








-
CRISPR/Cas systems have emerged as one of the most popular and advanced tools for genome editing. As early as in 1987, CRISPR sequences were first discovered in the iap gene of Escherichia coli, which functions in providing resistance against bacteriophages and enhancing adaptive immune responses in prokaryotes[1,2]. Repurposing CRISPR/Cas systems as a gene editing tool in eukaryotic cells has revolutionized the field of genome engineering[3,4]. This technology has been rapidly expanding and applied to form translational medicine, diagnostics and therapeutics to other fields[5−7].
In 2013, the CRISPR/Cas9 system was first established in Arabidopsis thaliana[8], Nicotiana benthamiana[9], rice (Oryza sativa) and wheat (Triticum aestivum)[10] showing prospects of wider application of CRISPR/Cas technologies in plants. Since then, continuous improvements in CRISPR/Cas systems, including CRISPR/Cpf1[11], base editors[12] and prime editors[13], have made plant genome editing easier and they have become more widely used as well as being low-cost and highly efficienct precision genetic manipulation tools. An ever-increasing number of crops and other plant species have been modified by genome editing for gene functional validation, trait improvement, and plant virus detection, among other uses[14−16]. CRISPR/Cas technologies thus have the potential to change the landscape for conventional breeding by aiding the introduction of target traits more accurately, in less time. More importantly, CRISPR/Cas differs from transgenic breeding technology in the fact that there is no transfer of exogenous genes into the target genome, paving the way to reduced biosafety concerns. Here, we describe a series of CRISPR/Cas systems and summarize the recent advances in those systems and their applications in plant genome editing.
-
CRISPR/Cas systems exist in various species of bacteria and archaea providing adaptive immunity against viruses, and they vary in composition and mechanisms of action. The defense process is mainly accomplished by Cas proteins and CRISPR array through three sequential steps: adaptation, expression and interference[17]. CRISPR systems have been divided into two classes that were additionally grouped into six types based on the easily recognizable signature of Cas proteins[18,19] (Fig. 1). Typically, class 1 CRISPR/Cas systems (comprising types I, III, and IV; represent about 90% of the CRISPR-Cas loci) employ multi-subunit protein complexes to target DNA or RNA for processing and cleavage. By contrast, class 2 systems (comprising types II, V, and VI; represent about 10% of the CRISPR-Cas loci) employ a single multidomain protein in complex with crRNAs or alternative tracrRNA[20,21]. The extensive diversification of the CRISPR/Cas systems offers potential expansion of the gene editing toolbox from the original single system CRISPR/Cas9 to a rich toolbox.
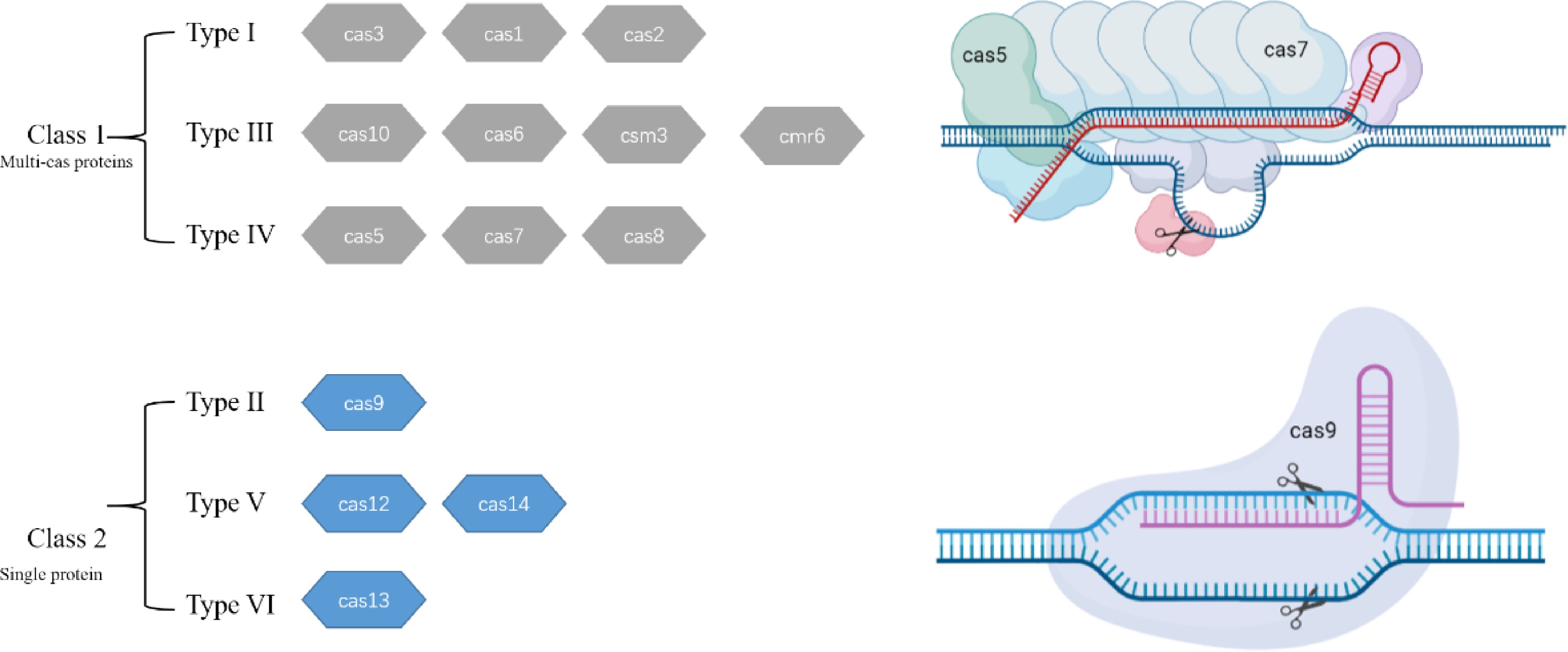
Figure 1.
The classification of CRISPR/Cas systems. Representative operons for each type are shown here. Gray represents Cas systems employing multi-subunit protein complexes to cut target DNA or RNA; blue represents Cas employing a single multidomain protein in complex for sequence cleavage.
CRISPR/Cas9
-
CRISPR/Cas9 system belongs to the type II of class 2 CRISPR systems and is the most well-characterized, most popular and widely used system for genome editing in a number of organisms[20,22]. Among the CRISPR/Cas systems, the Cas9 protein was the first used system to introduce DNA double-strand break (DSB) in target DNA by an RNA duplex formed by crRNA and tracrRNA[17]. Cas9 nuclease contains two domains RuvC-like and HNH, which cleave the non-complementary and complementary strands of the target DNA, respectively[23] (Fig. 2). The dual RNA, crRNA and tracrRNA, designed a single guide RNA (sgRNA) together with the Cas9 protein which makes up the simplified two-component system. The binding of sgRNA by Cas9 in cells results in the formation of a ribonucleoprotein complex, which is then guided by 20 nt sgRNA to bind to protospacer adjacent motif (PAM, 5'-NGG-3') that achieves cleavage of target DNA. There are two major pathways for cellular DNA repair for DSBs caused by DNA cleavage[24]. One is homologous recombination (HR) that recombines and integrates donor DNA into the position of DSBs, and the second is non-homologous end joining (NHEJ) that is error-prone to induce insertion, deletion or substitution of individual bases. Under natural conditions in eukaryotic cells, HR is of low probability, whereas NHEJ happens more often. Therefore, gene modification and genome editing can be achieved through DSBs and subsequent DNA repair.
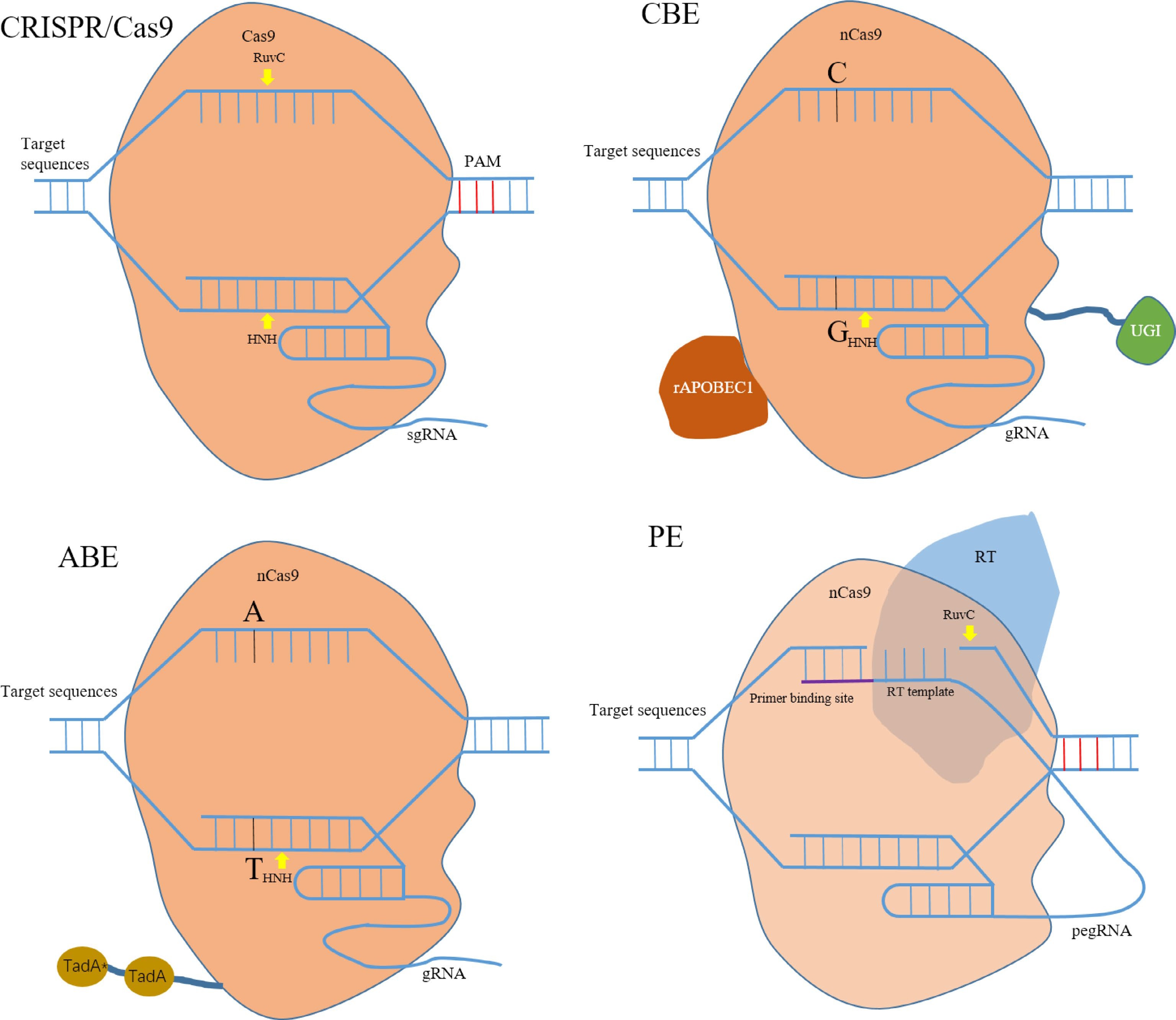
Figure 2.
Composition of CRISPR/Cas system. CRISPR/Cas9, the Cas9 system is composed of a Cas9 protein and a sgRNA. The yellow arrow represents DNA double-strand break (DSB) induced by RuvC and HNH domains. CBE, a base editor is a fusion of catalytically deactivated Cas9 domain and deaminase domain. CBE deaminates cytosine to generate uracil (C-U), then converts the uracil into thymine (U-T) by DNA repair response, eventually effecting a C-T (or G-A) substitution. ABE deaminates adenosine to generate inosine (A-I), which is read as guanosine (G) by polymerase enzymes, eventually effecting a A-G (or T-C) substitution through DNA repair or replication. PE, Prime editor consist of two simple operational components, a fusion protein made by the fusion nickase Cas9 protein (nCas9) and reverse transcriptase (RT) and a pegRNA contains sgRNA, primer binding site and reverse transcription template.
CRISPR/Cas12
-
The characteristics of Cas12 is similar to Cas9 with the difference that Cas12 proteins have multiple domains, but contain a single RuvC-like endonuclease domain[25]. Cas12a system, also known as Cpf1 because of its discovery in Prevotella and Francisella 1, belongs to type V-A of class 2[11]. Compared to recognizing to G-rich PAMs by the Cas9 system, Cas12a induces target DNA cleavage preferably by binding to 5′-TTTN-3′ (T-rich) PAMs (Table 1). Additionally, Cas12a cleaves target DNA using Cas12a-crRNA complex without the requirement of tracrRNA that produces a staggered DSB (sticky ends) compared to blunt ends produced by Cas9[11,26]. This unique ability in crRNA processing has been utilized for multiplex gene editing[27]. Thus, the discovery of Cas12a systems expands the CRISPR toolbox providing an alternative genome editing tool for other application scenarios.
Table 1. Comparative analysis between Cas9, Cas12, and Cas13.
Cas9 Cas12 Cas13 Class 2 2 2 Type II V VI Subtypes II-A,B,C V-A(Cas12a); V-B(Cas12b); V-C(C2c3) VI-A(Cas13a/C2c2); VI-B(Cas13b); VI-C(Cas13c);
VI-D (Cas13d)Targets dsDNA ssDNA RNA Nuclease domain(s) RuvC and HNH RuvC Two HEPN Guide RNA crRNA and tracrRNA crRNA crRNA Cleavage type Blunt ended dsDNA break 5′ overhang dsDNA break ssRNA Size 1000−1600aa 1100−1300aa 900−1300aa Guide spacer 18−24nt 18−25nt ~64nt PAM NGG (SpCas9) TTTN (FnCas12a) 3′ non-G PFS (LasCas13a) NGN (SpCas9-NG) TYCV (LbCas12a-RR) D-(PS)-NAN/NNA (BzCas13b) NNG (ScCas9) TATV (LbCas12a-RVR) NNGRRT (SaCas9) NNNNGATT (NmCas9) NNNVRYM (CjCas9) NNAGAAW (StCas9) PAM-less (SpRY) Repair mechanism NHEJ and HDR NHEJ and HDR No Applications Most widely used Majorly used in epigenetic editing Majorly used in disease detection and diagnostics In addition to the Cas12a system, the variants of Cas12, AaCas12b, BhCas12b, AacCas12b and BthCas12b derived from different bacteria, have been harnessed and engineered for editing in mammalian genomes and plant genomes with high targeting specificity[28−30]. Cas12e (also known as CasX) derived from Deltaproteobacteria (DbCas12e) was proven to produce extensive 5ʹ-overhangs at cleaved targets in mammalian genomes[31]. Cas12d (also known as CasY) recognizes a 5ʹ-TA PAM and cleaves dsDNA[32]. Cas12g, smaller than other Cas12 effectors that only contain ~800 amino acids, is an RNA-guided ribonuclease with collateral RNase and single-strand DNase activities[25,33]. The characteristic structures of Cas12h and Cas12i are similar to that of Cas12a in that they use a single crRNA to target dsDNA without tracrRNA[33]. Cas12f is a miniature CRISPR/Cas system that cleaves dsDNA rely on PAM recognition, such as AsCas12f1 (422 amino acids), which recognizes 5ʹ T-rich PAMs and create staggered DSB to target DNA[34,35]. The most recent report found that Cas12c directly induces DSB in a unique mechanism without DNA cutting, which may be repurposed to represses transcription[36].
CRISPR/Cas13
-
The Cas13 system, which belongs to type VI of class 2, is further classified into Cas13a (also known as C2c2), Cas13b, Cas13c and Cas13d[21]. Cas13 employs a single crRNA to cleave target single-strand RNA by its two HEPN domains (Table 1). Furthermore, Cas13 has two distinct catalytic activities; an RNA recognition and a cleavage activity[37,38]. These characteristics make Cas13 a versatile RNA editing tool and differs from other DNA manipulation tools. RNA Editing for Programmable A to I Replacement (REPAIR) platform was designed to edit nucleic acids at the RNA level in mammalian cells using the complex obtained by fusing adenosine deaminase acting on RNA type 2 (ADAR2) with catalytically-inactive Cas13 (dCas13)[39]. Cas13 orthologs, such as LwaCas13a, PspCas13b and LshCas13a, have been engineered for RNA knockdown and binding[40]. Among them, LshCas13a was optimized through plant codon and had been engineered to effect RNA interference targeting plant virual RNA sequences of Turnip Mosaic Virus (TuMV)[41]. Besides, the platform specific high-sensitivity enzymatic reporter unlocking (SHERLOCK) which is developed on CRISPR/Cas13a system, has made great achievements in the field of virus detection, especially in response to the recent SARS-CoV-2 outbreak[7].
Other CRISPR/Cas systems
-
Recently, additional CRISPR/Cas systems have been discovered and characterized, which differ from well-known Cas9, Cas12 and Cas13 systems. Cas3 system, which belongs to type I of class 1 CRISPR systems, recruiting CRISPR-associated complex for antiviral defense (Cascade) in trans for processive DNA degradation to programmable genome deletion and large-scale genome engineering[42,43]. Repurposing Cas3 systems to genome editing in Zea mays, shows its potential to become a powerful tool for plant genome streamlining[44]. Besides, a type I-D (TiD) system using Cas10d nuclease instead of Cas3 nuclease that engineered to generate small indels, long-range deletions and bi-directional DNA deletions in the tomato genome[45]. Cas10 system belongs to class 1 Type III CRISPR systems and the standout feature is its unique cas10 protein[46]. CRISPR/Cas10 system, a complex that is composed of multiple subunits and binds to crRNAs, has been used to edit phage genomes[47]. Cas14 is particularly unique for possessing a single-stranded DNA targeting activity and distinguishing single-nucleotide polymorphisms as its half the size of typical class 2 CRISPR effectors[48]. Thus, the CRISPR/Cas14 system has been engineered as an ideal tool for detecting ssDNA viruses in plant, and may be used widely in the diagnostics field[49].
-
Even CRISPR/Cas9, the most widely adopted CRISPR/Cas technology, has some flaws and negatives, such as PAM dependence, off-target effect and so on. Generally, the optimization of CRISPR/Cas9 system in plants mainly involves Cas9 protein, sgRNA and regulatory elements for Cas9 and gRNA (e.g. promoter, codon and terminator). To increase the targeting range, Cas9 protein has been engineered to recognize different PAMs, such as SpCas9-NG (NGN), xCas9 (NG, GAA and GAT), EQR-Cas9 (NGAG), VQR-SpCas9 (NGAN and NGCG) and VRER-SpCas9 (NGCG)[50,51]. A large number of Cas9 orthologs from other bacteria provide potential candidates for engineering Cas9 proteins with different PAM sites, such as StCas9 (NNAGAAW), CjCas9 (NNNVRYM) and SaCas9 (NNNRRT), among others[52,53]. In particular, using an engineered SpRY Cas9 variant made highly efficient genome editing target relaxed PAM sites in rice[54]. Similarly in Cas12 systems, different PAMs, such as TYCV, TATV, CCCC, and TATG, were also recognized by Cas12a variants to expand the scope of targeting[26,55].
To decrease undesired mutagenesis at off-target sites, truncated gRNAs (17-18 nt), engineering a hairpin secondary structure on the 5' end of the sgRNA and chemical modification at the sgRNA sequence sites have been used and proven effective[56−58]. The off-target effects were also effectively reduced by fusing Cas9 protein with programmable DNA-binding domain[59]. With the continuous efforts of researchers, high-fidelity variants (SpCas9-HF1), enhanced-specificity variants (eSpCas9) and several more new variants have been engineered to improve targeting specificity and precision[60,61]. In addition, many bioinformatics tools were developed to evaluate off-target effects and help design highly specific sgRNAs, including E-CRISP, CRISPR-P, and CRISPOR[62−64] (Table 2).
Table 2. List of available web tools for designing CRISPR/Cas systems.
Name Advantage Web address BE-Designer Provide all possible base editor target sequence http://www.rgenome.net/be-designer/ BEtarget Design guide RNAs for base editing in plants http://skl.scau.edu.cn/betarget/ Breaking-Cas Design gRNAs for CRISPR/Cas experiments http://bioinfogp.cnb.csic.es/tools/breakingcas Cas-Analyzer Analyze next generation sequencing data http://www.rgenome.net/cas-analyzer/ Cas-Designer Design CRISPR targets http://www.rgenome.net/cas-designer/ Cas-OFFinder Search for potential off-target sites http://www.rgenome.net/cas-offinder CCTop Predict CRISPR/Cas9 target http://crispr.cos.uni-heidelberg.de CGD Predict the efficacy of gRNAs http://big.hanyang.ac.kr:2195/CGD CHOPCHOP v3 Identify CRISPR–Cas sgRNA targets https://chopchop.cbu.uib.no/ COSMID Searches genomes for potential off-target sites http://crispr.bme.gatech.edu CrisPam Detect PAMs for allele-specific targeting https://www.danioffenlab.com/crispam CRISPOR Find guide RNAs and evaluate potential off-targets http://crispor.org CRISPRdirect Select rational CRISPR/Cas targets http://crispr.dbcls.jp/. CRISPRscan Design sgRNAs for CRISPR/Cas9 targeting in vivo https://www.crisprscan.org/ CRISPy-web Design sgRNAs for microbial genome http://crispy.secondarymetabolites.org/ CRISPR-ERA Design sgRNAs for gene regulation http://CRISPR-ERA.stanford.edu CRISPR-P 2.0 Improved CRISPR-Cas9 tool for genome editing in plants http://cbi.hzau.edu.cn/CRISPR2/ E-CRISPR Design sgRNAs in a gene-by-gene fashion http://www.e-crisp.org/E-CRISP MMEJ-KO Design gRNAs for MMEJ-mediated fragment deletion http://skl.scau.edu.cn/mmejko/ Off-Spotter Identify all potential off-target sites https://cm.jefferson.edu/Off-Spotter/ PhytoCRISP-Ex Offer Cas9 target prediction using phytoplankton genomes http://www.phytocrispex.biologie.ens.fr/CRISP-Ex/ The addresses of some web tools could not be resolved, this could be a temporarily unable service so they were not included. However, off-target effects are not so unacceptable in plants compared to that in clinical practice[65,66]. In fact, off-target effects in plants are rare. An off-target effects analysis using whole-genome sequencing found off-target mutations were negligible in rice plants edited by Cas9 and Cpf1[67]. Furthermore, any unwanted mutations or negative-effect mutations produced by off-target events could be segregated out through genetic crosses and during sexual reproduction[68]. In crop molecular breeding, as long as it can lead to good traits, it is a good goal, whether the mutation is caused by on-targets or off-targets. Therefore, off-target effects will not be a great hurdle for genome editing to be applied in plants, particularly crop plants, which play a major role in agriculture.
Base editing
-
Base editing (BE) is a point mutation gene modification technology developed on the CRISPR/Cas9 system. Base editing induces R-loop formation to enable nucleotide substitutions without DSB and donor templates, thereby greatly reducing the random insertion, deletion, translocation and other genomic mutations caused by DNA repair[69,70]. A base editor is a fusion of catalytically deactivated Cas9 domain (Cas9 variants) and deaminase domain, usually divided into cytosine base editors (CBEs) and adenosine base editors (ABEs) according to their deaminase domain[71,72]. CBEs can only convert a C•G base pair to a T•A base pair, whereas ABEs convert an A•T base pair to a G•C base pair (Fig. 2). On the basis of the original prototype, a dual adenine and cytosine base editor (A&C-BEmax) and adenine and cytosine base editor (ACBE) were developed to simultaneously achieve C to T and A to G conersions in mammalian cells by fusing both cytosine and cytosine deaminases, whereas saturated targeted endogenous mutagenesis editors (STEMEs) were developed similar to rice[73−75]. Moreover, C-to-G base editors (CGBE), glycosylase base editor (GBE) and UNG-nCas9-AID base editors were developed to convert C to G in human cells, C to G in mammalian cells, and C to A in bacteria, respectively[76,77]. AGBE, a new type of dual deaminase-mediated base editing system that fusing a CGBE with ABE, simultaneously generated four types of base conversions (A to G and C to G / C to T / C to A) as well as indels, thereby greatly extending the diversity of mutations in the same DNA strand at the target sites[78]. Besides being used to mediate specific DNA transition mutations, base editors are also used to directly edit RNA transcript adenosine to inosine (A to I) or cytidine to uridine (C to U) by using a catalytically dead Cas13 (dCas13) and an enzyme adenosine deaminase acting on RNA (ADAR)[39,79]. A large number of base editors have been generated and optimized for different plant species to obtain excellent character trait variations and help crop improvement. Plant based editing systems were first tested in rice, tomato, wheat and maize, proving the feasibility of introducing altered phenotypes[12,80−82]. However, bystander mutations, indels, and off-target single nucleotide variations (SNV) were also observed in plants and animal cells after processing by BEs[83]. A higher number SNVs occur in the rice genome after editing by CBE compared to that after editing by ABE, which is supposed to be caused by random deamination[84,85]. Recently, a novel base editing system TAC9-ABE was developed for eliminating Cas9-dependent DNA off-target in human cells, which provides a potentially safe base editing tool for the breeding of gene edited plants and animals[86].
Prime editing
-
Prime editing (PE) is a major breakthrough beyond the CRISPR/Cas9 system, which can effectively generate the free conversion of all single bases, and can also effectively enable the precise insertion and deletion of multiple bases. Prime editor is significantly modified into two aspects; one, to transform Cas9 protein into nickase and then fuse it with reverse transcriptase to form a new protein, and two, to add a sequence including the primer binding site and the reverse transcription template at the 3' end of sgRNA. The newly formed sgRNA is referred to as pegRNA[13]. PE systems just consist of two simple operational components, the fusion protein and pegRNA (Fig. 2). Because of its simplicity, precision and versatility, PE was applied in plant research soon after its invention, especially in rice[87−89]. However, the editing efficiency of prime editors in plants is not high and at some sites even lower than that edited by base editors[87]. Therefore, PE needs further improvement to be applied more efficiently in plants.
To improve PE efficiency, several strategies have been used in plants including engineering pegRNAs and fusion protein, driving promoter replacement and optimizing editing conditions. A dual-pegRNA strategy greatly improved PE at most of the targets (13/15), which boosted PE efficiency 4.2-fold on average[90]. Through incorporating structured RNA motifs or fusing the 20-nt Csy4 recognition site at the 3' end of the pegRNA, PE efficiency was improved[91,92]. Strategic co-expressing MLH1dn to transiently inhibit DNA mismatch repair (MMR) enhances PE efficiency by at least 2.2-fold[93]. Replacing C-terminal reverse transcriptase Cas9 nickase fusion with N-terminal fusion could increase PE efficiency about 2- fold to 3-fold in rice[94]. Through engineering the fusion protein that removes the ribonuclease H domain of Moloney–murine leukemia virus reverse transcriptase (M-MLV RT) and incorporating a viral nucleocapsid protein with nucleic acid chaperone activity, can independently improve plant prime editing efficiency by ~1.8−3.4-fold in plant cells[95]. In addition, suitable high-temperature treatment could improve the PE efficiency in rice protoplast at least 2.8 fold[96]. While each modification at present independently improves PE efficiency several fold, combining these strategies may further improve the efficiency.
-
CRISPR/Cas technologies have developed to such an extent that they are transforming different fields of research, including agriculture and plant biology. CRISPR/Cas technology provides simple, highly efficient, low cost, and scalable tools for the research in plant biology and crop breeding, and has quickly been embraced by plant researchers. More importantly, the diversity of CRISPR/Cas tools is conducive to meeting the editing needs of arbitrary target sequences in different plant genomes.
Crop improvement and precision breeding
-
Although CRISPR/Cas technology has been the preferred tool for plant genome editing and genetic operation for a short time, it has already demonstrated great application value, especially in crop genetic improvement. Genome editing provides a new toolkit for plant precision breeding and offers more crop improvement strategies[97]. CRISPR/Cas systems and CRISPR/Cas-derived base editors have emerged as powerful tools not only for DSB-mediated genome editing, but also for generating programmable single DNA base changes. Furthermore, prime editors work as another precise genome editing tool that can enable all base conversions and targeted precise insertions. The precise deletion and insertion of DNA or RNA has made it possible to manipulate gene functions and stack crop traits. New alleles of betaine aldehyde dehydrogenase 2 (OsBADH2) have been created using CRISPR/Cas9 systems, which improved aroma in non-fragrant rice varieties[98]. Producing CRISPR/Cas9 mutants with knocked out OsERF922 in rice, enhanced rice blast resistance[99]. Through editing the uORF of FvebZIPs1.1 in strawberry (Fragaria vesca) by A3A-PBE, a series of novel genotypes with a continuum of sugar content variations were generated that achieved fine-tuning of quantitative traits[15]. In addition, CRISPR/Cas-based systems could be used to knock-up target genes in rice by editing structural variations to create new genes and traits[100].
Complex traits like growth and development are controlled by multiple genes that require multiplex genome editing and modification. CRISPR/Cas9 has been successfully used to knock-out flavanone-3-hydroxylase 1 (GmF3H1), -2 (GmF3H2) and flavone synthase II 1 (GmFNS II-1) in soya bean, leading to improved isoflavone content and resistance to soya bean mosaic virus (SMV)[101]. Utilizing CRISPR/Cas9-mediated system to directly knock-out 48 growth-related genes in maize created a batch of mutants with bigger leaf compared to the control[102]. Most of the important agronomic traits are significantly associated with single nucleotide polymorphisms (SNPs) and small indels, which are suitable for precise substitution and mutation using BEs and PEs[70]. Enhancing resistance to powdery mildew has been achieved through editing mildew resistance Locus O (MLO) family genes in various plant species, including wheat, tomato, and grapevine[14,103,104]. Enhancing broad-spectrum disease resistance to bacterial pathogens by using CRISPR/Cas9-mediated genome editing for DMR6 orthologue in banana[105]. To date, many traits including yield, quality, and biotic stress and abiotic stress resistance have been improved using CRISPR/Cas systems though knock-in or knock-out methods. Therefore, CRISPR/Cas technology is an effective approach, for improving haploid breeding in maize and rice[106,107], for shortening growth times in rice[108], for reducing cassava brown streak disease (CBSD) symptom severity and incidence in cassava[109], for increasing silique shatter resistance in Brassica napus[110] and overcoming self-incompatibility in diploid potato[111], to meet breeders’ requirements. Moreover, de novo domestication strategies with CRISPR/Cas genome editing technologies as the core, has been proposed and then applied in tomato, ground cherry and rice, which accelerated the process of domesticating crops[112−114]. There is no doubt that it is an exciting route and direction for future crop genetic improvement.
Epigenetic modification and regulation
-
Epigenetic mechanisms comprise DNA methylation, histone modifications, chromatin remodeling and RNA processing that play an essential role in the regulation of plant growth and development, underlie their complex traits and contribute to evolution[115,116]. Single cytosine methylome analyses of the CmROS1 knockout mutant that was induced by CRISPR/Cas9 systems revealed DNA methylation changes regulating fruit ripening in melon[117]. Constitutively expressing catalytically inactivated dCas9 fusion proteins further expand the application of CRISPR/Cas system for epigenetics studies without inducing DSBs[118]. Manipulated epigenetic modifications, such as CRISPR-mediated activation (CRISPRa) and CRISPR-mediated interference (CRISPRi), have already been achieved using dCas9-based tools in mammalian genome system[119]. Moreover, programmable epigenetic platform CRISPRoff triggers highly specific DNA methylation to silence genes, whereas CRISPRon removes DNA methylation to reverse silenced genes[120]. A dCas9-SunTag-VP64 system was adopted in Arabidopsis for activating DNA methylated genes, which provides a valuable tool for site-specific manipulation of the plant epigenome[121]. The MS2-CRISPR/dCas9 system was developed and validated, to enable manipulation of Arabidopsis flowering time by targeting modulate epigenetic status and transcriptional activity of FT gene[122]. Although only a few cases of CRISPR/dCas-mediated application were reported in plants and may be related to the off-target effects, it is conceivable that more studies will improve targeting specificity and efficiency of these epigenome editors and expand their application in plants in the future[123].
Nucleic acid detection and disease diagnostics
-
The CRISPR/Cas systems have been recognized for their promising applications in detecting different human and plant diseases and Cas9 have been used for miRNA detection[6,124]. In particular, the improvement of CRISPR/Cas12 and CRISPR/Cas13 has brought new dimensions to the field of disease diagnosis (Fig. 3). CRISPR/Cas toolkit reshape management of citrus greening disease through various strategies that precisely edit the host genome for improving resistance, rapidly detect the pathogen for early prevention and potentially use gene drive for insect population control[125]. The rapid, sensitive and specific nucleic acid detection is critical for agricultural applications, including trait detection, pest monitoring and pathogen identification during breeding process[126]. A modified version of the SHERLOCK platform has been used to rapidly quantify the level of glyphosate resistance genes in soybean mixtures and to detect multiple genes in a single reaction, which reveals its huge applications in detection and quantitation of genes in agriculture[127]. The geminiviruses, tomato leaf curl New Delhi virus (ToLCNDV, bipartite begomovirus) and tomato yellow leaf curl virus (TYLCV, monopartite begomovirus), can be both be detected in ~1 h by utilizing Cas12-mediated systems, making it suitable for point-of-use diagnostic applications[128]. By using the improvement of DNA endonuclease targeted CRISPR trans reporter (DETECTR), which is highly sensitive and specific, was used to detect potato witches' broom (PWB) and potato purple top (PPT) phytoplasmas[129]. The new strategy of these diseases-detection methods developed based on CRISPR/Cas technologies provides a practical and convenient approach for detecting early stage viral, bacterial and fungal infections in crop plants.
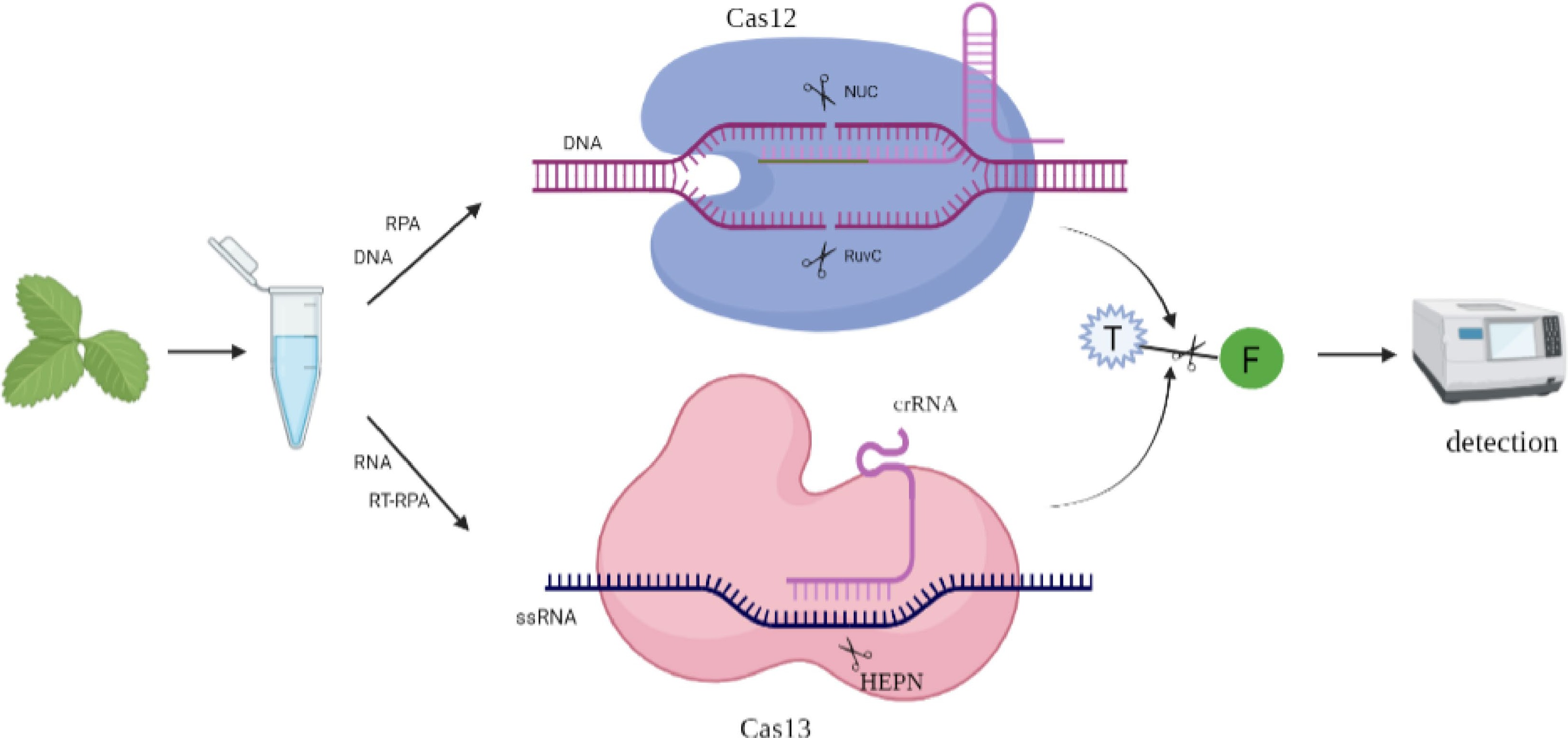
Figure 3.
Application of CRISPR/Cas12 and CRISPR/Cas13 in nucleic acid detection. Cas12 collateral cleaves a reporter nucleic acid, which is composed of a single stranded DNA (ssDNA) bearing a fluorophore, and the fluorophore is released and leads to the emission of a fluorescent signal. Cas13 : crRNA complexes activate and cleave fluorescent RNA sensors for RNA target detection.
-
Despite the latest technological advances in CRISPR/Cas in plant genome editing, it is still not perfect and may create undesired changes in a genome. Most obviously, off-target effects, massive DNA deletions and other rearrangements still exist in CRISPR/Cas-based editing of plants, and further research to address this propensity is required. Besides, unlike mammalian cells, the plant cell wall makes efficient delivery of CRISPR reagents to plant cells challenging. The current delivery systems, biolistic bombardment and Agrobacterium-mediated delivery, are highly dependent on plant species, genotypes, and tissue-type specificity[130]. Also, plant tissue culture and regeneration still limit the efficiency of obtaining edited plants, especially for those plants that have not yet established genetic transformation and regeneration system, which is a great challenge. Thus, how to create edited plants without going through plant tissue culture that sidesteps the tissue culture bottleneck will be a key direction of future research in plants. Another challenge for plant genome editing is to identify and understand gene function before editing. Genes for desirable traits in many plants have not been mapped clearly. Despite these limitations, CRISPR/Cas systems have great potential for improving plant design and synthetic biology. For example, artificial DNA sequences, including promoters, genes, transcriptional regulatory elements, and genome assemblies, can be inserted into plant genomes to alter cell or plant behavior to generate novel functions and products. Given that CRISPR/Cas systems have been expanding as a molecular manipulation toolbox, misuse of this toolbox could be a risk and danger, so a scientific hierarchical regulatory system might contribute to accommodate both existing and future technologies[131]. CRISPR/Cas, like polymerase chain reaction (PCR), is an epoch-making discovery in the field of life science that will be worth mastering by every researcher. Therefore, with continuous technological advancement and interdisciplinary applications of big data, CRISPR/Cas systems and their derivations will be the most widely used toolbox in plants in the near future and will revolutionize the future of agriculture.
-
The authors would like to thank Dr. Yifu Tian at Hainan Yazhou Bay Seed Laboratory, for his valuable comments and suggestions to improve the quality of the figures by using biorender (
https://biorender.com/ ) in this manuscript. The authors also thank Dr. Xinbo Li from Hainan Yazhou Bay Seed Laboratory for his good suggestions to improve the layout of this manuscript. -
The authors declare that they have no conflict of interest.
-
Received 16 November 2022; Accepted 26 January 2023; Published online 17 February 2023
-
# These authors contributed equally: Chao Dong, Yue Xi
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Dong C, Xi Y, Satheesh V, Lei M. 2023. Advances in CRISPR/Cas technologies and their application in plants. Tropical Plants 2:2 doi: 10.48130/TP-2023-0002 

Advances in CRISPR/Cas technologies and their application in plants
- Received: 16 November 2022
- Accepted: 26 January 2023
- Published online: 17 February 2023
Abstract: Discovery of the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-associated (CRISPR/Cas) system and its repurposing into a powerful genome editing tool has revolutionized genome engineering and generated excitement for innovative breeding technology. CRISPR/Cas can perform genetic operations such as targeted insertion, deletion and replacement of genes in plants by its simple two-component system comprising a Cas protein and a guide RNA. Here, we focus on the recent advances in CRISPR/Cas technologies that are available for genome editing, among which Cas9, Cas12, and Cas13 systems are widely used in the areas of botany and agriculture. We also describe the new high-precision genome editing tools, base editors and prime editors that are derived from the Cas9 system and beyond for altering the genome in living cells, without generating double-stranded breaks in DNA or requiring a donor. In addition, we summarize the differences between the different CRISPR/Cas systems and their broad applications in plants. We also discuss the challenges facing the use of CRISPR/Cas technologies and the future directions of CRISPR/Cas systems in plant genome editing.
-
Key words:
- CRISPR/Cas /
- Applications /
- Genome editing /
- Precision breeding