








-
Plants' responses to internal or external cues and control of flowering time are vital for their development, survival, and reproduction, which are regulated via complex mechanisms, including transcriptional and epigenetic regulation. In addition, epitranscriptomic RNA methylation, particularly N6-methyadenosine (m6A), has recently emerged as a potent regulatory mechanism of stress adaptation and floral transition[1−3]. The m6A modification (m6A markers) in a transcript is added by methyltransferases (writers), removed by demethylases (erasers), and recognized by m6A-binding proteins (readers). The domain of YT521-B homology (YTH)-containing proteins in animals and their orthologous proteins in plants, named EVOLUTIONARILY CONSERVED C-TERMINAL REGION (ECT) proteins, have been identified as m6A readers. The writing, erasing, and reading of m6A markers in mRNA play a crucial role in regulating mRNA's fate, including stability, splicing, transport, and translation. For overview of the m6A machinery and its function, refer to the reviews by Liang et al.[4], Tang et al.[2], Hu et al.[5], and Song et al.[6].
Liquid–liquid phase separation (LLPS) is a dynamic process where proteins and other biomolecules condense into membraneless organelles (e.g., stress granules, promyelocytic leukemia [PML]-nuclear bodies[7−9]), enabling spatiotemporal regulation of cellular functions[10−12]. These liquid-like droplets concentrate molecules while maintaining diffusibility, allowing rapid assembly and disassembly for efficient biological responses[13,14]. LLPS is driven by a complex interplay of multivalent interactions, sequence-encoded features (e.g., low-complexity repeats, charge patterns), and environmental factors (pH, ions, RNA)[15−18]. Similar to the post-translational modifications necessary for phase separation, m6A modifications can modulate phase separation by altering RNA–protein and RNA–RNA interactions and their structure[19,20].
Usually, m6A-modified mRNAs act as scaffolds for reader proteins, driving their condensation into phase-separated droplets. Many studies in animals have demonstrated that YTH proteins, the confirmed m6A readers in animals, play a crucial role in phase separation[21]. For examples, eIF3-S6 binds m6A-modified EcR/Cht10 mRNAs, forming LLPS condensates to stabilize molting-responsive mRNAs in locusts[22]. YTHDF2 undergoes LLPS when bound to m6A-modified PIK3R2 mRNA, promoting mRNA degradation in arsenic-induced stress[23]. IGF2BP1 forms LLPS condensates upon binding GPX4 mRNA, stabilized by lncRNA RUNX1-IT1 in breast cancer[24]. Similarly, the significance of m6A readers in phase separation is increasingly being unraveled in plants. However, the significance of m6A modifications in LLPS in response to phytohormones and the mechanistic and cellular role of m6A modifications in phase separation in plants remain largely unknown. Two recent studies have investigated m6A readers' function in phase separation during the abscisic acid (ABA) response in Arabidopsis and flowering in rice[25,26].
In one study, Wu et al. demonstrated that ABA signalling upregulates the m6A reader ECT8, triggers LLPS to form stress granules (SGs), where ECT8 temporarily sequesters m6A-marked transcripts of PYRABACTIN RESISTANCE 1-LIKE 7 (PYL7), reducing PYL7 mRNA for translation and negatively feedback regulating PYL7 levels and ABA signalling[26]. In another study, Cui et al. demonstrated that YTH07 interacts with Early Heading Date 6 (EHD6) in rice, sequestering the flowering repressor CONSTANS-like 4 (OsCOL4) mRNA into phase-separated condensates, resulting in early flowering[25]. Notably, as an RNA-binding protein (RBP), EHD6 physically interacts with YTH07 to enhance the m6A binding and relocation of YTH07 from the cytoplasm into ribonucleoprotein (RNP) granules through LLPS. This sequesters OsCOL4 mRNA and reduces its protein abundance[25] (Fig. 1).
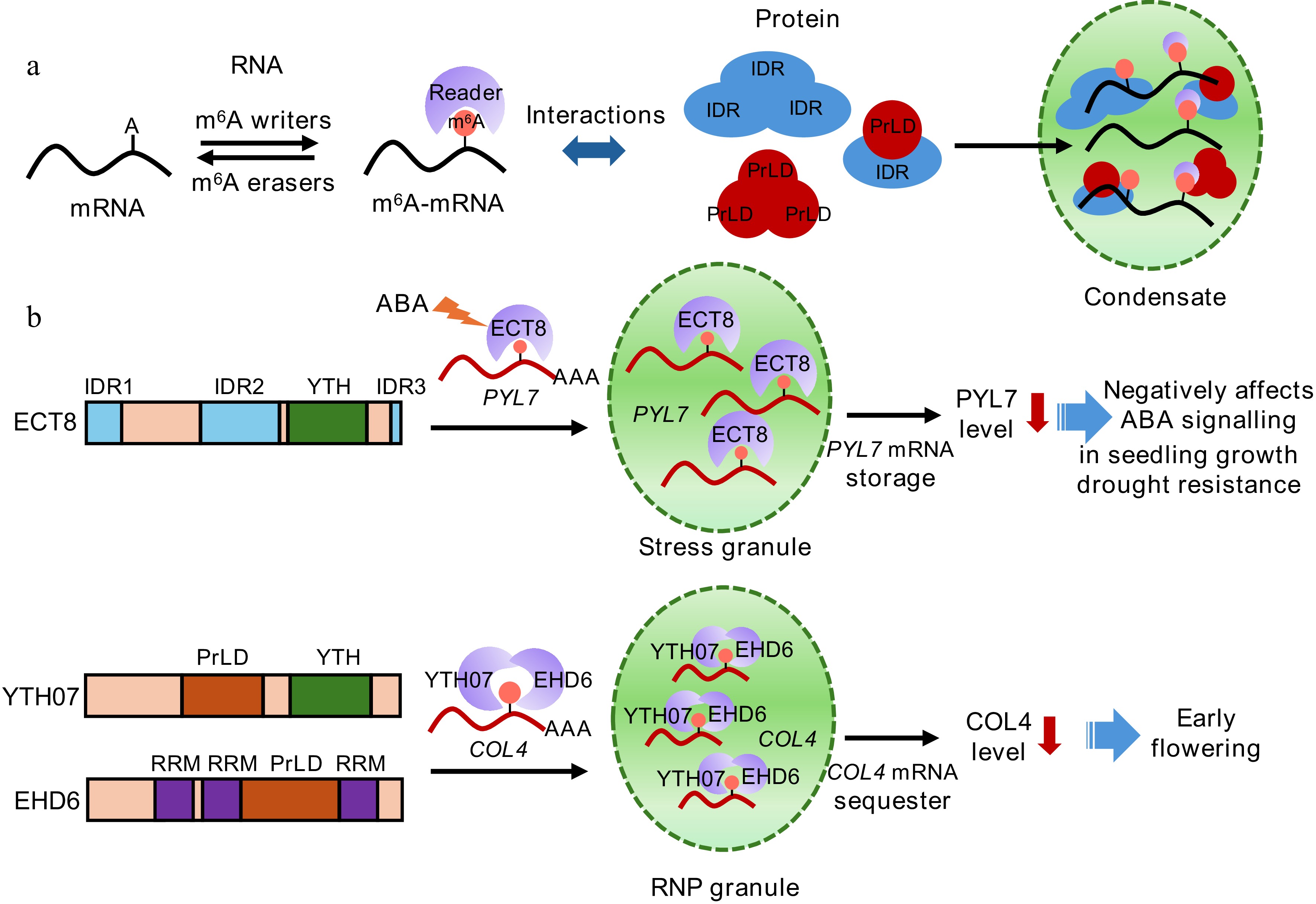
Figure 1.
The domain structures and phase separation mediated by m6A reader proteins. (a) m6A writers install m6A markers on mRNA, and m6A readers bind to the m6A-modified mRNA. The proteins harboring intrinsically disordered regions (IDRs) or prion-like domains (PrLDs) interact with the eraser-bound m6A-modified mRNA to form condensates via phase separation. (b) The m6A reader ECT8, YTH07, and EHD6 contain IDRs, PrLDs, RNA-recognition motifs (RRMs), or YT521-B homology (YTH) domains. The abscisic acid (ABA)-induced ECT8 binds to the m6A-modified PYL7 mRNA, an ABA receptor, sequestering PYL7 transcripts into stress granules and reducing PYL7 protein levels, and leading to a negative ABA response in seedling growth and drought resistance. The YTH07‒EHD6 complex binds to the m6A-modified COL4 mRNA, a flowering repressor, sequestering COL4 transcripts into ribonucleoprotein (RNP) granule and leading to early flowering.
Proteins undergoing LLPS generally harbor intrinsically disordered regions (IDRs), low-complexity sequence domains (LCDs), or prion-like domains (PrLDs)[21]. CPSF30, an Arabidopsis YTH m6A reader harboring IDRs or PrLDs, promotes the formation of liquid-like nuclear bodies, which is important for the floral transition and ABA response caused by the polyadenylation site choice[27]. Notably, prion-like amino acid composition (PLAAC) analysis (
http://plaac.wi.mit.edu ) indicated that ECT2‒8 and ECT10 in Arabidopsis contain putative PrLDs, and the condensate formation of some of these has been experimentally verified[28]. ECT1, ECT9, and ECT11 exhibit different functions depending on the properties of the YTH domains and IDRs[29]. Moreover, ECT1 undergoes LLPS, and its N-terminal PrLD is critical for forming biomolecular condensates in response to salicylic acid or bacterial pathogens[30]. These findings highlight that the IDRs or PrLDs of ECT proteins are crucial for their functionality and phase separation in response to phytohormones or pathogen infection.ECT8 contains three IDRs (Fig. 1). ECT8 forms condensates in response to ABA, and IDR2 is required for LLPS of ECT8 and its function in ABA signaling[26]. To determine the importance of m6A-binding ability of ECT8 in phase separation, Wu et al. mutated the tryptophan (W) in the YTH domain to alanine (A) residues and discovered the significance of m6A binding for ECT8-mediated LLPS. Notably, among the ABA receptors, including PYL7, PYL9, PYL1-1, and PYL4-1, ECT8 regulates ABA signaling, mainly through sequestering the PYL7 mRNA[26]. Furthermore, by analyzing the m6A levels and transcript levels of PYL7 mRNA in ect8 mutants complemented with the wild-type ECT8, IDR2-deficient ECT8, or W-to-A mutated ECT8, they confirmed that both the m6A-binding capacity and phase separation potential of ECT8 are crucial for its effects on PYL7 in SGs[26].
EHD6 harbors one PrLD and three RRMs, and YTH07 contains a PrLD (Fig. 1). Notably, EHD6 can bind to the m6A-modified RNAs, suggesting it as a novel m6A reader[25]. Interestingly, EHD6 and YTH07 interact together, strengthening the m6A-binding ability of the EHD6‒YTH07 complex. The PrLD domain is vital for the condensation of EHD6 that recruits YTH07 to RNP granules through LLPS, which sequesters OsCOL4 mRNA and represses the accumulation of OsCOL4 protein[25]. Importantly, this study shed new light on the mechanism of m6A recognition and phase separation by the concerted action of an RBP and m6A readers. This finding raises the intriguing questions of whether other m6A readers also interact with as yet unidentified proteins and whether the m6A reader‒protein interactions can influence the target mRNA recognition, m6A-binding, and phase separation capacity in response to development and stress cues.
In summary, the work by Wu et al.[26] demonstrated the m6A reader-mediated phase separation to be a key checkpoint for ABA signaling regulation, and the work by Cui et al.[25] revealed the m6A recognition and phase separation of the EHD6‒YTH07 complex as a pivotal regulatory mechanism for controlling flowering time. These works are valuable additions to our understanding of the m6A reader-mediated condensate formation in ABA response and flowering. Given that ECT2/3/4 stabilizes ABA response-related genes[31] and that ALKBH9B/10B-mediated m6A demethylation is associated with the ABA response in Arabidopsis[32,33], it will be of great interest to explore whether the regulation of m6A levels and the recognition of m6A markers by m6A readers and erasers, respectively, modulate phase separation of the m6A-modified mRNAs in response to ABA. In addition to the ABA response, another study revealed that salt stress-induced ECT8 undergoes LLPS to sequester m6A-modified mRNAs of negative salt stress regulators into processing bodies, leading to salt tolerance[34]. These findings highlight that ECT8-mediated phase separation is associated with both salt stress and ABA responses by targeting different m6A-modifed mRNAs depending on the phytohormones and stress conditions. However, the mechanism underlying the selectivity of ECT8 for target mRNAs during ABA and stress remains unresolved. Future research identifying other m6A readers undergoing LLPS and determining the mechanisms by which m6A readers regulate plants' development and responses to the environment will illuminate the significance of epitranscriptomic RNA modifications in plants. Furthermore, identifying and characterizing other protein factors interacting with m6A readers will be needed to deepen our understanding of the mechanistic and cellular role of m6A-mediated phase separation in plants' responses to developmental, environmental, and phytohormonal signals.
HTML
This work was funded by the Mid-Career Researcher Program through the National Research Foundation of Korea funded by the Ministry of Science, ICT and Future Planning (NRF-2021R1A2C1004187), Republic of Korea; the Basic Research Program of Jiangsu (BK20251928); the Natural Science Foundation of Jiangsu Higher Education Institutions of China (24KJA180004); and the Qing Lan Project of Jiangsu Province (2024), China.
-
The authors confirm their contributions to the paper as follows: writing the mauscript: Xu T, Tao J, Cai J, Kang H; reviewing the manuscript and providing advice: Kang H. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no datasets were generated in this study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Tao Xu, Jiaju Tao
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Xu T, Tao J, Cai J, Kang H. 2025. mRNA m6A reader-mediated phase separation as a potent regulator of abscisic acid response and flowering. Epigenetics Insights 18: e013 doi: 10.48130/epi-0025-0013


|