









-
Inefficient nitrogen utilization in ruminants is attributed to the excretion of 75% of nitrogen as ammonia and urea[1]. This leads to environmental pollution through nitrate leaching, NH3 volatilization, nitrification, or denitrification to N2O gas, which is 298 times more potent than CO2[2]. Enteric methane (CH4) is another rumen fermentation byproduct with a global warming potential 27 times greater than that of CO2[2], which accounts for approximately 39% of emissions from the livestock sector[3], and 2%−12% dietary energy loss[4].
The rumen of ruminants hosts various microbes, including bacteria, fungi, protozoa, and viruses that convert ingested feed into energy and carbon sources through fiber digestion. However, rumen protozoa support methanogenesis by producing hydrogen through hydrogenosomes or cytoplasmic hydrogenases[5]. Moreover, their bacterial predation leads to intraruminal protein recycling and increases wasteful ammonia release[6], despite their beneficial roles in pH homeostasis and fiber degradation. Although defaunation, the elimination of protozoa, can decrease ammonia and methane emissions, it impairs fiber degradation, highlighting the need for alternative strategies of protozoal modulation[7]. Due to the ban on antibiotic use in many countries, there is growing interest in using secondary plant compounds as mitigation strategies that may reduce nitrogen losses and CH4 emissions[8,9]. Using phytochemical-rich feed additives such as Acacia saligna, Leucaena leucocephala, and Atriplex halimus[10], Calotropis gigantea[11], Liquorice[12], and Citrus[13] have been substantially reported to suppress rumen protozoa populations, particularly the genus Entodinium, known as detrimental precursors of ammonia and enteric methane[14].
Calotropis gigantea (CG) belongs to the Apocynaceae family and is mainly found in the tropics and subtropics of Asia[15−17]. It is a tall, waxy flower, perennial undershrub, found chiefly in wastelands and commonly called crown flower, giant milkweed, wild cotton, apple of Sodom, and rubber tree[18,19]. Different parts of Calotropis gigantea and their extract are known for various medicinal properties[20]. It has been reported to have anti-rheumatic activity, antifungal activity, cytotoxic activity, antipyretic activity, activity against severe muscle spasms, malaria, asthma, paralysis, bronchitis, diabetes, dyspepsia, swellings, fever, epilepsy, and wound-healing activity[18,21−23].
Phytochemicals such as flavonoids have emerged as promising natural alternatives to antibiotics to improve productivity and mitigate environmental impacts[24] due to their rumen protozoa inhibitory activities[12,13]. Protozoa are the key drivers of ammonia release[25] and methane production[26,27]. Calotropis gigantea has been evaluated as a feed resource and feed additive for ruminants due to its bioactive effects on rumen protozoa and in-vitro fermentation. Daily intake is reduced in dairy cows supplemented with fresh leaves of CG due to unpalatability or the presence of toxic compounds[27]; however, ensiling or biotransformation reduces toxicity, increases milk yield and milk protein, and improves palatability for dairy cows while altering metabolite composition[14]. It is rich in flavonoids such as kaempferol, hesperidin, naringin, myricetin, and quercetin with documented antimicrobial[28] and protozoa inhibition effects (Entodinium), without affecting fermentation efficiency[11]. The plant was chosen based on its rich phytochemical content with medicinal properties, as well as the protozoa-inhibitory efficacy of the leaf powder. However, efficacy is influenced by dose, substrate, and complex interactions within the rumen microbiome[29]. Additionally, the effects of individual pure flavonoids present in whole mixed crude extracts, the potential synergistic interactions among flavonoid combinations, and the whole extract of Calotropis gigantea on microbial fermentation and rumen protozoa have not been investigated. This study aimed to assess the effects of pure flavonoid monomers (KAE, HES, NAR), their binary (KAE + NAR, KAE + HES, NAR + HES) and ternary combinations (KAE + HES + NAR), and Calotropis gigantea flavonoid-rich extract (CGFE) on rumen fermentation variables and rumen protozoa.
-
The Calotropis gigantea leaf samples were obtained from Honghe, Yunnan Province, China. The taxonomic name and authentication are preserved at the Kunming Institute of Botany, Chinese Academy of Sciences. The leaves were detached from the stems, left to dry in an open, shaded area in the laboratory, and then ground into powder using an electric grinder. The finely ground powder was stored in an airtight glass container until extraction.
Extraction process and quantification of major flavonoids
-
Ethanolic extraction of the air-dried leaf sample was carried out as described previously[13] with modifications. Briefly, 1 kg of Calotropis gigantea (CG) leaf powder was combined with 80% aqueous ethanol and extracted three times using a SCIENTZ-II D ultrasonic disruptor (Ningbo Scientz Biotechnology Co., Ltd, Ningbo, China) at 40 °C for 45 min with maximum power of 90%, and filtered using filter paper. The solvent in the extract was evaporated in a rotary evaporator at 40 °C and 80 rpm under gradually reduced pressure until it became concentrated. The concentrated extract residue was kept in a freeze dryer at −80 °C until it was dried, and then ground into a powder used for flavonoid analysis and in-vitro efficacy trials.
Major flavonoids in the Calotropis gigantea ethanolic extract were quantified using HPLC (Shimadzu Nexera-i LC-2040C 3D Plus series, Shimadzu Corporation, Kyoto, Japan), as described by Kingori et al.[30]. The oven temperature was maintained at 40 °C. The mobile phase comprised 0.1% aqueous phosphoric acid (mobile phase A) and HPLC-graded acetonitrile (mobile phase B). A gradient elution system was applied: 0.01 min (10% B), 30 min (55% B), 35 min (50% B), 38–40 min (10% B), with a flow rate of 1.0 mL/min and an injection volume of 20 μL. Hesperidin and naringin were quantified at 280 nm; kaempferol, quercetin, and myricetin at 371 nm. For analysis, 5 mg of dry ethanolic extract was dissolved in 5 mL of 80% (v/v) methanol to prepare a 1 mg/mL sample solution. A 1 mg/mL stock solution of standards was diluted to prepare working standard solutions (0, 50, 100, 200, 400, and 800 µg/mL), which were then filtered through a 0.22 µm membrane filter.
Artificial fermentation medium preparation
-
An artificial medium was prepared according to the methods described by Goering & Van Soest[31], with minor modifications. Briefly, a buffer solution was prepared by measuring 35 g NaHCO3 and 4 g NH4HCO3, and diluting with 1 L of distilled water. Then, macrominerals (5.7 g Na2HPO4, 6.2 g KH2PO4, and 0.6 g MgCl2) were weighed into another volumetric flask and diluted with 1 L of distilled water to make a solution. Finally, micromineral solutions were prepared by weighing 13.2 g CaCl2·2H2O, 10.0 g MnCl2·4H2O, 1.0 g CoCl2·6H2O, and 0.8 g FeCl2·6H2O, and diluting with 100 mL of distilled water. Additionally, a resazurin solution (0.1 g resazurin in 100 mL distilled water) was prepared and added to a round, flat-bottomed flask to monitor the reduction of the medium, indicated by a color change from red (oxidized) to colorless (reduced). The medium was bubbled overnight with CO2 and then reduced with a reducing agent (0.160 mg/dL NaOH and 0.625 g/dL Na2S·9H2O in 1,000 mL). The color change from pink to colorless indicated the reduction of the medium.
Rumen fluid collection, inoculum preparation, and in vitro fermentation
-
The rumen inoculum was collected 1 h before morning feeding from three non-lactating fistulated cows at the Changping Animal Experimental Base of the Institute of Animal Science, Chinese Academy of Agricultural Sciences. Rumen fluid was mixed at equal volume and transferred into pre-warmed thermos bottles. The thermally insulated bottles were filled and tightly capped to maintain anaerobic conditions and preserve microbial viability during transport to the ruminant nutrition laboratory. Upon arrival, the rumen fluid was filtered through four layers of cheesecloth into a plastic jar to remove larger feed particles, then placed in a 39 °C water bath under a continuous flow of CO2 to maintain anaerobic conditions.
Nine treatment groups, including control (CON), individual commercial flavonoids (Kaempferol [KAE], naringin [NAR], hesperidin [HES]; 4.5% dry matter [DM] each), their binary (KAE + NAR, KAE + HES, NAR + HES), and ternary (KAE + HES + NAR) combinations (1:1/1:1:1 ratios, 4.5% DM, respectively), and Calotropis gigantea flavonoid-rich extract (CGFE; 4.5% DM) were evaluated. Kaempferol, naringin, hesperidin, quercetin, and myricetin (all with purities greater than 98%) were purchased from Yuanye Biotechnology Co., Ltd, Shanghai, China. The dose was estimated from a study by Oskoueian et al.[32]. Each treatment had nine replicates. The dose for each treatment at 4.5% of DM was measured into individual 100-mL fermentation bottles containing 0.5 g total mixed ration (TMR) as a fermentation substrate. The filtered rumen fluid was mixed with the reduced artificial medium in a 1:2 (v/v) ratio under a continuous flow of CO2, transferred to the fermentation bottles, flushed with CO2, sealed with a rubber stopper, and incubated at 39 °C for 24 h.
Chemical analysis
-
Approximately 15 mL of cultured rumen fluid was centrifuged at 5,000 × g for 20 min at 4 °C, and the supernatant was collected to measure fermentation parameters, including pH, volatile fatty acids (VFA), and ammonium nitrogen (NH3-N). The pH value was measured using a digital pH meter (Sartorius, PB-10). VFA was analyzed by gas chromatography (Network GC System-6890N; Agilent Technologies). NH3-N concentrations were analyzed using an Agilent BioTek Epoch microplate spectrophotometer[33], in which a series of standard ammonia concentrations (0, 1, 2, 4, 8, 16, 32, and 64 mg/dL) was used for standard curve calibration. Methane was estimated using the stoichiometric equation: CH4 = 0.45 × (Acetate molar proportion) − 0.275 × (Propionate molar proportion) + 0.40 × (Butyrate proportion)[34,35]. Microbial crude protein (MCP) concentration was determined based on the standard procedures described by Makkar et al.[36].
Microscopic counting of rumen protozoa
-
Rumen protozoa were characterized and counted using a compound light microscope according to the methodology reported by Dehority[37] with minor modifications. Briefly, 1 mL of cultured rumen fluid was fixed with 1 mL of 50% formaldehyde to preserve the protozoan structure. Then, a 100 µL aliquot of brilliant green dye was added to each sample to stain the cells, left for at least 4 h for deep staining, and then diluted with 9 mL of 30% glycerol and mixed thoroughly. One mL of the mixture was placed in a Sedgewick-Rafter counting chamber (Thomas Scientific, Swedesboro, NJ, USA) and covered with a slide cover. Protozoal cells were counted at 100 × magnification. Fifty grids per sample were counted as provided in Supplementary Table S1, with cells touching the right or bottom grid lines excluded and those touching the left or top lines included. Protozoa were identified based on morphological characteristics, including ciliate zone localization, body size, spine and skeletal plate structure, and cilia tufts.
DNA extraction and quantification of total bacteria, and total methanogenic archaea
-
DNA extraction from a rumen fluid sample was conducted using a repeated bead-beating plus column purification method as described by Yu & Morrison[38]. The quality and concentration of extracted DNA were checked using a NanoDrop spectrophotometer (Invitrogen Corporation, Carlsbad, Calif, USA). Total bacteria absolute abundance was quantified by qPCR using universal bacterial primers (forward: 5'-CGGCAACGAGCGCAACCC-3'; reverse: 5'-CCATTGTAGCACGTGTGTAGCC-3') targeted 16S rRNA gene[39]. Total methanogens were quantified by qPCR using primers (forward: 5'-TTCGGTGGATCDCARAGRGC-3'; reverse: 5'-GBARGTCGWAWCCGTAGAATCC-3') specific to the Methyl coenzyme-M reductase A (mcrA) gene[40]. The concentration of DNA samples was standardized to 10 ng/μL. A final volume of 10 μL qPCR reaction containing 5 μL of SYBR Premix Ex Taq (2×), 0.25 μL forward primer, 0.25 μL reverse primer, 1 μL DNA template, and 3.5 μL nuclease-free water. Reactions were run in triplicate using a 7500 Real-Time PCR System (Applied Biosystems) under the following thermal cycling conditions: an initial denaturation at 95 °C for 2 min, followed by 40 cycles of denaturation at 95 °C for 15 s, and annealing at 60 °C for 1 min.
Statistical analysis
-
Analyzed data presented as mean ± standard error of the mean (SEM). Mean differences between rumen fermentation variables, microbial crude protein (MCP), protozoal counts, and bacterial and methanogen abundance were analyzed using one-way ANOVA, followed by Tukey's post hoc test for multiple comparisons. Spearman's correlation analysis was conducted to reveal the association between protozoal genera and fermentation variables. All statistical analyses were performed using R software (v4.4.0), with significance set at p < 0.05.
-
Calotropis gigantea leaf has a high dry matter content of (92.75 ± 0.26%) and substantial crude protein levels (17.02 ± 0.18%). It also contained considerable ether extract content (5.48 ± 0.32%), moderate fiber content (NDF: 30.30 ± 1.56%; ADF: 27.49 ± 0.24%), and low ash content (2.36 ± 0.01%) (Fig. 1a).
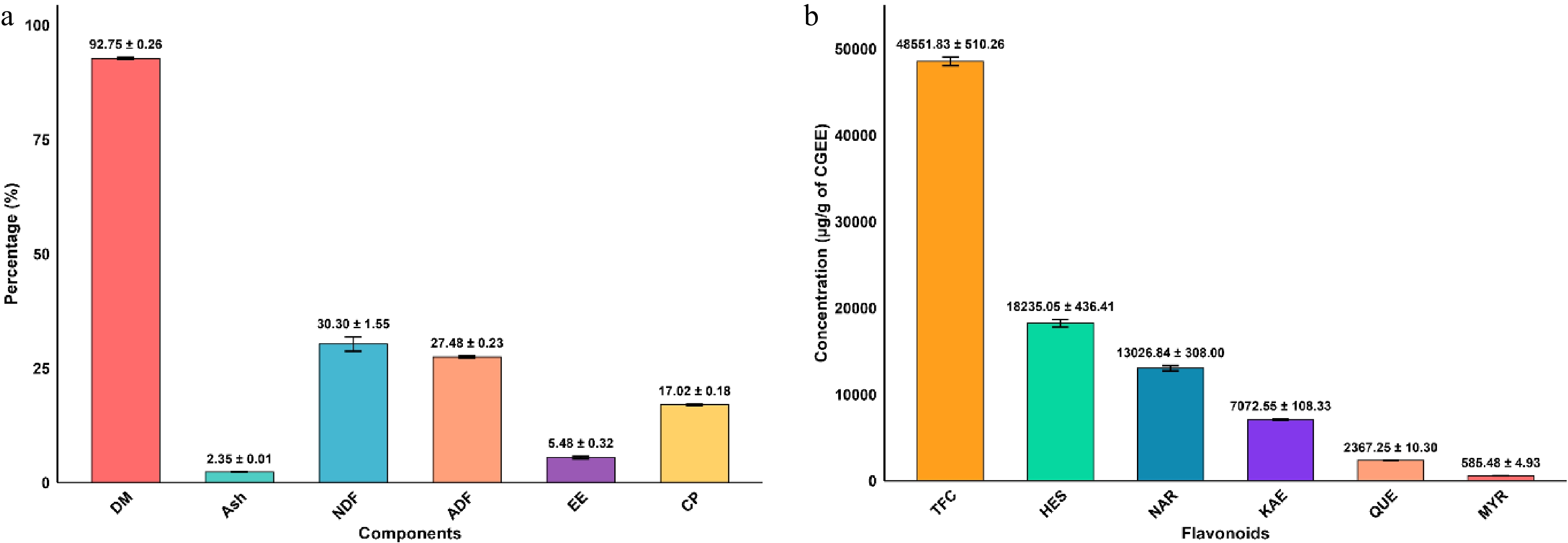
Figure 1.
(a) Proximate analysis of leaf powder showing dry matter (DM), ash, neutral detergent fiber (NDF), acid detergent fiber (ADF), ether extract (EE), and crude protein (CP) content. Descriptive statistical analysis of data presented in mean ± SD (% dry matter basis). (b) Flavonoid composition of Calotropis gigantea ethanolic extract, showing total flavonoid content (TFC) and individual flavonoid concentrations: hesperidin (HES), naringin (NAR), kaempferol (KAE), quercetin (QUE), and myricetin (MYR). Values presented as mean ± SD (µg/g extract).
The ethanol extract of leaves of Calotropis gigantea consisted of remarkably high total flavonoid content of 48,551.83 ± 510.26 quercetin equivalent (QE) µg/g, as shown in Fig. 1b. The HPLC analysis identified hesperidin as the predominant flavonoid (18,235.05 ± 436.41 µg/g), accounting for approximately 37.6% of total flavonoids, followed by naringin (13,026.84 ± 308.00 µg/g, 26.8%) and kaempferol (7,072.55 ± 108.33 µg/g, 14.6%). We also detected minor quantities of quercetin (2,367.25 ± 10.30 µg/g) and myricetin (585.48 ± 4.93 µg/g). These flavanone glycosides (hesperidin, naringin, and kaempferol), which were found at high concentrations, were used in the in vitro efficacy trial in this study.
Calotropis gigantea flavonoid-rich extract and ternary flavonoid blends reduce ammonia and methane production
-
CGFE and its bioactive components significantly modulated rumen fermentation (Table 1). Ruminal pH decreased in all treatments, although it remained within the normal physiological range (6.34–6.47). The most significant reduction was observed in CGFE (6.34), followed by NAR (6.37), and KAE + HES + NAR (6.40), as compared to the CON (6.47). Total volatile fatty acids (VFA) concentration increased in all treatments, with the highest values observed in CGFE (80.8 mM), and KAE + HES + NAR (62.4 mM). Acetate levels increased under KAE + HES (56.8), but declined significantly by HES + NAR (53.1), KAE + HES + NAR (53.3), and CGFE (47.3). Propionate proportion was decreased by KAE alone (17.4) but enhanced by CGFE (20.0). The enhancement of propionate production by CGFE is beneficial. Unlike acetate and butyrate, the propionate fermentation pathway does not liberate hydrogen; instead, it acts as a hydrogen sink and thereby may contribute to lower CH4 formation. Moreover, propionate is a gluconeogenic VFA and thus can increase the availability of energy to the mammary gland. Butyrate molar proportion increased in CGFE (27.9), with moderate increases in HES + NAR (25.4), and KAE + HES + NAR (24.7), relative to the CON (24.0). All branched-chain fatty acids and valerate were significantly increased in CGFE. The acetate-to-propionate ratio decreased markedly in CGFE (2.37), suggesting improved energy efficiency.
Table 1. Effect of flavonoid treatments on rumen fermentation characteristics, ammonia-N, methane, and microbial protein.
Parameters Treatments (4.5% DM) SEM p value CON KAE HES NAR KAE + HES KAE + NAR HES + NAR KAE + HES + NAR CGFE pH 6.47a 6.46ab 6.45ab 6.37cd 6.41abc 6.43abc 6.42abc 6.40bcd 6.34d 0.01 < 0.001 TVFA (mM) 45.2d 47.7cd 48.9cd 48.9cd 51.0cd 55.0bcd 56.2bc 62.4b 80.8a 2.17 < 0.001 Individual VFA (mol/100 mol) Acetate 54.3bc 55.7ab 56.1ab 54.8bc 56.8a 54.8bc 53.1c 53.3c 47.3d 0.38 < 0.001 Propionate 18.2b 17.4d 17.7bcd 18.0bc 17.5cd 17.8bcd 18.1bc 18.3b 20.0a 0.12 < 0.001 Butyrate 24.0bc 23.5b 22.8c 23.9bc 22.4c 23.9bc 25.4b 24.7b 27.9a 0.39 < 0.001 Isobutyrate 0.82b 0.78b 0.81b 0.76b 0.77b 0.82b 0.77b 0.82b 0.95a 0.02 < 0.001 Valerate 1.38c 1.35c 1.41bc 1.39c 1.34c 1.43bc 1.43bc 1.56b 2.17a 0.03 < 0.001 Isovalerate 1.25b 1.22b 1.21b 1.19b 1.18b 1.26b 1.24b 1.29b 1.58a 0.03 < 0.001 A:P ratio 2.99de 3.20ab 3.16abc 3.04cde 3.24a 3.08bcd 2.94de 2.91e 2.37f 0.03 < 0.001 NH3-N, mg/dL 19.4a 13.3cd 17.6ab 15.0bcd 13.2cd 13.7cd 15.9abc 12.9cd 12.0d 0.83 < 0.001 Methane*, mol/100mol 29.0bc 29.7a 29.5ab 29.3abc 29.7a 29.3ab 29.1bc 28.8c 27.0d 0.09 < 0.001 MCP, mg/mL 1.94bcd 2.01bc 1.10e 1.53cde 1.80bcd 1.77bcd 1.43de 2.13b 3.14a 0.11 < 0.001 * Estimated from standard stoichiometric equation: Methane = 0.45 × (Acetate proportion) − 0.275 × (Propionate proportion) + 0.40 × (Butyrate proportion)[1,2]. TVFA total volatile fatty acids, VFA volatile fatty acids, NH3-N ammonia nitrogen; MCP microbial crude protein. Fermentation parameters across treatments: control (CON), kaempferol (KAE), hesperidin (HES), naringin (NAR), their combinations, and Calotropis gigantea flavonoid extract (CGFE). Means with different superscripts differ (p < 0.05). SEM = standard error. Microbial crude protein (MCP) synthesis was significantly increased in CGFE (3.14 mg/mL, +61.9%) and KAE + HES + NAR (2.13 mg/mL, +9.8%), but was highly reduced by HES (1.01 mg/mL) and HES + NAR (1.43 mg/mL). NH3–N concentrations were markedly lower in all groups, with CGFE (12.0 mg/dL, −38.1%) and KAE + HES + NAR (12.9 mg/dL, −33.5%) exhibiting the most significant reductions relative to the CON (19.4 mg/dL).
CH4 was reduced in CGFE (27.0 mL/g DM, −6.9%) as compared to control (29.0 mL/g DM), consistent with its propionate-promoting effect, which is known as an alternative hydrogen sink pathway. It also reduced marginally by KAE + HES + NAR (28.8 mL/g DM). In contrast, methane was marginally increased by other treatments, likely due to elevated acetate production, which releases hydrogen as a substrate for methane synthesis.
Calotropis gigantea flavonoid-rich extract and ternary flavonoid combinations reduce protozoa
-
Rumen microbial populations were modified by the treatments (Table 2). Total protozoa was lower under CGFE (5.33 log10 cells/mL, −4.65%) as compared to the CON (5.59 log10 cells/mL). Similar reductions were observed with KAE + NAR (5.44 log10 cells/mL) and KAE + HES + NAR (5.33 log10 cells/mL).
Table 2. Effect of different flavonoid treatments on microbial populations.
Log10 cells/mL Treatments (4.5% DM) SEM p value CON KAE HES NAR KAE + HES KAE + NAR HES + NAR KAE + HES + NAR CGFE Total protozoa 5.59a 5.46bc 5.53ab 5.51abc 5.44c 5.55a 5.54ab 5.35d 5.33d 0.02 < 0.001 Isotricha 3.27 2.64 1.75 1.36 1.75 2.04 1.39 1.00 1.80 0.48 0.079 Dasytricha 1.33 0.97 0.68 0.65 0.68 0.68 0.68 0.65 0.65 0.46 0.977 Entodinium 5.59a 5.45bc 5.52abc 5.51abc 5.44c 5.54a 5.53ab 5.34d 5.32d 0.02 < 0.001 Epidinium 0.32 0.65 0.32 0.36 0.32 0.32 0 0.68 0.68 0.33 0.917 Diplodinium 0.97 1.72 1.00 1.82 0.71 0.71 0.32 0.68 1.00 0.48 0.433 Ophryoscolex 3.36 2.91 2.62 3.39 3.33 3.16 3.43 2.21 3.39 0.24 0.079 Prokaryotes (log10 copies/mL) Total bacteria* 11.9ab 12.6a 12.1ab 12.0ab 12.5ab 12.0ab 11.8ab 12.4ab 11.9ab 0.17 0.006 Total Methanogens⁑ 9.23d 10.1abc 9.76bcd 9.60bcd 9.37cd 10.3ab 10.2ab 10.6a 10.2ab 0.16 < 0.001 Control (CON), kaempferol (KAE), hesperidin (HES), naringin (NAR), Calotropis gigantea extract (CGFE). Means with different superscripts within rows differ (p < 0.05). SEM = standard error of the mean, * absolute abundance expressed as log10 copies of 16S rRNA genes per mL culture, ⁑absolute abundance expressed as log10 copies of mcrA genes per mL culture. At the genus level, CGFE, KAE, KAE + HES, KAE + HES + NAR, and CGFE treatments significantly reduced Entodinium, with CGFE (5.32 log10 cells/mL, −4.83%) and KAE + HES + NAR (5.34 log10 cells/mL, −4.47%) showing the lowest counts compared to the CON (5.59 log10 cells/mL). In contrast, Isotricha, Dasytricha, Epidinium, Diplodinium, and Ophryoscolex were not affected by any treatments.
Total bacterial abundance was increased by KAE (p = 0.006), while it remained unaffected by the other treatments. The absolute abundance of methanogens increased in all treatments, with the highest abundance observed in KAE + HES + NAR (10.6 log10 copies/mL, +14.8%), followed by moderate increases with CGFE (10.2, +10.5%), KAE + NAR (10.3, +11.6%), and HES + NAR (10.2, +10.5%). Minimal yet significant increases were observed with KAE (10.1, +9.4%), HES (9.76, +5.7%), NAR (9.60, +4.0%), and KAE + HES (9.37, +1.5%).
Correlation between protozoa and fermentation parameters
-
Correlation analysis revealed that Entodinium and total protozoa counts were positively correlated with several fermentation variables, as detailed in Fig. 2. They were positively associated with pH, NH3-N, acetate, and the A : P ratio and negatively correlated with propionate, butyrate, and microbial crude protein (MCP). The consistent negative association with MCP supports the hypothesis of protozoal predation on rumen bacteria impairing nitrogen utilization efficiency. Other protozoa genera, including Isotricha, Dasytricha, Diplodinium, Epidinium, and Ophryoscolex, showed no significant correlations with any measured fermentation parameters, indicating that Entodinium plays a predominant role in mediating protozoal-fermentation relationships under these conditions, although correlation cannot confirm causation.
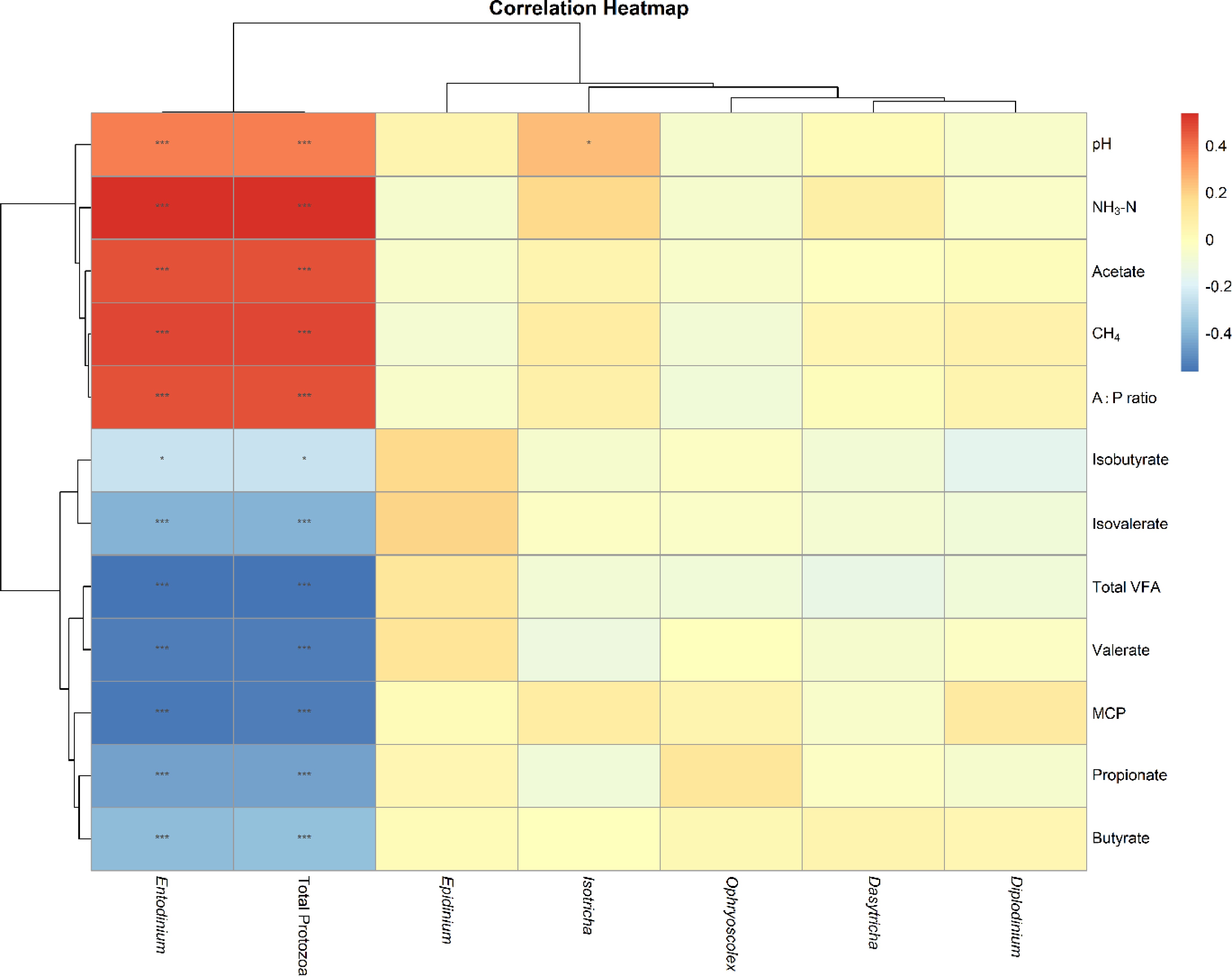
Figure 2.
Sperman's correlation analysis between protozoal genera and rumen fermentation parameters. Color intensity represents correlation strength (red: positive; blue: negative) with significance denoted as * p < 0.05, ** p < 0.01, and *** p < 0.001.
-
Rumen protozoa contribute to inefficient intraruminal nitrogen recycling through microbial protein degradation into ammonia, most of which is excreted as urea[41] and to methane production by supplying hydrogen to associated methanogens via interspecies hydrogen transfer[42]. Many countries ban antimicrobials, including ionophores, as feed additives, creating a pressing need for new, non-antimicrobial alternatives. The present study evaluated the effects of individual and combined flavonoid monomers, KAE, HES, and NAR, as well as CGFE, on key rumen fermentation parameters, including CH4 emission, NH3-N production, and protozoal populations. Among the nine treatment groups, KAE, KAE + HES, KAE + HES + NAR, and CGFE reduced total protozoa and Entodinium. NH3-N was reduced in all treatments. KAE + HES + NAR, and CGFE further reduced CH4 concentrations, acetate-to-propionate ratios, and pH, while increasing total VFA and MCP, indicating that they did not affect energy availability. Total methanogens increased in all treatments. Bacterial populations remained stable, except for significant increases in the KAE and the KAE + NAR groups. These findings support our hypothesis that ternary flavonoid blends and whole CGFE exhibit superior protozoa-inhibitory properties, compared with individual or binary flavonoid blends.
Flavonoid supplementation, particularly KAE + HES + NAR, and CGFE, markedly reduced ruminal NH3-N concentrations and decreased Entodinium populations, without altering total bacterial populations or impairing fermentation. The reduction in ammonia nitrogen is likely due to decreased protozoal-mediated microbial protein degradation[43]. Similarly, previous studies reported that pure flavonoids[32], flavonoid-rich citrus extracts[13], Eucalyptus globulus flavonoid-rich extracts[44], isoflavonoid-rich liquorice extracts[12], and many other plants[45] inhibited protozoa, particularly Entodinium, alongside reduced ammonia nitrogen. Moreover, Ayemele et al.[46] observed that dose-dependent inhibition of more than 50% of Entodinium species by giant milkweed leads to lowered NH3-N concentrations without impairing beneficial microbes. Protozoal inhibition, particularly of Entodinium, strongly correlates with reduced NH3-N[47], confirming their role as determinants of nitrogen waste via microbial protein degradation.
Up to 70% of dietary nitrogen in ruminants is excreted as urea[7,48], which can be volatilized as NH3, leached nitrate, nitrified, or denitrified as N2O to other reactive nitrogen species that are detrimental to air and water pollution[49,50]. Previous studies reported that urine patches deposited over 600 kg N/ha, with N2O emission factors 0.69% higher than those deposited and emitted from feces[51,52]. Thus, by inhibiting protozoal activity, particularly that of the predator Entodinium, flavonoids could decrease wasteful ammoniagenesis, enhance nitrogen utilization efficiency, and reduce environmental pollution.
In this study, the combination treatments KAE + HES + NAR and CGFE significantly reduced methane production. This might be due to the observed inhibition of total protozoa and the genus Entodinium, as well as the relatively low acetate proportion. In line with our findings, Kim et al.[53] reported that in-vitro supplementation with Camelia, Punica, Betula, Cudrania, and Ginkgo extracts reduced methane production, rumen protozoa, and some Gram-positive fibrolytic bacteria, such as Ruminococcus albus and Ruminococcus flavefaciens. Similarly, flavonoids of Mulberry leaf and resveratrol reduced methane emissions, while enhancing nutrient digestibility in sheep[54]. Moreover, an extract of liquorice, rich in isoflavonoids, reduced methane emissions by inhibiting protozoa[12]. This is further supported by findings that Mulberry leaf flavonoids reduced daily methane output alongside protozoa and methanogens in ewes, while increasing Fibrobacter succinogenes, Butyrivibrio fibrisolvens, and Ruminococcus albus fibrolytic bacteria[55]. The ternary flavonoid combination and CGFE further shifted metabolic fermentation toward propionate, resulting in a lower acetate to propionate ratio, which indicates the capture of hydrogen towards propionate synthesis, and enhances glucogenic energy supply[8]. Plant extracts such as garlic oil and secondary plant metabolites, benzyl salicylate, at doses of 300 and 3,000 mg/L, reduced acetate and increased propionate proportion with concurrent methane production reduction[56]. Various polyphenol-rich plant extracts reduce methane emissions and protozoa populations[45,57,58]. In line with this, Seradj et al.[59] observed that Bioflavex® and its pure flavonoids (for example, Neohesperidine, Naringine) reduced methane by decreasing the acetate-to-propionate ratio, and methanogens, without affecting the concentration of total volatile fatty acids.
Our findings indicated that supplementation with CGFE substantially reduced methane production, despite an increase in methanogen abundance. This is consistent with the findings of Newbold et al.[60], who observed a 47% methane reduction with (76%) removal of methanogens. This suggests that modulation of microbial fermentation and rumen protozoa, rather than the methanogen population, determines methane production. Inhibition of rumen ciliates, key producers of hydrogen for methanogens, is closely associated with enteric methane reduction[61]. Molecular evidence shows that all protozoa species have [FeFe]-hydrogenases regardless of the mitochondria-like organelle hydrogenomes, which are crucial to hydrogen metabolism[62,63]. This indicates that targeted inhibition of Entodinium is critical for CH4 mitigation. A meta-analysis by Fotsidie et al.[47] confirmed that a decrease in total protozoa population (25.6%) and Entodinium (74.4%) was significantly associated with reduced methane emission (200 g/d).
Targeted, rather than complete elimination of rumen protozoa is essential. Though defaunation can reduce methane production by 11%[64], it impairs fiber digestion by removing fibrolytic protozoa genera such as Epidinium and Polyplastron[65,66]. Therefore, selective inhibition of bacterivorous genera such as Entodinium, in our study, decreased ammonia and methane production, while maintaining microbial fermentation, which is critical for energy supply and rumen epithelial health.
In general, the results of the present study align with global efforts to achieve sustainability in developing climate-smart livestock production systems[67,68]. Flavonoid-rich plants, such as Calotropis gigantea, which was once considered a weed, are a natural, low-cost, drought-resistant, and potentially accessible alternative to antibiotic or synthetic feed additives like ionophores and monensin, thus raising food safety and health concerns among consumers[69]. The utilization of local, easily accessible, or underutilized plant resources, and low-cost feeding strategies by farmers further support circular bio-economy principles that enhance overall livestock production sustainability and resilience[70].
The present study provides evidence of the significant Entodinium-suppressing and acetate-reducing effects of CGFE and ternary flavonoid blends, leading to reduced ammoniagenesis and methanogenesis. However, there are also limitations. In the present study, we evaluated treatments at a single dose. Additionally, results from an in-vitro short-term study may not necessarily support findings in vivo or be extrapolated to other life stages. In addition, the persistence of effects, or effects that may emerge after a certain lag period, could not be properly evaluated. Therefore, further research should be conducted by incorporating multiple doses to assess the optimum dose response effect. Long-term animal studies are needed to confirm the persistence of the antiprotozoal, antimethanogenic, and ammonia-reducing effects, rumen microbiome adaptation or metabolic process changes, or any loss of efficacy over time.
-
This study demonstrated that total volatile fatty acids increased in all treatment groups. Flavonoid blends of KAE + HES + NAR, and CGFE significantly modulate microbial fermentation by increasing propionate proportions and microbial crude protein synthesis, while reducing acetate, ammonia waste, and methane. KAE, KAE + HES, KAE + HES + NAR, and CGFE inhibited the genus Entodinium, in which bacterial populations were maintained. The effects produced by CGFE and KAE + HES + NAR were superior to those made by individual pure flavonoids (KAE, NAR, and HES) and binary combinations, indicating the significance of synergistic interactions among bioactive compounds in modulating the rumen microbiome and fermentation metabolism. Therefore, CGFE is rich in flavonoids and may be a promising feed for ruminants to improve nitrogen utilization efficiency and mitigate environmental impacts. Future research should explore the long-term effects on digestibility, productivity, nitrogen metabolism, and methane emissions across different ruminant species.
-
The animal handling and care protocols during the collection of rumen fluid from dairy cows were approved by the Animal Care Advisory Committee of the Institute of Animal Science, Chinese Academy of Agricultural Sciences.
-
The authors confirm their contributions to the paper as follows: conceptualization, methodology: Abrhaley A, Bu D; data curation, investigation, formal analysis, software, writing-original draft, visualization: Abrhaley A; Validation: Abrhaley A, Zhan T, Bu D; writing -review editing: Abrhaley A, Zhan T, Ma L, Li X, Bu D; resources: Li X, Bu D; project administration, and funding acquisition: Ma L, Bu D; Supervision: Zhan T, Bu D. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
-
This study was supported by the National Key Research and Development Program of China (No. 2022YFD1301002), Beijing Innovation Consortium of Livestock Research System (BAIC05-2025), the Agricultural Science and Technology Innovation Program (ASTIP-IAS-17), the Central Public-interest Scientific Institution Basal Research Fund (No.2025-YWF-ZX-07), and the International Atomic Energy Agency Technical Co-Operation and Assistance Programme (No. CPR5029).
-
The authors declare that there are no financial or personal conflicts of interest that could have influenced the results presented in this paper.
-
accompanies this paper online at: https://doi.org/10.48130/cas-0026-0016.
- Supplementary Table S1 Raw counts and corresponding transformed concentrations (cells/mL) of rumen protozoa.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Abrhaley A, Zhan T, Ma L, Li X, Bu D. 2026. Calotropis gigantea flavonoid blends reduce wasteful ammoniagenesis and methanogenesis by inhibiting rumen protozoa. Circular Agricultural Systems 6: e015 doi: 10.48130/cas-0026-0016 

Calotropis gigantea flavonoid blends reduce wasteful ammoniagenesis and methanogenesis by inhibiting rumen protozoa
- Received: 16 February 2026
- Revised: 27 April 2026
- Accepted: 28 April 2026
- Published online: 29 June 2026
Abstract: Ammonia (NH3-N) and enteric methane (CH4) contribute to nutritional inefficiencies and environmental impacts. This study evaluated the effects of Calotropis gigantea flavonoid-rich extract (CGFE), individual flavonoids (kaempferol [KAE], hesperidin [HES], naringin [NAR]), and binary (KAE + NAR, KAE + HES, NAR + HES) or ternary combinations (KAE + HES + NAR), each at a dose of 4.5% dry matter (DM), on rumen fermentation parameters and rumen protozoa in an in-vitro batch fermentation system incubated for 24 h. Rumen protozoa were counted microscopically, while bacteria and methanogens were quantified by qPCR targeting 16S rRNA and mcrA genes. Data were analysed using one-way ANOVA followed by Tukey's test and Spearman correlation in R v4.4.0. CGFE has a total flavonoid content (TFC) of 48,552 ± 510 µg/g, with HES (37.6%) and NAR (26.8%) as the major components, followed by KAE (14.6%). CGFE and KAE + HES + NAR showed the most pronounced effects. Both increased total volatile fatty acids (VFA) and microbial crude protein (MCP), reduced the NH3-N, acetate-to-propionate ratio, and methanogenesis (p < 0.001). Entodinium was suppressed by CGFE (5.32), KAE + HES + NAR (5.34), KAE + HES (5.44), and KAE (5.45), as compared to the control (CON, 5.5) (p < 0.001), while other genera remained unaffected (p > 0.05). In conclusion, CGFE and ternary flavonoid combinations reduce ammonia and methane, likely through protozoa inhibition and acetate reduction, suggesting the potential of CGFE as a sustainable feed additive for ruminants.
-
Key words:
- Calotropis gigantea /
- Flavonoids /
- Methane /
- Rumen fermentation /
- Rumen protozoa