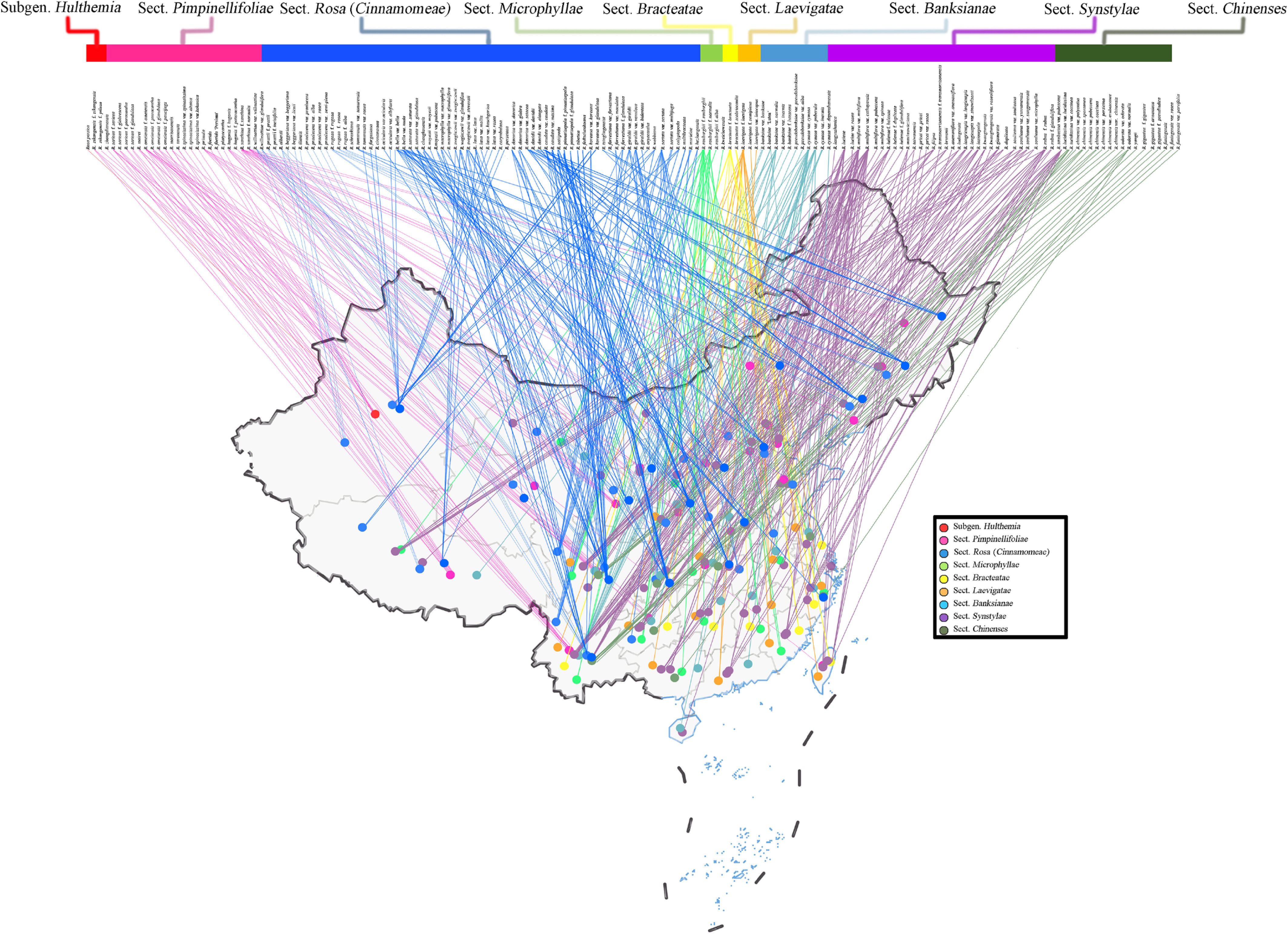
Figure 1.
Main distribution regions of Rosa taxa of different sections in China. Note: distribution is shown at the provincial level. Data were aggregated from the Chinese Virtual Herbarium (CVH), the Global Biodiversity Information Facility (GBIF), and published records. The complete species list and detailed distribution data are provided in Supplementary File 1.
Among these environmental factors, moisture is the most significant factor influencing species richness in this genus[27−29]. This distribution pattern is likely a result of a combination of the genus's evolutionary history, physiological adaptations, and human activity[30,31]. It is important to note that eight species of Rosa plants are currently listed in China's National Key Protected Wild Plants List (second-class). While most Rosa species are not endangered in terms of population size, protecting their habitats should be prioritized over merely conserving the species themselves.
-
Since the 18th century, botanists have worked to investigate the germplasm resources of Rosa. However, significant disagreements persist among scholars regarding the scope and classification of the genus. The most widely accepted classification system for Rosa today is the one published by Rehder in 1940s[20]. Although Rosa resources in China are abundant, classification research has lagged behind that of other countries.
Yu[6] developed one of the earliest classification systems for Rosa in China in Flora Reipublicae Popularis Sinicae (Vol. 7). This system categorizes 82 known native and introduced species of Rosa (including four alien species) into two subgenera, nine sections, and seven series. The genus Rosa was first divided into subgen. Hulthemia and subgen. Eurosa. Subgen. Eurosa was further divided into Sect. Pimpinllifoliae (Ser. Sericeae and Ser. Spinosissimae), Sect. Cinnamomeae (Ser. Beggeriana, Ser. Cinnamomeae and Ser. Webbianae), Sect. Synstylae (Ser. Multiflorae and Ser. Brunonianae), Sect. Chinenses, Sect. Banksianae, Sect. Laevigatae, Sect. Bracteatae, Sect. Microphyllae, and Sect. Rosa (now referred to as Sect. Gallicae) (Table 1). Ku & Robertson[1] revised and included 95 species (including two alien species) of wild Rosa native to China in the Flora of China, using Yu's classification system. They retained two subgenera, eight sections, and seven series while removing Sect. Rosa. This system has been adopted by various provincial floras and major researchers studying Rosa in China[32].
Table 1. Comparison of Rosa taxa in Flora Reipublicae Popularis Sinicae, Flora of China, and Genus Rosa L. in China.
Subgenus/section FRPS FOC GRLIC Subgen. Hulthemia One species One species One species Subgen. Eurosa Sect.
Pimpinellifoliae18 species, two varieties, eight forms, one species noted in discussion 17 species, three varieties, eight forms 13 species, two varieties, nine forms, one cultivar Sect. Rosa (Sect.
Cinnamomeae)31 species, 15 varieties, four forms,
four species noted in discussion36 species, 16 varieties, two forms 39 species, 25 varieties, five forms, six cultivars Sect. Microphyllae Three species, one form Three species, one deformation Two species, two forms Sect. Bracteatae One species, one variety One species, one variety One species, one variety Sect. Laevigatae One species, one form One species, one form One species, one variety, one form Sect. Banksianae Two species, two varieties, two
forms, 1 hybrid2 species, two varieties, two forms Three species, six varieties, one form, three cultivars Sect. Synstylae 18 species, 10 varieties, one form,
three species noted in discussion29 species, 11 varieties, three forms 19 species, 10 varieties, four forms, 22 cultivars Sect. Chinenses Three species, five varieties Three species, five varieties Six species, eight varieties, four forms, 28 cultivars Sect. Gallicae Four species / / FRPS is the abbreviation for Flora Reipublicae Popularis Sinicae; FOC is the abbreviation for Flora of China; GRLIC is the abbreviation for Genus Rosa L. in China. Luo et al.[33] included 86 species of Rosa in the monograph Genus Rosa L. in China, which encompasses all species recorded in Flora Reipublicae Popularis Sinicae and Flora of China, along with some newly published and newly revealed species, varieties, and forms. Some previously recognized species have been reclassified as synonyms, and 60 important cultivars related to classification have been incorporated. A key taxonomic revision in this work was the subdivision of Sect. Chinenses into Ser. Chinenses Luo et Yang and Ser. Odoratae Luo et Yang, to more accurately reflect species relationships. The primary distinction between these two series lies in their morphology: Ser. Chinenses comprises climbing or loose shrubs with 3–5 (7) leaflets and pubescent or glabrous new shoots producing ovoid or pyriform fruits, while in contrast, Ser. Odoratae are climbers with consistently glabrous new shoots, a higher leaflet count of 5–9, and typically globose or depressed globose fruits. This new classification resulted in a total of two subgenera, eight sections, and nine series (Supplementary Table S3). A new identification key for Rosa in China has been developed (Supplementary Table S4). This book significantly contributes to the conservation and utilization of Rosa resources, providing valuable reference materials for research and breeding of this important economic crop.
Overview of phylogenetic studies
-
Traditionally, extensive classification work on Rosa has been fundamentally based on phenotypic traits. For example, Hung & Wang[34] found that stipule shape, gynoecium, and the shape of terminal leaflets are key diagnostic characters for Rosa in Taiwan. Beyond macroscopic morphology, palynology and cytology have also proven valuable for taxonomic studies and in investigating evolutionary relationships. Rosa pollen morphology and exine pattern variation (e.g., perforate exine patterns representing more primitive types) provide significant taxonomic resolution[35,36]. Cytologically, Rosa taxa have a chromosome base number of x = 7. Frequent polyploidy (with observed chromosome counts of 2n = 3×, 4×, 5×, 6×, 8×, and 10×) and aneuploidy further complicate the genus[37]. By investigating karyotypes, researchers like Fang et al.[37] have hypothesized ancestral progenitors for species such as R. praelucens.
Driven by biotechnological advancements, molecular phylogenetics has continuously reshaped Rosa taxonomy. Early molecular studies primarily utilized DNA regions like ITS and matK to establish foundational phylogenetic trees, corroborating the morphological affinities between Sect. Synstylae and Sect. Chinenses[38]. Subsequently, molecular markers such as RAPD[39], AFLP[40], SSR[41,42], and CDDP[25,43] emerged as effective tools for evaluating genetic diversity and identifying phylogenetic relationships among numerous Rosa populations at a finer scale.
In recent years, the rapid development of high-throughput sequencing has transitioned Rosa phylogenetics into the genomic era, resolving many long-standing taxonomic conflicts. Phylogenetic analyses using complete chloroplast genomes have tested traditional morphological subgroupings[44] and confirmed close relationships among similar species[45,46]. More profoundly, nuclear genomic sequence analyses have become widespread. A recent phylogenomic study using extensive nuclear loci established the first robust backbone phylogeny of Rosa, revealing multiple hybridization events, including the hybrid origin of Sect. Caninae[47]. Furthermore, the publication of several Rosa reference genomes[5,48−52] has provided a crucial foundation for in-depth research. For instance, utilizing the R. chinensis genome, Wang[53] analyzed the genetic structure of the R. brunonii complex, shedding light on its speciation and historical population dynamics. Cheng et al.[26] performed population genomic analyses of 215 Rosa accessions, reconstructing nuclear and plastid phylogenies that offer molecular insights into the taxonomy of the genus.
-
Phylogenetic research in recent years suggests that the genus Rosa is a monophyletic group[4,54], but there are still numerous issues and challenges in the infrageneric classification of Rosa.
Limited reference data, synonyms, and homonyms
-
Descriptions of species in existing flora references are often outdated, lacking sufficient detail or specificity in characteristics. Quantitative traits are often narrowly defined, qualitative traits are frequently absent, and distribution records are incomplete. These gaps complicate specimen identification and field surveys, creating ambiguities in taxonomic references. Many species within Sect. Synstylae and Sect. Rosa (Sect. Cinnamomeae), for example, are not fully described regarding traits like flower number, presence and density of trichomes, or leaflet count[55,56].
A prime example of such taxonomic confusion is the recently described R. austrotibetana. Due to deceptively overlapping phenotypic traits—such as sparsely pubescent leaflets—it was historically misidentified as R. filipes in herbarium records; its distinct specific status was only uncovered when integrated multi-organ and molecular analyses exposed the severe limitations of the original morphological diagnosis[57].
Early introductions and exchanges of Chinese Rosa species by foreign botanical gardens also contributed to narrow, unrepresentative descriptions that complicate classification[2].
Incomplete research materials and unsystematic methods
-
Accurate and comprehensive research materials are essential for establishing a robust and credible classification system for Rosa. Currently, however, many studies are limited to specific sections, resulting in insufficient sample sizes and poor species coverage. Consequently, the taxonomic conclusions often lack genus-wide representativeness. Furthermore, genomic resources for Rosa remain scarce and fragmented. Although high-quality chromosome-scale assemblies exist for a few species[52−58], the genus lacks a comprehensive pangenome reference that can capture the full spectrum of interspecific genetic variation. As demonstrated in other plant groups, pangenome analyses can uncover substantial gene presence/absence and structural variations missed by single-reference genomes[59,60]. Without such integrated genomic evidence and broader sampling, species boundaries within Rosa remain poorly defined.
Conventional taxonomic frameworks for Rosa relied on diverse methodologies, including morphology, quantitative analysis, palynology, cytology, and basic molecular biology. While these approaches have laid the foundation for understanding the genus, their resolution and diagnostic accuracy frequently falter when confronted with the inherent biological complexity of Rosa[61−65]. Recent integrative studies have underscored that even high-throughput, unbiased morphometric analyses frequently fail to identify stable diagnostic traits in morphologically cryptic species when applied in isolation[66]. Furthermore, the evolutionary history of Rosa complicates matters at the genetic level. Phylogenomic analyses have revealed extensive reticulate evolution and ancient hybridization[47]. These complex evolutionary processes lead to widespread incomplete lineage sorting (ILS) and historical introgression, causing frequent and profound discordance between plastid-based and nuclear phylogenies[67,68]. Ultimately, traditional molecular markers—which capture only a fraction of the genome—and morphology simply lack the resolving power to disentangle such intricate evolutionary dynamics.
Additionally, as more methods are applied, the inconsistencies between results derived from divergent methodologies have become increasingly apparent. The frequent mismatch between phenotypic traits and underlying genetic structure underscores the peril of relying on any single line of evidence[66]. To overcome these limitations, advanced genomic tools—such as pangenomics—offer a transformative solution. For example, through a super-pangenome analysis of 61 tomato varieties, Yu et al.[69] successfully resolved long-standing taxonomic controversies. By capturing structural variations missed by linear genomes, they recalibrated Solanum galapagense as an ancestral lineage to wild tomatoes, fundamentally overturning traditional views of tomato evolution. This success in disentangling complex evolutionary histories demonstrates the immense potential for clarifying species boundaries in Rosa. Moving forward, resolving the complex evolutionary history of Rosa requires an integrated taxonomic framework. This framework must combine high-resolution genomic infrastructures, particularly pangenomes, with phenomics, cytology, and ecological niche modeling to achieve a holistic understanding of Rosa diversity.
Controversy over grouping and species boundaries
-
Disputes exist over the grouping of sections Chinenses and Synstylae in China’s Rosa taxonomy. Multiple studies indicate a close phylogenetic relationship between these sections, with Chinenses nested within Synstylae, jointly forming a monophyletic group on phylogenetic trees[4,70−72]. More recent phylogenomic research has provided evidence for this relationship, leading the authors to recommend merging Sect. Indicae (syn. Chinenses) with Sect. Synstylae[47].
The delineation of species has also been a source of significant controversy, like R. graciliflora, R. pseudobanksiae, R. praelucens, R. sericea, and R. omeiensis. R. graciliflora, for example, has been traditionally placed in Sect. Pimpinellifoliae due to its lack of bracts[6]. However, field observations reveal that flowers on the same plant can vary, exhibiting either bracted or non-bracted forms. Morphological characteristics[73], vitamin content in hips[74], and molecular evidence[75,76] all suggest that R. graciliflora should be reassigned to Sect. Rosa (Sect. Cinnamomeae). Whether it serves as a transitional species between Sect. Pimpinellifoliae and Sect. Rosa (Sect. Cinnamomeae) remains an open question requiring further investigation.
While phylogenomics has clarified many long-standing issues, it has also uncovered deeper complexities and presented new challenges. A core challenge today is addressing the conflict between phylogenetic signals derived from different genomes—namely, the nuclear and plastid genomes.
-
The taxonomy of Rosa remains a dynamic field of study, marked by debates over species boundaries, the application of various research methods, and the handling of taxonomic complexities. This section synthesizes key insights into the classification of the genus, spanning theoretical principles, methodological advancements, and case studies on specific taxonomic groups. By addressing these topics, the discussion aims to deepen understanding of Rosa diversity and foster systematic strategies for its conservation and utilization.
How are species boundaries defined in genus Rosa?
-
A primary challenge in Rosa taxonomy is the consistent delimitation of wild species from the vast number of cultivars, a distinction often blurred by centuries of cultivation and frequent hybridization[77]. To address this, we propose a more rigorous operational framework. Within this framework, a 'species' should be strictly defined as a wild type that meets several criteria: it must exhibit stable traits, exist in natural populations with a defined geographic distribution, and typically possess single-petaled flowers (allowing for occasional staminodes). Conversely, plants with double or multi-petaled flowers that lack a wild distribution should be classified as cultivars.
Based on this foundational distinction, we argue that interspecific delimitation must primarily rely on morphological evidence, requiring at least two distinct and corresponding character differences between putative species. This morphological baseline should then be tested and refined using supplementary evidence from cytology, palynology, and molecular systematics. Recent integrative taxonomic studies on morphologically confounding Rosa species within Sect. Pimpinellifoliae have demonstrated that even high-throughput, unbiased morphometric analyses based on large and representative sample sizes were insufficient to identify effective diagnostic traits when applied in isolation[66]. This finding directly echoes the problem previously raised that existing flora descriptions are often outdated, with narrowly defined quantitative traits and insufficient qualitative detail. It underscores that the morphological baseline must be cross-validated by complementary evidence, as the frequent mismatch between phenotypic traits and underlying genetic structure highlights the peril of relying on any single line of evidence[66].
Addressing the question of which traits should serve as primary criteria, phylogeny-based morphological evolution studies in Rosa sections Synstylae and Chinenses have identified six key characters with strong taxonomic resolution: stipule margin ornamentation, petal abaxial surface indumentum, style hairiness, leaflet number, leaflet texture, and leaflet abaxial hairiness. These characters should serve as primary diagnostic traits, while variable features such as flower colour or the presence of abaxial glands—which may vary within the same species or even the same population—should be treated as secondary or auxiliary traits[57].
Beyond morphology, we advocate integrating two additional evidence dimensions within our operational framework. First, genomic differentiation from closest relatives—evidenced by population structure analyses, fixation index (FST) values, and/or private alleles—is essential for detecting cryptic species that are morphologically similar but genetically divergent, as well as for merging morphologically distinct entities that may represent ecotypes or local variants rather than true species. Second, geographic and ecological niche assessments provide independent evidence for species status, as environmental heterogeneity plays a significant role in driving species differentiation. Ecological niche modelling and reciprocal transplant or common-garden experiments can help determine whether morphologically and genetically distinct populations represent genuinely separate species or merely intraspecific ecomorphs adapted to different habitats[66].
Therefore, resolving interspecific boundaries in Rosa requires: (1) establishing a clear hierarchy of primary vs secondary diagnostic traits and quantifying the standards for their assessment (e.g., the degree of difference in quantitative traits; the presence or absence of qualitative traits); and (2) subjecting these morphological hypotheses to cross-validation using genomic and ecological data. Only when a taxon is supported by at least two of the three evidence dimensions—morphology, genomics, and ecology—should it be recognised as a valid species. This standard mirrors the 'species-as-hypotheses' framework advocated in modern integrative taxonomy, wherein each species hypothesis must be explicitly testable and potentially falsifiable by independent data[78−80].
Establishing operational criteria for species delimitation is only the first step; the next challenge is to identify an array of analytical methods capable of generating the evidence required to validate these criteria in a genus as evolutionarily complex as Rosa.
What research methods are suitable for Rosa taxonomy?
-
The species boundaries within the genus Rosa are notoriously difficult to delineate due to a combination of complex biological factors, including frequent interspecific hybridization, widespread polyploidy, unique reproductive systems like the apomixis found in Sect. Caninae, and a long, intricate history of cultivation that blurs the lines between wild taxa and early cultivars. These challenges mean that relying on a single line of evidence, whether morphological or molecular, is often insufficient and can lead to conflicting taxonomic conclusions.
To address this challenge, some researchers advocate integrating various forms of evidence to delineate species boundaries from multiple perspectives, giving rise to the concept of integrative taxonomy[78,79]. This approach has proven effective for resolving hybridization issues, describing new taxa, studying cryptic species, and addressing challenging interspecific boundaries in previously described species[80]. For example, Li et al.[81] conducted a phylogeographic study of 590 Rhodiola Sect. Trifida samples and concluded that taxonomy based on variable leaf traits is unreliable, as it strongly conflicts with genetic data, showing that distinct species share genotypes, and that R. alterna and R. sacra are indistinguishable. Batista et al.[82] employed molecular phylogenetics, morphometrics, and data on morphological, ecological, and geographic distribution to define species within the Cleistes rosea complex (Orchidaceae).
Currently, studies employing multiple taxonomic methods for species identification on Rosa already exist. Deng et al.[83] demonstrated significant differences between R. tomurensis and R. laxa through analyses of geographic distribution, morphological traits, palynological features, karyotype, and genomic features, confirming R. tomurensis as a distinct species. While such studies represent a crucial step forward, we propose that a more comprehensive integrative framework is required to operationalise the aforementioned three-dimensional framework.
To generate robust morphological data, standardised measurement protocols are essential. Recent integrative studies employing high-density phenotyping demonstrate the power of systematic phenomic data for species delimitation, revealing that phenomic and genomic data must be analysed on an equal footing[84]. For genomic validation, whole-genome resequencing at the population level provides the most powerful approach, enabling calculation of fixation indices (FST), identification of private alleles, and detection of population structure. When whole-genome data are unavailable, targeted capture of conserved orthologue sets—such as the RosCOS markers employed by Jeon et al.[55] to analyse 137 East Asian Synstylae accessions—provides a cost-effective alternative. Given the prevalence of hybridisation and incomplete lineage sorting in Rosa[47,67,68], coalescent-based species tree methods and phylogenetic network analyses are essential for distinguishing shared ancestral polymorphism from post-divergence gene flow. For ecological assessment, ecological niche modelling using occurrence records and environmental variables can quantify niche differentiation between putative species. Reciprocal transplant and common garden experiments can disentangle genetic differentiation from environmentally induced plasticity.
These three evidence dimensions must be synthesised through explicit analytical workflows. Recent revisions combining quantitative morphological traits, principal component analysis, karyotyping, and phylogenetic reconstruction demonstrate how cryptic species can be uncovered within a unified framework[85]. Even within this framework, certain Rosa lineages exhibit such extensive morphological intergradation and low genetic differentiation that conventional species-level treatment becomes impractical, suggesting that the species complex concept may offer a more biologically realistic classification.
Taxonomic implications of complexes in Rosa
-
The extensive and often continuous morphological variation within Rosa challenges traditional species delimitation. To better reflect the biological reality of these closely related and intergrading taxa, we advocate for the application of the 'species complex' concept, where a group of morphologically diverse, but genetically cohesive taxa are treated as a single, variable entity.
The R. sericea complex serves as a well-documented example. Although the taxa within this group (such as R. sericea and R. omeiensis) show significant morphological heterogeneity, population genetic studies have revealed low genetic divergence and a weak phylogenetic structure among them[86]. This pattern of morphological diversity coupled with genetic uniformity suggests these are not distinct species but rather a single, polymorphic complex that underwent recent, rapid radiation. Supported by compelling genetic evidence, we treat these taxa as a single cohesive unit, and we propose adopting the name 'R. omeiensis complex' (rather than the 'R. sericea complex'), because this name better reflects its morphological diversity.
The species complex framework presented here offers a conceptual response to the species-boundary controversies previously catalogued. Rather than forcing intergrading taxa into artificial species-level bins, this framework acknowledges that morphological diversity can coexist with genetic cohesion when divergence is recent, and gene flow persists.
However, adopting the species complex concept carries inherent risks: collapsing distinct morphotypes may obscure substructure driven by local adaptation or partial reproductive isolation. For instance, within the R. omeiensis complex, subdivisions based on indumentum and foliolate traits (Table 2) hint at underlying ecological or genetic heterogeneity. Whether these subgroups represent incipient speciation, ecotypic variation, or mere phenotypic plasticity remains an open empirical question. Resolving this requires shifting focus from broad phylogenomic patterns to population-level processes; specifically, evaluating gene flow uniformity and the correlation between ecological gradients and genetic clines.
Table 2. Potential complexes among Rosa taxa.
Name First-level classification Second-level classification Included taxa R. omeiensis complex Densely-indumentum type R. Sericea, R. sericea f. glandulosa, R. mairei Glabrous or sparsely indumentum type Multifoliolate subtype R. Omeiensis, R. omeiensis f. glandulosa, R. omeiensis f. pteracantha Few-foliolate subtype R. omeiensis f. paucijuga, R. morrisonensis, R. taronensis R. sertata complex Single-flowered,
small-leaf typeSingle-flowered, few-leaflet, small-leaf subtype R. farreri Single-flowered, multi-leaflet, small-leaf subtype R. graciliflora Multi-flowered,
large-leaf typeMulti-flowered, few-leaflet,
large-leaf subtypeR. banksiopsis Multi-flowered, multi-leaflet, large-leaf subtype R. sertata var. multijuga R. multiflora complex R. multiflora, R. multiflora var. cathayensis, R. multiflora var. pubescens, R. multiflora var. spinosa R. soulieana complex R. soulieana, R. soulieana var. yunnanensis, R. soulieana var. Sungpanensis, R. soulieana var. microphylla R. lucieae complex R. lucieae, R. kwangtungensis, R. taiwanensis, R. pricei R. brunonii complex R. Brunonii, R. helenae, R. filipes, R. ludingensis, R. glomerata etc. Importantly, this complex-level perspective also provides taxonomic space to recognize infraspecific diversity without inflating species counts. The 22 new varieties and forms described from our field investigations (Table 3) fit naturally within this framework: rather than erecting new species for every morphological variant, we can accommodate observed diversity at the varietal level within broadly defined complexes. This approach aligns with the growing recognition in plant taxonomy that species-level inflation obscures true evolutionary relationships and complicates conservation prioritisation.
Table 3. Taxa of Rosa discovered and recorded in Genus Rosa L. in China.
Section Species Variety Form Cultivar Sect. Pimpinellifoliae R. hugonis f. pteracantha Sect. Rosa (Sect. Cinnamomeae) R. tomurensis var. Tomurensis[83] R. praelucens var. alba R. prattii f. incisifolia R. rugosa 'Danban Danfen' R. praelucens var. rosea R. praelucens var. semi-plena R. tomurensis var. rosea R. laxa var. rosea R. forrestiana var. maculata[87] Sect. Banksianae R. banksiae var. inermis R. banksiae 'Wuci Chongbanbai' R. pseudobanksiae var. alba R. cymosa var. inermis Sect. Synstylae R. multiflora var. pubescens R. multiflora 'Yinbei Taohong Fentuan' R. multiflora var. spinosa R. multiflora 'Baibei Zihua Fentuan' Sect. Chinenses R. yangii[88] R. chinensis var. pubescens R. gigantea f. armeniaca R. chinensis 'Zhai Ye' R. funingensis f. funingensis[89] R. chinensis var. coccinea R. chinensis 'Xian E' R. chinensis var. persicina R. chinensis 'Banchongban Xian-e' R. chinensis var. erubescens R. chinensis 'Zi Hong' R. chinensis var. multijuga R. chinensis 'Chongban Taohong' R. lucidissima var. coccinea R. chinensis 'Shaoci Meihong' R. odorata var. normalis R. chinensis 'Xiao Ye' R. funingensis var. rosea R. chinensis 'Duo Tou' R. funingensis var. parvifolia R. chinensis 'Chongban Xinghong' R. × odorata Sweet 'Tao Yun' R. × odorata Sweet 'Fenhong Mudan' R. × odorata Sweet 'Xiaoye Fenhua' R. × odorata Sweet 'Rou Fen' A recent, robust phylogenomic framework for the entire genus[26] provides a powerful tool for identifying and evaluating such complexes. The nuclear SNP tree from that study, for instance, generally supports the close phylogenetic relationships and recent divergence among taxa within several of our proposed complexes (e.g., the R. soulieana and R. brunonii complexes). Building on the available molecular evidence and our extensive morphological observations, we propose a working framework of several potential species complexes within Chinese Rosa (Table 2). We emphasize that these groupings function as testable hypotheses; determining whether they should be maintained, subdivided, or merged will ultimately require dedicated studies integrating the aforementioned population genetics with broader phylogenomic contexts.
Proposals and publications of new taxa in Rosa
-
Over years of field survey and research conducted during the preparation of the book Genus Rosa L. in China, we identified several new taxa of Rosa, 4 already published (three species and one variety), and 22 newly described in the book (19 varieties and three forms; Table 3). In addition, 17 formerly unrecorded cultivars were also described, most of which exist in a semi-wild state and remain underutilized. Based on the species definition principles discussed before, these cultivars have been designated accordingly. They represent critical intermediate materials for Rosa breeding and classification, broadening perspectives on the evolution and taxonomy of the genus.
Phylogenetic relationships between Sect. Chinenses and Sect. Synstylae
-
During our investigations, we found that Sect. Chinenses and Sect. Synstylae are closely related, displaying similarities in stipules, inflorescences, and hip shapes. Moreover, some species within Sect. Chinenses also possess connate or semi-connate styles. The overlapping distributions of these two sections create opportunities for natural hybridization. Species in Sect. Synstylae such as R. multiflora, R. lucieae, R. longicuspis, R. sambucina (R. henryi), R. brunonii, R. rubus, etc., are all sympatric taxa with taxa in Sect. Chinenses. For example, R. funingensis has been suggested as a natural hybrid between Sect. Chinenses and Sect. Synstylae[89]. Multiple studies have shown that materials from Sect. Chinenses and Sect. Synstylae always jointly form a monophyletic group in phylogenetic trees, indicating the possibility of introgression[44,38]. A phylogenomic study using nuclear single-copy genes strongly supports that Sect. Chinenses (syn. Indicae) and Sect. Synstylae form a well-supported clade[47]. More recent genomic research provides even stronger evidence, showing that the genetic differentiation between Sect. Chinenses and Sect. Synstylae is extremely low (FST < 0.05), suggesting that Sect. Chinenses likely originated from Sect. Synstylae in a relatively recent divergence event. Accordingly, it has been proposed that the two sections be merged into a single section, with each retained as a separate series[26].
Synthesis and future perspectives
-
The challenges catalogued before—ambiguous species boundaries, fragmented methodological practices, and unresolved controversies surrounding sectional classification—are formidable but not insurmountable. They demand a deliberate shift toward integrative, quantitative, and collaborative taxonomy. The operational species-delimitation framework, the integrative methodological workflow, the species-complex hypotheses, the 22 newly described taxa, and the evidence supporting the merger of Sect. Chinenses and Sect. Synstylae collectively represent a research-based response to the diagnostic problems identified at the outset of this review.
Translating these insights into a stable, widely accepted classification system will require a sustained effort across three strategic fronts. First, a broader and more representative sample across all sections and geographic regions is essential. Many previous studies have been limited to specific sections with insufficient sample sizes and poor species coverage, compromising the genus-wide representativeness of their conclusions. Future efforts should prioritize population-level sampling, the expansion of genomic resources—particularly the development of a comprehensive pangenome reference that captures interspecific genetic variation—and the establishment of standardized morphological measurement protocols to ensure quantitative and reproducible phenotypic data. Second, integrative analytical approaches must be strengthened and standardized. Given the pervasive conflict between plastid and nuclear phylogenetic signals in Rosa, future studies should routinely employ coalescent-based species-tree methods and phylogenetic network analyses to distinguish shared ancestral polymorphism from post-divergence gene flow. Machine learning provides a promising complementary tool in this context: by training models on large-scale phenotypic datasets, researchers can objectively identify diagnostic characters that are often obscured by continuous morphological variation, quantify subtle interspecific differences within species complexes, and develop predictive models to assist in species delimitation[90]. Third, the theoretical frameworks proposed herein require rigorous empirical validation. The species-complex hypotheses (Table 2), as well as the proposed merger of Sect. Chinenses and Sect. Synstylae, should be treated as working hypotheses subject to verification through dedicated population-genetic studies, hybridization assessments, and common-garden or reciprocal-transplant experiments. Determining whether morphological variants within complexes represent incipient speciation, ecotypic differentiation, or phenotypic plasticity remains a critical unresolved question.
-
This review systematically summarizes the progress in the distribution, survey, and classification of Rosa resources in China. Decades of field investigations have established that Chinese Rosa species are concentrated in two primary diversity centers—the southwestern Hengduan Mountains and the northwestern Tianshan–Altai region, where favorable hydrothermal conditions and habitat heterogeneity support the highest species richness. The trajectory of Rosa taxonomy in China has evolved from early morphology-based systems, exemplified by Flora Reipublicae Popularis Sinicae and Flora of China, to increasingly integrative approaches that combine palynology, cytology, molecular phylogenetics, and phylogenomics.
Despite these advances, substantial challenges remain. Existing flora references frequently provide outdated or incomplete species descriptions, and many studies continue to suffer from limited sample sizes and narrow taxonomic coverage. Methodological inconsistencies—particularly the reliance on single lines of evidence—have produced conflicting results regarding sectional boundaries and species delimitation. Phylogenomic studies have clarified certain long-standing controversies, such as the close relationship between Sect. Chinenses and Sect. Synstylae, yet have also revealed deeper complexities arising from hybridization, incomplete lineage sorting, and nuclear-plastid phylogenetic discordance.
A growing body of evidence supports the adoption of more rigorous operational frameworks for Rosa taxonomy. Integrative approaches that cross-validate morphological, genomic, and ecological data offer the most promising path forward for resolving ambiguous species boundaries. The application of the species-complex concept provides a pragmatic alternative for accommodating morphologically diverse but genetically cohesive groups, and the continued expansion of genomic resources—including population-level resequencing and pangenome references—will be essential for disentangling reticulate evolutionary histories. The documentation of previously unrecognized taxa and semi-wild cultivars further underscores that the morphological diversity of Chinese Rosa remains incompletely characterized.
Moving forward, the resolution of Rosa taxonomy will depend on sustained collaborative efforts to expand representative sampling, standardize methodological protocols, and empirically validate proposed taxonomic hypotheses. Accurate classification and systematic clarification are essential prerequisites for the conservation of Rosa germplasm, the construction of core collections, and the continued genetic improvement of roses for horticultural and commercial applications.
-
All data generated or analyzed during this study are included in this published article and its Supplementary Information files. Additional data are available from the corresponding author upon reasonable request.
-
The authors confirm their contributions to the paper as follows: conceptualized the study and designed the overall framework: Luo L; collected and analyzed the literature: Zhang C, Pan H; provided supporting data: Yu C, Yang Y; drafted the manuscript: Zhong J, Zhang C; revised and refined the manuscript: Luo L, Zhang Q, Yuan T, Bai J; created the visualizations: Lyu P, Zhong J. All authors reviewed the results and approved the final version of the manuscript.
-
This research was funded by the Major Science and Technology Special Projects of Yunnan Province China, (Grant No. 202502AE090068), the Fundamental Research Funds for the Central Universities, (Grant No. QNTD202306), the National Key Research and Development Project of China, (Grant No. 2023YFD1200105), the National Key Research and Development Project of China, (Grant No. 2019YFD1001001), the Horizontal Project commissioned by Enterprises and Institutions for Scientific and Technological Projects, (Grant Nos 2023-HXFW-428, SSTLAB-2023-1).
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
accompanies this paper online at: https://doi.org/10.48130/opr-0026-0012.
-
# Authors contributed equally: Junfeng Zhong, Chenjie Zhang, Peifeng Lyu
- Supplementary Table S1 Surveys of Rosa resources across Chinese regions. Focused research on specific Rosa species in China.
- Supplementary Table S2 Distribution and altitudinal ranges of Rosa sections in China.
- Supplementary Table S3 Taxonomic comparison between the Flora Reipublicae Popularis Sinicae (FRPS) and the Flora of China (FOC). Taxonomic comparison between the Flora of China (FOC) and Genus Rosa L. in China (GRLIC).
- Supplementary Table S4 Identification key to Rosa in China.
- Supplementary File 1 Species list and documented occurrence of Rosa in China.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhong J, Zhang C, Lyu P, Pan H, Yu C, et al. 2026. Genus Rosa L. in China: advances in resources and taxonomy. Ornamental Plant Research 6: e023 doi: 10.48130/opr-0026-0012 

Genus Rosa L. in China: advances in resources and taxonomy
- Received: 17 January 2026
- Revised: 12 April 2026
- Accepted: 22 April 2026
- Published online: 05 June 2026
Abstract: Rosa is considered one of the most challenging and difficult groups in taxonomy and phylogenetics due to its wide distribution, extensive variability, and high hybridization potential, which leads to unclear interspecific boundaries. This paper summarizes and analyzes the progress in the distribution, survey, and classification of Rosa resources in China, based on years of research on the genus and a review of relevant literature. The latest distribution data for Rosa in China was updated, highlighting two key diversity centers where species richness is highest—regions with favorable hydrothermal conditions, minimal seasonal climatic variation, and high habitat heterogeneity. Additionally, 22 previously unpublished Rosa taxa (19 varieties and three forms) and 17 significant cultivars have been introduced. A new identification key for the genus Rosa in China has also been given. We also address issues in the classification of Rosa, offering fresh perspectives on interspecific boundaries, classification methods, and key taxonomic groups. This provides theoretical support for the identification, classification, and evolutionary research of Rosa species, and offers valuable references for the conservation and utilization of Rosa resources.
-
Key words:
- Rosa taxonomy /
- Resource distribution /
- New taxa proposal /
- Rosa complex