









-
Antibiotic resistance has become a pervasive concern worldwide, posing a significant threat to public health[1]. It is estimated that 4.71 million deaths in 2021 were associated with bacterial antibiotic resistance[2]. Antibiotic resistance genes (ARGs) are the fundamental determinants that encode antibiotic resistance, which confer on bacteria the ability to survive antibiotic treatment. Although ARGs are ancient and ubiquitous in the natural environment, decades of inappropriate consumption of antibiotics and non-antibiotic chemicals in humans and animals have largely facilitated the widespread dissemination of ARGs across the three sectors of One Health—humans, animals, and the environment[3]. Due to improper waste disposal and wastewater discharge, polluted environments such as water and soil serve as key reservoirs and conduits for ARG dissemination[4]. Indeed, ARGs are not only found in pathogens but also widely distributed among commensal and environmental microbiota[5,6].
Among various environments, surface water has drawn significant attention for its role in disseminating ARGs. Surface waters, especially rivers, which serve as dynamic connectors between diverse habitats through water flow, are highly vulnerable to pollution from various known and emerging sources, including wastewater discharged from residential communities[7], hospitals[8], industries[9], as well as livestock[10] and crop farms[11]. A global-scale study[12] of antibiotic and non-antibiotic pharmaceuticals in 258 rivers across 137 geographic regions has revealed the occurrence of a diverse range of pharmaceuticals in the aquatic environment. The most contaminated sites are typically located near urban areas, especially in low- and middle-income countries (LMICs) that lack adequate wastewater and waste management infrastructure. Such selective pressures have driven the emergence and dissemination of ARGs and their host bacteria in the water environment worldwide[13−16]. River surveillance in different regions has revealed that their resistome profiles are largely shaped by the level of urbanization and the presence of wastewater discharge. For example, ARGs and antibiotic-resistant bacteria in large subtropical rivers—the Pearl River[14] and the Yangtze River[16] in China—were highly associated with human- and animal-related wastewater discharge. A three-year longitudinal metagenomic monitoring of rivers in Hong Kong revealed that resistome patterns are significantly correlated with fecal pollution[13]; rivers in LMICs lacking hygiene infrastructure are compounded by more diverse sources and pollutants, as observed in India[17], Africa[18], and Latin America[19]. This might create an ideal environment for enriching ARGs through mechanisms such as selection and co-selection[4].
Various types of wastewater act as potential pollution sources in urban water environments. Unlike domestic wastewater—the most well-known source—hospital wastewater has a much higher load of multi-resistant pathogens and ARGs, especially those of clinical importance[20−23]. This has been well exemplified by the consistent differences in ARG profiles between hospital and residential wastewaters across European countries (e.g., Denmark, Spain, and France)[22,24], even though they have similar microbial community structures. The level of β-lactamase genes in hospital wastewater was found to be over 5-fold[22], even 26-fold[24] higher than that in residential wastewater. A similar contrast resistome was also observed between the two wastewater types in Asian countries, such as Singapore[25] and China[26]. In particular, hospital wastewater quality seems to vary by region and hospital type, as observed in China, probably due to differences in wastewater management[27]. Water pollution by wastewater discharge, especially hospital wastewater, has thus become a critical public concern.
The management of hospital wastewater, therefore, has garnered significant global attention. The WHO guidelines stipulate that hospital wastewater must be treated before discharge[28]. This can be done either in specifically designed treatment systems or in general wastewater plants after the wastewater has been disinfected and mixed with domestic sewage. Despite the guidelines, levels of implementation vary among countries[27,29,30]. In contrast to well-established systems in high-income countries, LMICs' capacity for implementation is often not ready. Previous comparative studies have revealed an obvious increase in antibiotics, antibiotic-resistant bacteria, and ARGs downstream of hospitals in LMICs like China[31,32], India[33], and Mali[34]. However, unlike other LMICs, both legislation and infrastructure in China have evolved more rapidly in recent years, including the management system for hospital wastewater. This has been further strengthened by relevant measures enforced during the COVID-19 pandemic[35]. The Chinese government has mandated that local authorities build and upgrade dedicated wastewater treatment facilities for hospitals[36]. This policy requires the separate collection and treatment of medical wastewater to ensure it meets standards before discharge, strictly prohibiting the release of untreated effluent. This raises two key questions: Is this ban being uniformly implemented across China? What is the impact of hospitals on resistome profiles in the downstream water environment in China under current wastewater management policies?
To understand the current status of the resistome in surface water within the proximity of hospitals in China, we collected samples from surface water near hospitals in 11 cities. Using metagenomic approaches, we analyzed the ARG profiles across geographical regions and hospital types. Resistome profiles in hospital-adjacent water bodies tend to be shaped more by geographical location than by proximity to the hospital. Despite high-risk ARGs, host pathogenicity, and mobility potential identified in the studied hospital wastewater, the overall ARG abundance (0.47 copies per cell) and diversity levels (389 ARG subtypes) in the adjacent water environment were similar to those of typical rivers in China and other regions. However, significantly higher ARG abundance (2.60 copies per cell) was observed in the hospital-adjacent river of Nepal, which is likely due to the poor management of hospital wastewater in the region. This study uses China as a representative case of an evolving wastewater management framework to assess the current status of water pollution from hospitals across regions. Our findings suggest that the effective implementation of WHO guidelines for managing hospital wastewater could substantially mitigate its associated environmental impact, extending from a local to a global scale.
-
Surface water samples were obtained from 11 cities in China and one city in Nepal. Sampling locations were strategically placed near hospitals, specifically at sites ≤ 1,000 m downstream from the hospital areas (Fig. 1a). These sites were grouped into two categories based on downstream distance from the hospitals: ≤ 500 m and 500–1,000 m. Sampling was conducted in urban areas of major cities in China, including Beijing, Shanghai, Shenzhen, Hangzhou, Tianjin, Wuhan, Hefei, Suzhou, Nanning, Kunming, and Baoding (Fig. 1b). A total of 100 surface water samples were collected from 11 cities in China. Among them, 11 samples were collected from lakes and 89 samples from rivers (Supplementary Table S1). Additionally, five upstream samples corresponding to the five downstream samples were collected from Suzhou (logistical constraints prevented upstream sampling in other cities). We also collected 24 non-hospital-adjacent surface water samples, comprising 13 from rivers and 11 from lakes. A site was defined as non-hospital-adjacent if no hospital was present within a 2,000 m radius. Two untreated hospital wastewater samples were also collected in Hefei (Supplementary Table S2). In Nepal, a total of 21 samples were collected in Kathmandu, including 17 hospital-adjacent samples (upstream and downstream) and four untreated hospital wastewater samples (Supplementary Table S3).
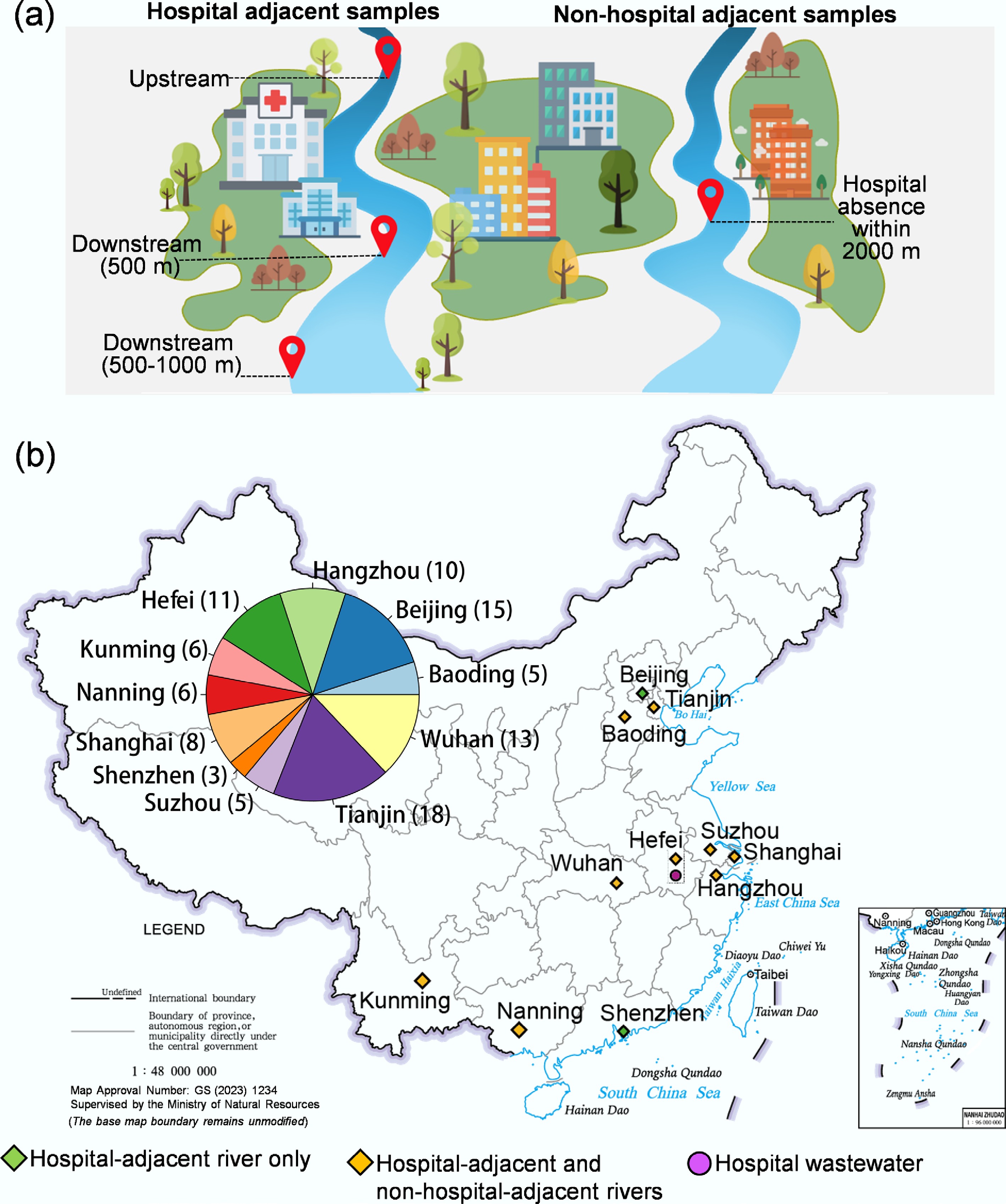
Figure 1.
Sampling design and geographical distribution of sampling sites. (a) Conceptual diagram illustrating the sampling strategy. (b) Geographical distribution of sampling sites across provinces and cities (blue shaded area). Symbol shapes and colors denote the sample types collected from the cities.
DNA extraction and high-throughput sequencing
-
Water samples with a volume of 500 mL were collected, filtered through 0.45 μm cellulose nitrate membranes, and stored at −80 °C prior to DNA extraction. Total DNA was extracted using the DNeasy PowerSoil Pro Kit (Qiagen, Hilden, Germany). After purification, DNA concentrations and quality were determined using micro-spectrophotometry (NanoDrop ND1000; Wilmington, DE). The construction and sequencing of shotgun libraries were conducted on samples with sufficient DNA amount using the Illumina NovaSeq6000 platform and the PE150 strategy by Shanghai Majorbio Bio-pharm Technology Co., Ltd., at an average depth of ~10 Gb.
Annotation of ARGs and mobile genetic elements (MGEs), and taxonomy at the short-read level
-
All raw short-read sequences were initially processed using SOAPnuke with parameters "-n 0.001 -l 20 -q 0.5 --adaMis 3 --minReadLen 150" to ensure high-quality data for subsequent analyses. The filtered reads were retained for downstream analysis (Supplementary Table S4). ARG abundance was quantified using ARGs_OAP (v3.2.4) with default settings, aligning the reads against the Structured Antibiotic Resistance Gene (SARG) database (v3.2.1)[37]. The ARGs' abundances were normalized to copies per cell. Similarly, MGEs abundance was assessed by aligning reads to a published MGEs database[38] using the same normalization procedure as with the SARG database, but replacing it with the MGEs database. For bacterial community composition analysis, the filtered reads were processed using Kraken2 (v2.0.8)[39] using the default database and confidence value of 0.05. The resulting taxonomic classifications were further refined and corrected using Bracken[39] to improve accuracy. Taxa with a relative abundance below 0.01% were filtered out to minimize false positives in the downstream analysis. To compare the profiles of ARGs across different regions, publicly available datasets from river samples in Mainland China (n = 114)[40], Hong Kong (n = 152)[13], South Korea (n = 45)[41], and Switzerland (n = 18)[42] were obtained and analyzed using the standardized pipeline as described above (Supplementary Table S5).
Gene annotation, genomic localization, and taxonomic classification of contigs
-
The contigs were assembled using the MEGAHIT software (v1.2.9)[43] with the 'meta' parameter. Following assembly, quality filtering was performed to retain only contigs longer than 1,000 base pairs (bp), with shorter contigs being discarded. Open reading frames (ORFs) were subsequently predicted from these filtered contigs using Prodigal software (v2.6.3)[44].
For functional annotation, all predicted ORFs were aligned against two specialized databases as above for ARGs and MGEs identification (as detailed in methods above). These alignments were performed using DIAMOND software (v2.1.11)[45] with default parameters. All alignment results were subjected to stringent filtering criteria, retaining only hits with sequence identity ≥ 80%, coverage > 80%, and selecting the best-scoring hit for each ORF. To determine the genomic localization of the contigs (plasmid or chromosomal origin), we employed the geNomad software (v1.11.0)[46]. For taxonomic classification, the contigs were analyzed using Kraken2 (v2.0.8)[39] with GTDB[47] as the reference database.
Assembly and taxonomic detection of metagenome-assembled genomes
-
Metagenome-assembled genomes (MAGs) were reconstructed using the BASALT software (v1.1.0)[48] with default parameters, incorporating all assembled contigs (including those shorter than 1,000 bp) and clean reads. The resulting MAGs were subjected to quality filtering based on completeness and contamination thresholds, retaining only those with a completeness ≥ 90% and contamination ≤ 5%. For functional annotation, ARGs and MGEs were identified in the filtered MAGs using the same pipeline as described for contigs. Taxonomic classification of the MAGs was performed using GTDB-Tk[49] with GTDB[47] as the reference database.
Statistical methods
-
Principal coordinates analysis (PCoA), based on Bray–Curtis dissimilarity, was employed to assess differences in ARG profiles across various samples. ANOSIM (Analysis of Similarities) was used to evaluate the significance of groupings based on sample metadata. A t-test was employed to assess the difference between two groups. ANOVA (Analysis of Variance) was employed to assess the difference between two or more groups. Network analyses were performed to investigate the co-occurrence patterns between ARGs and microbial taxa. Connections in the network analysis were defined as those with a significant correlation (Spearman's ρ > 0.4, p < 0.05). All statistical analyses were performed using R, with results visualized using the ggplot2 package. Network analyses were visualized in Gephi.
-
The abundance of total ARGs (copies per cell) in surface water samples collected near hospitals varied significantly across 11 cities in China. The average abundance was 0.47 copies per cell. Shenzhen showed the highest ARG levels, with an average of 1.11 copies per cell, which was nearly eight times higher than the lowest level observed in Wuhan (0.14 copies per cell) (Fig. 2a, b).
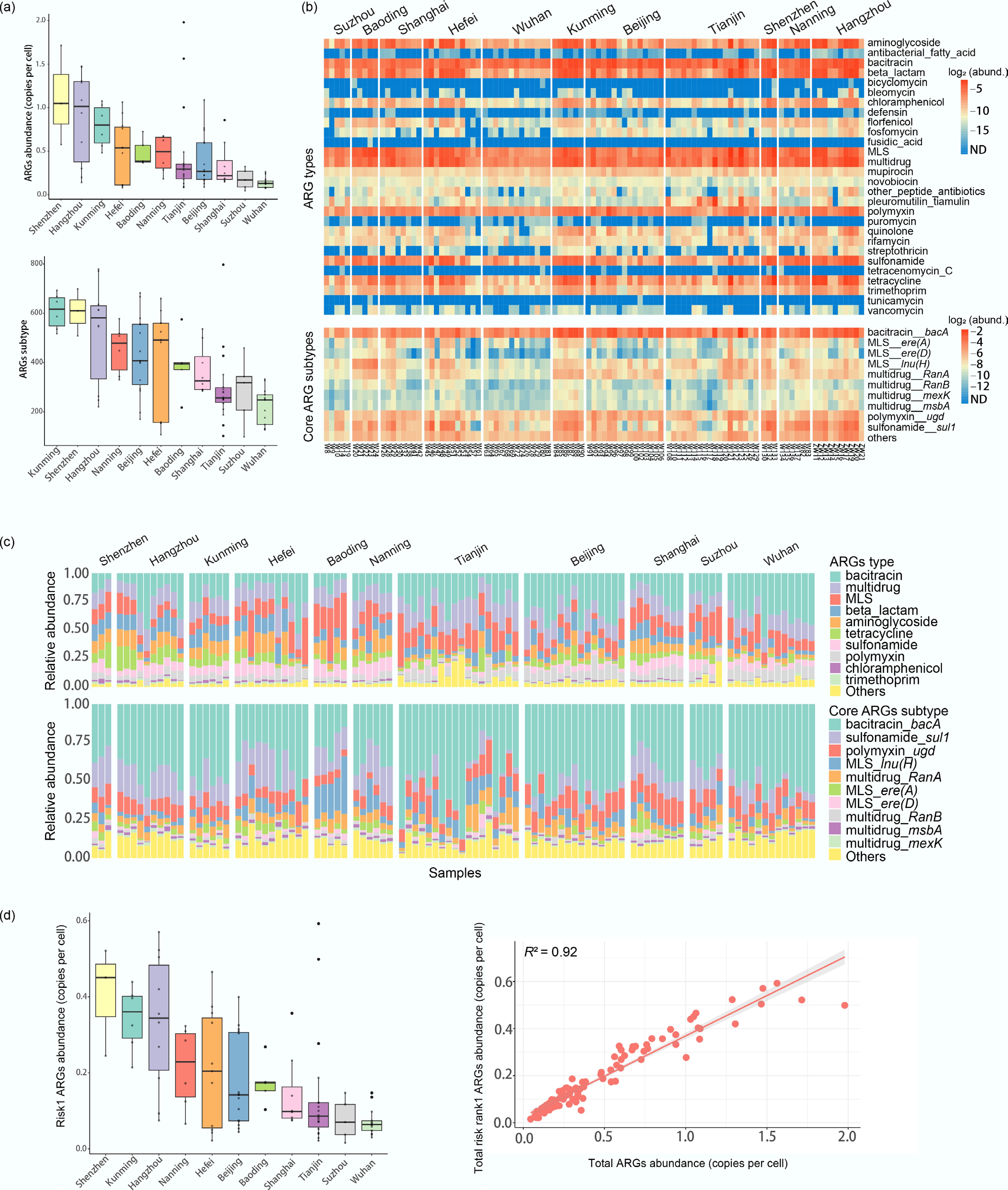
Figure 2.
Profiles of ARG abundance and diversity across samples. (a) Box plots of ARG abundance (copies per cell) and diversity (number of unique ARG subtypes) among the sampling cities (center line: median; box limits: 1st and 3rd quartiles). (b) Heatmap of the top 10 most dominant and core ARG subtypes. Abundance is displayed as log2-transformed copies per cell, with an offset of 1/10 of the minimum non-zero value added to each measurement to permit log transformation. (c) Compositional profiles of ARG types and core subtypes. The top 10 are labeled, with the remainder grouped as 'Others'. (d) Abundance of risk Rank 1 ARGs and the linear correlation between total ARGs and Risk I ARG abundances (p < 0.01).
The average ARG richness was 389 subtypes across all samples. Kunming's samples displayed the greatest average diversity of ARGs (608 subtypes), while Wuhan's samples had the lowest average diversity (222 subtypes) (Fig. 2a). Correlation analysis showed that ARG subtype diversity tended to decrease as total ARG abundance declined (r = 0.84, p < 0.01) (Supplementary Fig. S1). In total, 1,723 ARG subtypes across 28 types were identified in the hospital-adjacent water environment, out of 2,843 subtypes from 32 types listed in the reference database. The most diverse ARG types included β-lactam (976 subtypes), aminoglycoside (139 subtypes), multidrug (112 subtypes), and macrolide-lincosamide-streptogramin (MLS) (109 subtypes).
Diversity at the ARG subtype level varied across sampling sites, yet several core ARG types were consistently present, including aminoglycoside, bacitracin, β-lactam, chloramphenicol, MLS, multidrug, mupirocin, novobiocin, polymyxin, sulfonamide, tetracycline, and trimethoprim (Fig. 2b, c). These 12 core types accounted for 96.42% of the total ARG abundance at each location on average. Additionally, 29 ARG subtypes were common to all river water samples, such as resistance genes for bacitracin (bacA, bcrA), polymyxin (arnA, rosB, ugd), and sulfonamide (sul1). Among these ARG subtypes, bacA, ugd, sul1, and lnu(H) exhibited higher average abundances compared to other ARG subtypes, which were less than 0.01 copies per cell. For each sampling point, these core subtypes contributed to more than 50.08% of the total abundance on average.
The Rank I ARGs, which pose a potential threat to human health, were obtained from the study by Zhang et al.[50]. Among those 225 Rank I ARG subtypes, 113 subtypes were detected in the samples of our study. Similar to the overall resistome, the highest total abundance of Rank I ARGs was observed in Shenzhen, with an average of 0.41 copies per cell. The lowest total abundance was 0.07 copies per cell in Wuhan. The average total abundance of Rank I ARGs across all samples was 0.19 copies per cell, and the abundance profile was significantly correlated with total ARGs (Fig. 2d). Three Rank I ARGs, bacA, ere(A), and sul1, were detected in all samples. Among these, bacA—which confers resistance to bacitracin—showed the highest abundance, with an average of 0.08 copies per cell, accounting for approximately 42.11% of the total average abundance (Supplementary Fig. S2). The highest average abundance of bacA was recorded in Kunming (0.16 copies per cell), while the lowest was detected in Suzhou (0.03 copies per cell) (Supplementary Fig. S3).
The influence of hospital characteristics and proximity on ARG profiles
-
We analyzed surface waters adjacent to hospitals of varying sizes (large: > 500 beds, n = 91 samples; small-to-medium: 100–500 beds, n = 9 samples) and service types (general, n = 65 samples; specialized, n = 35 samples), as classified by the Chinese Ministry of Health guidelines. Analysis revealed that neither hospital service type nor its size significantly shaped the adjacent water resistome (Fig. 3a, b). Principal coordinates analysis showed no significant difference in resistome composition between general and specialized hospitals (ANOSIM: R = 0.01, p > 0.01). Consistently, total ARG abundance was not significantly different, although the mean was slightly higher for general hospitals (0.46 vs 0.41 copies per cell). Similarly, hospital size had no significant effect on total ARG abundance (ANOSIM: R = 0.04, p > 0.01), despite a higher mean abundance in the water environment near small-to-medium hospitals (0.58 vs 0.46 copies per cell).
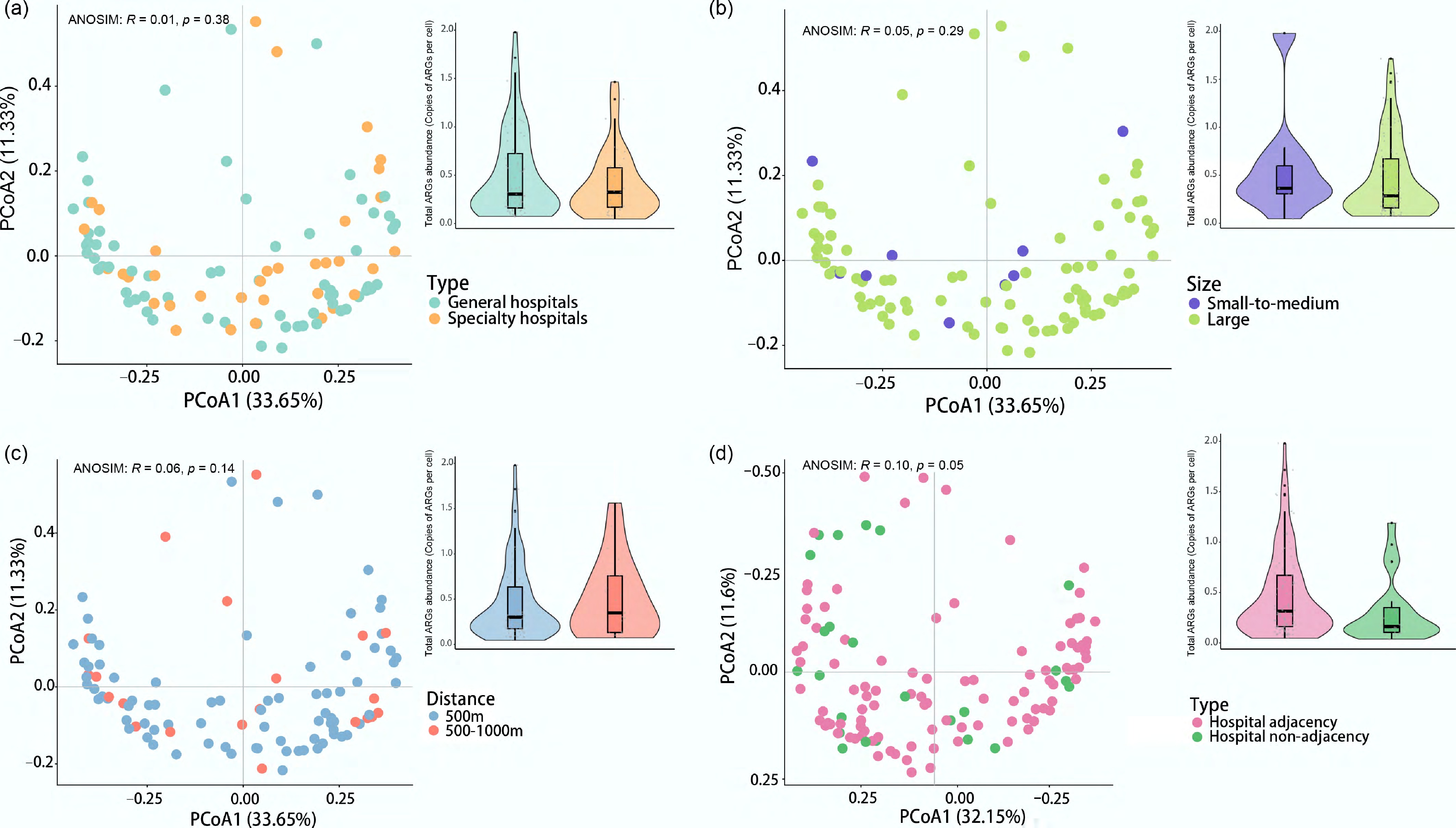
Figure 3.
Influence of hospital characteristics and proximity on the ARG profile. Panels show the effect of (a) hospital type (general vs specialty), (b) hospital size (large: > 500 beds; small-to-medium: 100–500 beds), (c) sampling distance downstream (near: ≤ 500 m; far: 500–1,000 m), and (d) hospital adjacency (hospital present within 2,000 m; hospital absent). In each panel, the right plot is a PCoA showing ARG profile clustering, and the left plot illustrates ARG abundance (copies per cell) as a boxplot (center line: median; box limits: 1st and 3rd quartiles).
The proximity of sampling sites to the upstream hospital also showed no significant effect. While samples were categorized as near (≤ 500 m, n = 79 samples) or far (> 500 m, n = 21 samples) (Fig. 3c), PCoA indicated that ARG patterns were not structured by the sampling distance from hospitals (ANOSIM: R = 0.06, p > 0.01). In contrast, geographical location (sampling city) was a significant factor shaping the ARG profiles (ANOSIM: R = 0.24, p < 0.01).
This limited influence of hospital presence was further supported by comparisons with non-hospital-adjacent surface waters (Fig. 3d). We collected 24 control-like non-hospital-adjacent samples from nine Chinese cities, with sampling sites defined as having no hospital within a 2,000 m radius. The cities included were Baoding, Hangzhou, Hefei, Kunming, Nanning, Shanghai, Suzhou, Tianjin, and Wuhan. Although these control sites were not fully isolated from the influence of distant hospitals and lacked pairwise matching with hospital-adjacent samples, they still provide a relevant baseline for assessing the impact of hospital presence. In these control samples, the average total ARG abundance was 0.31 copies per cell, ranging from 0.04 (Baoding) to 1.19 (Tianjin). No significant difference in total ARG abundance was found between the hospital-adjacent and non-hospital-adjacent groups (ANOSIM: R = 0.10, p > 0.01). Despite the limited sample size, the comparable ARG abundance between paired upstream and downstream samples, further suggests minimal impact from hospitals (Supplementary Fig. S4).
We further investigated the role of ecosystem type by comparing ARG profiles between river (n = 89) and lake (n = 11) samples. A significantly lower average ARG abundance and diversity were found in lake samples compared to river samples (0.13 vs 0.51 copies per cell and 209 vs 411 subtypes) (ANOSIM: R = 0.15, p = 0.05; Supplementary Fig. S5). Rivers, being more dynamic and flowing water systems, are generally more susceptible to pollution from urban residential areas. However, the results may be biased by the sampling design of this study, as there were more river samples than lake samples.
In contrast to surface water, hospital wastewater is expected to contain far higher loads of ARGs. This was confirmed by samples from an infectious disease hospital in Hefei, where both treated and untreated effluent exhibited significantly higher total ARG abundance (6.60 and 9.77 copies per cell, respectively) and diversity (1,010 and 1,162 subtypes, respectively) compared to any hospital-adjacent surface water from the same city (0.08–1.06 copies per cell and 107–406 subtypes) (Supplementary Fig. S6).
Regional comparison of ARG profiles in urban surface waters
-
To contextualize our findings, we performed a comparative analysis of river resistomes from other regions, either with or without a hospital within proximity. This included hospital-adjacent sample datasets from Nepal (collected and sequenced using our protocol, as detailed in methods), as well as available datasets from typical rivers in Mainland China, Hong Kong, South Korea, and Switzerland. For this analysis, 'typical rivers' were defined as those where sampling was not designed based on hospital proximity and where source information did not explicitly consider the hospital factor; the non-hospital-adjacent river resistomes in this study were included in this part as 'typical rivers'.
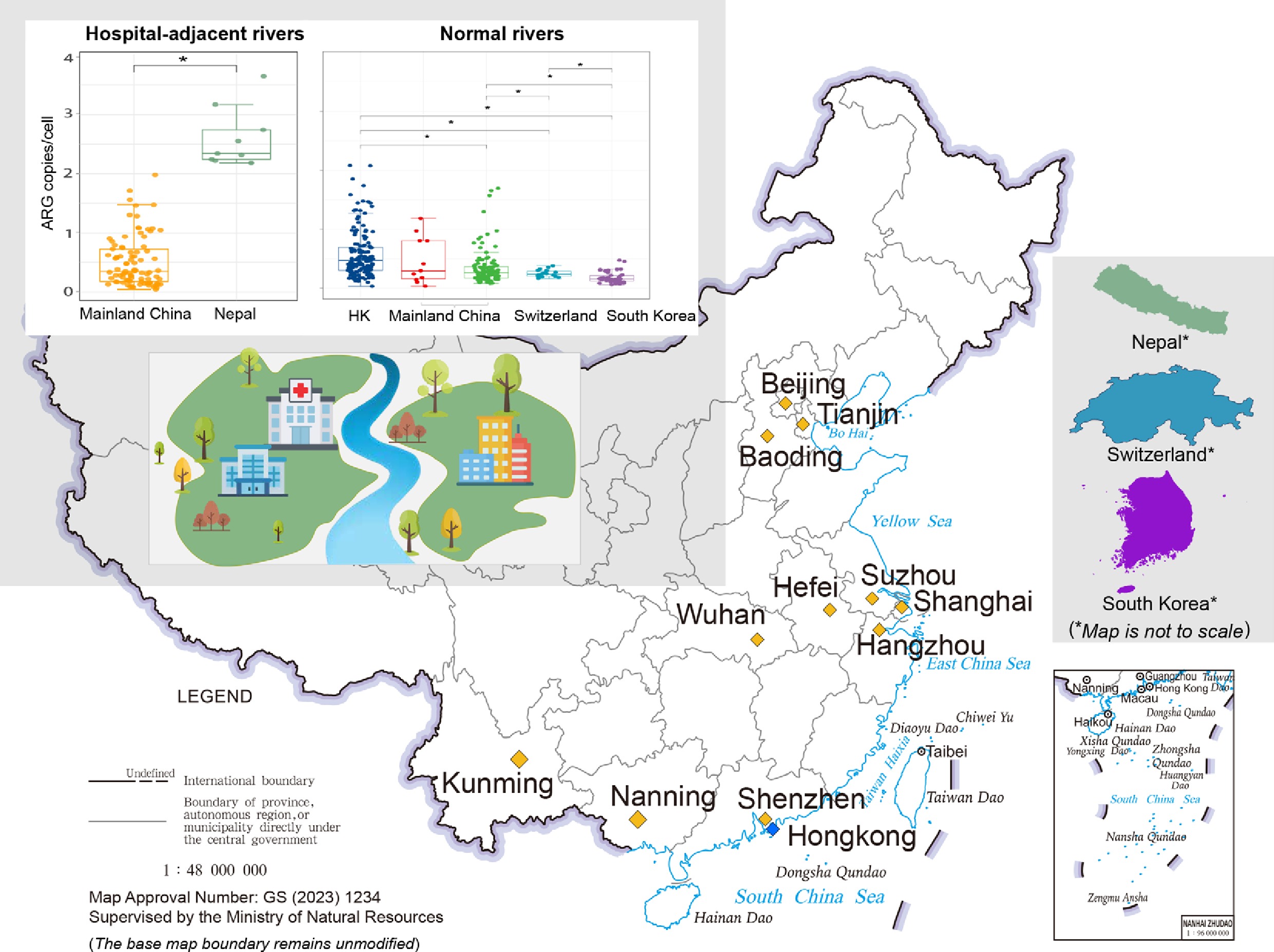
We first compared the influence of hospital proximity on river resistomes between Nepal (n = 9) and China (n = 89) (Fig. 4a). We acknowledge that this two-region comparison is limited; however, to the best of our knowledge, no other regional-scale studies on hospital-adjacent river resistomes are available, despite our efforts to incorporate a broader geographical scope. The resistome profile of samples in Nepal exhibited significantly higher average ARG abundance (2.60 copies per cell) compared to the hospital-adjacent rivers in China (0.51 copies per cell), as well as significantly higher average ARG diversity (727 subtypes vs 411 subtypes) (p < 0.01). Furthermore, the lack of upstream-downstream differences in Nepal (Supplementary Fig. S7) suggests a background of pervasive urban pollution. Based on these findings, we hypothesize that (1) the relatively low ARG abundance in China's hospital-adjacent rivers, compared to the high levels found in Nepal, is likely due to the more effective wastewater management practices implemented at the sampled Chinese hospitals than in Nepal; (2) widespread environmental pollution in Nepal may contribute to a high background level of ARGs. Additional survey data on pollution sources in close proximity would further support these hypotheses; however, such data are currently lacking.
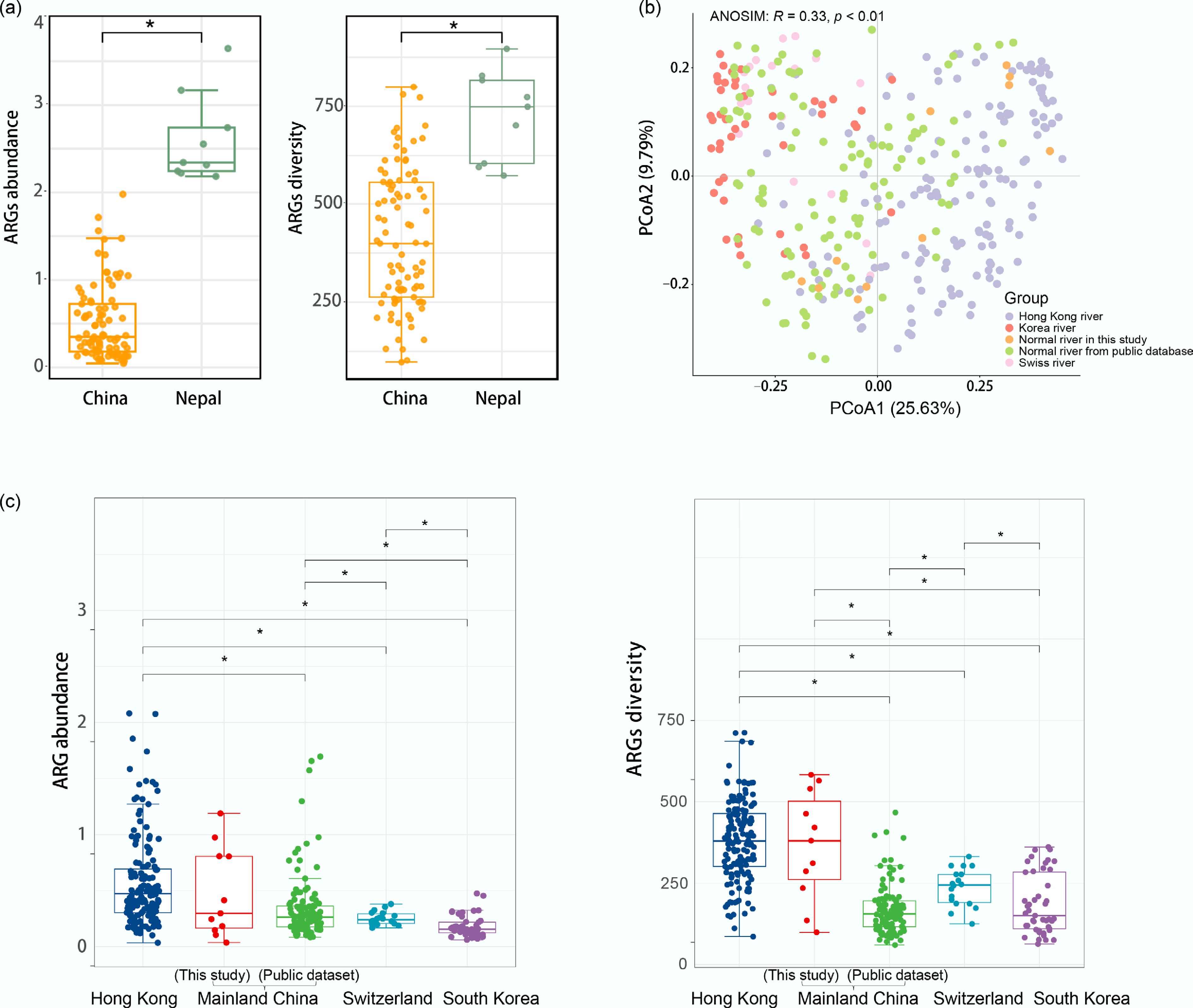
Figure 4.
Comparative analysis of ARG profiles across different geographical regions. (a) ARG abundance and diversity in hospital-adjacent rivers from China and Nepal. (b) PCoA plot based on Bray–Curtis dissimilarity, showing the overall similarity of ARG profiles in typical rivers among the different regional groups, including Mainland China (this study and public dataset[40]), Hong Kong[13], South Korea[41], and Switzerland[42]. (c) ARG abundance and diversity in typical rivers from different regional groups. In both panels, boxplots display the median (center line) and interquartile range (box). Significant differences between groups, determined by t-test, are indicated by asterisks (*).
We then profiled the resistomes of typical rivers across multiple regions, including Mainland China (incorporating both data from this study [non-hospital-adjacent river resistome] and online public sources), Hong Kong, South Korea, and Switzerland. A significant difference was observed among these regional river resistomes (ANOSIM, R = 0.33, p < 0.01) (Fig. 4b, c). The average ARG abundance followed a hierarchical trend (with mean abundance [copies per cell] and diversity [number of unique ARG subtypes] in brackets): Hong Kong (0.58 and 380) > Mainland China—this study (0.47 and 365) > Mainland China—public datasets (0.34 and 167) > Switzerland (0.25 and 237) > South Korea (0.18 and 184).
Mobile genetic elements and the mobilization potential of ARGs
-
The overall average abundance of MGEs varied from 0.31 copies per cell in Wuhan to 3.99 copies per cell in Hangzhou (Fig. 5a). The composition of MGE types was similar across different cities. Transposase and IS91 were the two most prevalent MGEs in all samples, accounting on average for 87.21% of MGE abundance per sample. A significant positive correlation was observed between the total relative abundance of ARGs and MGEs in all surface water samples in this study (R2 = 0.69, p < 0.01), indicating that MGEs are a key driver of ARG transmission (Fig. 5b). Among all 33,076,795 assembled contigs, a total of 4,175 contigs carrying 20 different ARG types were obtained (Fig. 5c). Among these contigs carrying ARGs, 77.65% originated from chromosomes, whereas 22.28% originated from plasmids (Fig. 5d). Co-occurrence of ARGs and MGEs was found on 471 contigs. Regarding the ARG-carrying plasmids, 5.06% were identified as mobilizable, while 94.94% were classified as non-mobilizable. The ARGs located on plasmid contigs were predominantly associated with resistance to MLS (20.38%), tetracycline (18.89%), β-lactam (17.80%), aminoglycoside (16.39%), and sulfonamide (7.99%).
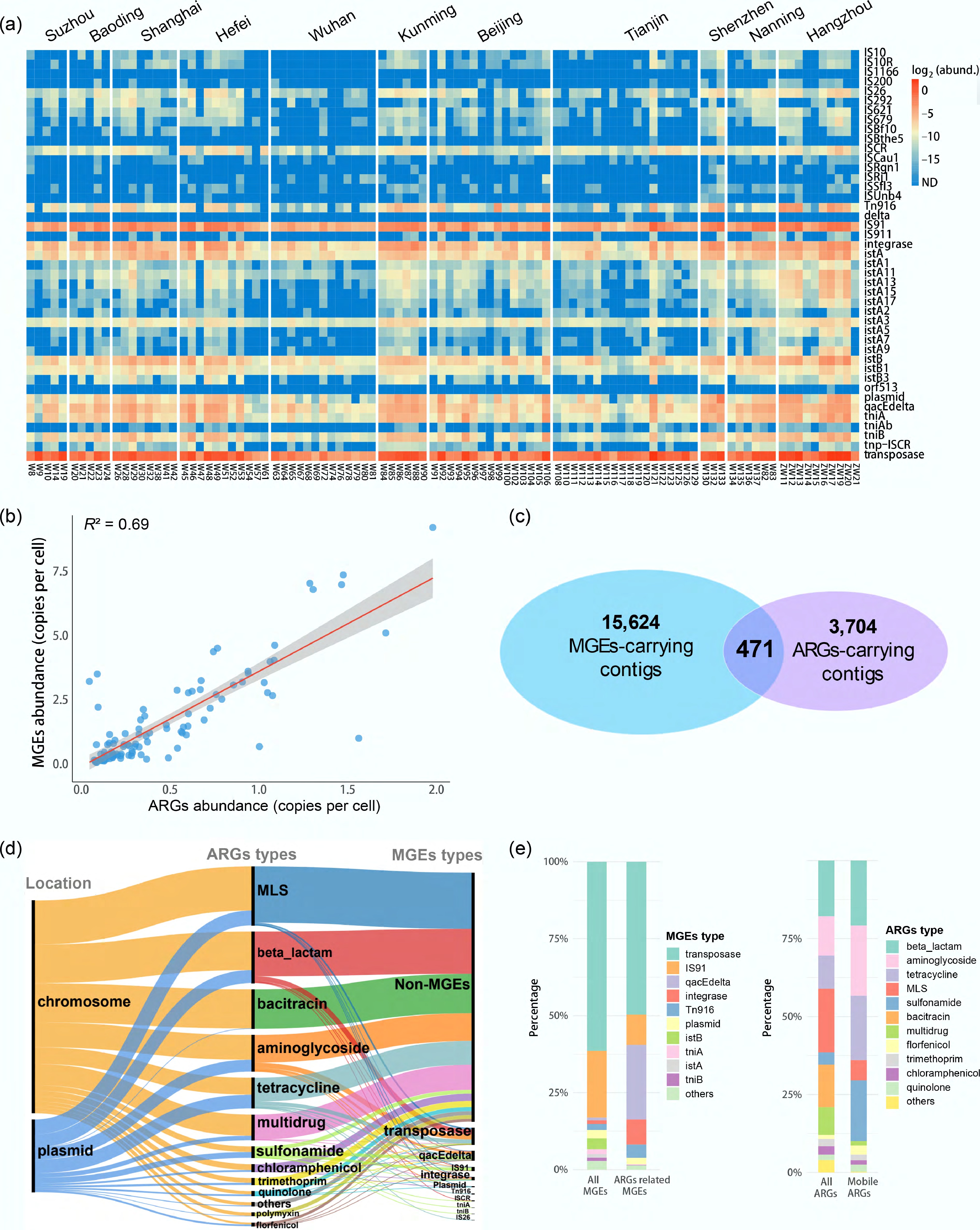
Figure 5.
MGEs profiles and their co-occurrence with ARGs. (a) MGEs' abundance across all samples. Abundance is displayed as log2-transformed copies per cell, with an offset of 1/10 of the minimum non-zero value added to each measurement to permit log transformation. (b) Linear correlation between the abundance (copies per cell) of ARGs and MGEs. (c) Venn diagram showing the number of contigs carrying MGEs, ARGs, or both. (d) Sankey diagram showing the distribution of contigs by their origin (chromosomes and plasmids), carried ARG types, and carried MGE types. (e) Composition of MGEs on all contigs vs on ARG-carrying contigs (left), and composition of ARGs on all contigs vs on MGEs-carrying contigs (right). The top 10 types are labeled; the remainder are grouped as 'others'.
The MGEs' abundance and diversity profile across all assembled contigs closely resembled that from unassembled short reads, with transposase and IS91 accounting for 61.23% and 21.74%, respectively. However, based on co-occurrence analysis of contigs, the ARG-associated MGEs profile differed notably from that of total MGEs on contigs, showing increases in qacEdelta, integrase, and Tn916, and a reduction in transposase abundance. Similarly, the distribution of MGE-associated ARGs differed from that of total ARGs on the contigs (Fig. 5e). For example, the abundance of aminoglycoside resistance reached 22.55% of the MGE-associated ARGs, which, however, was only 12.82% in total ARGs. On the contrary, MLS resistance, which was dominant among all ARGs on contigs (20.69%), was less common among MGE-associated ARGs on contigs (6.50%). Additionally, 10 ARG types were in physical proximity (≤ 5 kbp distance) to qacEdelta genes, and 11 ARG types were linked to transposase genes. Among the ARGs associated with transposase genes, the majority were genes conferring resistance to β-lactam (33.85%), aminoglycoside (26.15%), and tetracycline (20.51%), indicating a high potential for mobility driven by transposases for these genes. Notably, Tn916 was specifically enriched in contigs carrying tetracycline resistance genes.
Microbial community composition and identification of potential ARG hosts
-
Short reads were used to examine overall community composition across all samples. At the class level, more than 70% of the reads were assigned to Betaproteobacteria (27.85%), followed by Gammaproteobacteria (15.12%), Cyanophyceae (14.10%), Alphaproteobacteria (10.41%), and Actinomycetes (7.71%). The taxonomic composition at the order level indicated that Chroococcales had a higher relative abundance on average in samples from Suzhou (37.94%), Shanghai (19.13%), Tianjin (15.78%), Hefei (7.75%), Wuhan (4.94%), compared to other samples where its abundance was lower (0.05%–0.95%) (Fig. 6a). Since Chroococcales are an order of cyanobacteria, their relatively high abundance is likely due to elevated nutrient levels and the potential for algal blooms induced by nearby human activities. Across all samples, significant differences were observed in community composition (ANOSIM: R = 0.20, p < 0.01) (Fig. 6b).
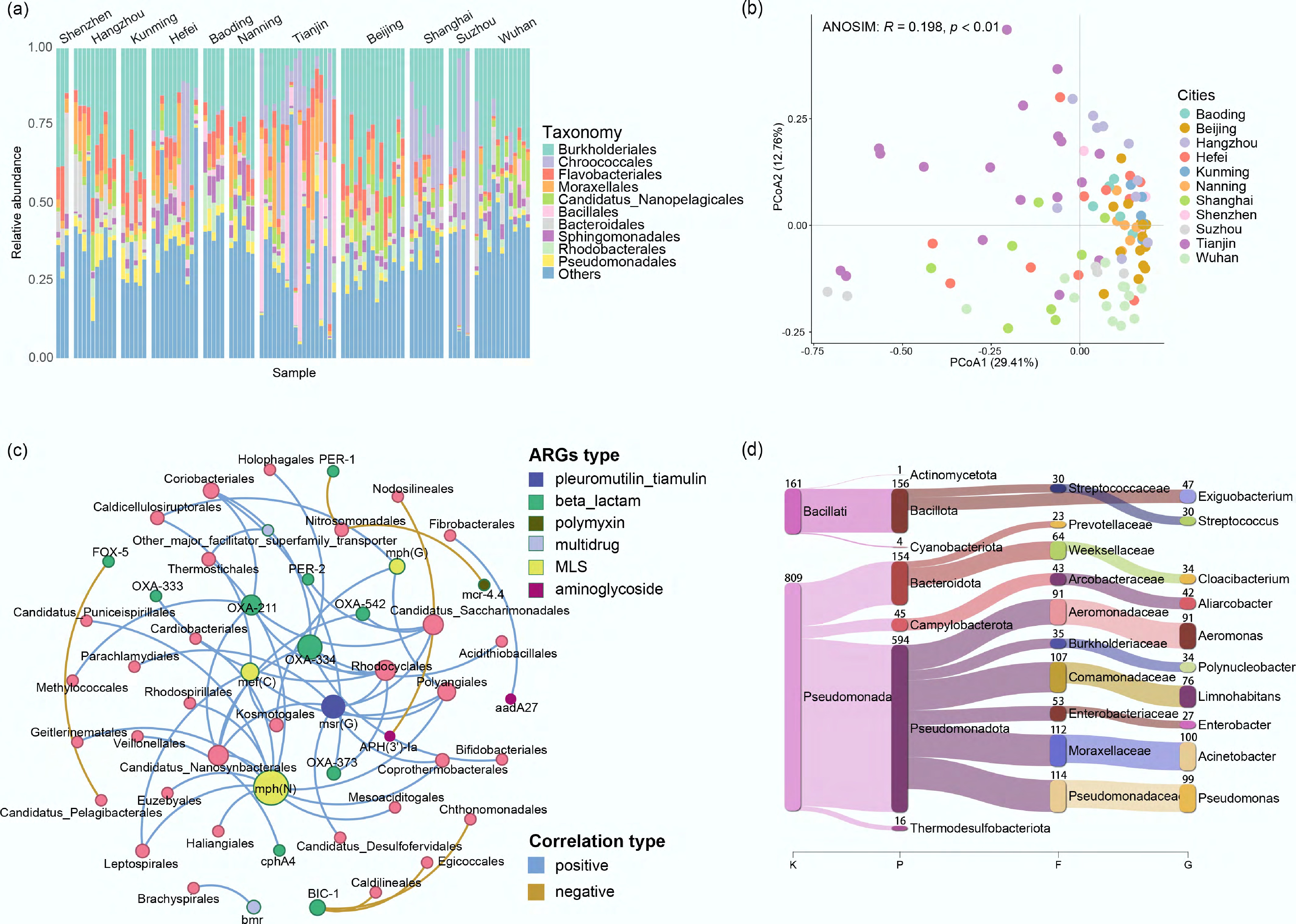
Figure 6.
Microbial community profiles and potential ARG hosts across sampling sites. (a) Relative abundance of the microbial community at the order level. The top 10 groups are labeled; the rest are grouped as 'Others'. (b) PCoA of the microbial community profile based on Bray–Curtis dissimilarity. (c) Co-occurrence network of ARGs and their potential hosts at the order level (Spearman's ρ > 0.4, p < 0.05). (d) Sankey diagram showing the host taxonomy of ARG-carrying contigs, with taxonomy names and contig numbers labeled on the bars.
Hosts of ARGs were retrieved using two complementary approaches: (1) a correlation network analysis linking the ARG profile to the microbial community at the order level, and (2) the direct taxonomic annotation of contigs that carried ARGs. Network analysis showed that Coriobacteriales, Rhodocyclales, and Polyangiales were the potential ARG hosts (Fig. 6c). Compared to read-level analysis, long-fragment (contig) based analysis provided a higher resolution of genomic context. This method identified 193 host species, predominantly Pseudomonas aeruginosa (7.53%), Limnohabitans sp. 103DPR2 (4.33%), Aeromonas media (3.91%), Acinetobacter johnsonii (3.81%), and Aliarcobacter cryaerophilus (3.51%) (Fig. 6d). Notably, 57 of the identified host species were pathogens, collectively carrying 13 different types of ARGs. Among these, several species harbored a particularly high number of ARGs (more than 18 per species), including Escherichia coli, A. media, A. caviae, Acinetobacter johnsonii, Alcaligenes faecalis, A. veronii, and Enterobacter huaxiensis.
Furthermore, contigs carrying Rank I ARGs were frequently identified and were found to originate from both chromosomes and plasmids. Of these, 534 Rank I ARGs were located on chromosomal contigs (64.49% of the total), while 292 were located on plasmid contigs (35.27%). Taxonomic annotation at the family level revealed that the hosts of these contigs were primarily Aeromonadaceae (42 contigs), Enterobacteriaceae (29), and Geobacteraceae (13) (Supplementary Fig. S8).
Using hybrid assembly and binning, we reconstructed 80 high-quality MAGs with < 5% contamination and > 90% completeness. Within these MAGs, we identified 134 ARG subtypes spanning 14 different ARG types (Supplementary Fig. S9). Fifteen MAGs were identified as potential pathogenic hosts carrying ARGs, geographically distributed across Tianjin (5 MAGs), Kunming (3), Shenzhen (2), Hangzhou (2), Beijing (1), Hefei (1), and Wuhan (1) (Supplementary Fig. S10). Notably, we identified a multi-resistant E. coli MAG in Tianjin that carried six types of ARGs (excluding multidrug resistance genes): bacitracin (bacA), quinolone (QnrS9), microcin (yojI), β-lactam (FONA-6, LAP-2, ampC), MLS (macA, macB, mph[B]), and polymyxin (arnA, eptA, pmrF, rosB). From the same city, an Enterobacter huaxiensis MAG was found to carry six ARG types: bacitracin (bacA), quinolone (QnrE2), microcin (yojI), polymyxin (pmrF), MLS (macA), and β-lactam (ACT-29, OmpK37).
-
Through a specifically designed sampling campaign, for the first time, this study revealed the variation in ARG abundance and diversity patterns in the surface water environment adjacent to hospitals across different regions of China. Based on previous sporadic relevant observations[18,31,32,34], a relatively higher ARG load was expected in the surrounding rivers of hospitals than in those without hospitals within proximity; however, the results of our study revealed that ARG patterns in hospital-adjacent rivers were more likely impacted by non-hospital factors than by hospitals located upstream. By extensive comparative analysis between different sample groups collected in this study and from previous studies, we present three key lines of evidence indicating a minimal impact of hospitals on the resistome of the studied Chinese rivers: first, no significant difference was observed in ARG levels between the studied hospital-adjacent and non-hospital-adjacent rivers in China (0.47 vs 0.31 copies per cell); second, neither proximity to hospitals (i.e., at varying distances) nor hospital characteristics (i.e., sizes and types) demonstrated a significant effect on resistome profiles of adjacent rivers; third, cross-regional comparisons further contextualized these findings: ARG levels in Chinese hospital-adjacent rivers were significantly lower than in Nepalese samples (which are likely impacted by untreated hospital wastewater) (0.47 vs 2.6 copies per cell), yet were comparable to those in other Chinese rivers. Furthermore, the ARG levels in typical Chinese rivers remained significantly higher than those in high-income countries like Switzerland and South Korea (0.47 vs 0.25 and 0.18 copies per cell, respectively), which likely benefit from advanced infrastructure that substantially mitigates wastewater pollution.
We demonstrated that under China's current wastewater management policies, hospitals may have a limited impact on downstream river resistomes. Instead, the driving factors behind these observed river resistomes are more likely related to geographical factors. For example, a wide range in abundance (1.11 to 0.14 copies per cell) and diversity (222 to 608 ARG subtypes) was observed across sampling cities, and their clustering pattern was significantly shaped by geographical location. Notably, significant variation was observed even at the city scale, indicating that water pollution is influenced by both local environmental factors and regional factors. For instance, within-city differences in ARG abundance in cities like Tianjin, Beijing, and Hangzhou reached 5 to 10-fold. Indeed, this pattern of dynamic and variable river resistomes is consistent with previous studies[13,14,41,51]. Research on the Pearl River documented similar fluctuations, with more pronounced variation during dry seasons[14], while a three-year seasonal profiling in Hong Kong revealed that high spatial variation—with 10-fold differences in abundance and diversity between polluted and unpolluted sites—is common[13]. This high variation is primarily due to a multitude of natural and anthropogenic factors that affect rivers. Both explicit and implicit human activities, including diverse agricultural and industrial practices along rivers, contribute substantially through point and non-point source discharges. These combined inputs drive resistome dynamics and complicate source tracking[52].
Despite the minimal direct impact from upstream hospitals observed in this study, a concerning level of environmental deterioration—marked by the frequent occurrence of ARGs in rivers—was evident across the studied Chinese cities, consistent with previous research[13,14,41,53]. This includes high-risk (Risk Rank I) ARGs such as bacA, ere(A), and sul1, which were frequently identified alongside their potential pathogenic hosts from families like Aeromonadaceae and Enterobacteriaceae. This pattern highlights the extensive dissemination of ARGs and their associated pathogenic hosts, demonstrating their ability to transmit across geographic, ecological, and phylogenetic boundaries. Worse yet, MAGs of multi-resistant Escherichia coli and Enterobacter huaxiensis were recovered from Tianjin rivers, carrying six distinct types of ARGs (e.g., bacitracin, β-lactam, and polymyxin). The successful assembly of these pathogens from metagenomic data indicates their significant prevalence in the river environment. Indeed, the detection of such high-risk ARGs is a common finding in previous resistome surveys of the aquatic environment, spanning regions of all income levels. For instance, mobilizable high-risk ARGs like mcr and tet(X) have been reported in the Pearl River delta[14], and diverse ARGs and host pathogens in surface water from Hong Kong[13], South Korea[41], and Switzerland[53]. These findings collectively demonstrate that antibiotic-resistant bacteria and their genes have become pervasive, making public exposure through the environment inevitable. This urgent situation necessitates the implementation of standardized, spatiotemporal surveillance to track dynamic resistome profiles, generate comparable data, and inform effective mitigation strategies—a critical need that the scientific community has long advocated.
The high load of clinically relevant ARGs and pathogens in hospital wastewater poses a significant threat to public and environmental health. This risk is exacerbated in regions with poor wastewater management, where the dissemination of antibiotic resistance is amplified[7]. The elevated ARG abundance and diversity in Nepal's hospital-adjacent rivers, for instance, are likely a consequence of inadequate wastewater treatment infrastructure. This cross-regional comparison highlights the efficacy of China's stringent policies, which mandate the pre-discharge treatment of hospital effluent and have successfully mitigated the environmental release of ARGs. The situation of insufficient infrastructure is critical in many LMICs, where the burden of antibiotic resistance is most severe. In these regions, the lack of basic sanitation exacerbates environmental degradation. A strong scientific consensus affirms that establishing basic hygiene and sanitation systems is one of the most cost-effective measures to combat the environmental spread of antibiotic resistance[54]. When coupled with routine environmental surveillance, such infrastructure investments create a framework for evaluating the effectiveness of control strategies. This data-driven approach is essential for informing policy; for example, it enables the assessment of various wastewater treatment technologies and management policies—from hospitals and other sources—in reducing the spread of antibiotic resistance.
This study offers valuable insights into the resistome profiles of hospital-adjacent surface waters in China, yet a few limitations must be acknowledged. First, its cross-sectional design and single-time-point sampling may not fully capture the temporal dynamics of ARGs dissemination, particularly given the known seasonal fluctuations in river resistomes demonstrated by longitudinal metagenomic studies of major Chinese rivers[14,16,55]. Future studies incorporating seasonal sampling would further elucidate temporal patterns. Second, while the findings suggest a limited hospital influence—potentially reflecting China's recent stringent wastewater policies—this conclusion may lack robustness. The study's scope was constrained by the absence of pollution source surveillance within the sampling radius, the limited number of cities sampled, an unbalanced regional distribution, and insufficient representation of underdeveloped areas. Specifically, only two untreated hospital wastewater samples from Hefei and five upstream samples from Suzhou were included, introducing uncertainties regarding the contributions of upstream sources and hospital discharges to the downstream water body. Furthermore, comparative analysis may be biased by the use of public data derived from different methodologies. Future research should employ longitudinal sampling to track temporal dynamics, expand geographical comparisons to include diverse settings (e.g., developed vs underdeveloped regions), and integrate geographical data with microbial ecology to better elucidate the links between environmental factors, policy, and the dissemination of antibiotic resistance.
While this study focuses on hospital-derived resistance, broader regional surveillance is crucial for identifying all potential sources of antibiotic resistance. Under current intensive anthropogenic pressure, the inherent connectivity of the One Health triad—humans, animals, and the environment—readily drives the spread of resistance far beyond any single sector[56]. Therefore, routine surveillance should be implemented within each sector and integrated across them to systematically address the environmental dimension of antibiotic resistance[57]. In the long term, comprehensive and sustained surveillance will provide more robust evidence to link environmental data with clinical outcomes, thereby informing public health strategies.
-
This study presents a systematic evaluation of ARG dissemination in hospital-adjacent surface waters across China. Our findings provide crucial insights into the roles of geographical variation and wastewater management policies on the resistome in urban water environments. The data demonstrate that under China's current stringent regulations, resistome profiles near hospitals are comparable to those of typical urban surface waters, indicating that regional environmental factors, rather than proximity to hospitals, are the primary drivers of ARG distribution. However, the detection of clinically significant ARGs, often linked to MGEs, indicates the persistent risk and need for long-term monitoring. These results partially validate China's policy-driven approach in curbing the spread of antibiotic resistance via wastewater, establishing a critical benchmark for future surveillance of hospital-adjacent water environments in other regions, particularly in LMICs.
We thank all the individuals who assisted with field sampling, sample preparation, and DNA extraction.
-
It accompanies this paper at: https://doi.org/10.48130/biocontam-0026-0008.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: Rong Zhang, Liguan Li: conceiving and initiating the study, providing overall supervision; Wudi Gai, Xiaoyang Ju: writing the main manuscript text and preparing all figures; Xuemei Mao: providing metagenomic data from Hong Kong, South Korea, and Switzerland; Zelin Yan, Yuchen Wu, Yi Sun, Hanyu Wang: sample collection, DNA extraction, sequencing coordination; Haicai Cheng, Yuxin Tao, Guhang Shi: data analysis and visualization; Xuemei Mao, Yanping Mao, Ke Yu, Tong Zhang: providing critical feedback on the manuscript. All authors reviewed and approved the final version of the manuscript.
-
Raw sequencing data for the metagenomes have been deposited in the China National Center for Bioinformation under BioProject number PRJCA051371. The accession numbers for the public metagenomic datasets included in this manuscript are listed in the supplementary tables.
-
This work is supported by the General Research Fund (Grant Nos 17202522 and 28200525), the National Natural Science Foundation of China (Grant No. 22193062), and the Dean's Research Fund of the Education University of Hong Kong (0405M).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xiaoyang Ju, Wudi Gai
Full list of author information is available at the end of the article. - The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ju X, Gai W, Yan Z, Wu Y, Sun Y, et al. 2026. What leaves the hospitals can be contained: a China-wide resistome study reveals limited impact of hospitals on adjacent surface waters. Biocontaminant 2: e011 doi: 10.48130/biocontam-0026-0008 

What leaves the hospitals can be contained: a China-wide resistome study reveals limited impact of hospitals on adjacent surface waters
- Received: 30 March 2026
- Revised: 22 April 2026
- Accepted: 11 June 2026
- Published online: 26 June 2026
Abstract: The frequent occurrence of bacteria, particularly human pathogens, carrying antibiotic resistance genes (ARGs) has become the norm rather than the exception. This is primarily due to decades of selective pressure, which has driven the widespread dissemination of ARGs across humans, animals, and the environment. Among transmission pathways, wastewater-polluted surface waters are notorious for spreading ARGs. Especially, water bodies near hospitals are more likely to receive hospital-derived ARGs in the absence of proper wastewater management. Resistome surveillance of hospital-adjacent surface waters can provide a basis for understanding and managing the hospital environment's ARG transmission chain. In this study, water samples were collected from hospital-adjacent surface waters in 11 cities across China. Results were compared with metagenomic data from rivers in other regions, including Nepal, Hong Kong, South Korea, and Switzerland. Resistomes of hospital-adjacent surface waters varied significantly across the studied Chinese cities; Shenzhen exhibited the highest ARG abundance. While ARG diversity differed across sites, similar core ARG types accounted for > 96% of total ARG abundance in each sample. Through extensive comparisons of regions, sampling locations, and hospital characteristics, we found that proximity to hospitals did not significantly influence resistome profiles in Chinese surface waters, where geographical factors were the primary drivers of ARG distribution. In contrast, rivers near hospitals in Nepal—a country with likely inadequate wastewater management—showed substantially elevated ARG levels. These findings demonstrate that, in the studied Chinese cities, with adequate infrastructure and policy-driven management, the environmental impact of hospital wastewater has been substantially mitigated. These results provide a critical framework for guiding the development of infrastructure and subsequent policies for managing hospital wastewater in low- and middle-income countries.
-
Key words:
- Hospital-adjacent surface water /
- Resistome /
- Wastewater /
- Management