










-
Tea anthracnose, which mainly occurs in Japan, China, and Sri Lanka, is an economically devastating tea disease that is prevalent in warm and humid conditions and severely affects tea yields[1−3]. The disease is widespread in all tea-producing provinces in China, especially in the Jiangnan tea region, wherein it is severe and ranks as the top among the three major tea leaf diseases such as tea anthracnose, brown blight, and grey blight[1,4].
Previous studies have identified 23 pathogens responsible for tea anthracnose[5,6], which can be broadly categorized into two groups: Discula theae-sinensis and fungi from the genus Colletotrichum (Colletotrichum spp.), with C. camelliae being the predominant species. Discula theae-sinensis was first discovered by Miyake in Japan in 1907 and initially named Gloeosporium theae-sinensis Miyake[7]. After two revisions, it was officially named D. theae-sinensis in 2009[8,9]. In the past decade, Colletotrichum spp. has garnered increased attention from researchers. Liu et al.[10] isolated 11 species of Colletotrichum from tea plants and proposed that C. camelliae is the dominant pathogen of tea trees in China. Since then, researchers have regarded Colletotrichum spp. as the primary pathogens of tea anthracnose and have conducted extensive studies on them[11−16].
Although researchers have increasingly regarded Colletotrichum spp. as the primary pathogens of tea anthracnose, there is still controversy over whether they cause tea anthracnose. Firstly, the most studied C. camelliae was initially reported as the pathogen of brown blight rather than tea anthracnose in tea plants[1,17]. In addition, some researchers have also noticed that the necrotic lesions caused by Colletotrichum spp. in some experiments do not match those of tea anthracnose[18]. Secondly, during the name revision process, D. theae-sinensis was once classified under the genus Colletotrichum[19], which may have misled later researchers. Additionally, in recent years, the majority of the studies related to Colletotrichum spp. have used tissue isolation methods to isolate pathogens. Although this method is easy to perform, it can easily lead to misidentification without careful confirmation of the pathogenicity of the isolated pathogens. Therefore, it is necessary to re-confirm the pathogens of tea anthracnose.
To identify the pathogen causing tea anthracnose, we conducted a detailed study comprised of the isolation of the tea anthracnose pathogen using the spore isolation method and the pathogenicity assay of the isolates through indoor and field inoculation tests, with and without wounding. Finally, the pathogen of tea anthracnose was confirmed to be S. theae-sinensis. Additionally, the symptomatic manifestations of the inoculation of S. theae-sinensis and C. camelliae were also compared. This study accurately identified the pathogenic fungus causing tea anthracnose and established a scientific foundation for subsequent research on tea anthracnose and green prevention and control technology.
-
Diseased leaves with typical anthracnose symptoms and the healthy leaves used for inoculation were all collected from the 'Longjing 43' variety of Camellia sinensis in the Tea Research Institute, Chinese Academy of Agricultural Sciences in Hangzhou, Zhejiang Province, China.
For pathogen culture, potato dextrose agar (PDA) or Tea Dextrose Agar (TDA) media was used, and the composition of the media was as follows: PDA: 200 g potato power, 20 g glucose, 20 g agar, and water 1000 mL; TDA: 200 g tea shoots, 20 g glucose, 20 g agar, and water to make the volume 1,000 mL.
The strain of Colletotrichum camelliae used in this study was previously preserved in our lab.
Isolation and culture of the pathogen
-
The spore isolation method[20] was adopted as follows: firstly the leaves with typical lesions of anthracnose were collected from the field, and these leaves were placed in a petri dish with wet filter paper at the bottom and kept in an incubator (25 °C temperature for 12 h in light, 23 °C for 12 h in dark, and relative humidity 75%) for 1−2 d until the cream to pale yellow conidial masses exuded from ostioles of acervuli on the lesion. Then the conidial masses were picked up with a sterilized dissecting knife under the dissecting microscope and streak cultured on a PDA or TDA plate. After 3−4 d of cultivation in the incubator, mycelial transfers were made from the colony periphery onto a new PDA or TDA plate. About 2−3 weeks later, when the yellowish to cream conidial drops appeared in the colonies on the plate, the conidium was picked with the sterilized needle to retransplant the culture, and the pure isolates were obtained. Purified isolates were preserved at 4 °C for further use. Cultivation of the isolates was conducted on both PDA and TDA at 25 °C, and the colony diameter was measured after 15 d of cultivation in the incubator.
Pathogenicity tests
Inoculation assay in the lab
-
Before inoculation tests, young leaves (the second or third leaves below the bud) were collected from healthy tea trees in the field, washed with tap water, and air dried, then sprayed with 70% alcohol twice. Afterward, three types of inoculation tests were conducted. (a) Inoculation with mycelial disc on wounded leaves: First, a sterilized screwdriver was used to puncture the leaves from the dorsal side to make crosswise wounded spots (2 mm × 2 mm) in the middle parts of each leaf on both sides[21]. Subsequently, 7 mm mycelial discs derived from the colonies that had been cultivated on PDA for 12 d were placed over every wound of tea leaves. A blank PDA disc was employed as a control. (b) Inoculation with spore suspension on wounded leaves: The spore suspension of the isolate has been prepared using 5% sterilized tea decoction[17] flushing the spores from the TDA plate cultivated in the incubator for 25 d. Spore concentration was measured with a hemocytometer, and adjusted to 1.4 × 107 spores/mL. Detached tea leaves were wounded by sterile needles and a sterile screwdriver as previously mentioned respectively. Sterilized needles (0.5 mm diam.) were used to wound four times from the dorsal side in the middle parts of each leaf on both sides. Thereafter, 10 μL of conidial suspension was deposited on each side of the tested leaves using a pipette, with 10 μL of 5% sterilized tea decoction acting as a control. (c) Inoculation with spore suspension on non-wounded leaves: A constant quantity of spore suspension (1.4 × 107 spores/mL in 5% sterilized tea decoction, 0.5 mL/leaf) was uniformly smeared onto the leaf surface of both upper and dorsal side with defatted cotton balls, while the same quantity of 5% sterilized tea decoction was applied as a control. Following inoculation, the leaves were wrapped at the petiole with moistened cotton balls, and placed in a 9.0 cm diameter, 2.2 cm height plastic petri dish, and the petri dish was wrapped with the cling film. Subsequently, the petri dishes were incubated in a biological incubator (light cycle for 12 h at 25 °C, dark cycle for 12 h at 23 °C, and relative humidity 75%), and the cling film was removed after 48 h. A total of three replicates were conducted for each treatment, with two leaves per replicate. The experiment was repeated three times. After 2 d post-inoculation, the leaves were examined for symptom development daily. The longitudinal diameter of the lesion was measured and the incidence rate for each treatment was calculated according to the following formula: Incidence rate (%) = [L1/L2] × 100, where L1 indicates the number of symptomatic leaves after inoculation and L2 indicates the total number of leaves for testing. To complete Koch's postulates, the pathogen was re-isolated from the symptomatic lesion of the inoculated leaves and identified via molecular and phylogenetic analysis.
Inoculation assay in the field
-
The experiment was conducted in the experimental base of the Tea Research Institute, Chinese Academy of Agricultural Sciences in Shengzhou, Shaoxing, Zhejiang, China (20°48'48" E, 29°45'3" N, altitude 60.25 m) from May to June 2023. Meteorological information during the field inoculation experiment as listed in Supplementary Table S1. The section of the tea garden was approximately 20 years old, with the 'Longjing 43' tea variety. The spore suspension of the isolate (1.4 × 107 per mL in 5% sterile tea decoction) was prepared for inoculation. The study employed two inoculation methods, including inoculation with and without wounding, with three replications per treatment. Each replication consisted of five tea shoots, each shoot inoculated with a total of three leaves under the bud: the 1st leaf, the 2nd leaf, or the 3rd leaf. Briefly, tea shoots with similar growth and tenderness were selected, and then the surface of the leaves was disinfected with 70% alcohol. Wounds were conducted with a sterilized needle (0.5 mm diam.) by inserting one time on both sides of the leaf. Subsequently, inoculation by smearing spore suspension with a defatted cotton ball was performed, with 5% sterilized tea decoction acting as control. Immediately following inoculation, the inoculated tea shoots were covered with a gram line bag. One wet cotton ball was placed inside the bag to moisturize it, and the bag was then secured tightly. Forty-eight hours later, the plastic bag was removed. The inoculated leaves were examined and recorded at 7, 14, and 20 d post-inoculation, respectively. The incidence rate was calculated according to the following formulas separately: Incidence rate of shoots (%) = [S1/S2] × 100, where S1 indicates the number of shoots with symptomatic leaves after inoculation and S2 indicates the total number of shoots for testing. Incidence rate of leaves (%) = [L1/L2] × 100, where L1 indicates the number of symptomatic leaves after inoculation and L2 indicates the total number of leaves for testing. The pathogen was re-isolated from the symptomatic lesion of the inoculated leaves and identified via molecular and phylogenetic analysis.
Molecular identification of isolate strains
-
The strain A230402 isolated from typical lesion of anthracnose in the section on 'Isolation and culture of the pathogen' was used in pathogenicity tests. The isolates AL1, AL2, AL3, and AL4 were obtained from infected leaves subjected to such treatment as inoculation with spore suspension on non-wounded leaves, on pinprick wounded leaves, on cross wounded leaves, and inoculation with mycelium disc respectively, and the isolates AF1 and AF2 were obtained from diseased leaves of the treatments with and without wounding in field assay, respectively.
The mycelia and spores of these isolates were scraped into 2 mL sterile tubes for DNA extraction following the instructions of the Plant Genome DNA Kit (DP305-02, TIANGEN, China). The concentration and quality of DNA samples were detected by NanoDrop 2000 (Thermo Fisher Scientific, USA). Then genes of internal transcribed spacer (ITS)[22], large subunit ribosomal RNA (LSU)[23] and translation elongation factor 1-alpha (TEF)[24] were amplified by PCR. The forward and reverse primer sequences of three above genes are as follows: ITS1: 5'-TCCGTAGGTGAACCTGCGG-3', ITS4: 5'-TCCTCCGCTTATTGATATGC-3'; LSUF: 5'-GCATATCAATAAGCGGAGG-3', LSUR: 5'-GGTCCGTGTTTCAAGACGG-3'; TEFF: 5'-CATCGAGAAGT-TCGAGAAGG-3', TEFR: 5'-TACTTGAAGGAACCCTTACC-3'. PCR was performed in a 50 μL reaction mixture containing 25 μL Green Taq Mix (Vazyme, China), 2 μL forward primer, 2 μL reverse primer, 100 ng DNA, and sterile deionized water. The PCR reaction program contained the following steps: 95 °C for 3 min; 35 cycles of denaturation at 95 °C for 15 s, annealing at 58 °C for 15 s, and extension at 72 °C for 40 s, with a final extension at 72 °C for 5 min. PCR products were detected by 1% agarose gel electrophoresis to determine the correctness of DNA bandings, and sequenced. The resulting ITS, LSU, and TEF sequences were blasted in the NCBI database (
www.ncbi.nlm.nih.gov ) to confirm the genus of isolates. The sequences of the ITS, LSU, and TEF genes of reference strains[5,9,25] and Ceratosphaeria aquatica and Pyricularia grisea[25] as outgroup strains were downloaded from the GenBank database of NCBI. All these sequences along with those obtained in this study were recombined by ITS-LSU-TEF order. The phylogenetic analysis of these sequences used the Maximum Likelihood method in MEGA 7.0. GenBank accession numbers of all sequences are listed in Supplementary Table S2.Symptom comparison between pathogen isolates and C. camelliae
-
Mature leaves (the fourth or fifth leaves below the bud)[12] were collected from healthy tea trees for inoculation of detached leaves. After making crosswise wounds (2 mm × 2 mm) in the middle parts of each leaf on both sides with a sterilized screwdriver, 10 μL of spore suspension was dripped on the wounds to inoculate the leaves with spores at a concentration of 6.6 × 107 spores/mL (D. theae-sinensis) and 5.0 × 105 spores/mL (C. camelliae), with 10 μL of 5% sterilized tea decoction acting as a control, respectively. A total of three replicates were conducted for each treatment, with six leaves per replicate. After 5 d post-inoculation, the symptoms were observed and recorded daily.
Statistical analysis
-
The data was processed using Excel 2017 and statistically analyzed with SPSS (Statistical Products and Services Solutions, IBM, USA) using Duncan's Multiple Range Test at a significance level of p < 0.05. All experiments were conducted with at least three independent biological replicates. The data is presented as the mean ± standard deviation (SD) of three biological replicates.
-
Tea anthracnose primarily affects mature leaves of tea trees, particularly the fourth to sixth leaves beneath the bud (Fig. 1), although it can also occur on young and old leaves. The symptoms gradually appear in a consistent manner, with the initial stage characterized by the presence of dark green water-soaked lesions, which often spread and expand along the leaf veins, eventually turning brown or reddish brown. In a later stage, grayish-white lesions emerge, accompanied by the presence of numerous black prominent granules on the upper side of the lesions.

Figure 1.
Symptoms of naturally occurring tea anthracnose in the field. (a) Initial lesion. (b) Initial lesion enlargement. (c) Typical lesion. (d) Advanced stages of diseases.
After incubation in moistened and warm conditions for 24−48 h, the cream to pale yellow conidial masses exuded from ostioles of acervuli on the lesions of diseased leaves (Supplementary Fig. S1). Pathogen isolates were obtained after purification by picking up the conidial masses and transferring them to PDA or TDA for culture. The morphology of the colonies on PDA and TDA medium was essentially identical, whereas the growth of fungus was slower and the number of pycnidia was higher on TDA. The colony diameter was 72.7 ± 0.9 mm (PDA) and 70.0 ± 2.6 mm (TDA) after 15 d of incubation. The colonies were white, dense, and exhibited an undeveloped aerial mycelium. In the later stage, the yellowish to cream conidial drops appeared in the colonies on the plate. Following a 25-d incubation period, a considerable number of conidia were produced on the TDA. The conidia are predominantly fusiform or long ellipsoid, measuring 4.48−6.98 μm × 2.11−3.37 μm, with an average ± standard deviation of 5.57 ± 0.53 μm × 2.59 ± 0.30 μm, with a L/W ratio of 2.18 (n = 45). They typically contain two oil droplets (Fig. 2).
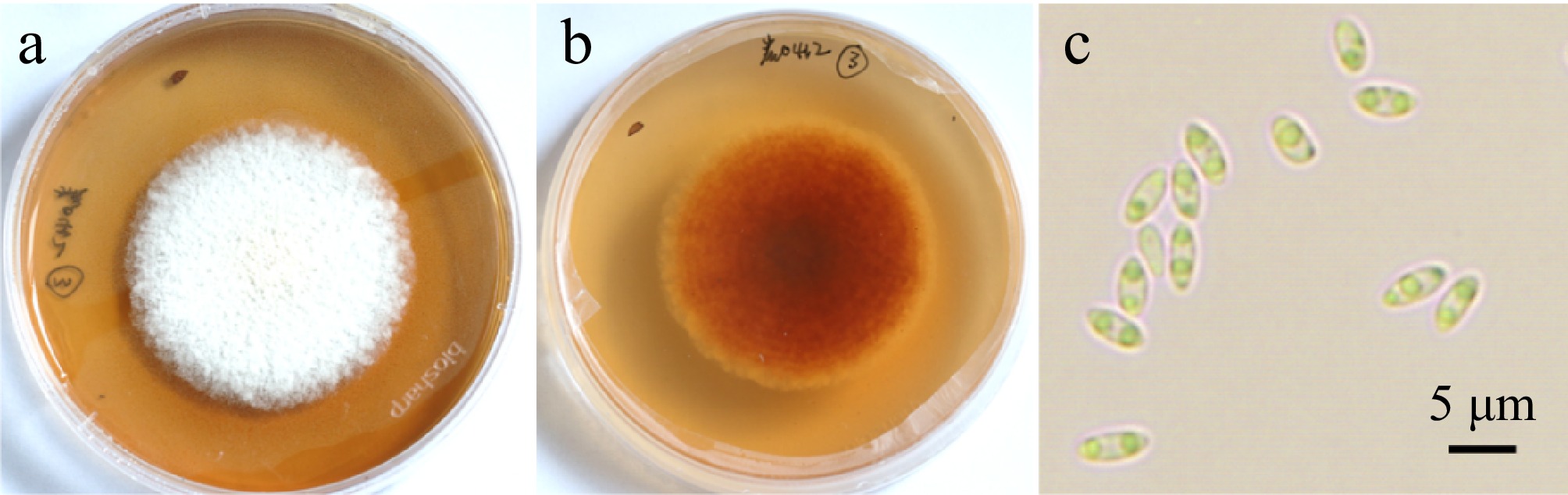
Figure 2.
Colony morphology and spore morphology of pathogen isolate cultured on TDA medium for 15 d. (a) Upper side of colonies. (b) Reverse side of colonies. (c) Conidia. Scale bar = 5 μm.
Determination of pathogenicity of pathogen isolates
Laboratory inoculation
-
The results of the laboratory inoculation tests demonstrated that anthracnose could be successfully induced by inoculation with and without wounding and that the symptoms of the disease were fully consistent with those observed in the field (Fig. 3a, b). Inoculation of the mycelium disc on wounded leaves resulted in the initial manifestation of disease three days post-inoculation. While inoculation with spore suspension on wounded leaves resulted in the onset of disease becoming visible to the naked eye four days after inoculation. The initial lesions manifested as yellowing of the leaf color around the wound, browning of the leaf veins, and localized water loss and wilting. These changes intensified day by day until typical lesions appeared on the 7th day (Fig. 3c, d). Subsequently, large wilting lesions emerged after 12−13 d, accompanied by increased acervuli on the lesions. Furthermore, the symptoms observed in inoculated cross-wounded leaves and pinprick-wounded leaves were identical, and the disease manifested on all inoculated leaves, while none of the control leaves developed the disease, with the exception of one small dead spot on one wound of the mycelium disc inoculated control leaves. The inoculation with spore suspension without wounding initiated the symptoms of the disease after five days, and the symptoms exhibited by the disease were not distinguishable from those observed in the inoculation with wounding. However, in comparison to the inoculation with wounding, the incidence of the inoculation without wounding was much lower, with only one-third of the inoculated leaves developing disease and none of the control leaves developing disease. Koch's postulates were confirmed by reisolating the same fungi and verifying its colony and morphological characteristics.
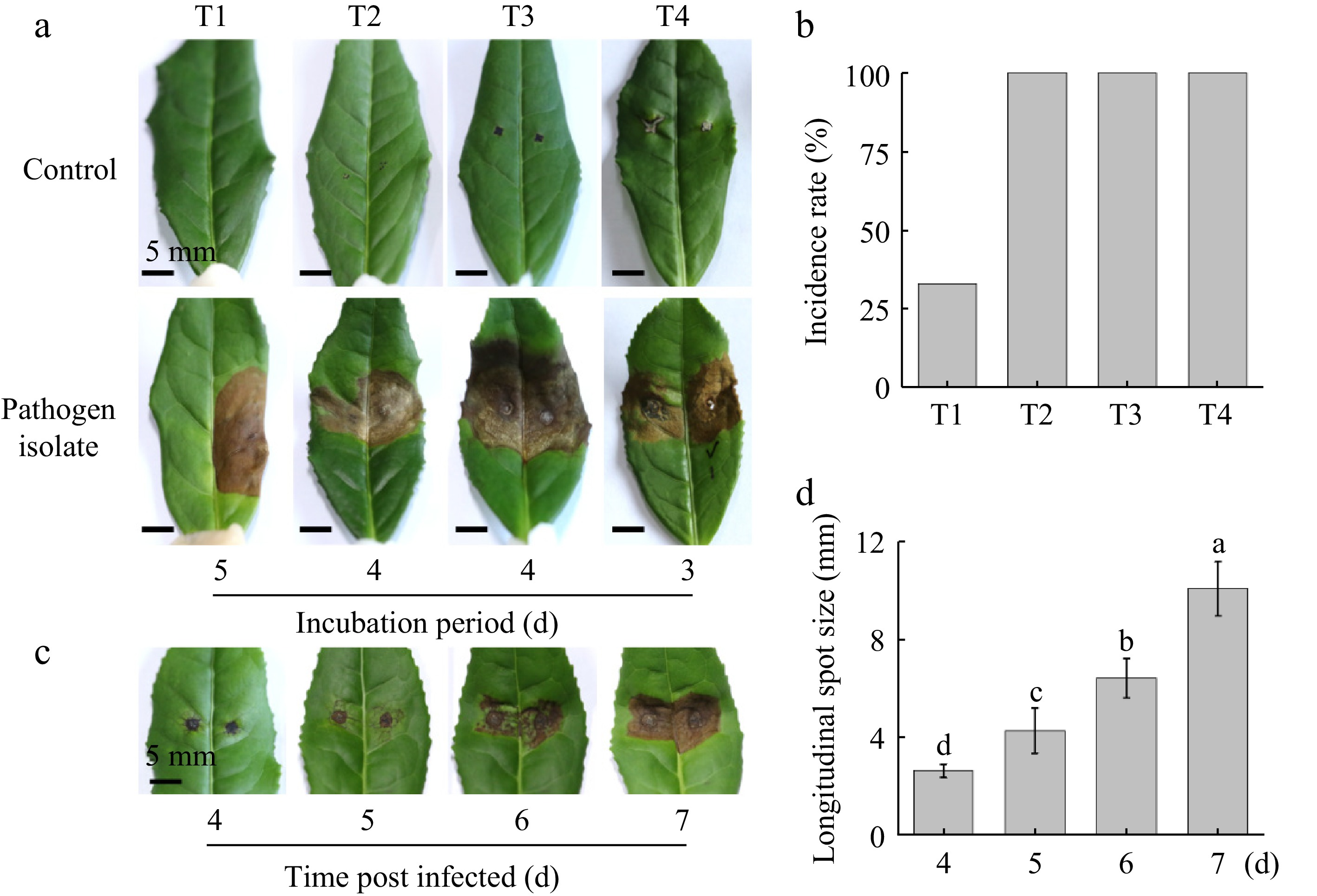
Figure 3.
Pathogenicity of the isolate tested in the laboratory. (a) Symptoms and incubation period of different inoculation methods, T1: Inoculation with spores on non-wounded leaves; T2: Inoculation with spores on pin prick wounded leaves; T3: Inoculation with spores on cross wounded leaves; T4: Inoculation with mycelium disc. T1−T3: Symptoms on the 13th-d post inoculation; T4: Symptoms on the 7th-day post inoculation. (b) Disease incidence rate of different inoculation methods. (c) Appearance changes of lesion post inoculation with T3 method. (d) Longitudinal diameter of the lesion post inoculation with T3 method. The mean denoted by the different lowercase letters indicates statistically significant differences between the treatments according to Duncan's Multiple Range Test (DMRT) at p < 0.05. Scale bars: (a) = 5 mm.
Field inoculation
-
Inoculation by spore suspension with and without wounding resulted in disease development (Table 1). Furthermore, the lesions phenotypes observed in the inoculated tea plants were identical to those observed in naturally infected tea plants in the field (Supplemental Fig. S2). While no discernible disease symptoms were observed 7 d after inoculation in the field, investigations at 14 d after inoculation revealed the presence of typical lesions in all inoculation treatments. Among the treatments, the average incidence of shoots in the treatment of inoculation without wounding was 20%, while the incidence of inoculation with wounding was significantly higher at 66.7%. The average incidence of leaf in both treatments was 8.9% and 26.7%, respectively. At 20 d after inoculation, a second investigation of incidence was conducted. It was found that the leaf incidence of inoculation without wounding had increased to an average of 13.3%, while that of inoculation with wounding remained stable at 26.7%. The leaf incidence of inoculation without wounding was primarily concentrated in the second and third leaves below the bud, with the third leaf exhibiting the highest incidence. In contrast, the incidence of inoculation with wounding was observed in all the first to third leaves, with the second leaf exhibiting the highest incidence, followed by the third leaf. In the control treatment, the tea leaves remained healthy all the time after inoculation and no incidence was observed. The fungi were re-isolated from the lesions and cultured on TDA to verify Koch's postulates.
Table 1. Disease incidence via different inoculation methods in the field.
No. Inoculation method Incidence of shoots (%) Incidence of leaves (%) 7 d 14 d 20 d 7 d 14 d 20 d 1 Without wounding 0 20.0 ± 0b 33.3 ± 11.5b 0 8.9 ± 3.8b 13.3 ± 6.7b 2 With wounding 0 66.7 ± 11.5a 66.7 ± 11.5a 0 26.7 ± 6.7a 26.7 ± 6.7a Each treatment has three replications. Each replication consisted of five tea shoots; each shoot inoculated three leaves under the bud. Different lowercase letters in same column indicate significant difference at 0.05 levels. Molecular identification of pathogen isolates
-
The ITS, LSU, and TEF sequences of isolate A230402 were submitted to the NCBI database (Accession Nos PQ586126, PQ611015, and PQ586797, respectively). The BLAST analysis indicated our sequences showed 100% homology with Sinodiscula theae-sinensis and Discula theae-sinensis. According to the latest taxonomic revision[25], D. theae-sinensis is the synonym of S. theae-sinensis. Based on the morphological and molecular data, the isolate A230402 was identified as Sinodiscula theae-sinensis. In the phylogenic tree established based on the combined data, the isolate A230402 and the reisolated strains AL1, AL2, AL3, AL4, AF1, and AF2 were clustered in the same clade. It was revealed that the reisolated strains were completely consistent with the original strain (Fig. 4).
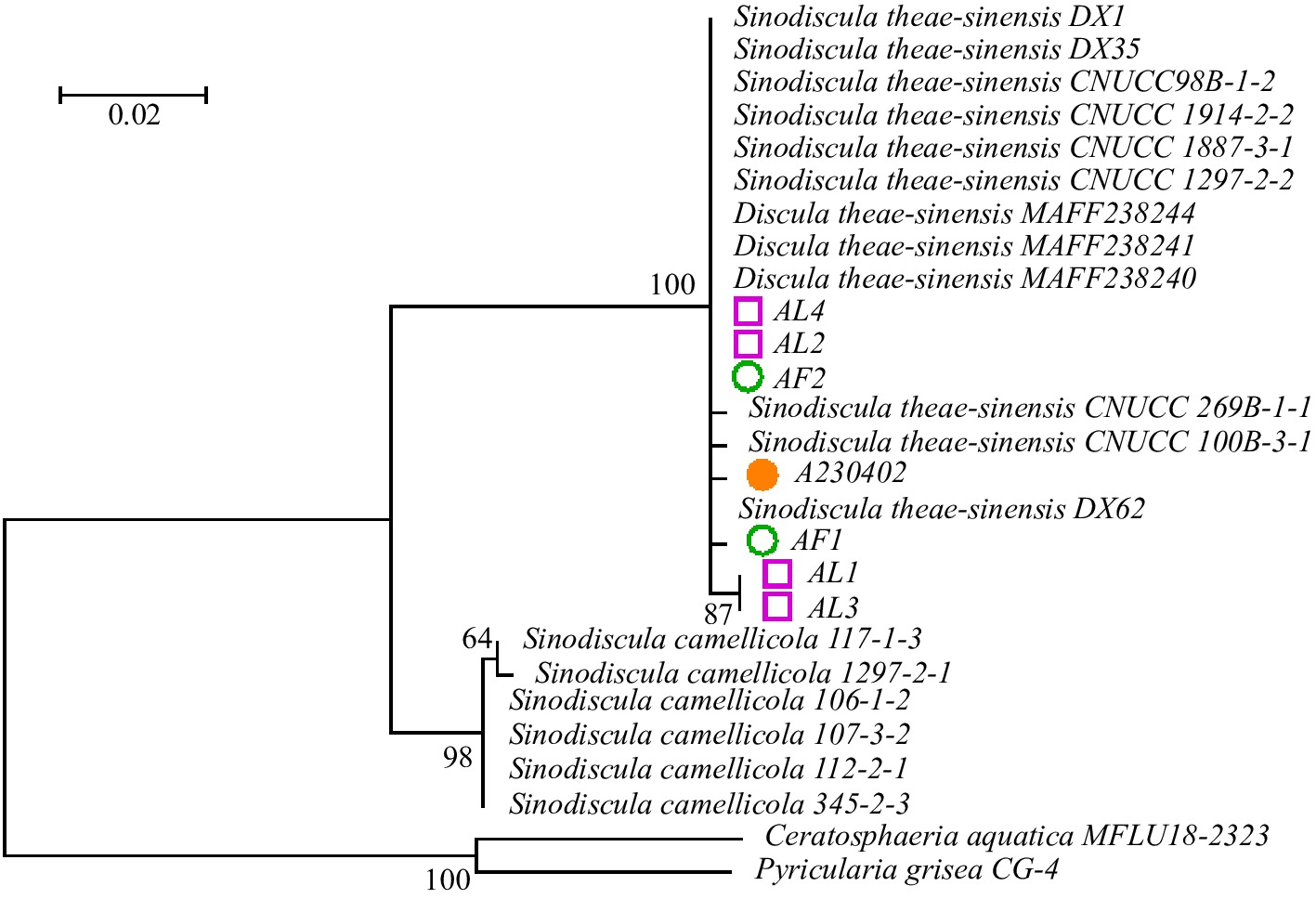
Figure 4.
Phylogenetic analysis of seven isolates obtained in this study and related isolates. The phylogenetic tree was constructed using maximum likelihood (ML) method based on the combined dataset of ITS, LSU, and TEF sequences of Discula sp. isolates and related species. The bootstrap values generated from the ML analyses are indicated at nodes.
Comparison of pathogen isolates with C. camelliae inoculation symptoms
-
Both S. theae-sinensis and C. camelliae could induce the necrotic lesions on the leaves post-inoculation with spore suspension under identical culture conditions, and none necrotic lesions appeared in the control leaves. However, the necrotic lesions induced by the two pathogens species exhibited a significant difference (Fig. 5a). The initial symptoms induced by S. theae-sinensis appeared at 6 d after inoculation manifesting as an inconspicuous watery stain expanded from the invasion point to the surroundings, that gradually lead to the necrosis of leaf veins, same as Figs 1a & 3c. Then the large lesion appeared about 10 d after inoculation, which was irregularly shaped and reddish-brown in color, with a gradual whitening of the lesions in the later stages and the gradual appearance of acervuli. While initial symptoms induced by C. camelliae exhibited the appearance of dark brown, round lesions 5 d after inoculation. The center of these lesions was grayish white, surrounded by dark brown, with regular edges. A considerable number of acervuli were observed in the vicinity of the inoculation wounds, and concentric whorls were frequently observed on the lesions at a later stage. The conidial masses were fleshy red and were often arranged in a concentric whorl-like pattern (Fig. 5b). These symptoms are more consistent with those of tea brown blight rather than anthracnose. The results revealed that the symptoms induced by C. camelliae are distinct and distinguishable from tea anthracnose.

Figure 5.
Comparison of symptoms caused by S. theae-sinensis and C. camelliae. (a) Leaf symptoms induced by S. theae-sinensis and C. camelliae 10-d post-inoculation. (b) Conidial masses of C. camelliae exuded from acervuli on the lesion. Scale bars: (a) = 5 mm, (b) = 100 μm.
-
A greater number of cases of anthracnose-like spots on leaves are observed in tea gardens. These include instances of sunburned or frozen leaves during the summer months, which are prone to the development of large brown spots. To determine whether these similar spots are anthracnose, it is necessary to conduct a careful identification of the disease, examine the morphology of the pathogen, and finally confirm it by isolation and identification of the pathogen and determination of pathogenicity.
Revisiting pathogens of tea anthracnose
-
Adherence to Koch's postulates is of great importance in the identification of disease pathogens. In this study, S. theae-sinensis was isolated from typical anthracnose-diseased leaves and then inoculated with its spore suspension in both the laboratory and field. The incidence of leaves by laboratory and field inoculation with wounding was 100% and 26.7%, respectively, while the incidence in laboratory and field inoculation without wounding was 33.3% and 13.3%, respectively. The inoculation-induced lesions exhibited characteristic features and were consistent with those observed in anthracnose field-onset disease, and the fungus was similarly isolated from inoculated diseased leaves. It can be stated with certainty that S. theae-sinensis is the pathogen of anthracnose. In some previous studies, the spore suspension of D. theae-sinensis did not bring about any obvious disease lesions after inoculation in pathogenicity tests[5,13,25]. Such a discrepancy with our results may be related to a variety of factors, such as the vigor of the inoculated spores, the inoculation conditions, and the observation time.
In this study, we also found that the necrotic lesions induced by C. camelliae were significantly different from tea anthracnose, while similar to tea brown blight[1,17]. Li et al.[5] proposed that tea anthracnose is caused by the synergistic action of D. theae-sinensis and Colletotrichum spp. However, in their test results, the lesions inoculated with a mixture of the two pathogens, D. theae-sinensis and C. fructicola, were only blighted lesions with a diameter of less than 6 mm, and no typical lesions of tea anthracnose appeared. Therefore, the evidence for this conclusion seems insufficient. It is possible that multiple pathogens may co-exist in the field to cause certain symptoms. Some studies have indicated that secondary infestations of C. camelliae can exacerbate the infection of tea net blister blight[17]. We also observed that a prevalence of S. theae-sinensis acervuli intermingled with individual C. camelliae acervuli on the same lesion of anthracnose leaf occasionally. It is postulated that this phenomenon may be attributed to a secondary infestation of tea leaves infected with anthracnose by C. camelliae, or alternatively, the presence of C. camelliae in the leaves as an endophyte.
Methods of isolation of tea anthracnose pathogens
-
In this study, the spore isolation method was employed for the isolation of pathogenic fungi, which is often used to isolate some fungi with large spore production[20]. We exploited the property that a large number of spore masses overflow from the acervuli after moistening and warming, then directly picked the spore masses for isolation and purification. This method is both rapid and highly efficient[26]. However, the conventional tissue isolation method is less efficient and frequently results in the isolation of other microorganisms with strong putrefactive properties, which can lead to confusion and the misidentification of the target pathogens[26,27]. This indicates that the isolation method employed in our study is an efficient method for the acquisition of tea anthracnose pathogens.
Symbiosis refers to different organisms coexisting, and the relationship between plants and endophytic fungi can be mutualistic or parasitic[28]. The genus Colletotrichum is a common fungal species found in tea plants, which is an important component of the endophytic fungi of the tea trees[29]. Therefore, fungi of this genus are often unintentionally isolated when using tissue isolation methods. A single, smaller spot was observed on the control leaf of the fungus discs inoculation test in this study. From the spot we obtained a fungus strain of C. fioriniae, which was a possible endophyte of the tea leaf (data not shown).
Disease name of tea anthracnose
-
The pathogen of tea anthracnose was first identified by Miyake in 1907 near Tokyo, Japan. While the disease name anthracnose was not finalized until 'anthracnose' was used in the book 'Tea Diseases in Japan' by Hara Kanesuke in 1931[17] and has been used ever since. C. camelliae was first identified by Massee in 1899, which causes a disease known as tea brown blight and its sexual generation belongs to Glomerella cingulata[1,17]. It also causes anthracnose in oil tea (Camellia olifera)[30] and anthracnose in Camellia japonica[31]. This study demonstrated that the lesions caused by C. camelliae are not typical anthracnose in the conventional sense, but rather tea brown blight. Given that C. camelliae is the dominant species among Colletotrichum spp. causing 'anthracnose', we suppose that the other species of Colletotrichum spp. may also lead to tea brown blight[32,33].
Guo et al.[25] suggested that the disease caused by fungi of the genus Colletotrichum be referred to as 'anthracnose' while those caused by S. theae-sinensis as 'the tea leaf blight'. However, we find ourselves in disagreement with such categorization. At present, it is still unclear what are the distinctions in symptoms caused by these two pathogens in the field and we believe that it is more reasonable to discuss if changing the disease name or not only after confirming the symptoms caused by its corresponding agent.
-
In the present study, pathogenic isolates were collected from anthracnose-diseased leaves through the spore isolation method and verified for pathogenicity using various inoculation methods. The same isolates were subsequently re-isolated from the inoculated diseased leaves, thereby providing indisputable evidence that the causal organism of anthracnose in Camellia sinensis is S. theae-sinensis. The symptoms induced by C. camelliae are distinct from those observed in tea anthracnose. This paper presents a detailed account of the symptomatic pictorial view of changes in tea leaves following inoculation with S. theae-sinensis and the onset of the disease. This provides a foundation for the identification of tea anthracnose and paves the way for intelligent disease diagnosis in the future.
This work was supported by the Central Public-Interest Scientific Institution Basal Research Fund (1610212022007), the Key Research and Development Plan of Xizang Autonomous Region (XZ202301ZY0014N), and the Innovation Project of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2019-TRICAAS).
-
The authors confirm contribution to the paper as follows: study conception and design: Tang M, Li X, Xiao Q; experiments performing and data analysis: Wang W, Zhang L, Zhou X, Li Z, Guo H; data discussion: Tang M, Li X, Wang W, Li Z; manuscript writing: Tang M, Wang W, Ahammed GJ, Li X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are not publicly available due to privacy but are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Wenli Wang, Xin Li
- Supplementary Table S1 Meteorological information during field inoculation trials.
- Supplementary Table S2 The GenBank accession numbers of different strains for ITS, LSU and TEF sequences.
- Supplementary Fig. S1 (a) Magnified view of lesions in infected tea leaves after incubation in moistened and warm conditions for 24−48 h, (b) The pale yellow conidial masses exuded from ostioles of acervuli on the lesions of diseased leaves.
- Supplemental Fig. S2 The lesion phenotypes observed in the inoculated tea plants in the field and disease incidence in different leaf positions in the field. (a, c) inoculation without wounding; (b, d) inoculation with wounding.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang W, Li X, Li Z, Zhang L, Ahammed GJ, et al. 2025. Revisiting causal organisms of tea anthracnose: pathogen isolation and pathogenicity identification. Beverage Plant Research 5: e016 doi: 10.48130/bpr-0025-0005 

Revisiting causal organisms of tea anthracnose: pathogen isolation and pathogenicity identification
- Received: 24 September 2024
- Revised: 22 January 2025
- Accepted: 19 February 2025
- Published online: 10 June 2025
Abstract: Tea anthracnose is a major disease of tea plant (Camellia sinensis) in all tea-growing regions in China, with the southern tea region being particularly susceptible. Over the past decade, more than 20 species belonging to Colletotrichum have been identified as causal agents of tea anthracnose besides its actual pathogen Discula theae-sinensis. Although researchers have increasingly regarded Colletotrichum spp. as the primary pathogens of tea anthracnose, there is still controversy over whether they cause tea anthracnose. By employing spore isolation methods, here we identified a fungal strain causing typical tea anthracnose disease but not belonging to the genus Colletotrichum. Upon spore isolation of tea anthracnose pathogens, various methods were employed to determine the pathogenicity of the isolate cultures. Eventually, a single isolate was obtained through the spore isolation method, which was subsequently identified as Sinodiscula theae-sinensis (Discula theae-sinensis) based on its morphological and molecular characteristics. The pathogenicity assay revealed that with either wound or non-wound inoculation the isolate exhibited typical lesions that were identical to those observed on the original diseased leaves. Furthermore, all the re-isolation strains obtained from the inoculated leaves was identical to the original strain. The pathogen of tea anthracnose was further confirmed as S. theae-sinensis by Koch's validation. Additionally, the symptomatic manifestations of the inoculation by S. theae-sinensis and Colletotrichum camelliae were also compared. The symptoms induced by C. camelliae are distinct from those observed in anthracnose. This study clarified the causal pathogen of tea anthracnose, thus a crucial foundation for the identification, prevention, and control of tea anthracnose.
-
Key words:
- Anthracnose /
- Camellia sinensis /
- Discula /
- Fungal pathogen /
- Koch's postulates /
- Spore separation method