









-
Rubber, a high-molecular-weight polymer composed of cis-polyisoprene (CPI) and trans-polyisoprene (TPI), serves as an indispensable industrial material that supports national industrialization and technological development[1−4]. With the development of high-throughput sequencing technologies, reference genomes of rubber-producing plants have been successively assembled, significantly enhancing the understanding of natural rubber biosynthesis[5−9]. In general, the core enzyme cis-prenyltransferase (CPT) interacts with the small rubber particle protein (SRPP) and the rubber elongation factor (REF) to form a rubber synthesis complex (RSC) that regulates the process of natural rubber biosynthesis[10−12].
E. ulmoides is an important strategic resource tree species in China[13]. Unlike Hevea brasiliensis, which is the primary source of CPI, E. ulmoides produces high-quality TPI in its peels, leaves, and bark. It has evolved a distinct biosynthetic pathway for long-chain TPI production mediated by farnesyl diphosphate synthases (FPSs)[6]. The E. ulmoides genome contains five FPS paralogs (EuFPS1-5), with EuFPS1, EuFPS3, and EuFPS5 forming an evolutionarily distinct clade that diverges from EuFPS2, EuFPS4, and all FPS genes from H. brasiliensis[6,7]. Heterologous expression of EuFPS5 in tobacco confirmed its capacity to produce TPI[6,14], demonstrating its role in this specialized biosynthetic pathway. In addition, the expansion of the SRPP/REF gene family is tightly and positively correlated with rubber yield in rubber-producing plants[5,8]. E. ulmoides possesses 13 EuSRPP/REF family members, showing a notable expansion compared to most plants, which typically contain no more than six SRPP/REF members except Hevea, which has 18 members[6,7]. These lineage-specific expansions and functional diversification of both the FPS and SRPP/REF gene family represent key evolutionary innovations that distinguish the TPI biosynthesis pathway in E. ulmoides from the CPI production system of H. brasiliensis.
Transcription factors and epigenetic regulators play critical roles in the regulation of rubber biosynthesis. In H. brasiliensis, transcription factors such as HbWRKY1 and HbWRKY83 directly bind to the promoter of HbSRPP genes to suppress their expression[15−17], while HbWRKY14 recruits the histone deacetylase HbHDA3 and the regulatory protein HbVQ4 to form a transcriptional repression complex that further modulates HbSRPP expression and negatively regulates rubber biosynthesis[18,19]. In addition, the HbMYB2-like transcription factor binds to and represses HbFPS1, resulting in reduced rubber biosynthesis activity but increased rubber molecular weight, accompanied by abnormal rubber particle development[20]. Similarly, HbMYB44 interacts with HbbHLH13 to regulate the expression of rubber synthase genes and HbSRPP genes; heterologous overexpression of HbMYB44 in guayule significantly enhances rubber content[21]. Through yeast one-hybrid screening and dual-luciferase assays, researchers revealed that transcription factors EuHDZ25, EuWRKY30, and EuTGA1 positively regulate rubber biosynthesis by directly binding to the EuFPS1 promoter and activating its transcription[22−24].
In this study, high-yielding rubber triploid E. ulmoides plants were generated via polyploid breeding. Through molecular biology analysis, EuCHR11 and EuCHR17 were identified as two key ISWI-family chromatin remodelers, as negative regulators of TPI biosynthesis. Further investigation revealed that EuCHR11 and EuCHR17 directly bind to the promoter of EuSRPP8 and suppress its expression, thereby negatively regulating TPI biosynthesis. These results uncover a previously uncharacterized epigenetic regulatory pathway controlling rubber biosynthesis and provide valuable insights into the molecular mechanisms underlying polyploidy-enhanced rubber production.
-
Fresh seeds of E. ulmoides were treated with 0.05% colchicine solution for 72 h, followed by thorough rinsing with water. After germination, the relative DNA content of variant seedlings was measured using flow cytometry, leading to the successful induction of tetraploid lines. Through phenotypic evaluation and ploidy verification, two outstanding tetraploid plants were identified. A single tetraploid plant was used as the male parent and crossed with elite diploid E. ulmoides lines ‘Huazhong No.10’, which acted as the female parent. Triploidy characterization and phenotypic observation were then performed in the progeny. One-year-old grafted seedlings of diploid, tetraploid, and their triploid hybrids were used as experimental materials and grown under identical environmental conditions. The materials were conserved in the National Germplasm Resources Bank of Major Famous Tree Species in the North (34°55′22″ N, 113°46′16″ E), Yuanyang, Henan Province, China.
Quantitative RT-PCR and data analysis
-
Total RNA was extracted from the sixth and seventh leaves of E. ulmoides collected at three developmental stages: mid-to-late April (small leaf, S1), mid-to-late May (young leaf, S2), and mid-to-late June (mature leaf, S3), using the FastPure Plant Total RNA Isolation Kit (Vazyme, RC401-1). cDNA was synthesized using the 5 × All-In-One RT Master Mix (Abm, G492). For the qRT-PCR assay, SYBR® Green Premix Pro Taq Kit (Rox Plus) was chosen, and EuACT2 was selected as the reference gene. The relative gene expression level was calculated by the 2−ΔΔCᴛ method. Three independent biological replicates were used to ensure reliability and reproducibility.
Detection of the TPI content in E. ulmoides leaves
-
The sixth and seventh leaves of E. ulmoides at stage S3 were randomly harvested, dried, and ground into powder. The powder was treated with 10% NaOH in a heated water bath for 3 h. After drying, the mass was recorded as m1. Eucommia rubber was then extracted with petroleum ether for 10 h. The extract was transferred to a pre-weighed centrifuge tube, dried at 37 °C, and washed repeatedly with anhydrous ethanol until white rubber was obtained. The dried rubber mass was recorded as m2. The rubber content was calculated as (m1−m2)/m1 × 100%.
Domain prediction, sequence alignment, phylogenetic analysis, and chromosome location
-
Protein sequences were obtained from the NCBI databases. The domains of the target proteins were predicted using the SMATT database (
https://smart.embl.de/ ). Sequence alignments were performed using DNAMAN (version 7), and the phylogenetic tree was constructed via the neighbor-joining method in MEGA (version 7). Based on the genome information of E. ulmoides, the gene names of chromatin remodeling enzymes were identified, and their chromosomal locations were mapped using TBtools software.Protein purification and electrophoretic mobility shift assay
-
For protein purification in vitro, the DNA-binding domain of EuCHR11 was cloned into a modified pET30a vector in fusion with MBP and His tags. The primer sequences used for vector construction are listed in Supplementary Table S1. The confirmed vector was transformed into the BL21(DE3) strain for protein expression and purification. Protein purification was performed as previously described[25]. Briefly, the protein expression was induced with 0.1 mM IPTG. The bacterial cells were resuspended in lysis buffer (20 mM Tris–HCl [pH 7.5], 150 mM NaCl, 1 mM DTT, and 1 mM PMSF), sonicated, centrifuged, filtered, and incubated with MBP Resin (Smart-life Sciences, SA077005) for 2 h at 4 °C. The beads were washed with lysis buffer to remove non-specific proteins. Finally, the protein was eluted with lysis buffer containing 10 mM maltose. Protein quality was confirmed by Coomassie blue staining.
For the electrophoretic mobility shift assay (EMSA), the truncated EuCHR11 protein was incubated with 1.25 μM double-stranded DNA labeled with Cy3 fluorescent dye. The protein-DNA complex was incubated in binding buffer (25 mM HEPES–KOH [pH 7.6], 12.5 mM MgCl2, 50 mM KCl, 100 μM EDTA, 0.5% BSA, 5% glycerol, and 1 mM DTT) for 30 min at room temperature. The protein-DNA mixture was separated on a 10% native PAGE gel in 0.5 × TBE at 4 °C for 180 min. The results were detected using a Bio-Rad scanner.
Virus- induced gene silencing (VIGS)
-
The conserved CDS sequence of EuCHR11 was inserted into the pTRV2 vector. The primer sequences used for vector construction are listed in Supplementary Table S1. The recombinant vector was confirmed by sequencing and transformed into the Agrobacterium tumefaciens strain GV3101 for leaf infiltration. For the virus-induced gene silencing (VIGS) assay, young E. ulmoides seedlings were first transplanted into soil and grown under consistent conditions. Agrobacterium cultures containing pTRV1 and those containing either the empty pTRV2 or the pTRV2-EuCHR11 construct were resuspended to an OD600 of 0.8 in a buffer containing 10 mM MES, 10 mM MgCl2, and 100 μM acetosyringone. The pTRV1 suspension was then mixed in a 1:1 ratio with the pTRV2 or pTRV2-EuCHR11 suspension. The cotyledons of the seedlings were chosen as the injection site. Following infiltration, the seedlings were kept in darkness for 3 d, and the silencing efficiency was detected by quantitative real-time PCR after 15 d.
Yeast one hybrid assay (Y1H)
-
For the yeast one hybrid (Y1H) assay, the EuSRPP8 promoter was cloned into the pHis2 vector to create the pHIS2.1-EuSRPP8 plasmid. Concurrently, the coding sequence for the DNA-binding domain (DBD) of EuCHR11 was cloned into the pGADT7 vector. The pGADT7-EuCHR11-C and pHIS2.1-EuSRPP8 plasmid were co-transformed into the Y187 yeast strain using the lithium acetate (LiAc)/polyethylene glycol (PEG) method at 30 °C for 3 d. The transformants were screened on SD/-Trp/-Leu/-His medium containing 50 mM 3-AT to select for positive interactions while inhibiting background growth.
-
In this work, new tetraploid germplasm of E. ulmoides was generated through colchicine-induced mutagenesis and subsequently created a new triploid variety by genetic hybridization, using ‘Huazhong No.10’ as the female parent and the colchicine-induced tetraploid as the male parent. Flow cytometry analysis is performed to determine the DNA content of the hybrid, which exhibited a distinct 3 C peak, whereas the diploid and tetraploid controls displayed characteristic 2 C and 4 C peaks, respectively (Fig. 1a, b). By assessing one-year-old grafted E. ulmoides seedlings, it is observed that a progressive increase in TPI content from diploid to tetraploid and then to triploid plants (Fig. 1c), highlighting the potential of the triploid variety as valuable germplasm for high-yield rubber breeding and further mechanistic studies.
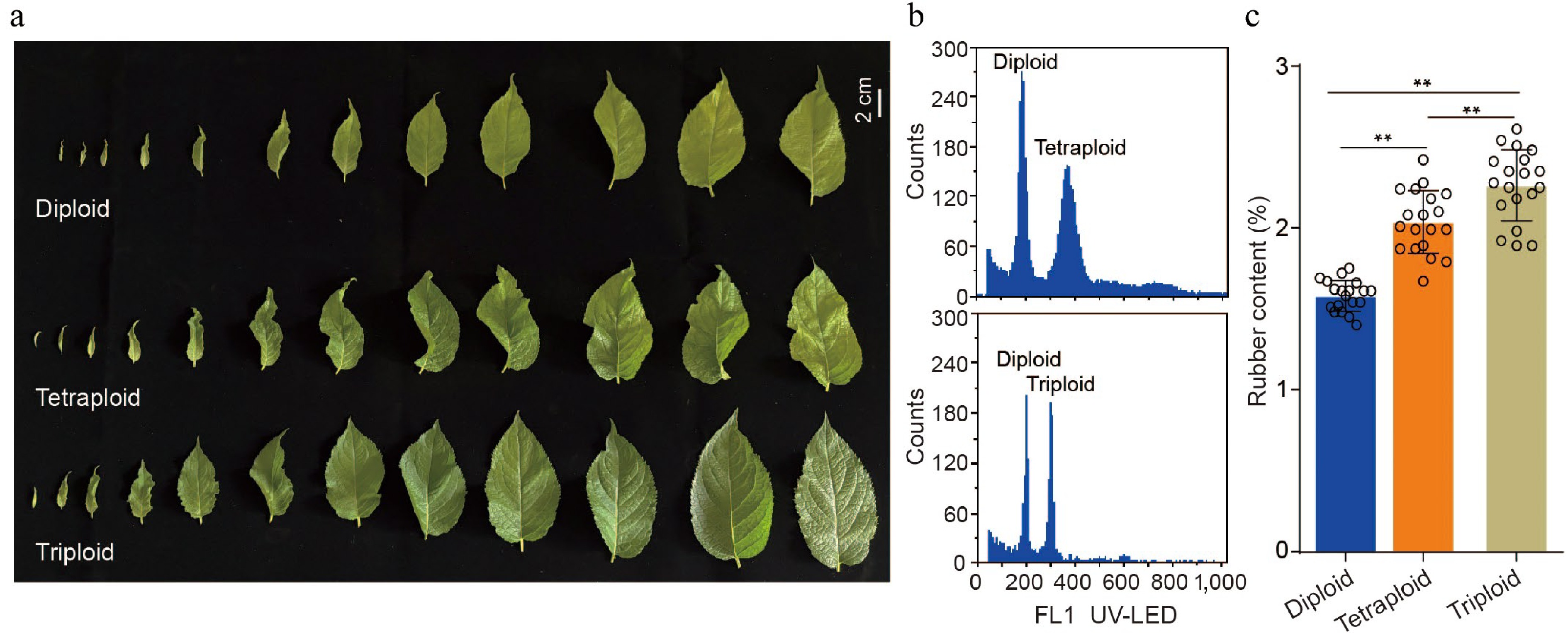
Figure 1.
Analysis of TPI content in leaves of E. ulmoides with different ploidy. (a) Phenotypes of diploid, tetraploid, and triploid leaves collected from the same position at stage S3. (b) Ploidy identification by flow cytometry. The top histogram shows a mixture of diploid and tetraploid nuclei, and the bottom shows a mixture of diploid and triploid nuclei. (c) Statistical analysis of TPI concentration in leaves with different ploidy at stage S3. All p-values were obtained by two-tailed t-tests. * p < 0.05 and ** p < 0.01 represented the highly significant and significant difference, respectively.
The expression levels of EuCHR11 and EuCHR17 continuously decreased throughout the TPI synthesis stage.
-
To identify epigenetic regulators of TPI biosynthesis, chromatin remodelers were focused on. Through genome-wide sequence alignment and phylogenetic footprinting, nine chromatin remodeling enzymes were identified in E. ulmoides, which were distributed across five distinct chromosomes (Supplementary Fig. S1a, S1b). Previous studies demonstrated that the TPI content in E. ulmoides leaves increases significantly from stage S1 (mid-to-late April) to S3 (mid-to-late June)[23,24]. Leaf samples were therefore collected at stages S1, S2 (mid-to-late May), and S3 from diploid, triploid, and tetraploid E. ulmoides, and real-time PCR analysis was performed to detect the expression of the genes encoding these chromatin remodeling enzymes. The results showed that the transcript levels of EuCHR11 and EuCHR17 were consistently downregulated during the transition from stage S1 to S3 in all varieties (Fig. 2a), whereas TPI content increased rapidly over the same period[23,24], suggesting a negative correlation between EuCHR11/17 expression and TPI accumulation in E. ulmoides. In contrast, the expression of other chromatin remodeling genes displayed variable patterns from stage S1 to S3 in diploid, triploid, and tetraploid E. ulmoides plants (Fig. 2a).
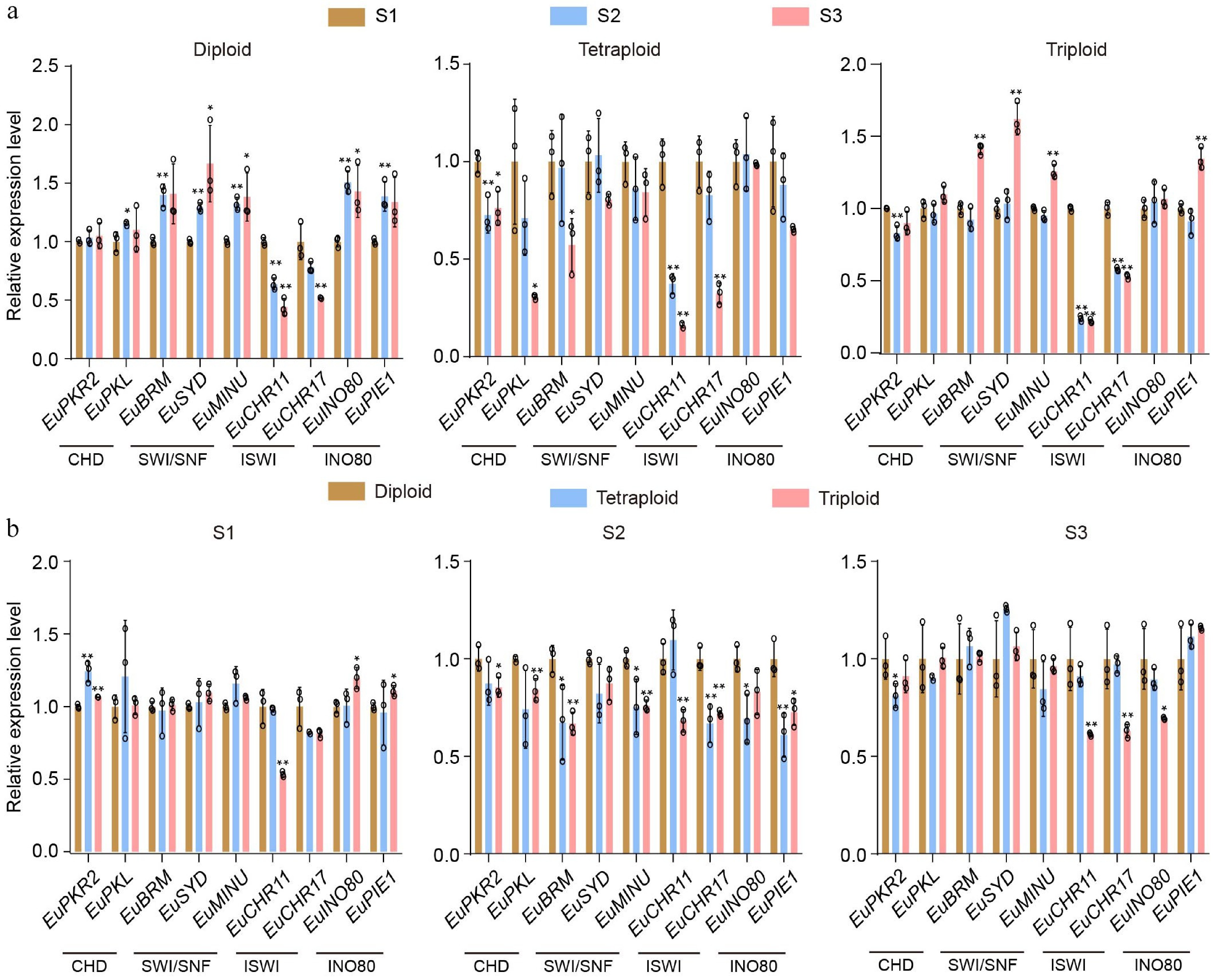
Figure 2.
Transcript profiles of chromatin-remodeling enzymes in E. ulmoides. (a) The relative expression dynamics of chromatin-remodeling enzymes across three leaf developmental stages (S1: mid-to-late April; S2: mid-to-late May; S3: mid-to-late June) in diploid, tetraploid, and triploid plants. (b) The relative expression levels of chromatin-remodeling enzymes among diploid, tetraploid, and triploid at each individual developmental stage (S1 to S3). Values are means ± SD of three biological replicates. All p-values were obtained by two-tailed t-tests. * p < 0.05, and ** p < 0.01 represented the highly significant and significant difference, respectively.
In addition, considering that the triploid variety contain higher rubber content than diploid and tetraploid, the expression levels of EuCHR11 and EuCHR17 in diploid, triploid, and tetraploid plants at stages S1, S2, and S3 were compared. The transcript level of EuCHR11 was found to be significantly lower in triploid than in either diploid or tetraploid at all stages (Fig. 2b). Similarly, EuCHR17 expression was significantly reduced in triploid compared to diploid throughout all stages and was also significantly lower in triploid than in tetraploid plants specifically at stage S3 (Fig. 2b). These findings further confirm that EuCHR11 and EuCHR17 are negative regulators of TPI biosynthesis in E. ulmoides. Their pronounced downregulation contributes to the elevated rubber content observed in the triploid variety.
EuSRPP8 positively regulates TPI biosynthesis in E. ulmoides
-
In rubber-producing plants, the SRPP and REF family genes are involved in rubber biosynthesis[6−9]. The expansion of these gene family is often associated with enhanced rubber yield[5,8]. To explore whether TPI biosynthesis in E. ulmoides is regulated by SRPP and REF genes, BLAST and phylogenetic analyses were performed using SRPP and REF protein sequences from rubber-producing plants, including Hevea brasiliensis, Lactuca serriola, and Taraxacum kok-saghyz, as well as the model plant A. thaliana, to identify orthologs in E. ulmoides. Eight EuSRPP and five EuREF genes were identified, demonstrating a notable expansion of these gene families in E. ulmoides compared to Arabidopsis (Fig. 3a).
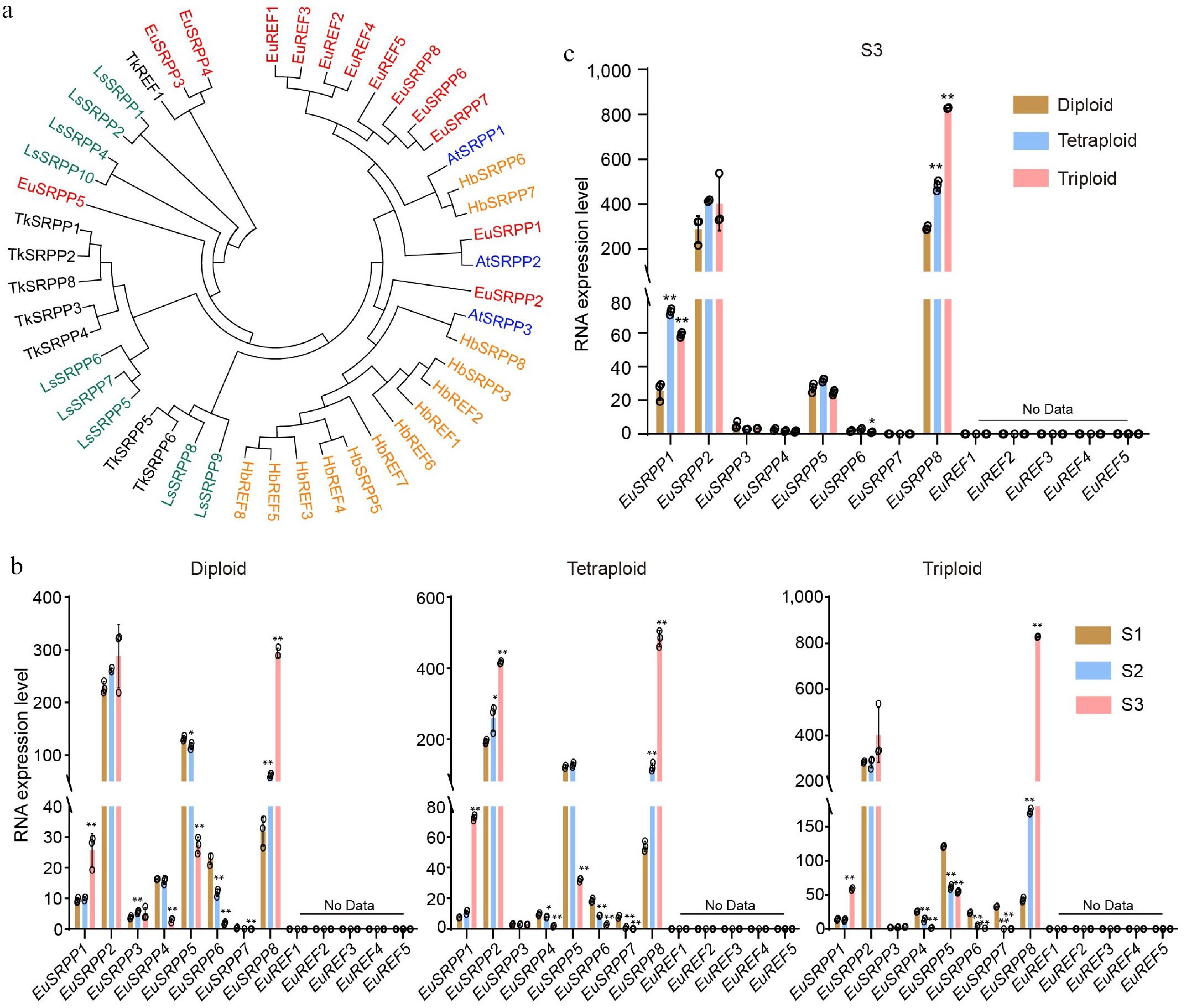
Figure 3.
Transcript profiles of the EuSRPP/REF family genes in E. ulmoides. (a) Phylogenetic relationship of SRPP/REF genes in rubber synthesis plants (Hb: Hevea brasiliensis; Ls: Lactuca serriola; Tk: Taraxacum kok-saghyz; At: Arabidopsis thaliana). (b) The relative expression dynamics of EuSRPP/REF genes across leaf developmental stages (S1: mid-to-late April; S2: mid-to-late May; S3: mid-to-late June) in diploid, tetraploid, and triploid plants. (c) The relative expression levels of EuSRPP/REF genes among diploid, tetraploid, and triploid at stage S3. Values are means ± SD of three biological replicates. All p-values were obtained by two-tailed t-tests. * p < 0.05 and ** p < 0.01 represented the highly significant and significant difference, respectively.
The expression of EuSRPP and EuREF genes were analyzed at stages S1, S2, and S3 in diploid, triploid, and tetraploid by real-time PCR. Among EuSRPP genes, EuSRPP3 showed negligible expression across all ploidies (Fig. 3b). EuSRPP2 was highly expressed and showed an increasing trend from S1 to S3, although not significant (Fig. 3b). EuSRPP1 expression was low at S1 and S2 but increased sharply at S3 (Fig. 3b). Notably, EuSRPP8 exhibited a progressively increasing expression from S1 to S3 (Fig. 3b), a pattern that was positively correlated with TPI accumulation, suggesting its role as a positive regulator of rubber biosynthesis. In contrast, EuSRPP4, EuSRPP5, EuSRPP6, and EuSRPP7 exhibited declining expression trends during the same period (Fig. 3b), inconsistent with the rising trend in TPI content. Given the different levels of TPI content in diploid, tetraploid, and triploid E. ulmoides (Fig. 1c), the expression levels of SRPP genes across these ploidies were compared. Among these genes, only EuSRPP8 exhibited a continuously increased expression that paralleled the elevated TPI content (Fig. 3c), demonstrating a positive correlation between its expression level and TPI accumulation. Furthermore, all EuREF genes, including EuREF1, EuREF2, EuREF3, EuREF4, and EuREF5, exhibited undetectable expression levels (Fig. 3b, c), suggesting that they are not critical regulators of TPI biosynthesis in E. ulmoides, in contrast to their roles in other rubber-producing plants. Together, these results demonstrate that EuSRPP8 may be involved in TPI biosynthesis in E. ulmoides.
CHR11 and CHR17 negatively regulate TPI biosynthesis by repressing SRPP expression
-
Using BLAST and phylogenetic analysis, the EuCHR11 and EuCHR17 are identified in E. ulmoides as orthologs of Arabidopsis AtCHR11 and AtCHR17, respectively. Multiple sequence alignment of the amino acid sequences of the proteins was then performed and revealed that the proteins share high sequence similarity, as well as conserved functional domains, including an ATPase domain and a DNA-binding domain with a 89.49% degree of similarity (Supplementary Fig. S2). These suggest that EuCHR11/17 and AtCHR11/17 likely originated from ancient duplication events and may retain similar molecular functions. In Arabidopsis, the Atchr11/17 double mutant exhibits severe developmental defects[25−27], highlighting the essential roles of AtCHR11 and AtCHR17 in maintaining normal plant growth, and implicating important functions of EuCHR11 and EuCHR17 in E. ulmoides.
To investigate whether the negative regulatory effects of CHR11 and CHR17 on TPI accumulation depend on the expression level of the rubber biosynthesis-related SRPP genes, the changes in expression of SRPP homologs in the Atchr11/17 Arabidopsis mutant were then analyzed using published RNA sequencing (RNA-seq, GSE139465) data[25]. The transcript levels of AtSRPP1, AtSRPP2, and AtSRPP3 were found to be significantly upregulated in the Atchr11/17 mutant, showing increases from 3.4- to 8.7-fold relative to the WT (Fig. 4a). These results suggest that AtCHR11 and AtCHR17 repress the expression of these SRPP genes. Moreover, based on published chromatin immunoprecipitation followed by sequencing (ChIP-seq, GSE139465) data[25], AtCHR11 was found to be enriched at the genomic loci of AtSRPP1, AtSRPP2, and AtSRPP3 (Fig. 4b), indicating that AtCHR11 directly regulates the expression of these genes in Arabidopsis.
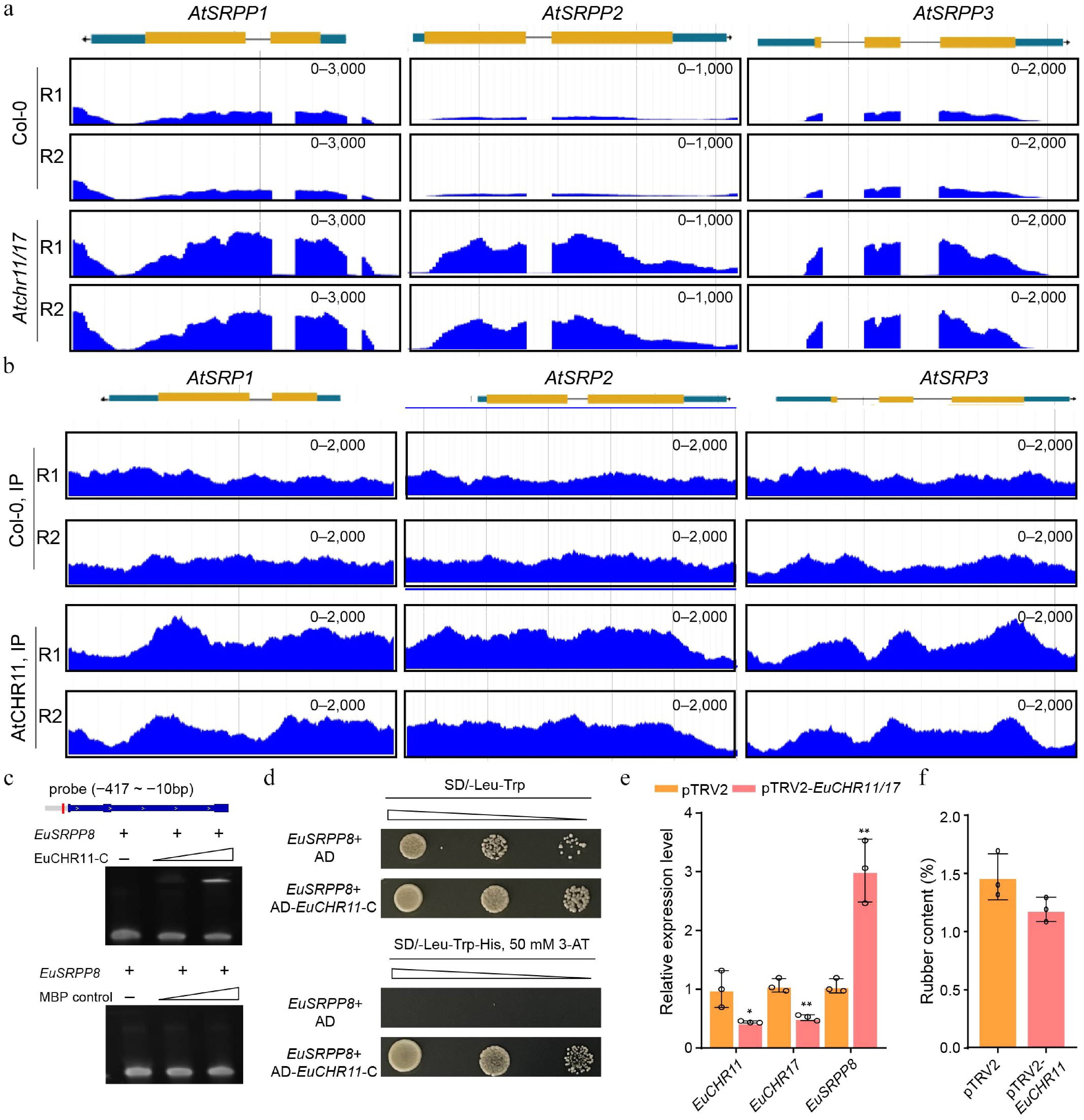
Figure 4.
CHR11 negatively regulates the expression of the SRPP family genes in Arabidopsis and E. ulmoides. (a) Genome browser view of RNA-seq signals of AtSRPP1, AtSRPP2, and AtSRPP3 in the WT and Atchr11/17 double mutant. RPKM scale is shown for each panel. Data from two biological replicates are shown. (b) Genome browser view of AtCHR11 ChIP-seq peaks at the genomic loci of AtSRPP1, AtSRPP2, and AtSRPP3. RPKM scale is shown for each panel. Two replicates are shown. (c) EMSA showing the binding of EuCHR11-C to the EuSRPP8 promoter. The empty MBP protein was used as a negative control. (d) Y1H showing the binding of EuCHR11-C to the EuSRPP8 promoter. The empty AD vector was used as a negative control. (e) The relative expression levels of EuCHR11, EuCHR17 and EuSRPP8 genes infiltrated with TRV2 or TRV2-EuCHR11/17 vector. (f) Statistical analysis of TPI concentration in leaves infiltrated with TRV2 or TRV2-EuCHR11/17 vector. Values are means ± SD of three biological replicates. All p-values were obtained by two-tailed t-tests. * p < 0.05 and ** p < 0.01 represented the highly significant and significant difference, respectively.
Given the sequence and structural conservation of CHR11 and CHR17 proteins between A. thaliana and E. ulmoides, it is speculated that EuCHR11 and EuCHR17 may also directly suppress the expression of SRPP genes, thereby negatively regulating TPI biosynthesis. To verify this hypothesis, the recombinant C-terminal region of the EuCHR11 protein (EuCHR11-C, 750-950 aa) is purified, which contains the DNA-binding domain (Supplementary Fig. S3), and an electrophoretic mobility shift assay (EMSA). The result showed that EuCHR11-C directly binds to the promoter of EuSRPP8, whereas the empty MBP control shows no interaction (Fig. 4c). This result was further corroborated by a Y1H assay, which confirmed the binding capability of the EuCHR11 DNA-binding domain (Fig. 4d).
To further elucidate the function of EuCHR11 and EuCHR17, a TRV2-based vector targeting EuCHR11 is constructed, which shares higher sequence similarity with EuCHR17 (EuCHR11/17-TRV2, Supplementary Fig. S4). Real-time PCR analysis confirmed that the expression of both EuCHR11 and EuCHR17 was successfully knocked down in plants infected with the EuCHR11/17-TRV2 vector compared to those with the empty vector (Fig. 4e, f). Concomitantly, the expression level of EuSRPP8 was upregulated in the knockdown leaves. Furthermore, the suppression of EuCHR11/EuCHR17 led to a decrease in TPI content. These findings support the notion that CHR11 and CHR17 proteins negatively regulate TPI biosynthesis by directly repressing the expression of critical SRPP genes across plant species.
-
Rubber plays an essential role in promoting national industrialization, sustaining social stability, and supporting the progress of human society. The biosynthetic pathway of rubber is regulated by a complex network involving enzymes, scaffold proteins, and epigenetic factors[1,5−7]. The divergence among rubber-producing species in polyisoprene stereochemistry (cis- vs trans-configuration) and associated cellular mechanisms represents a major challenge in elucidating the rubber biosynthetic pathway in plant biology and metabolic engineering. With the rapid development of biotechnology, key regulatory components and pathways of rubber production have been uncovered, contributing to a deeper understanding of the molecular mechanisms of rubber biosynthesis[10,14].
Polyploidy and its role in enhancing rubber yield
-
Recent progress in long-read sequencing, chromatin architecture analysis, and CRISPR/Cas9-mediated genome editing has facilitated the convergence of conventional breeding and molecular design[28]. In E. ulmoides, traditional improvement approaches are largely dependent on cross-breeding and phenotypic screening and are constrained by extended breeding timelines. Although decades of selective breeding have produced elite diploids, induced tetraploids, and innovative triploid hybrids, rubber content in E. ulmoides has stagnated under traditional breeding frameworks. The triploid hybrid obtained in this study exhibited not only characteristic gigas features, including thickened stems, but also a 1.7-fold increase in TPI accumulation relative to diploid controls (Fig. 1c). Flow cytometric analysis confirmed a stable triploid genome (3C DNA content). The enhanced rubber yield aligns with earlier reports that triploidy can decouple cell division from expansion, resulting in increased cytoplasmic volume that contributes to rubber particle accumulation[23,24]. Significantly, the findings in this study imply that this phenotypic gain is modulated by epigenetics rather than solely by gene dosage, as triploid plants exhibited the lowest transcript levels of EuCHR11 and EuCHR17 among all polyploids across all developmental stages (Fig. 2b).
The roles of EuCHR11 and EuCHR17 in regulating rubber biosynthesis
-
Chromatin remodeling plays a pivotal role in regulating chromatin structure, gene expression, development, and metabolic processes[29−32]. In this study, EuCHR11 and EuCHR17 were identified as negative regulators of TPI biosynthesis in E. ulmoides. These genes belong to the ISWI subfamily of ATP-dependent chromatin remodelers, which are conserved across plant species. Findings reveal that EuCHR11 directly targets the promoter of EuSRPP8, a key rubber biosynthesis gene, through its C-terminal DNA-binding domain. This interaction is associated with reduced transcriptional silencing of EuSRPP8. This mechanism is consistent with observations in Arabidopsis, where loss-of-function mutations in AtCHR11 and AtCHR17 derepress downstream target genes AtSRPP1/2/3[25]. Findings also extend this paradigm to woody plants, providing new insights into the conserved role of chromatin remodeling in regulating rubber biosynthesis.
EuSRPP8 as the functional hub of rubber biosynthesis
-
Rubber particles (RPs) are unique organelles that serve as the sites for both the elongation and storage of rubber molecules in rubber-producing plants[1,6,33]. Proteomic analyses have identified SRPPs and REFs as critical regulators of rubber biosynthesis[34]. However, the functions of these proteins remain poorly understood in E. ulmoides. In this study, EuSRPP8 is identified as the dominant isoform driving TPI biosynthesis in E. ulmoides. Phylogenetic analysis revealed lineage-specific expansion of the SRPP/REF family in E. ulmoides, with eight EuSRPP and five EuREF genes. During the rapid rubber-synthesis stage of leaves (S1–S3), EuSRPP8 expression showed the strongest correlation with TPI content. These findings mirror the hierarchy reported for H. brasiliensis, where HbSRPP3 is the dominant isoform driving CPI elongation[33]. It is proposed that EuSRPP8 nucleates the rubber-transferase complex in E. ulmoides, while other EuSRPP isoforms play accessory or compensatory roles. This discovery provides a critical foundation for understanding the regulatory network of rubber biosynthesis in E. ulmoides and opens new avenues for genetic improvement to enhance rubber yield.
Implications and future directions
-
This study uncovers the molecular mechanisms underlying TPI biosynthesis in E. ulmoides, with particular emphasis on the roles of polyploidy, chromatin remodeling, and rubber particle-associated proteins. The findings have important implications for the genetic improvement of rubber-producing plants and the sustainable production of natural rubber. The identification of EuSRPP8 as a functional hub of rubber biosynthesis and the elucidation of its regulatory network provide new targets for metabolic engineering. Furthermore, the discovery of epigenetic regulators for TPI accumulation opens new avenues for epigenome-editing strategies to enhance rubber yield. Future research should focus on validating these findings under field conditions and exploring the potential of E. ulmoides as a rubber-producing crop. By integrating classical breeding with molecular design, the development of high-yielding, sustainable rubber varieties for the benefit of agriculture and industry can be accelerated.
-
In summary, this work elucidates a central epigenetic regulatory module in which EuCHR11 and EuCHR17 constrain TPI biosynthesis by suppressing the key catalyst EuSRPP8. It is demonstrated that the yield superiority of triploid E. ulmoides plants is not from ploidy alone but from epigenetic reprogramming mediated by reduced EuCHR11 and EuCHR17, rather than from a simple gene dosage effect. This discovery establishes EuCHR11 and EuCHR17 as promising targets for epigenome editing, offering a strategic foundation for the precision breeding of high-yielding E. ulmoides varieties.
-
The authors confirm their contributions to the paper as follows: study conception and design: Du P, Wang L, Wu CJ; experimental work: Du P, Zhang J, Li ZG, Qiu TY; bioinformatics analysis: Li ZG, Wang Y; data analysis: Du P, Wang L, Wu CJ; draft manuscript preparation: Du P, Wang L, Wu CJ. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. The raw RNA-seq, and ChIP-seq data from that work have been deposited in the Gene Expression Omnibus (GEO) database under the accession number GSE139465[25] (www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE139465).
-
This work was supported by the Natural Science Foundation of Southwest University of Science and Technology (Grant No. 23zx7138), and the Sichuan Innovation Team of National Modern Agricultural Industry Technology System (Grant No. SCCXTD-2024-1).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Ping Du, Jie Zhang
- Supplementary Table S1 All primer sequences used in this study.
- Supplementary Fig. S1 Analysis of the chromosomal localization (a) and physico-chemical characteristics (b) of the chromatin remodeling enzymes in E. ulmoides.
- Supplementary Fig. S2 Amino acid sequences alignment of the CHR11 and CHR17 proteins in Arabidopsis and E. ulmoides. The conserved ATPase domain and DNA binding domain are labeled. Alignment was performed using DNAMAN software.
- Supplementary Fig. S3 Purification of the recombinant EuCHR11-C protein expressed in E. coli. The red arrow indicates the EuCHR11-C protein.
- Supplementary Fig. S4 Sequences alignment of the EuCHR11 and EuCHR17 CDS in E. ulmoides. The conserved CDS sequence for VIGS assay is labeled. Alignment was performed using DNAMAN software.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Du P, Zhang J, Li ZG, Qiu TY, Wang Y, et al. 2026. EuCHR11 and EuCHR17 negatively regulate trans-polyisoprene biosynthesis by repressing EuSRPP8 expression. Epigenetics Insights 19: e001 doi: 10.48130/epi-0025-0018 

EuCHR11 and EuCHR17 negatively regulate trans-polyisoprene biosynthesis by repressing EuSRPP8 expression
- Received: 04 September 2025
- Revised: 28 November 2025
- Accepted: 19 December 2025
- Published online: 22 January 2026
Abstract: Eucommia ulmoides could produce high-quality trans-polyisoprene (TPI), providing a new and alternative industrial material for natural rubber. However, the molecular mechanisms underlying TPI biosynthesis remain poorly understood. In this study, triploid E. ulmoides plants were generated through polyploid breeding and shower higher TPI content compared to diploid and tetraploid plants. Using real-time PCR analysis, it was found that the expression of two imitation switch (ISWI)-family chromatin remodelers, EuCHR11 and EuCHR17, were downregulated in high-TPI-yielding triploid plants during the period of TPI biosynthesis, whereas the expression of the key rubber biosynthesis gene EuSRPP8 was significantly upregulated under the same conditions. In Arabidopsis thaliana, it was demonstrated that the orthologs AtCHR11 and AtCHR17 directly bind to the genomic loci of AtSRPP1, AtSRPP2, and AtSRPP3 and repress their expression. Similarly, EuCHR11 also directly binds to the promoter of EuSRPP8 in E. ulmoides. These results together indicate that EuCHR11 and EuCHR17 negatively regulate TPI biosynthesis by repressing EuSRPP8 expression. The findings in this study uncover a previously uncharacterized epigenetic regulatory pathway mediated by chromatin remodeling factors that modulates rubber biosynthesis and enhances the understanding of the molecular mechanisms behind rubber production.
-
Key words:
- Eucommia rubber /
- Trans-polyisoprene /
- EuCHR11 /
- EuCHR17 /
- EuSRPP8