









-
Salt stress is considered a major abiotic factor that could limit plant productivity. It inhibits the normal growth and development of plants through osmotic stress, ion stress and other secondary stresses such as oxidative stress[1, 2]. Under abiotic stress, calcium acts as an important secondary messenger to transmit stress signals. Calcineurin B-like proteins (CBLs) are one of the main sensors of Ca2+, which cooperate with CBLs-interacting protein kinases (CIPKs) to induce cellular responses[3]. CBL-CIPK modules play central roles in Ca2+ conversion to physiological adaptations by phosphorylating downstream targets such as ion channels and transporter proteins to maintain ion balance. Moreover, the CBL-CIPK modules have been demonstrated to function in multiple abiotic stresses such as salinity, drought and disease[4].
CBLs were first identified in Arabidopsis thaliana and share high similarities to Calcineurin B (CNB) in animals and Neuronal Calcium Sensors (NCS) in yeast[5]. CBLs are characterized as four EF (elongation factor)-hand domains with constant spacing, containing proteins with an N-terminal cellular localization motif and a C-terminal phosphorylation motif[6, 7]. CIPK proteins consist of a Kinase catalytic domain at the N-terminus, a short variable junction domain and a regulatory domain at the C-terminus. The C-terminal regulatory domain contains a conserved protein phosphatase interaction (PPI) motif and a highly conserved FISL (NAF) motif, which are required for CBLs to activate the catalytic activity of CIPKs[8−11]. Genome-wide analyses have identified 10 CBLs and 26 CIPKs in Arabidopsis and 10 CBLs and 30 CIPKs in rice[12, 13]. The first identified CBL-CIPK signaling module was established in the Salt Overly Sensitive (SOS) signaling pathway. CBL4/SOS3 was found to interact with CIPK24/SOS2 to regulate SOS1, which encodes a plasma membrane-localized Na+/H+ antiporter protein by forming a protein complex[14−16]. The sos3/cbl4 mutant in Arabidopsis is highly sensitive to salt stress for the imbalance between Na+ and K+[17, 18]. CBL4/SOS3 also functions in auxin supply, lateral root primordia initiation and anthocyanin regulation[19, 20]
Cucumber (Cucumis sativus L.) is a major vegetable crop with important economic value and is especially vulnerable to high salt environments[21, 22]. Generally, cucumbers are produced under protected cultivation, which is more prone to secondary salt damage[23, 24]. Salinity stress has a significant effect on the yield and quality of cucumber fruit[25]. Using comparative genomic methods, six CBLs were identified in cucumber. However, no direct experiments were conducted to verify the functions of CBLs in cucumber[26]. Presently, molecular research on genes that function in salt stress in cucumber focuses mainly on transcription factors and oxygen-related proteins[27, 28].
Although CBL-CIPK signaling modules have been extensively studied and shown to play crucial roles in responses to various environmental stresses in Arabidopsis, little is known about their functions in cucumber. Therefore, we identified CsCBL4, encoding a Calcineurin B-like protein similar to AtCBL4, and found that CsCIPK6 interacted with CsCBL4. Silencing of CsCBL4 or CsCIPK6 in cucumber increased salt sensitivity, while the overexpression of CsCBL4 increased the salt tolerance of cbl4 mutant in Arabidopsis. Collectively, this study indicates that the CsCBL4-CsCIPK6 module plays a crucial role in the resistance to salt stress. Unraveling the CBL-CIPK signaling module in cucumber provides vital information for breeding cucumber with greater stress tolerance.
-
Studies have demonstrated that the Calcineurin B-like protein, CBL4, plays an important role in salt stress in Arabidopsis. To determine the function of CBL4 in cucumber, we performed a BLAST search in the cucumber genome (
http://cucurbitgenomics.org ) using the protein sequence of AtCBL4 (AT5G24270.1), obtained from the Arabidopsis genome (www.Arabidopsis.org ). The result showed that the protein encoded by CsaV3_3G019930 had the highest sequence similarity with AtCBL4. The full length of CsaV3_3G019930 (designated as CsCBL4) is 4,814 bp and includes 8 exons and 7 introns (Fig. 1a). Sequence analysis revealed that CsCBL4 encodes a protein of 212 amino acids with four EF-hand Ca2+-binding motifs similar to AtCBL4 (Fig. 1b). According to previous studies, 6 CBL genes were identified in cucumber. To determine the relationship between CBLs, a phylogenetic tree of 10 CBLs from Arabidopsis and 6 CBLs from cucumber was constructed using MEGA6. We classified the CBLs into three groups, with CsCBL4 (CsaV3_3G019930) sharing a close relationship with the AtCBL4 (AT5G24270) (Fig. 1c). Overall, structure and phylogenetic analysis indicated that CsCBL4 and AtCBL4 were highly homologous.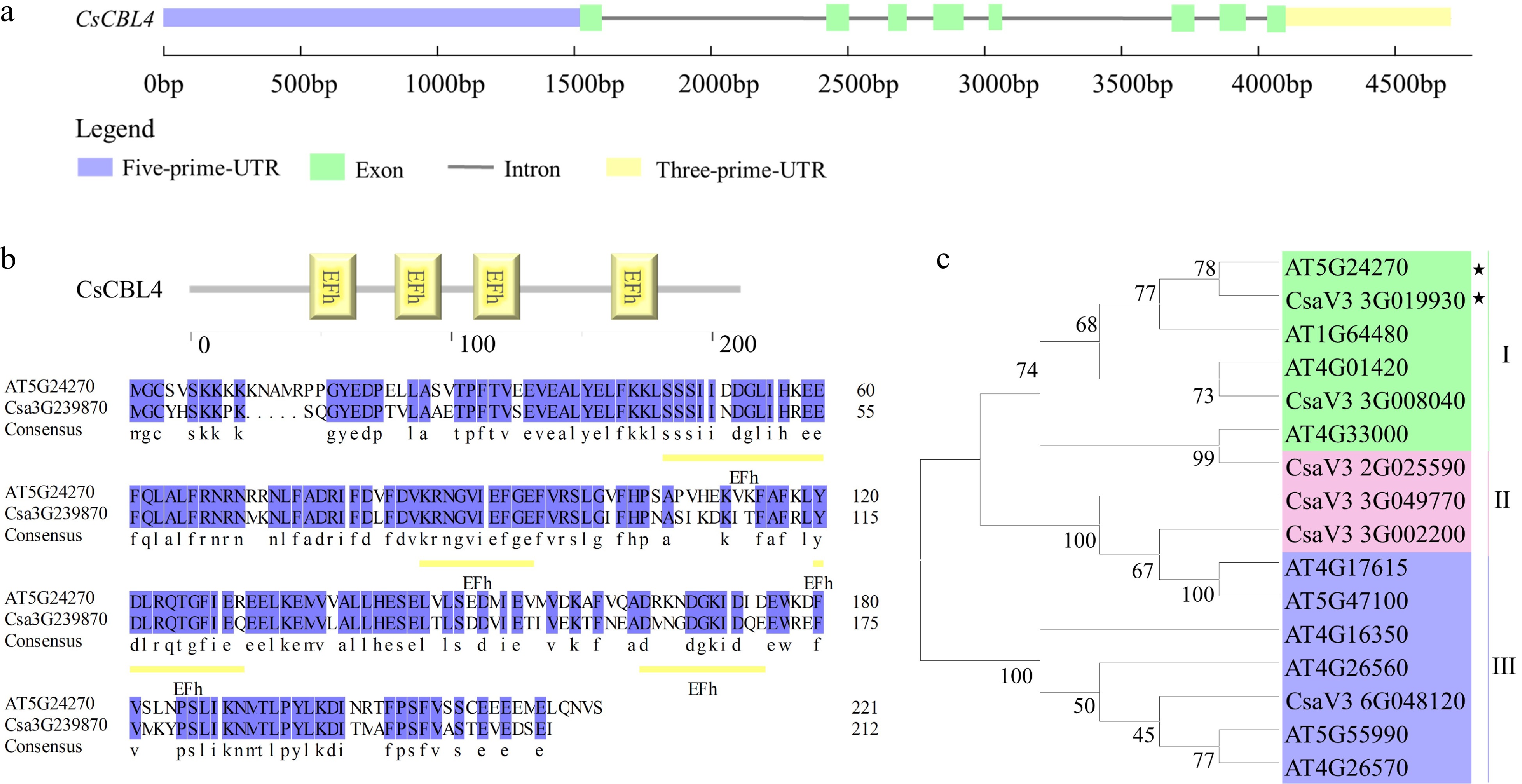
Figure 1.
Isolation and sequence analysis of CsCBL4. (a) The exon-intron structure of CsCBL4. (b) The EF-hand domains of CBL4 in Arabidopsis and cucumber. Yellow underlines indicated the position of domains. (c) The phylogenetic tree of CBLs in cucumber and Arabidopsis. * indicates CBL4 genes of Arabidopsis and cucumber respectively.
Expression analysis of CsCBL4
-
To investigate the function of CsCBL4 in salt stress, we performed quantitative real-time PCR (qRT-PCR) using root samples treated with different salt concentrations. qRT-PCR results show that the expression of CsCBL4 was induced by salt stress (Fig. 2a). To examine the spatial expression patterns of CsCBL4, we performed qRT-PCR analyses using various cucumber tissues. The results indicate that expression levels were highest in male buds, female buds and roots (Fig. 2b), which confirms that CsCBL4 has specific expression patterns in different tissues. The function of genes is closely related to the subcellular localization of their proteins. Therefore, we fused the full-length coding sequence of CsCBL4 without the stop codon to green fluorescent protein (GFP) and expressed it in mesophyll cells of N. benthamiana. The green fluorescent signal emitted by the CsCBL4-GFP fusion protein was observed on the membrane of mesophyll cells, while the empty pSuper-1300 vector was used as a control (Fig. 2c). These results suggested that CsCBL4 might be related to salt tolerance through activating ion transporters on membranes in cucumber roots.
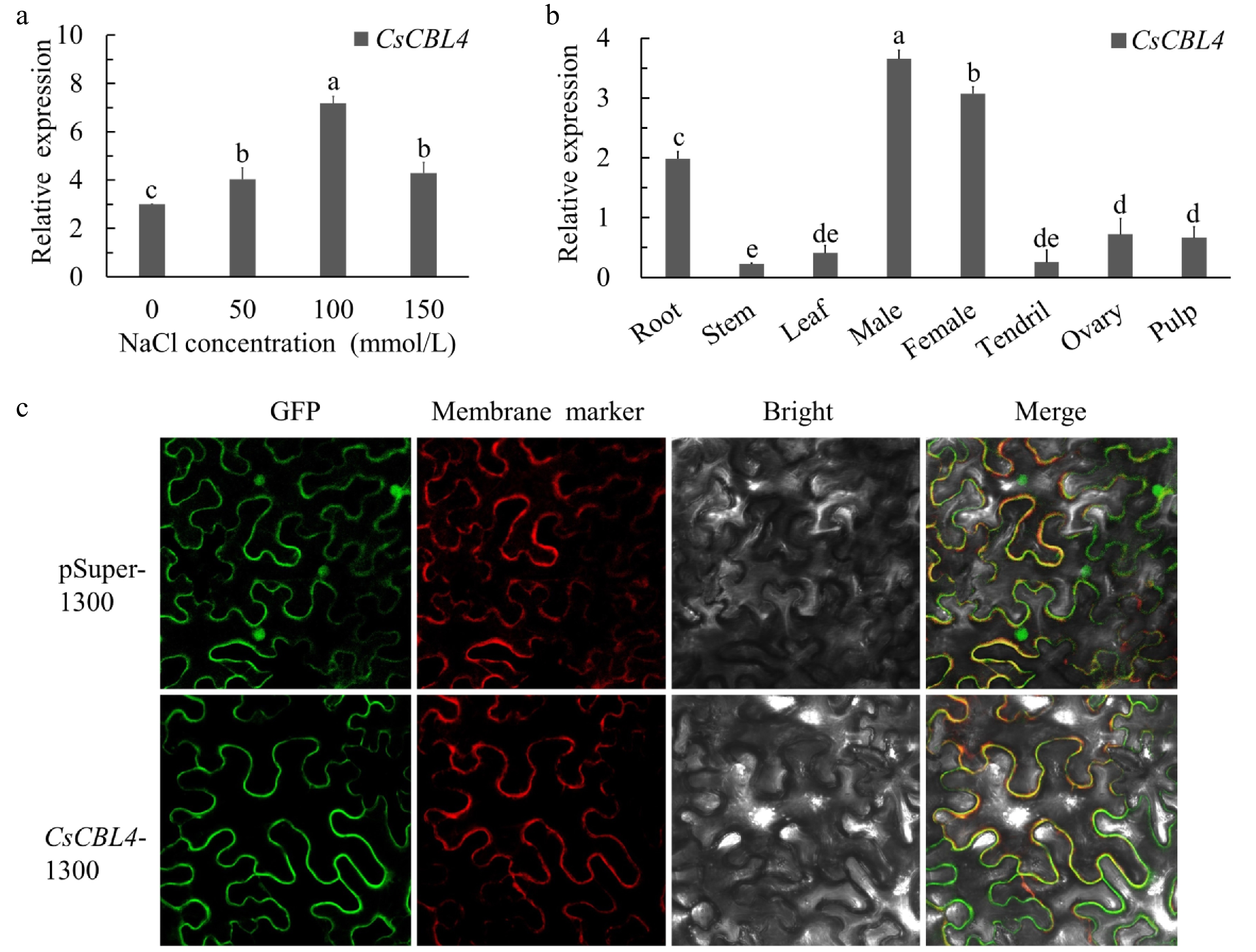
Figure 2.
The relative expression levels and subcellular localization of CsCBL4. (a) The expression levels of CsCBL4 under different salt treatments. (b) The relative expression levels of CsCBL4 in different tissues (root, stem, leaf, male bud, female bud, tendril, ovary at the first day of flowering and pulp at 7 days after flowering) of cucumber. (c) Subcellular localization of the CsCBL4-GFP fusion protein in N. benthamiana. Each value is the mean SE (n = 3). Different icons indicate significant differences between treatments (p < 0.05).
CsCBL4 physically interacts with CsCIPK6
-
To elucidate the regulatory mechanism of CsCBL4 in response to salt stress, we first identified interacting proteins by performing a DUAL membrane system screen. Since the pTSU2-APP is well expressed and interacts strongly with the pNubG-Fe65, the positive control transformation grew robustly under selection conditions, while the negative control transformation with pTSU2-APP and pPR3-N yielded considerably fewer colonies. The reaction results confirmed that the bait was functional in the DUAL membrane assay. Also, we found the optimal concentration of 3-AT(3-Amino-1,2,4-triazole) to optimize the basic screening conditions in the pilot screen (Fig. 3a & b). Based on the collation of the screening results, we discovered that the protein encoded by CsaV3_2G003670 was a putative interacting protein of CsCBL4 (Supplemental Table S2). Sequence alignment found that it has the highest similarity with AtCIK6. To confirm the interaction between CsCBL4 and CsCIPK6, we performed a yeast two-hybrid (Y2H) analysis. Y2H results indicate that CsCIPK6 interacts with CsCBL4 (Fig. 3c). In addition, the results of the LCI assay showed that the luminescent signals were strongly generated by co-expression of CsCBL4 and CsCIPK6, while no luminescent signals appeared in the control (Fig. 3d). Altogether, these results proved that CsCBL4 physically interacted with CsCIPK6.
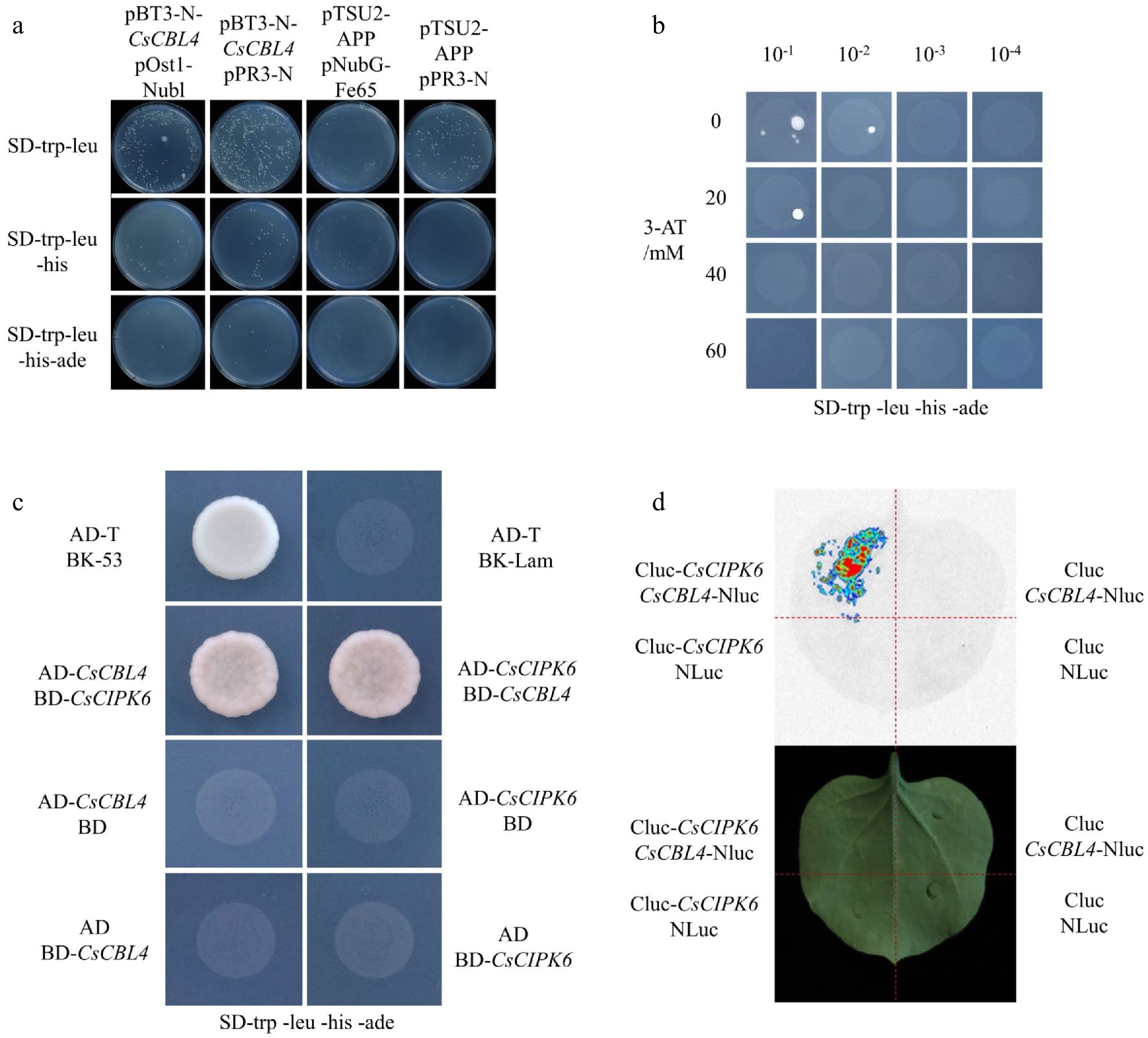
Figure 3.
CsCBL4 interacted with CsCIPK6. (a) The transformation efficiency of the DUAL membrane assay. (b) The background concentration of 3-AT to inhibit the self-activation of pBT3-N-CsCBL4. (c) The interaction between CsCBL4 and CsCIPK6 indicated by Y2H. (d) The interaction between CsCBL4 and CsCIPK6 in vivo showed by LCI assay.
Previous studies revealed that AtCIPK6 was involved in salt stress, the perception of pathogen-associated microbial patterns (PAMPs) and the regulation of auxin and Abscisic Acid (ABA)[29−31]. CBL-CIPK interactions are believe to play a role in response to salt stress. A single CIPK can interact with several CBLs. In Arabidopsis, CIPK6 can interact with CBL2, CBL4 and CBL10[32−34]. Previous studies have confirmed that CsCBL4 interacts with CsCIPK6, however, it is unknown whether CsCIPK6 responds to salt stress in cucumber.
Expression analysis provides evidence for the interaction between CsCBL4 and CsCIPK6
-
The full length of CsaV3_2G003670 (designated as CsCIPK6) is 1857bp and includes one exon without an intron (Fig. 4a). Sequence analysis indicated that CsCIPK6 encodes a protein of 433 amino acids in length with a highly conserved NAF domain at the C-terminus required for interaction with CBL proteins (Fig. 4b). A BLAST search for CIPKs identified 20 CIPKs in the cucumber genome (
http://cucurbitgenomics.org ). A phylogenetic tree including 20 CIPKs in cucumber and 26 CIPKs in Arabidopsis was constructed using MEGA6. As expected, CsCIPK6 shares a close relationship with AtCIPK6 (Fig. 4c). Collectively, these results indicated the CsCIPK6 and AtCIPK6 were highly homologous.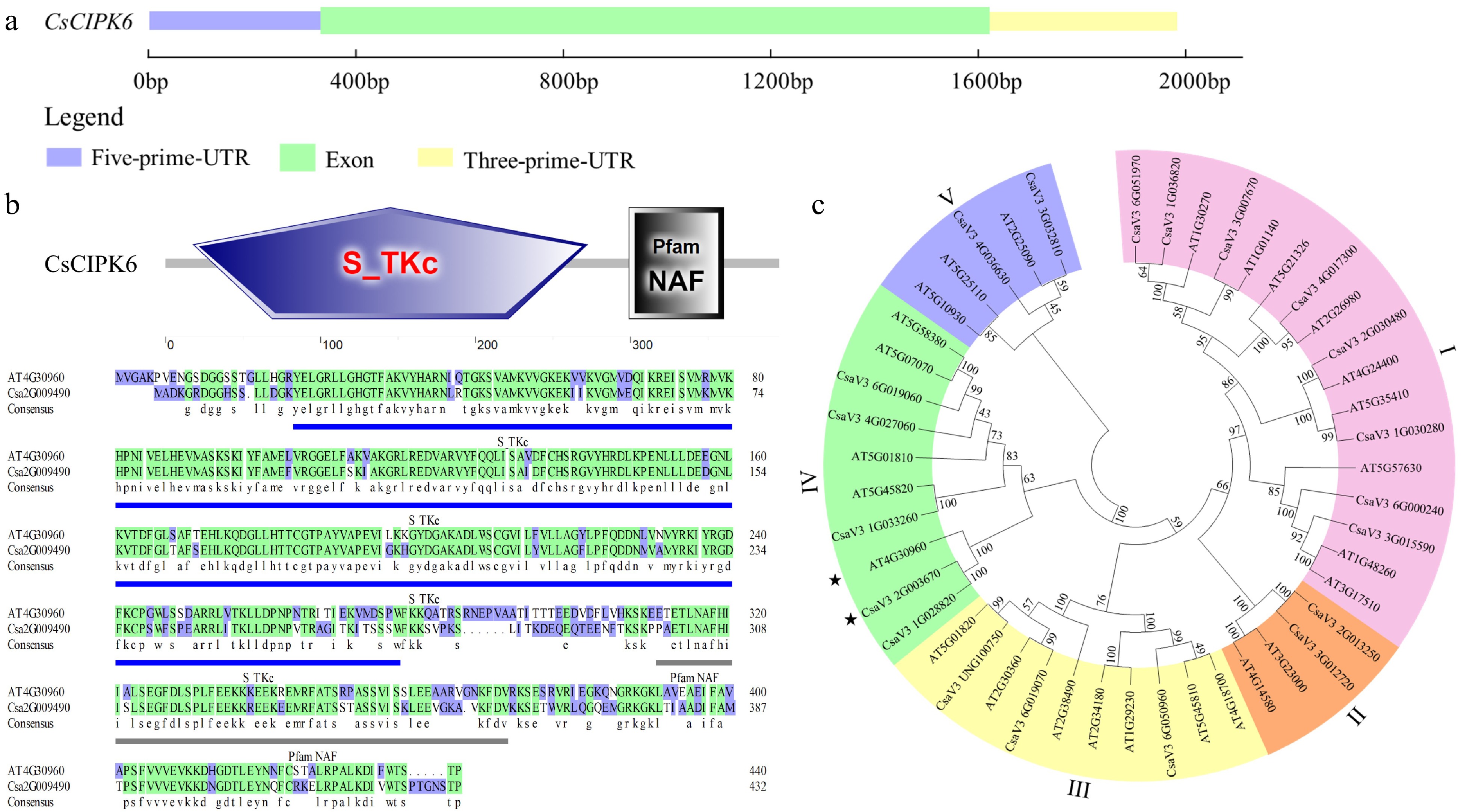
Figure 4.
Phylogenetic tree and conservative motifs analysis of CsCIPK6. (a) The exon-intron structure of CsCIPK6. (b) The domains of CsCIPK6 in Arabidopsis and cucumber. Blue underlines indicate the position of the Pfam NAF domain. (c) The phylogenetic tree of CIPKs in cucumber and Arabidopsis. * indicates the CIPK6 genes of Arabidopsis and cucumber respectively.
To determine if CsCIPK6 responds to salt stress in cucumber, we conducted qRT-PCR using root samples treated with different salt concentrations. The results suggested that CsCIPK6 expression could be induced by salt stress (Fig. 5a). To determine the spatial expression patterns of CsCIPK6, we performed qRT-PCR analyses on different cucumber tissues. The results showed that the expression level of CsCIPK6 was highest in female buds, pulps (7 days after flowering), stems and roots (Fig. 5b). Subcellular localization in mesophyll cells of N. benthamiana revealed that CsCIPK6 localizes the nucleus and membrane (Fig. 5c). The results of both qRT-PCR and subcellular localization of CsCIPK6 were different from that of CsCBL4. However, both CsCBL4 and CsCIPK6 were highly expressed in the roots and localized in the membrane. These analyses provide evidence for the CsCBL4-CsCIPK6 interaction.
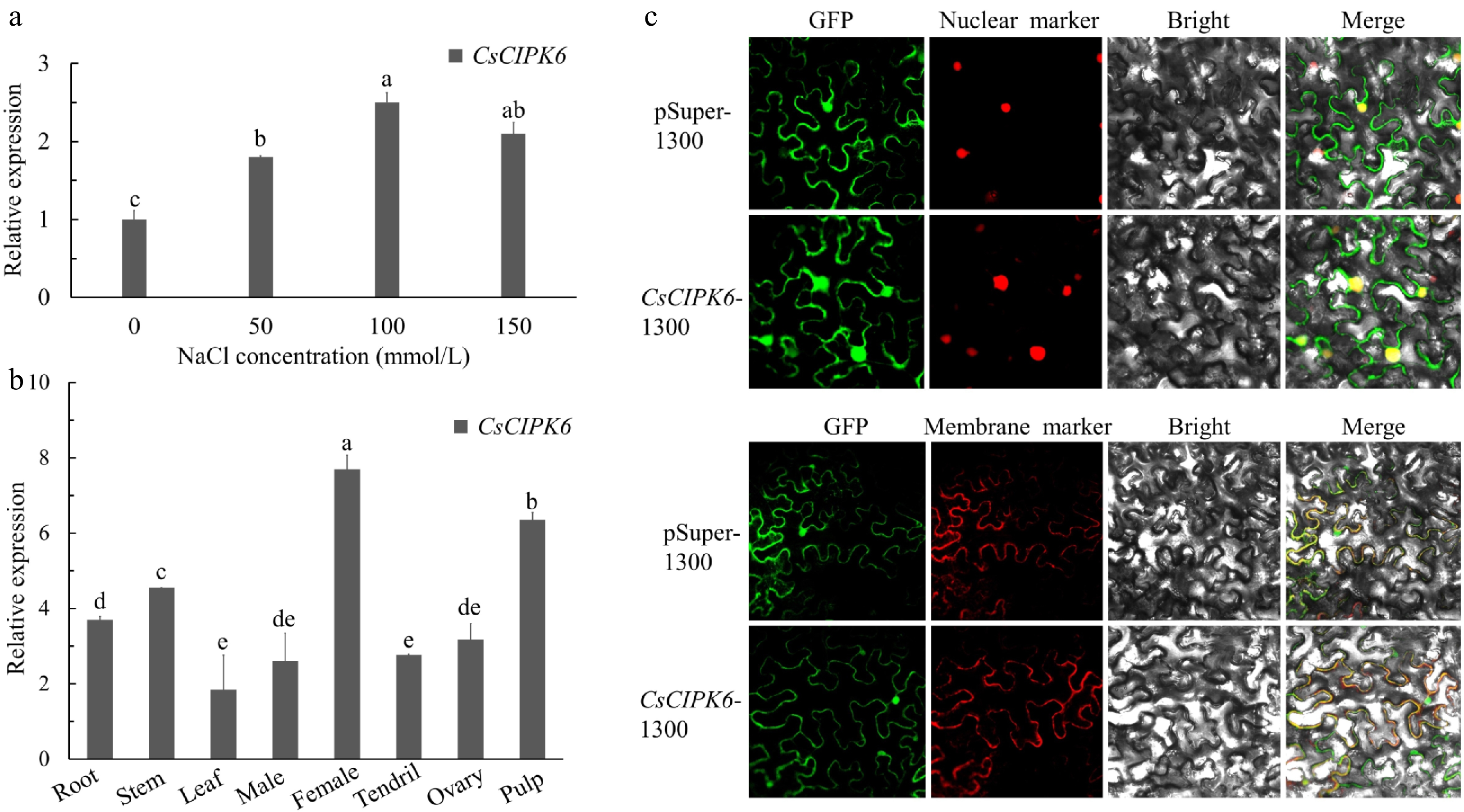
Figure 5.
Relative expression levels and subcellular localization of CsCIPK6. (a) The expression levels of CsCIPK6 under different salt treatments. (b) The relative expression levels of CsCIPK6 in different tissues (root, stem, leaf, male bud, female bud, tendril, ovary at the first day of flowering and pulp at 7 days after flowering) of cucumber. (c) Subcellular localization of the CsCIPK6 -GFP fusion protein in N.benthamiana. Each value is the mean SE (n = 3). Different icons indicate significant differences between treatments (p < 0.05).
Silencing of CsCBL4 or CsCIPK6 greatly reduce salt tolerance
-
To elucidate the biological functions of CsCBL4 and CsCIPK6 in cucumber, we used the tobacco ringspot virus (TRSV)-based virus-induced gene silencing (VIGS) system mediated by Agrobacterium to silence CsCBL4 and CsCIPK6. TRSV2-CsPDS (the phytoene desaturase gene) and empty vector TRSV2 were used as positive and negative controls, respectively[35]. Two weeks after Agrobacterium-mediated injection, cucumber leaves began to display the albino phenotype of the positive control. When the positive control plants showed the albino phenotype, the TRSV2-CsCBL4, TRSV2-CsCIPK6 and TRSV2 plants were treated with 0, 50, 100 and 150 mmol/L NaCl solutions for 3 weeks. Under normal conditions, the CsCBL4-silenced plants were significantly smaller or grew worse than the negative control plants, and the poor growth persisted with increasing salt concentrations. Whereas, CsCIPK6-silenced plants grew better than CsCBL4-silenced plants but worse than the control plants (Fig. 6a). Under salt stress, the leaf width (LW), root length (RL) and dry weight (DW) of CsCBL4-silenced plants were significantly lower than those of control plants. Furthermore, CsCBL4-silenced plants suffered more damage under the low NaCl concentration (50 mmol/L). The LW, RL and DW of CsCIPK6-silenced plants were similar to those of CsCBL4-silenced plants (Fig. 6b−d). We also calculated the ratio of phenotype (LW, RL and DW) values at 50, 100 and 150 mmol/L NaCl concentrations to those at 0 mmol/L. The results showed that the DW changes of CsCBL4-silenced plants and CsCIPK6-silenced plants were the most pronounced. However, CsCBL4-silenced plants were more sensitive to salt stress and exhibited reduced LW under 150 mmol/L NaCl concentration (Fig. 6b & d). The expression levels of CsCBL4 and CsCIPK6 were also significantly lower in the silenced plants (TRSV2-CsCBL4, TRSV2-CsCIPK6) (Fig. 7a). Salt stress is known to induce the accumulation of ROS (Reactive Oxygen Species) which leads to oxidative damage[36]. To determine the effect of salt stress on ROS accumulation, we detected total O2− content in the leaves of TRSV2, TRSV2-CsCBL4 and TRSV2-CsCIPK6 plants using NBT (Nitro Blue Tetrazolium). The results showed that the total content of O2− in TRSV2-CsCBL4 or TRSV2-CsCIPK6 plants were higher than those in TRSV2 plants. The quantitative measurement of ROS accumulation suggests that silencing of CsCBL4 or CsCIPK6 increased salt sensitivity in cucumber (Fig. 7b & c). These results indicate that the CsCBL4-CsCIPK6 network responded to salt stress synergistically.
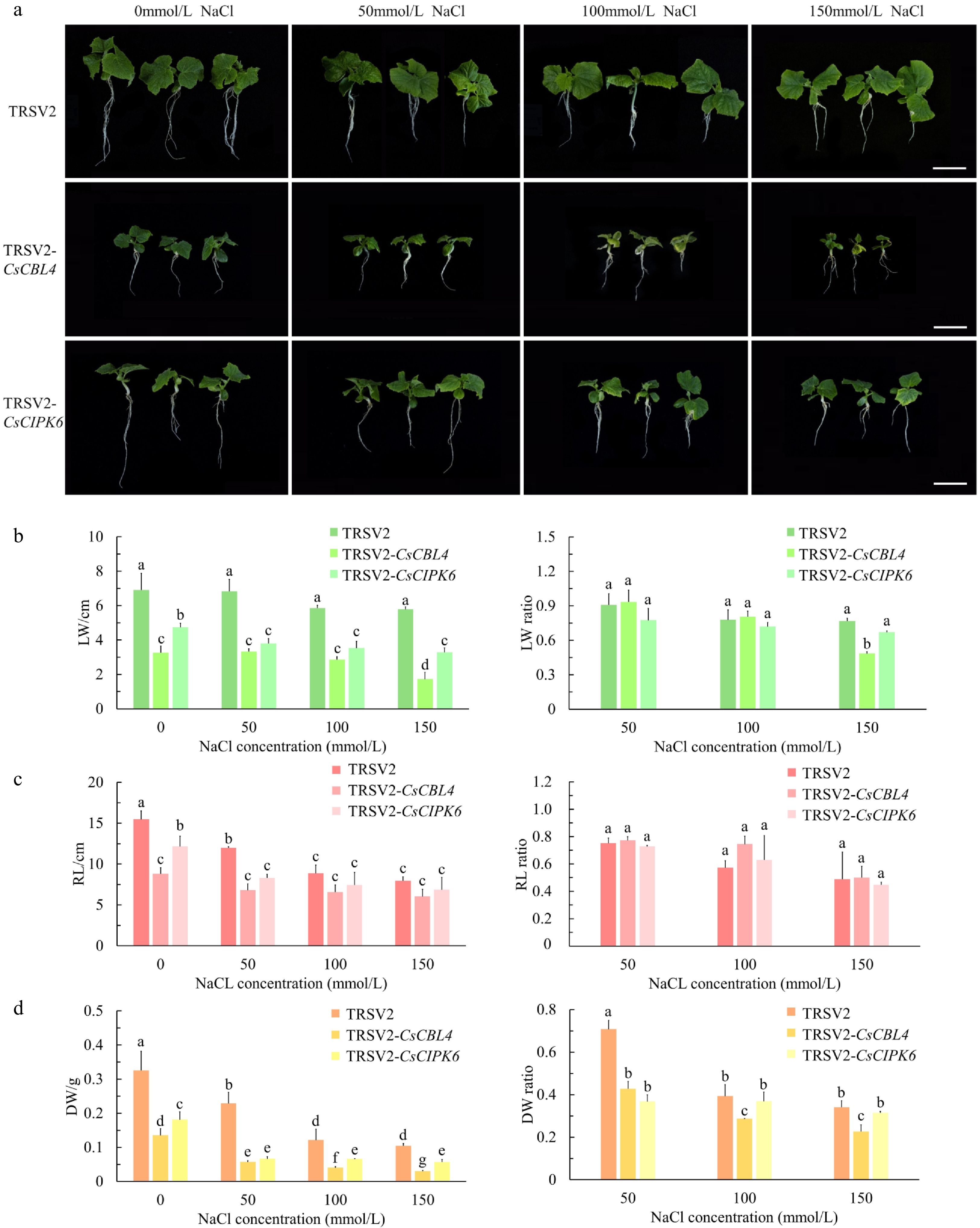
Figure 6.
Silencing of CsCBL4 or CsCIPK6 both reduced the salt tolerance in cucumber seedlings. (a) Phenotype of the silenced plants. Empty TRSV2 was used as a control. (Bar = 5 cm). (b) The leaf width (LW) of the control and the silenced plants in different treatments, and the ratios of plants with salt treatment to plants without salt treatment. (c) The root length (RL) of the control and the silenced plants in different treatments, and the ratios of plants with salt treatment to plants without salt treatment. (d) The dry weight (DW) of the control and the silenced plants in different treatments, and the ratios of plants with salt treatment to plants without salt treatment. Each value is the mean SE (n = 3). Different icons indicate significant differences between treatments (p < 0.05).
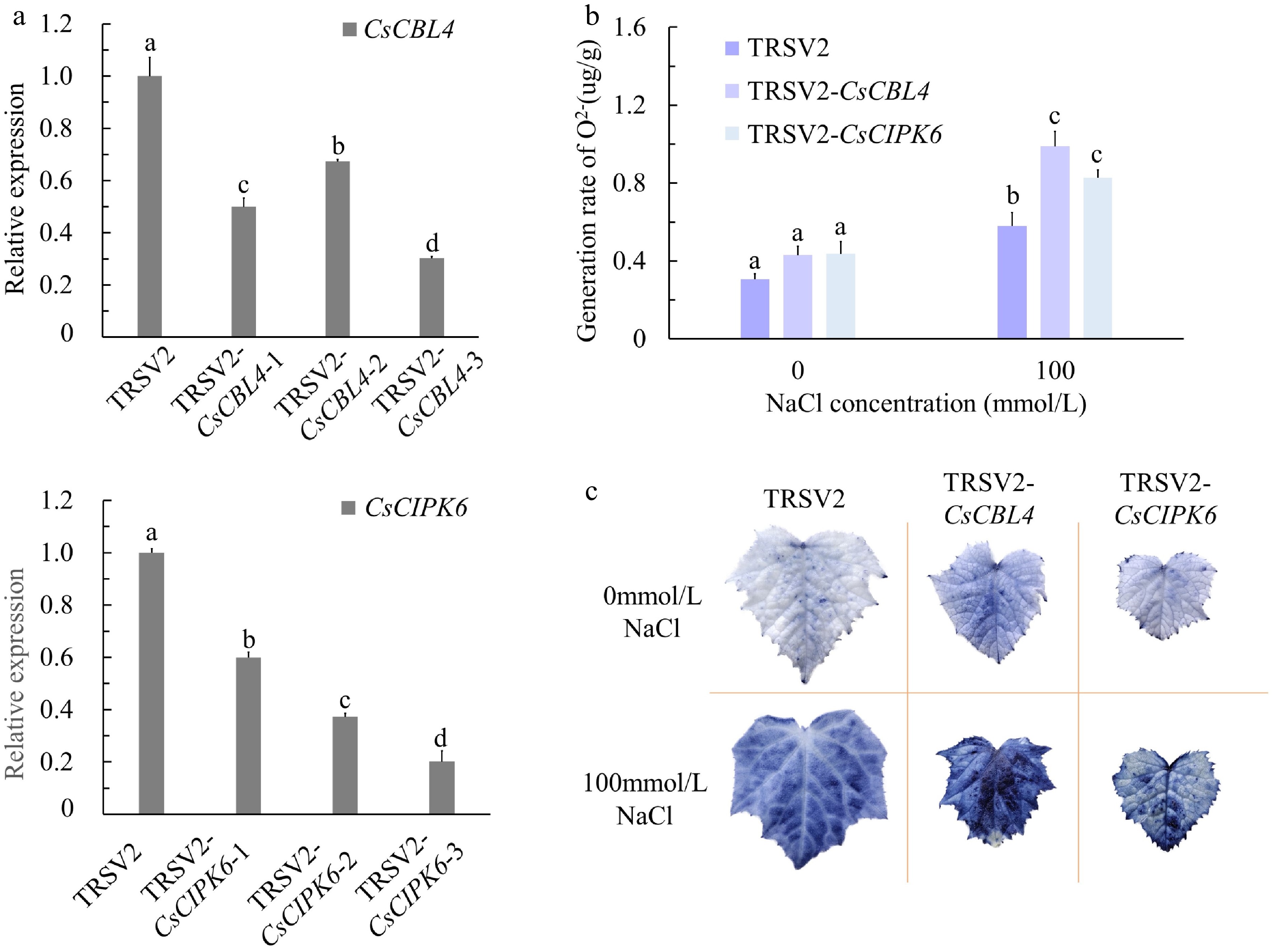
Figure 7.
The silenced degrees and oxygen damage of CsCBL4-silenced plants and CsCIPK6-silenced plants. (a) Relative expression levels of CsCBL4 and CsCIPK6 in roots of CsCBL4 and CsCIPK6 silenced plants. (b) The content of O2− of control and the silenced plants in different treatments. (c) The generation rate of O2− of the control and the silenced plants in different treatments. Each value is the mean SE (n = 3). Different icons indicate significant differences between treatments (p < 0.05).
CsCBL4 overexpression complements the salt sensitive phenotype of cbl4 (gl1) in Arabidopsis
-
To further explore the function of CsCBL4, the full-length coding sequence of CsCBL4 was used for overexpression experiments in Arabidopsis. The cbl4 mutant was obtained in the background of the gl1 mutant, which we referred to as the wild type (WT). WT (gl1), cbl4 (gl1) and 35S-CsCBL4 (cbl4) seeds were grown on 1/2 MS medium without NaCl (the control group) and 1/2 MS medium with 100 mmol/L NaCl (the experimental group). The germination rates were calculated after 5 days at 25 °C. In the experimental group, the germination rate of 35S-CsCBL4 (cbl4) plants was slightly lower than that of WT (gl1) plants but significantly higher than that of cbl4 (gl1) plants (Fig. 8a & b). The germinated seeds were then transferred to the 1/2 MS medium without NaCl and 1/2 MS medium with 100 mmol/L NaCl. After vertical placement for 5 days, there was no difference in the root elongation phenotype. However, under salt stress, the root lengths of cbl4 (gl1) plants were significantly shorter than that of WT (gl1) and 35S-CsCBL4 (cbl4) plants (Fig. 8c & d). Arabidopsis plants were then grown in soil and no obvious differences between the lines were observed under the normal growth conditions. However, when treated with salt (200 mmol/L NaCl) for 2 weeks, the growth of cbl4 (gl1) plants was inhibited, and the anthocyanin accumulation increased in their leaves (Fig. 8e). Furthermore, the dry weights of WT (gl1) and 35S-CsCBL4 (cbl4) plants were more than that of cbl4 (gl1) plants. These results indicate that overexpression of CsCBL4 in cbl4 (gl1) improved the salt tolerance of Arabidopsis.
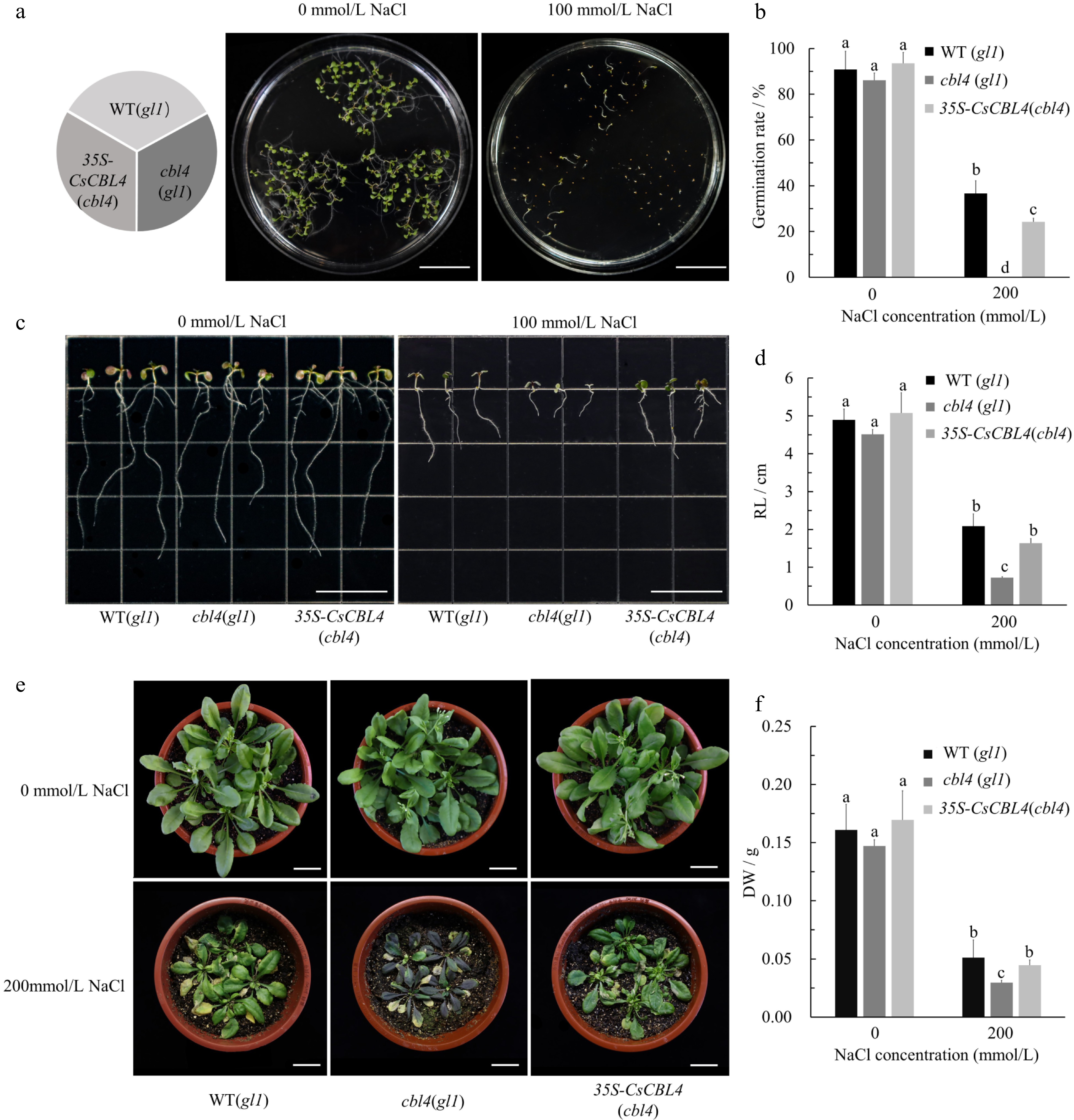
Figure 8.
Overexpression of CsCBL4 could improve the salt tolerance of cbl4(gl1) in Arabidopsis. (a) The germination conditions of WT (gl1), cbl4 (gl1) and 35S-CsCBL4 (cbl4) on the mediums with different NaCl concentrations (Bar = 2 cm). (b) Statistics of germination in different mediums. (c) The root elongation of WT (gl1), cbl4 (gl1) and 35S-CsCBL4 (cbl4) plants on the mediums with different NaCl concentrations (Bar = 2 cm). (d) Statistics of root length on different mediums. (e) Growth of WT (gl1), cbl4 (gl1) and 35S-CsCBL4 (cbl4) plants treated with different salt concentrations (Bar = 2 cm). (f) The dry weight of rosettes and roots in WT (gl1), cbl4 (gl1) and 35S-CsCBL4 (cbl4) under different salt treatments. Each value is the mean SE (n = 3). Different icons indicate significant differences between treatments (p < 0.05).05).
-
Previous studies revealed that CBL-CIPK modules play important roles in salt stress. CBL4-CIPK6 complexes could mediate the Ca2+ signal and activate AKT2 K+ channels by phosphorylation[32]. cbl4 mutants are specifically sensitive to salt stress. Under salt stress conditions, the primordia of lateral roots and the auxin transport in cbl4 mutant are significantly reduced. A decrease in auxin polar transport in the cbl4 mutant could result in less auxin supply, which causes defections of the lateral roots and cell divisions. The cipk6 mutant exhibits developmental damage, such as swollen hypocotyls and compromised lateral roots[19]. Furthermore, several genes involved in auxin transport and the responses to abiotic stress are expressed lower in the mutant plants[37]. Our research shows that the CsCBL4-CsCIPK6 network modulates salt tolerance and provides new evidence for the conservative function of CBLs and CIPKs in cucumber. However, we did not study their functions in lateral root development and auxin transport. Indeed, the number of lateral roots in CsCBL4- and CsCIPK6-silenced plants decreased significantly, but this data is not shown in this paper. In the future, we plan to investigate the molecular basis of CsCBL4-CsCIPK6 in regulating lateral root and auxin transport.
In this study, a DUAL membrane system was used to screen for CsCIPK6. Additionally, proteins that can interact with CsCBL4 were screened (Supplemental Table S2). After screening and sequencing, we identified 114 genes related to ROS (reactive oxygen species), chlorophyll, lateral roots and plant hormones. However, the interactions between the screened genes and CsCBL4 were not determined. To further explore the mechanism of salt resistance in cucumber, the interactions and functions of the screened genes could be determined.
Salt stress is known to have a significant effect on the development of cucumber. Currently, cucumber CBL and CIPK genes have been identified, but the interactions between those genes are unclear. In Arabidopsis, numerous advancements have been made in elucidating the functions of CBL-CIPK modules, which are central regulatory networks that decode Ca2+ signals of stresses[38]. It will be interesting to determine if other CBL-CIPK modules exist in cucumber and their possible function in abiotic stresses. Furthermore, CBL-CIPK modules have been shown to interact with a phosphatases protein, a 14-3-3 protein and a chaperone-like protein DNAJ in Arabidopsis[39−41]. To explore the regulatory mechanisms of the CsCBL4-CsCIPK6 complex, it will be useful to understand how the CsCBL4-CsCIPK6 complex obtains upstream Ca2+ signals and activates downstream target proteins. Furthermore, the characterization of additional phosphorylation targets of CsCIPK6 will be an important step in understanding the functions of CsCBL4 in salt stress.
-
The present study indicates that the CsCBL4-CsCIPK6 module had a positive effect on salt stress tolerance in cucumber. CsCBL4 interacts with CsCIPK6 to enhance salt tolerance in response to Ca2+ signals. Silencing of CsCBL4 or CsCIPK6 significantly affected the growth of cucumber plants. These results provide a better understanding of the molecular mechanism regulated by the CBL-CIPK network in cucumber.
-
Cucumber plants used in this study were cv. XinTaiMiCi. Seeds were surface sterilized and then plated on MS (Murashige and Skoog) medium (pH 5.6−5.8) containing 0.2% phytagel (Gellan Gun). After stratification at 28 °C for 5 days, the seedlings were potted in soil and placed in an environment‐controlled growth chamber with long-day conditions.
RNA extraction and qRT-PCR
-
Total RNA was extracted from various tissues of WT and transgenic plants using the Huayueyang RNA extraction kit (Huayueyang, P. R. China), and then 2 ug of total RNA extracted was reverse transcribed using PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Japan) following the manufacturer’s protocol. qRT-PCR was conducted in 96-well plates with an Applied Biosystems 7500 real-time PCR system (Applied Biosystems, USA) using SYBR Premix Ex Taq (TaKaRa, Japan). The α-TUBULIN gene (Supplemental Table S1) served as the internal control gene[42].
DUAL membrane system screen
-
The fundamental requirement of the DUAL membrane system is the Cub-LexA-VP16 module of the bait and the NubG module of the prey located in the cytosol. For this reason, the pBT3-N was chosen as the DUAL membrane starter kit. After constructing the bait, it was transformed into the reporter strain NMY51 at the optimal 3-AT concentration of 40 mM/L to restrain background growth on SD-trp-leu-his-ade. Transformants appear after 3−4 days after screening against a NubG-fused cDNA library.
Yeast two-hybrid assays
-
The full-length CDS of CsCBL4 was cloned into the pGADT7 vector and the full-length CDS of CsCIPK6 was cloned into the pGBKT7 vector using primers listed in Supplemental Table S1. The CsCBL4-pGADT7 and CsCIPK6-pGBKT7 constructs were transferred into Y2HGold chemically competent cell simultaneously. Transformants were selected on SD-trp-leu plates and tested for growth on SD-trp-leu-his-ade plates at 30 °C to identify protein−protein interaction.
Luciferase complementation imaging
-
The full-length CDS of CsCBL4 was cloned into the pCAMBIAI1300-cLUC (cLUC) vector to generate the CsCBL4-cLUC construct, and the full-length CDS of CsCIPK6 was cloned into the pCAMBIA1300-nLUC (nLUC) vector to generate the nLUC-CsCIPK6 construct using primers listed in Supplemental Table S1. Next, 1 ml samples of GV3101 cells harboring nLUC-CsCIPK6 and CsCBL4-cLUC were mixed equally for transient expression in N. benthamiana. After 48 h in the dark, the signals were detected in plant leaves sprayed fluorescein by CCD (Charge Coupled Device) imaging system.
Subcellular localization
-
The full-length coding sequence of the target gene excluding the stop codon was cloned into the vector pSuper-1300 to form the fusion protein. All recombinant vectors were confirmed by sequencing and transformed into GV3101. For transient expression in N. benthamiana mesophyll cells, the mixed liquid concluding target vector, marker vector and P19 were infiltrated into leaves. After 1 day in the dark and 2 days in the light, fluorescence was observed with a confocal microscope.
VIGS
-
TRSV-based VIGS was used to analyze the potential roles of genes in cucumber[43]. The unique 300-500 bp CDS sequences of each target gene (Supplemental Table S1) were inserted into pTRSV2, and then the constructs were transformed into GV3101. Four milliliter pTRSV2 and 4 ml pTRSV1 were mixed equally to infect cucumber seeds with tiny root hairs. The seeds were put on 1/2 MS solid medium with 100 uM acetosyringone for 5 days. They were then potted in soil and placed in a growth chamber.
Overexpression
-
The full-length coding sequence of the target gene was cloned into the vector pSuper-1300, and the recombinant vector was confirmed by sequencing. After confirmation by sequencing, the recombinant vector was transformed into A. tumefaciens strain GV3101. Arabidopsis was infected by the dipping method and the seeds of the T0 generation were selected by hygromycin. The seeds of the T1 generation were treated with salt stress to observe the salt sensitivity of the overexpressed plants.
Salt stress treatment
-
Cucumber phytoene desaturase (TRSV2-CsPDS) was used as a marker for VIGS. When the PDS plants began to exhibit an albino phenotype, the salt tolerances of VIGS-silenced cucumber plants were determined by treating soil with NaCl solutions at various concentrations (0, 50, 100 and 150 mM) for 3 weeks.
Reaction oxygen species (ROS) analysis
-
For ROS analysis, 1 g of leaf tissue was quickly triturated with 5 ml of 50 mM phosphate buffer (pH 5.8), diluted to 10 ml and then incubated at 4 °C for 15 min. After incubation, 0.5 ml of the supernatant was mixed with an equal volume of phosphate buffer and 1 ml of 1 mM hydroxylamine hydrochloride. After standing at 25 °C for 1 h, 1 ml of 17 mM aminobenzene sulfonic acid and 1 ml 7 mM α-naphthylamine were added to the mix. The absorbance was determined at 530 nm for 20 min.
This research was supported by the Hainan Provincial Joint project of Sanya Yazhou Bay science and technology city (320LH042); Sanya institute of China Agricultural University (SYND-2021-18; SYND-2022-20); and the Construction of Beijing Science and Technology Innovation and Service Capacity in Top Subjects (CEFF-PXM2019_014207_000032). The authors are grateful to members of Ren’s lab for technical assistance and discussions.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The primers involved in the study process
- Supplemental Table S2 Some possible interaction proteins screened by DUAL membrane
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang M, Yang S, Sun L, Feng Z, Gao Y, et al. 2022. A CBL4-CIPK6 module confers salt tolerance in cucumber. Vegetable Research 2:7 doi: 10.48130/VR-2022-0007 

A CBL4-CIPK6 module confers salt tolerance in cucumber
- Received: 06 February 2022
- Accepted: 27 May 2022
- Published online: 27 June 2022
Abstract: Soil salinization is a major threat to cucumbers grown under protected cultivation. Under stressful environments, calcineurin B-like proteins (CBLs) can sense and bind Ca2+ signals and regulate CBL-interacting protein kinases (CIPKs) to transmit signals and induce cellular responses. Although CBL-CIPK modules play central roles in plant development and response to various abiotic stresses in Arabidopsis, little is known about their functions in cucumber. In this study, we demonstrate that CsCBL4 interacts with CsCIPK6, which exhibited similar responses to salt stress in cucumber. Furthermore, salt stress resulted in greater accumulation of CsCBL4 and CsCIPK6. Comprehensive phenotype analysis demonstrated that silencing CsCBL4 or CsCIPK6 reduced the salt tolerance of cucumber, and overexpression of CsCBL4 increased the salt tolerance of Arabidopsis. Collectively, these results indicate that the CsCBL4-CsCIPK6 module plays an important role in the resistance of cucumber to salt stress. The information provides insights for the genetic breeding of salt tolerance in cucumber in the future.
-
Key words:
- Cucumber /
- CBL4 /
- CIPK6 /
- Salt tolerance